Introduction
Cancer/testis (CT) antigens are a category of
tumor-associated antigens expressed in a proportion of many
different types of cancers, but not in normal tissues, except for
testis and placenta (1–4). Also, CT antigens are frequently
mutated in cancers, and many CT antigens were identified by using
sera of cancer patients (3,5–9),
implying they are highly antigenic. CT antigens are classified into
two categories based on the chromosomal location (3). CT-X antigens are located on the X
chromosome, are often members of gene families, and represent more
than half of all CT antigens identified. In contrast, non-X CT
antigens are distributed throughout the genome and mostly
single-copy genes. To date, >200 CT antigens, including MAGE,
NY-ESO-1, GAGE, AKAP3, SSX, and LAGE, have been identified, but the
function of most is largely unknown (3,10–12).
Recent studies indicated that members of CT antigens
are likely involved in cell cycle regulation, transcriptional
control, cell survival, and apoptosis (5,13–21).
CT45A1, a member of cancer/testis antigen-45 family (CT45),
functions as a novel proto-oncogene by upregulating various
oncogenic and metastatic genes and activating ERK and CREB
signaling pathways to trigger oncogenesis and metastasis (17). Another study on functions of a CT
antigen reported that the growth promoting and survival properties
of SSX are mediated in part through modulation of MAPK/Erk and Wnt
signaling pathways (22).
Among CT antigens, NY-SAR-35 was previously
identified by SEREX analysis from human sarcoma patients. The
NY-SAR-35 gene is located on chromosome Xq28 like several other CT
antigens and not a member of a multigene family. In addition, the
putative NY-SAR-35 protein is expected to have a signal peptide
domain, a transmembrane domain, and a cystein-rich trefoil/P-domain
(23). Despite the expression of
NY-SAR-35 in various malignancies, it is expressed with low
frequency or not expressed in some cancer cells (3,24).
In our previous study, treatment with the DNA demethylating agent
can also lead to an increase in the absent or downregulated
expression of NY-SAR-35, and transcriptional silencing of NY-SAR-35
is caused by hypermethylation of its promoter (25). This finding proposed
hypomethylation of CpG islands within the promoter region be a
primary mechanism for transcriptional activation of NY-SAR-35 gene
in cancer. However, the functions of this antigen remain
unexplored.
In the present study, the functional effect of
NY-SAR-35 on cell growth, proliferation, and invasion was assessed
by an in vitro cell culture system in HEK293 cell line.
Materials and methods
Cell culture
The human embryonic kidney (HEK) 293 and the murine
fibroblast cell line NIH 3T3 cells were cultured in DMEM
(Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% (v/v)
FBS, 2 mM L-glutamine, 100 U/ml penicillin, and 100 μg/ml
streptomycin. The cells were obtained from American Type Culture
Collection (ATCC) and cultured at 37°C in a humidified atmosphere
containing 5% CO2. The study was conducted under an
approved protocol from the Ethics Committee of Pusan National
University.
RNA isolation and RT-PCR
Total RNA was extracted from cells by using the
RNeasy mini kit (Qiagen, Hilden, Germany). The total RNA of cDNA
was synthesized from 1 μg total RNA by using M-MLV reverse
transcriptase (Promega, Fitchburg, WI, USA). Primers used were
NY-SAR-35-forward (5′-CTTGGTGCGA TCAGCCTTAT-3′) and
NY-SAR-35-reverse (5′-TTGATGCA TGAAAACAGAAC-3′). PCR amplification
was carried out as follows: denaturation for 5 min at 94°C, then 35
cycles consisting of 30 sec at 94°C, 30 sec at 55°C, and 1 min at
72°C and then 10 min at 72°C for final extension. PCR products were
analyzed by agarose gel electrophoresis and visualized by ethidium
bromide.
Western blot analysis
Cells were lysed in lysis buffer and cellular debris
was removed by centrifugation at 15,000 rpm for 10 min. Proteins
were separated by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis, transferred onto nitrocellulose membranes
(Hybond-ECL, GE Healthcare, UK), blocked with 5% skim milk and
incubated with anti-V5 or anti-His (Life Technologies, Carlsbad,
CA, USA) and anti-β actin antibodies (Sigma-Aldrich). Membranes
were then incubated with horseradish peroxidase-conjugated goat
anti-mouse-IgG secondary antibodies for 1 h at room temperature and
detected by enhanced chemiluminescence (Perkin-Elmer Life Science,
MD, USA).
Generation of recombinant NY-SAR-35
protein
The open reading frame (ORF) cDNA inserts of
NY-SAR-35 (AY211917) were selected to generate His-tagged NY-SAR-35
protein. The primers for the partial protein were GGAATTCCATATGCT
GCCCAACAGT (forward) and CCGCTCGAGCATTTGCA TCATCT (reverse)
containing NdeI and XhoI restriction enzyme sites.
PCR products were subcloned into the pET21a expression plasmid
containing a polyhistidine-tag. The recombinant NY-SAR-35 fusion
protein was induced by isopropyl β-D-thiogalactoside (IPTG) with a
final concentration of 0.4 mM at 37°C. Affinity chromatography
using Ni-NTA agarose (Qiagen) was performed to purify NY-SAR-35
recombinant protein. The purified recombinant protein was
determined by SDS-PAGE and Coomassie Blue staining.
Production and purification of polyclonal
NY-SAR-35 antibodies
Antibodies against NY-SAR-35 were raised in a naïve
New Zealand white rabbit immunized subcutaneously with 200 μg of
recombinant NY-SAR-35 protein. Three booster injections were given
using the same antigen, the first booster three weeks after the
first immunization and then each week. The rabbit was then
sacrificed and the crude antiserum was collected. Especially for
the isolation of NY-SAR-35 purified antibodies, affinity
purification was performed using cyanogen bromide (CNBr)-activated
sepharose beads (GE Healthcare). Normally, coupling to beads was
achieved at pH 8.3, while elution occurred at pH 2.8.
Neutralization by adding Tris buffer with pH 8.8 was performed as
soon as possible after elution to prevent damage of the
antibody.
Construction of stable and transient cell
lines
To generate stably transfected cell lines, ORF of
NY-SAR-35 including 5′-untranslated region was cloned into
pcDNA3.1/V5-HisA vector which has C-terminal fusion tag (V5 and 6X
His epitopes) containing EcoRI and XhoI sites. Then,
HEK293 cells (1×106 cells in 60-mm dishes) were
transfected with 4 μg of cloned pcDNA3.1/V5-HisA-NY-SAR-35 using
Lipofectamine LTX (Life Technologies). Transfected cells were
selected in medium containing G418 (1 mg/ml). The selected clones
were maintained in medium containing G418 (0.3 mg/ml).
For transient transfection, the cDNA of complete
coding region of NY-SAR-35 was generated by PCR amplification using
primers containing XhoI and PstI restriction sites.
The PCR products were then inserted in-frame within the pEGFP-N1
vector. The NIH 3T3 cell was transfected with cloned
pEGFP-N1-NY-SAR-35.
siRNA transfection
The small interefing RNA (siRNA) used for the
targeted silencing of NY-SAR-35 (5′-GCCUUAUCCUGGU AUGUCUTT-3′) was
purchased from Qiagen. In brief, cells (2×105 cells/ml)
were seeded on 6-well plates and transfected with 200 nM of siRNA
using oligofectamine reagent, according to the manufacturer's
instructions (Invitrogen, Carlsbad, CA, USA). Cells were then
incubated at 37°C for 4 h in serum-free media. The transfected
cells were collected after 48 h and NY-SAR-35 expression levels in
the cells were determined using RT-PCR.
Immunohistochemistry
Staining was conducted with the peroxidase-based
EnVision Detection kit (DakoCytomation, USA) by following the user
manual. Briefly, tissue sections were deparaffinized and hydrated
in xylene and graded alcohol series. Then, the sections were
incubated with the purified NY-SAR-35 antibody for 1 h at room
temperature. After incubation with EnVision solution, the 3,
3′-diaminobenzidine (DAB) substrate was added followed by
hematoxylin counterstaining and mounting.
Cell counting
Cell numbers were determined as follows: cells
(5×105/90-mm culture plate) were cultured for 4 days in
standard culture medium supplemented with 1 or 10% FBS. After
incubation for 4 days, adherent and vital cells were counted in a
trypan blue exclusion assay.
Bromodeoxyuridine (BrdU) incorporation
assay
Cell proliferation was measured by
5-bromo-2′-deoxyuridine (BrdU) incorporation using the Cell
Proliferation ELISA, BrdU kit (Roche Diagnostics, Germany)
according to the manufacturer's instructions. In brief, cells were
grown in 96-well plates and labeled with BrdU for 2 h.
Subsequently, anti-BrdU-POD Fab fragments and substrates were added
to the medium. The optical density (OD) was determined at 450 nm
using an ELISA reader (Bio-Tek Instruments, VT, USA).
Immunofluorescent staining
Cells grown on coverglass-bottom confocal dish were
fixed with 4% paraformaldehyde. Then, cells were blocked in 3%
FBS/PBS and incubated with anti-PCNA (proliferating cell nuclear
antigen) or Ki-67 antibody. The cells were washed three times with
PBS, incubated with FITC-coupled secondary antibodies. Images were
captured using confocal microscopy (Olympus).
Cell migration and invasion assay
To measure the cell migration activity, transwell
chamber assays were performed using a transwell plate
(Becton-Dickinson, MA, USA). The lower surface of the filters was
coated with 1% gelatin. Cells were resuspended in serum-free medium
and then added to the upper chamber at a density of
5×104 cells/insert. The medium containing 10% FBS was
added to the lower chamber. After the cells were incubated at 37°C
for 18 h, the number of cells that migrated to the lower side of
the upper chamber was counted by staining with crystal violet. The
invasive potentials of the cells were analyzed using a
Matrigel-coated modified Boyden chamber (Becton-Dickinson). The
medium containing 10% FBS was added to the lower chamber as an
attractant. The incubations of the cells were continued for 24 h.
Inserts were then stained with crystal violet and the number of
invaded cells was counted.
Results
NY-SAR-35 protein is located in the
cytoplasm
To investigate the role of NY-SAR-35 gene,
NY-SAR-35-transfected HEK293 cells were established with
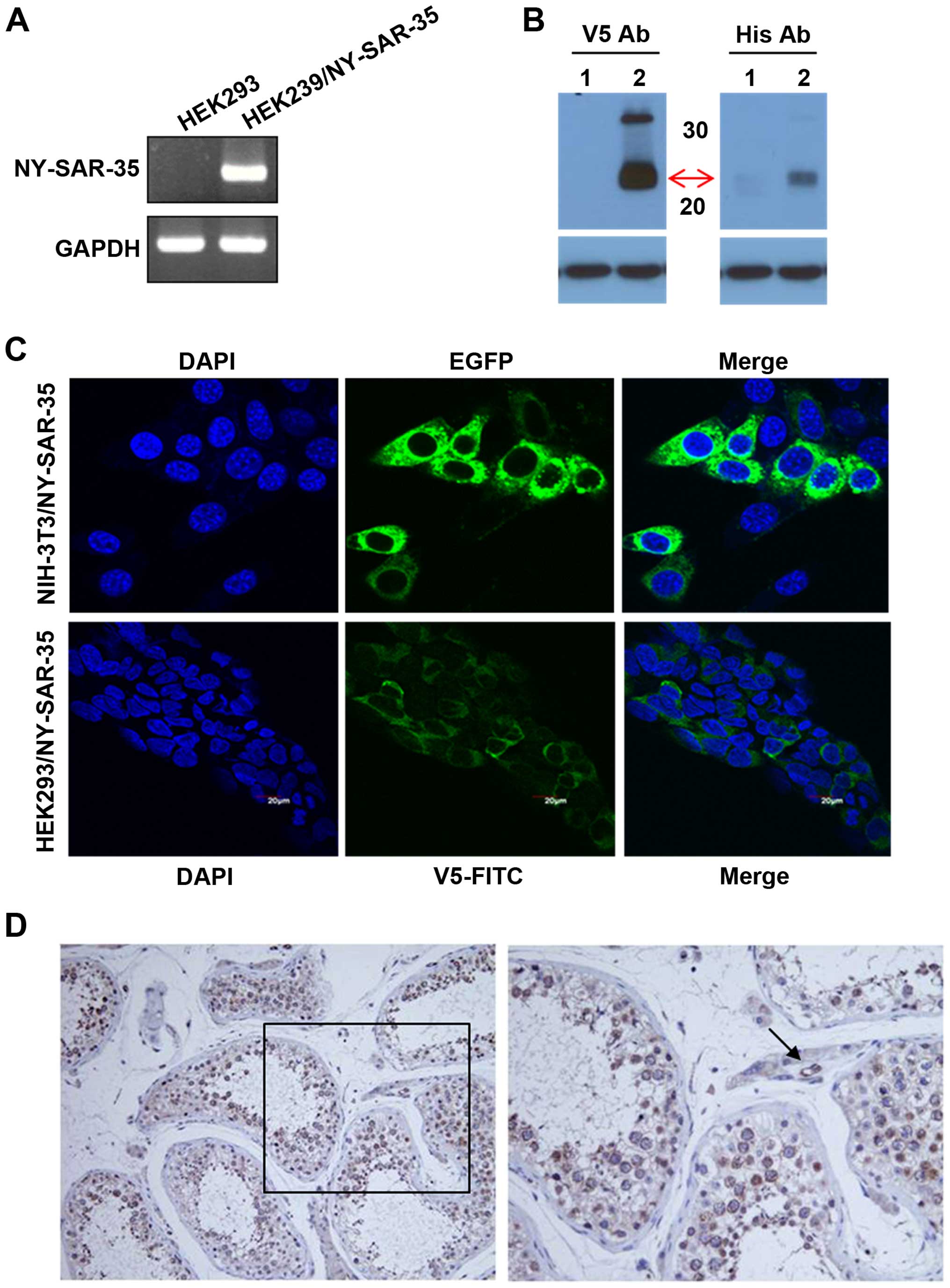
pcDNA3.1/V5-HisA-NY-SAR-35 vector and named HEK293/NY-SAR-35.
Expression of NY-SAR-35 in HEK293/NY-SAR-35 cells was confirmed
with RT-PCR (Fig. 1A) and western
blot analysis (Fig. 1B). In
addition, immunofluorescent staining with EGFP or V5-FITC showed
that NY-SAR-35 was predominantly observed at the cytoplasm in
NIH3T3/NY-SAR-35 and HEK293/NY-SAR-35 cells, respectively (Fig. 1C).
Using purified antibody, the expression of NY-SAR-35
protein in normal testis tissue was analyzed using
immunohistochemical techniques. Expression of NY-SAR-35 was mainly
observed in spermatogonia and spermatocytes, as cytoplasmic
proteins. In addition, a positive reaction was seen in vascular
endothelial cells, but not Leydig cells in interstitium (Fig. 1D).
NY-SAR-35 enhances cell growth and
proliferation in HEK293 cells
Because NY-SAR-35 protein seemed to be involved in
mitosis during spermatogenesis (Fig.
1D), we tested whether the NY-SAR-35 could regulate cell growth
or proliferation. The increase in cell number of HEK293/NY-SAR-35
cells was determined by trypan blue exclusion assay.
HEK293/NY-SAR-35 cells grew on average 2.2 times more rapidly than
HEK293 cells within the time period of 4 days (Fig. 2A).
Further examination of cell proliferation by BrdU
assay showed an increase in DNA synthesis within three days of
in vitro growth in HEK293/NY-SAR-35 cells (Fig. 2B). Also, positive staining for
Ki-67 (green, Fig. 2C) and PCNA
(green, Fig. 2D), markers of cell
proliferation, was found in HEK293/NY-SAR-35 cells. Ki-67 staining
was restricted to the nucleus. In contrast, PCNA positivity showed
staining intensity with both the nuclei and the cytoplasm. It was
noted that NY-SAR-35 enhanced cell proliferation.
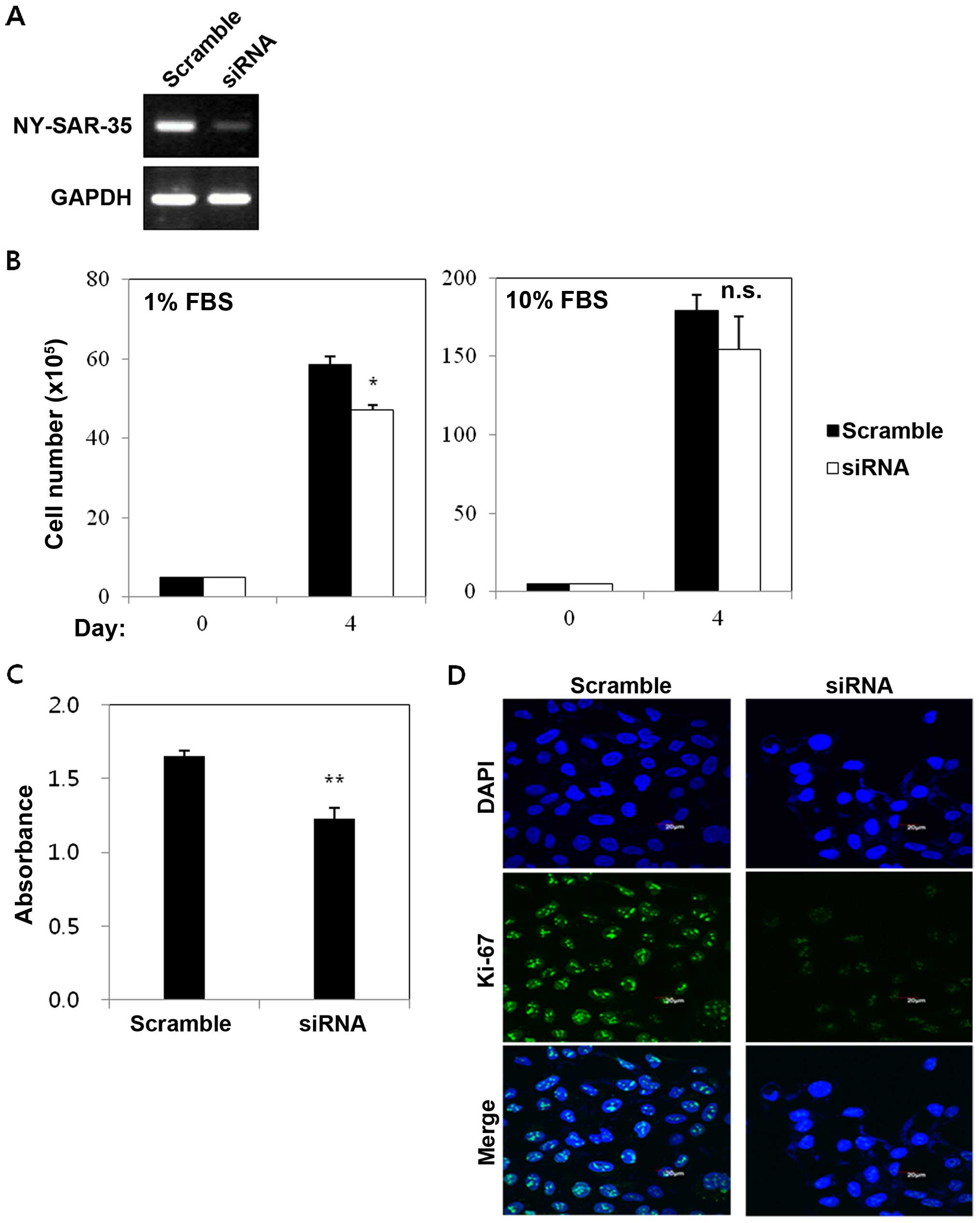
Furthermore, to confirm these effects of NY-SAR-35,
knockdown of NY-SAR-35 was performed using RNAi molecule specific
for NY-SAR-35. RT-PCR showed that NY-SAR-35-siRNA effectively
silenced NY-SAR-35 expression in HEK293/NY-SAR-35 cells (Fig. 3A). For growth assay, the control
scrambled- and NY-SAR-35-siRNA treated cells were incubated for 4
days. The percentage of cell numbers dropped to 20% in NY-SAR-35
knocked-down cells compared to HEK293/NY-SAR-35 cells under media
containing 1% FBS (Fig. 3B). Cell
proliferation assay showed that DNA synthesis decreased in
NY-SAR-35-silenced cells compared with HEK293/NY-SAR-35 cells
(Fig. 3C). Immunofluorescent
staining also showed that knocking down NY-SAR-35 expression
significantly reduced positivity of Ki-67 (Fig. 3D).
NY-SAR-35 promotes cell migration and
invasion in HEK293 cells
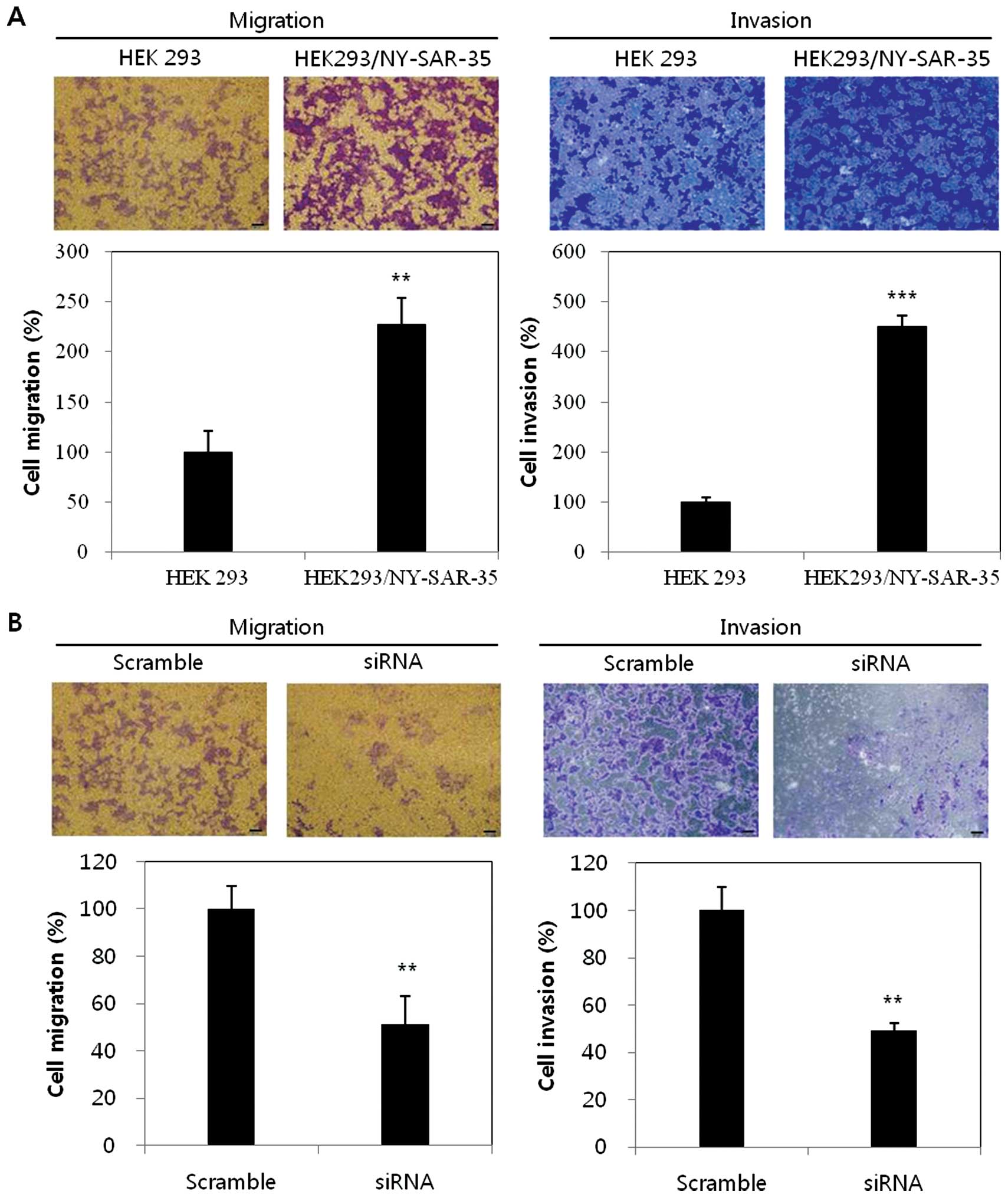
To understand the relationship between the
expression of NY-SAR-35 and cellular motility, migration and
invasion activity, we were examined in transwell assays using a
Boyden chamber. HEK293/NY-SAR-35 cells showed that the migratory
ability was increased to 2.3-fold and invasive ability was also
enhanced to 4.5-fold higher than the control HEK293 cells (Fig. 4A).
Furthermore, to confirm the migratory and invasive
effects of NY-SAR-35 expression, NY-SAR-35 was knocked down in the
HEK293/NY-SAR-35 cells and migration and invasion assays were
performed. NY-SAR-35 silencing reduced migration and invasion of
HEK293/NY-SAR-35 cells by ~50% (Fig.
4B). These findings indicated that NY-SAR-35 expression
promoted cell migration and invasion in HEK293 cells.
Discussion
Many studies have analyzed CT antigens for their
potential use in cancer immunotherapy because of their
immunogenicity and restricted expression patterns (1–4,10).
This study provided evidence that overexpression of
NY-SAR-35 increased cell proliferation, migration, and invasion in
HEK293 cells. We have for the first time demonstrated that
NY-SAR-35 impacts cell proliferation, migration, and invasion.
Although the role of NY-SAR-35 was not directly evaluated, these
experiments illustrated the potential contribution of the CT
antigen NY-SAR-35 in proliferative and migratory function.
During spermatogenesis, different CT antigens
uniquely expressed in different germ cell types, demonstrating
involvement in different phases of spermatogenesis (3,12,26).
Immunohistochemical staining in the testis tissue showed that
NY-SAR-35 was mainly expressed in spermatogonia and primary and/or
secondary spermatocytes, which was observed in most of CT antigens
located on X chromosome (26).
NY-SAR-35 was also present in the cytoplasmic compartment; however,
predominant cytoplasmic expression was rare, with CT47, another CT
antigen, as an example (27). This
finding demonstrated NY-SAR-35 was associated with spermatogonia
and spermatocyte and had possible roles during mitotic division of
spermatogenesis.
We also found that the Ki-67 was localized within
the nucleus of HEK293/NY-SAR-35 cells. The Ki-67 antigen has been
detected in proliferating cells in all phases except G0 phase of
the cell-division cycle of normal and neoplastic cells. Equal to
Ki-67, PCNA detection was increased in HEK293/NY-SAR-35 cells.
Possibly NY-SAR-35 was able to perform a number of functions within
proliferating cells.
In addition, the functional study of NY-SAR-35 in
HEK293 cell lines demonstrated that HEK293/NY-SAR-35 cells have
greater migratory and invasive abilities compared to control cells;
whereas suppression of NY-SAR-35 showed reduction in migration and
invasion capabilities of HEK293/NY-SAR-35 cells.
The regulations of NY-SAR-35-mediated cell
proliferation and migration were determined, however, various
questions need to be addressed and investigated in further studies,
including: what are the potential mechanisms to increase the cell
proliferation and migration of HEK293 cells by expression of
NY-SAR-35? How do the patterns of gene expression change in HEK293
cells by overexpression of NY-SAR-35? Also it is important to
explore whether the phenotypic changes are shown in cancer cell
types by overexpression of NY-SAR-35.
In conclusion, the above data demonstrated that the
expression of NY-SAR-35 stimulated the cell growth and
proliferation in vitro assay. Also, NY-SAR-35 enhanced cell
migration and invasion, suggesting that NY-SAR-35 gene appears to
be able to mediate proliferative and migratory signals. In
addition, NY-SAR-35 may be a useful tool in research on the linkage
between embryology and tumor biology.
Acknowledgements
This study was supported by Basic Science Research
Program through the National Research Foundation of Korea (NRF)
funded by the Ministry of Education (NRF-2012R1A1A2041573).
References
|
1
|
Scanlan MJ, Gure AO, Jungbluth AA, Old LJ
and Chen YT: Cancer/testis antigens: An expanding family of targets
for cancer immunotherapy. Immunol Rev. 188:22–32. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Scanlan MJ, Simpson AJ and Old LJ: The
cancer/testis genes: Review, standardization, and commentary.
Cancer Immun. 4:12004.PubMed/NCBI
|
|
3
|
Simpson AJ, Caballero OL, Jungbluth A,
Chen YT and Old LJ: Cancer/testis antigens, gametogenesis and
cancer. Nat Rev Cancer. 5:615–625. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zendman AJ, Ruiter DJ and Van Muijen GN:
Cancer/testis-associated genes: Identification, expression profile,
and putative function. J Cell Physiol. 194:272–288. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen YT, Scanlan MJ, Sahin U, Türeci O,
Gure AO, Tsang S, Williamson B, Stockert E, Pfreundschuh M and Old
LJ: A testicular antigen aberrantly expressed in human cancers
detected by autologous antibody screening. Proc Natl Acad Sci USA.
94:1914–1918. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Song MH, Choi KU, Shin DH, Lee CH and Lee
SY: Identification of the cancer/testis antigens AKAP3 and CTp11 by
SEREX in hepatocellular carcinoma. Oncol Rep. 28:1792–1798.
2012.PubMed/NCBI
|
|
7
|
Chen YT, Chadburn A, Lee P, Hsu M, Ritter
E, Chiu A, Gnjatic S, Pfreundschuh M, Knowles DM and Old LJ:
Expression of cancer testis antigen CT45 in classical Hodgkin
lymphoma and other B-cell lymphomas. Proc Natl Acad Sci USA.
107:3093–3098. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Song MH, Ha JM, Shin DH, Lee CH, Old L and
Lee SY: KP-CoT-23 (CCDC83) is a novel immunogenic cancer/testis
antigen in colon cancer. Int J Oncol. 41:1820–1826. 2012.PubMed/NCBI
|
|
9
|
Song MH, Ha JC, Lee SM, Park YM and Lee
SY: Identification of BCP-20 (FBXO39) as a cancer/testis antigen
from colon cancer patients by SEREX. Biochem Biophys Res Commun.
408:195–201. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fratta E, Coral S, Covre A, Parisi G,
Colizzi F, Danielli R, Nicolay HJ, Sigalotti L and Maio M: The
biology of cancer testis antigens: Putative function, regulation
and therapeutic potential. Mol Oncol. 5:164–182. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ong SE and Mann M: A practical recipe for
stable isotope labeling by amino acids in cell culture (SILAC). Nat
Protoc. 1:2650–2660. 2006. View Article : Google Scholar
|
|
12
|
Dobrynin P, Matyunina E, Malov SV and
Kozlov AP: The novelty of human cancer/testis antigen encoding
genes in evolution. Int J Genomics. 2013:1051082013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Doyle JM, Gao J, Wang J, Yang M and Potts
PR: MAGE-RING protein complexes comprise a family of E3 ubiquitin
ligases. Mol Cell. 39:963–974. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Epping MT, Wang L, Edel MJ, Carlée L,
Hernandez M and Bernards R: The human tumor antigen PRAME is a
dominant repressor of retinoic acid receptor signaling. Cell.
122:835–847. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Whitehurst AW, Xie Y, Purinton SC, Cappell
KM, Swanik JT, Larson B, Girard L, Schorge JO and White MA: Tumor
antigen acrosin binding protein normalizes mitotic spindle function
to promote cancer cell proliferation. Cancer Res. 70:7652–7661.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cappell KM, Sinnott R, Taus P, Maxfield K,
Scarbrough M and Whitehurst AW: Multiple cancer testis antigens
function to support tumor cell mitotic fidelity. Mol Cell Biol.
32:4131–4140. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shang B, Gao A, Pan Y, Zhang G, Tu J, Zhou
Y, Yang P, Cao Z, Wei Q, Ding Y, et al: CT45A1 acts as a new
proto-oncogene to trigger tumorigenesis and cancer metastasis. Cell
Death Dis. 5:e12852014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bai S, He B and Wilson EM: Melanoma
antigen gene protein MAGE-11 regulates androgen receptor function
by modulating the interdomain interaction. Mol Cell Biol.
25:1238–1257. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Monte M, Simonatto M, Peche LY, Bublik DR,
Gobessi S, Pierotti MA, Rodolfo M and Schneider C: MAGE-A tumor
antigens target p53 transactivation function through histone
deacetylase recruitment and confer resistance to chemotherapeutic
agents. Proc Natl Acad Sci USA. 103:11160–11165. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cilensek ZM, Yehiely F, Kular RK and Deiss
LP: A member of the GAGE family of tumor antigens is an
anti-apoptotic gene that confers resistance to Fas/CD95/APO-1,
Interferon-gamma, taxol and gamma-irradiation. Cancer Biol Ther.
1:380–387. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hamer G, Gell K, Kouznetsova A, Novak I,
Benavente R and Höög C: Characterization of a novel
meiosis-specific protein within the central element of the
synaptonemal complex. J Cell Sci. 119:4025–4032. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
D'Arcy P, Maruwge W, Wolahan B, Ma L and
Brodin B: Oncogenic functions of the cancer-testis antigen SSX on
the proliferation, survival, and signaling pathways of cancer
cells. PLoS One. 9:e951362014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee SY, Obata Y, Yoshida M, Stockert E,
Williamson B, Jungbluth AA, Chen YT, Old LJ and Scanlan MJ:
Immunomic analysis of human sarcoma. Proc Natl Acad Sci USA.
100:2651–2656. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim YD, Park HR, Song MH, Shin DH, Lee CH,
Lee MK and Lee SY: Pattern of cancer/testis antigen expression in
lung cancer patients. Int J Mol Med. 29:656–662. 2012.PubMed/NCBI
|
|
25
|
Park JH, Song MH, Lee CH, Lee MK, Park YM,
Old L and Lee SY: Expression of the human cancer/testis antigen
NY-SAR-35 is activated by CpG island hypomethylation. Biotechnol
Lett. 33:1085–1091. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cheng YH, Wong EW and Cheng CY:
Cancer/testis (CT) antigens, carcinogenesis and spermatogenesis.
Spermatogenesis. 1:209–220. 2011. View Article : Google Scholar
|
|
27
|
Caballero OL and Chen YT: Cancer/testis
(CT) antigens: Potential targets for immunotherapy. Cancer Sci.
100:2014–2021. 2009. View Article : Google Scholar : PubMed/NCBI
|