Cancer is the second leading cause of mortality
worldwide after cardiovascular disease (1). Among men, the most common cancers
include lung, liver, prostate, colorectal and gastric cancers
(2). Lung cancer, broadly
classified into small-cell (SC) and non-SC lung cancer (NSCLC),
remains the leading cause of cancer-related mortality in males and
the second highest cause in female patients, following breast
cancer globally (3). NSCLC
represents the major histological subtype of lung cancer (4-7),
accounting for >80% of all cases. It comprises two primary
subtypes: Lung squamous cell carcinoma (LUSC) and lung
adenocarcinoma (LUAD) (8). The
global incidence of lung cancer is projected to increase from 1.7
million new cases in 2018 to 3.8 million/year by 2070 (9,10).
Although multiple approaches have decreased lung
cancer mortality, the 5-year overall survival (OS) rate remains
low, at 4-17%, depending on the disease stage and geographical
factors (10). Surgery, targeted
therapy, immunotherapy and radiotherapy are recommended therapeutic
regimens for patients with NSCLC. For advanced NSCLC and earlier
stages, the standard first-line treatment remains systemic
platinum-based chemotherapy (11). Platinum-based chemotherapy is
often combined with pemetrexed, vinorelbine/taxanes and gemcitabine
as a first-line regimen for NSCLC. However, the effectiveness of
platinum-based chemotherapy is limited by chemoresistance, which
varies between individuals (12,13). Therefore, novel therapeutic
mechanisms are needed (14,15).
Ferroptosis is a recently discovered type of
regulated cell death (RCD) characterized by iron-dependent toxic
accumulation of lipid peroxides on cellular membranes (16-18). Conventional therapies, including
chemotherapy, radiotherapy, immunotherapy, and targeted cancer
therapy, exert anti-tumor effects by inducing ferroptosis (16,19-22). Thus, targeting ferroptosis offers
a promising therapeutic strategy, especially for cancer resistant
to conventional treatments, including NSCLC (13,23). Understanding the molecular
mechanisms that regulate ferroptosis in NSCLC may enable
development of more effective therapeutic strategies for this
disease.
Non-coding RNAs (ncRNAs) are a class of RNA
molecules with no or limited protein-coding potential (24). ncRNAs, which include circular RNAs
(circRNAs), long ncRNAs (lncRNAs) and microRNAs (miRNAs or miRs),
are key epigenetic regulators of ferroptosis (25-31). ncRNAs regulate tumor malignancy by
modulating ferroptosis in various types of cancers, including NSCLC
(32,33). However, the mechanism underlying
ncRNA-mediated epigenetic regulation of ferroptosis in NSCLC
remains unclear. The present review summarizes the core molecular
mechanisms of ferroptosis, how epigenetic mechanisms modify
ferroptosis in NSCLC and research progress on ncRNA-mediated
regulation of ferroptosis in NSCLC to clarify understanding of
ncRNA-mediated epigenetic modulation of ferroptosis in NSCLC,
highlighting the ncRNA-ferroptosis axis as a potential therapeutic
target for NSCLC.
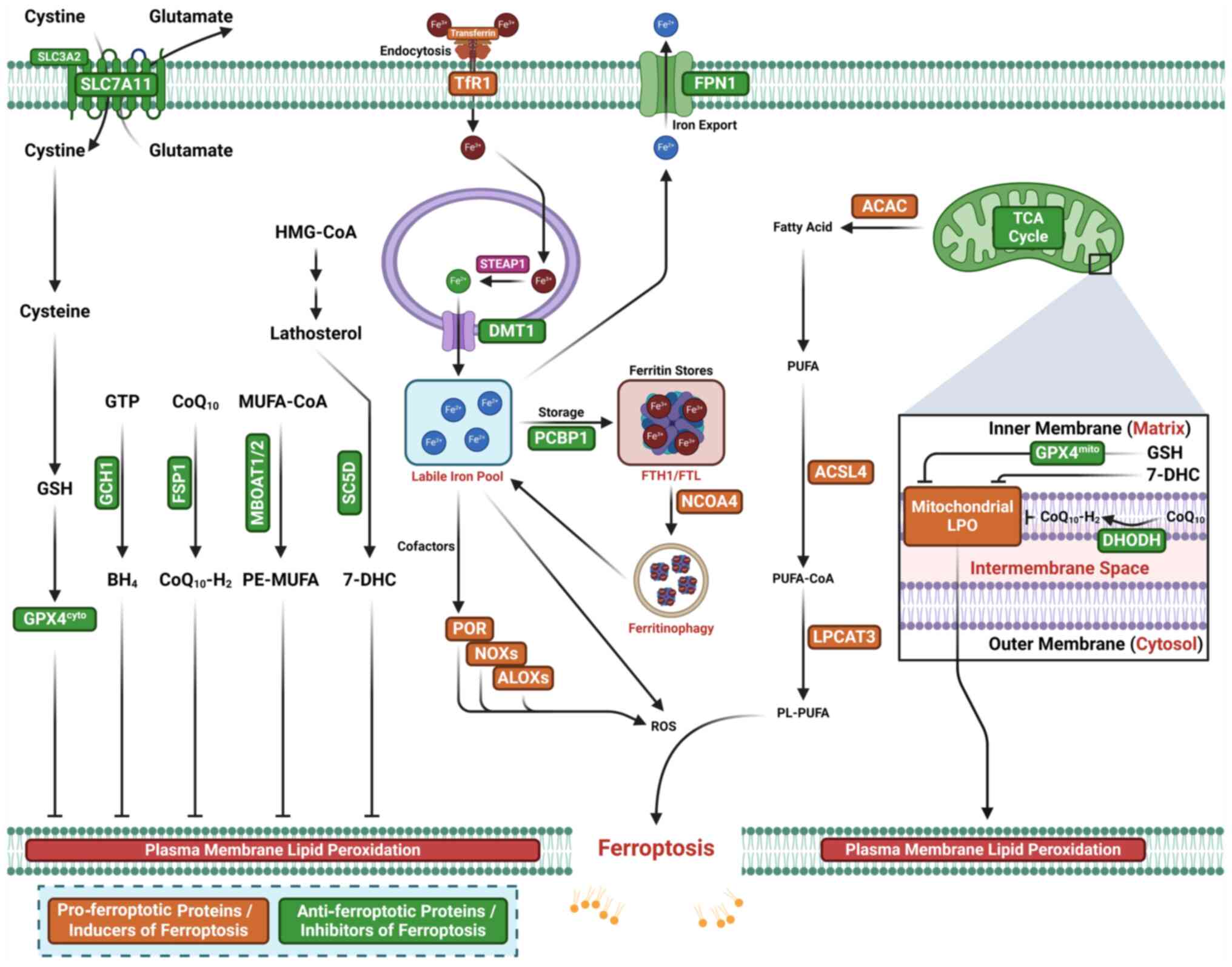
Ferroptosis was first identified in 2012 as a form
of RCD driven by iron-dependent lipid peroxidation (LPO) (34-39) (Fig.
1). This discovery stemmed from studying the mechanisms by
which small molecules induce cancer cell death. Research has since
identified specific small-molecule inhibitors of ferroptosis, which
have revealed additional intermediate nodes in the ferroptosis
signaling cascade (40). Three
essential elements, reactive oxygen species (ROS), oxidizable
lipids and LPO, drive the initiation and induction of ferroptosis
(41) (Fig. 2). An imbalance between ferroptosis
inhibitors and inducers enhances accumulation of lethal lipid
peroxides (lipid hydroperoxides) on cell membranes, leading to
membrane rupture and ferroptotic cell death (16,42-44).
The synthesis and peroxidation of polyunsaturated
fatty acid-containing phospholipids (PUFA-PLs), along with iron and
mitochondrial metabolism, are the primary inducers that initiate
ferroptosis (16,45-47).
Phospholipid peroxidation, a process that depends on
transition metal iron, PUFA-PL and ROS, induces ferroptosis
(34,48,49). Both LPO and the accumulation of
peroxidized lipids trigger ferroptosis (44,50). Iron chelation studies have
clarified the link between iron and ferroptosis, highlighting
interplay between lipids and iron (48,49,51). PUFA-PLs, which are highly
susceptible to peroxidation, serve as effective substrates for LPO
via enzymatic and non-enzymatic mechanisms (43). Initiation, propagation and
termination constitute the mechanism underlying PUFA-PL-mediated
LPO (52,53). The incorporation and formation of
peroxidized lipids in membrane phospholipids trigger ferroptosis
(54,55). In the first step of PUFA-PL
synthesis, acyl-CoA synthetase long-chain family member 4 (ACSL4)
ligates coenzyme A (CoA) to fatty acids, with a high preference for
arachidonic acid (AA) (54,55). Arachidonoyl-CoA is then esterified
in membranes by lysophosphatidylcholine acyltransferase 3, which
preferentially binds lysophosphatidylethanolamine and
lysophosphatidylcholine to produce phospholipids in the endoplasmic
reticulum (ER) (56).
Lipoxygenases (LOXs), NADPH oxidases (NOXs),
oxidoreductase cytochrome P450 reductase (POR) and NADH-cytochrome
b5 reductase (CYB5R1) are enzymes that generate oxidants to
initiate and induce LPO (57-64). LOXs, including 15-, 12- and 5-LOX,
are a class of enzymes that catalyze reactions through the
involvement of iron and other metals. LOXs facilitate double
oxidation of PUFAs, exacerbating cell membrane instability and
triggering ferroptosis (64,65). Specifically, 15-LOX, which has a
high selectivity for PUFA-PLs, oxidizes PUFA-PLs to PL-PUFA-OOH
(PL-OOH). POR and CYB5R1 induce ferroptosis by transferring
electrons to oxygen to produce hydrogen peroxide
(H2O2), which damages cell membranes by
oxidizing membrane phospholipids via Fenton reaction (57). NOXs directly produce ROS and cause
LPO (66). Mitochondria produce
substantial amounts of ROS, further contributing to initiation of
LPO and promoting ferroptosis (47,67). The interaction of iron with lipids
leads to LPO, producing lipid peroxides, PUFA-PL hydroperoxides,
peroxidized PUFA-PLs (PUFA-PL-OOH) and derivatives such as
4-hydroxynonenal and malondialdehyde (MDA) (44).
Iron induces ferroptosis by directly driving LPO via
a non-enzymatic Fenton reaction and serving as an essential
cofactor for iron-dependent peroxidases (the enzymatic LPO pathway)
(16,50,53,68,69). Iron exists in two states: Ferric
(Fe3+) and ferrous iron (Fe2+) (70). PUFA-PLs can react with ROS, such
as lipid peroxyl (LO•) or hydroxyl radicals
(HO•), via the Fenton reaction to produce PUFA-PL-OOH,
triggering LPO (71-73). When PUFA-PL-OOH is not neutralized
promptly, it can propagate peroxidation to neighboring PUFA-PLs in
the presence of labile iron. Inhibiting the iron exporter
ferroportin (74-76), promoting transferrin uptake
(77) or inducing autophagic
degradation of ferritin (78,79) increase the labile iron pool in
cells, enhancing cellular sensitivity to ferroptosis (50). In the enzymatic LPO pathway,
Fe2+ serves as a key cofactor for iron-dependent
peroxidases, initiating dioxygenation of membrane PUFA-PLs
(80,81). In this pathway, LOXs and POR use
labile iron and O2 to peroxidize PUFA-PLs, forming
PUFA-PL-OOH (43,58,62). Previous reviews provide further
details on lipid resources involved in ferroptosis: ACSL4 that
activates long-chain fatty acids by converting them into acyl-CoA
esters, lysophosphatidylcholine acyltransferase 3 (LPCAT3) that
incorporates PUFA into phosphatidylethanolamines (PEs), whereas the
other activates sterol O-acyltransferase 1-producing
PUFA-cholesteryl esters instead of PUFA-PEs), proxisomes that is
involved in fatty acid breakdown, hydrogen peroxide production and
PUFA plasmalogen biosynthesis, and lipophagy that selectively
degrades lipid droplets, releasing lipids for peroxidation
(41,53,82).
Specific ferroptosis inhibitors typically suppress
LPO to prevent unwanted ferroptosis (69). Ferroptosis inhibitors or defense
systems that directly neutralize lipid peroxides comprise
GPX4-dependent or -independent surveillance mechanisms with
specific subcellular localization (35).
MBOAT1/2-MUFA system is a newly identified
ferroptosis inhibitor independent of GPX4 and FSP1 (102). MBOAT1/2 inhibits ferroptosis in
this system (102).
Phosphatidylethanolamine (PE)-PUFAs are the preferred substrates
for LPO and dictate the ferroptosis sensitivity (103,104). MBOAT2 is a lyso-PL
acyltransferase that selectively transfers MUFAs to lyso-PE,
resulting in increased cellular PE-MUFA levels and decreased
cellular PE-PUFAs, thereby suppressing ferroptosis induction
(102).
The SC5D-7-DHC axis is a recently identified
ferroptosis inhibitor, with 7-DHC serving as a natural inhibitor of
ferroptosis (105,106). 7-DHC, generated in the ER, is
found in mitochondria and cell membranes along the cholesterol
synthesis pathway. It diverts the LPO pathway from phospholipids
and traps radicals, suppressing LPO and subsequent ferroptosis in
both mitochondria and the plasma membrane.
Increasing evidence indicates that dysregulated
epigenetic modifications contribute to cancer initiation and
progression through aberrant gene expression, protein alteration
and malignant transformation (107-109). ncRNAs have recently emerged as
key regulators of ferroptosis (28). Studies suggest that epigenetic
modifications modulate ferroptosis at transcriptional,
post-transcriptional and post-translational levels (39,110). Targeting epigenetic and
post-translational modifications that modulate ferroptosis may
offer new directions for cancer treatment (39,110). Recent research has shown that
ncRNAs modulate ferroptosis by regulating glutathione metabolism,
mitochondria-associated proteins, LPO and iron metabolism, thereby
impacting cancer biology (25-31). In cancer, ncRNAs influence
ferroptosis by modulating genes associated with ferroptosis
inducers or inhibitors, affecting pathways such as the SLC7A11-GPX4
axis, lipid, glutamine and iron metabolism and Nrf2 signaling axis
(28).
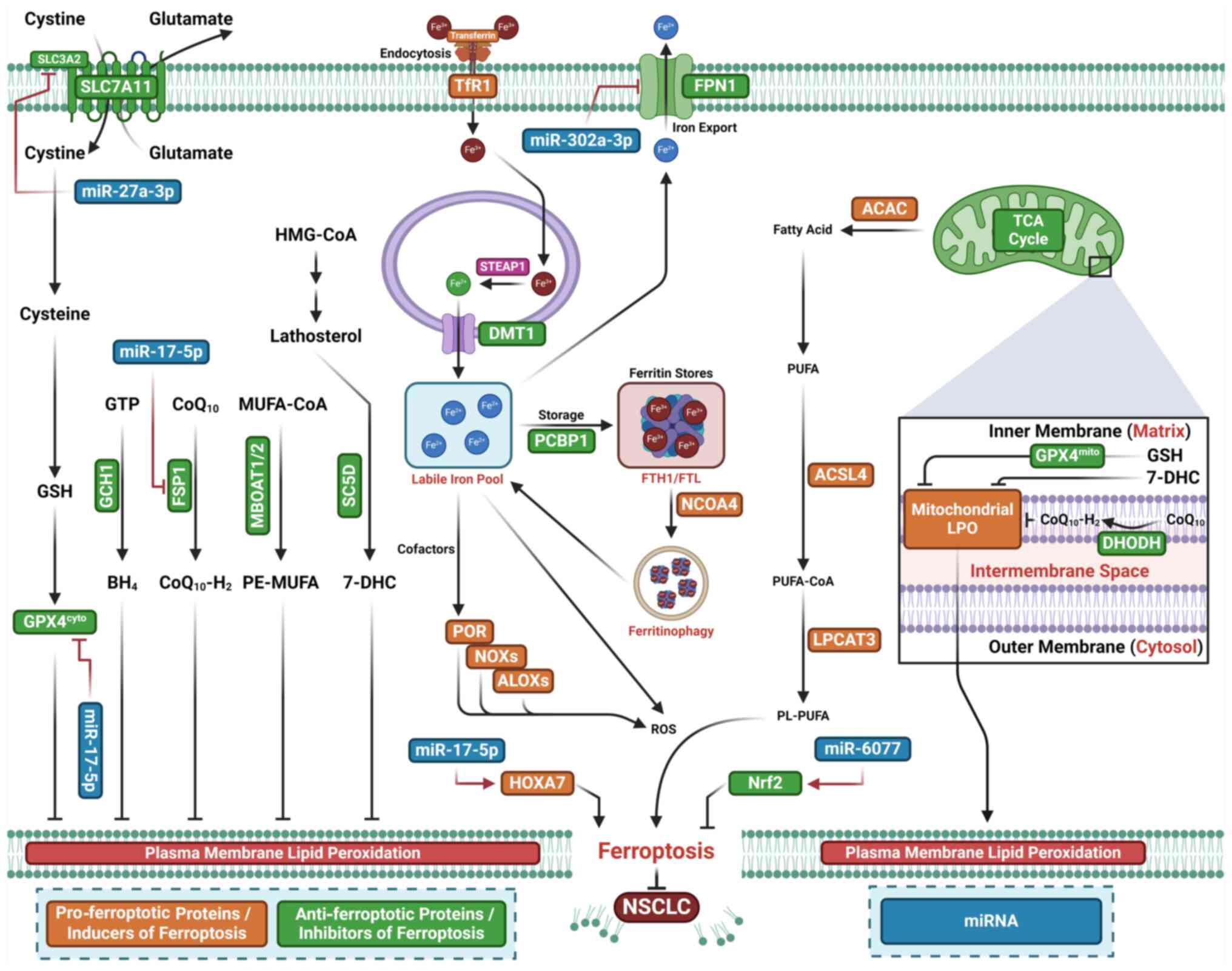
Decreased miR-324-3p expression is observed in
cisplatin-resistant A549 human lung cancer cells(DDP-A549)
(111) (Fig. 3; Table I). Overexpression of miR-324-3p
overcomes DDP resistance by targeting GPX4, where increased GPX4
expression reverses the miR-324-3p-induced sensitivity of DDP-A549
cells to DDP (111). miR-324-3p
enhances DDP-triggered ferroptosis in DDP-A549 cells and GPX4
inhibitor RSL3 mimics miR-324-3p by increasing DDP-A549 cell
sensitivity to DDP (111). Thus,
miR-324-3p overcomes DDP resistance by promoting ferroptosis and
suppressing GPX4 expression in NSCLC cells. Upregulation of
miR-4443 has also been observed in exosomes derived from
DDP-resistant NSCLC tumor tissues (112). Exosomal miR-4443 promotes
resistance in recipient cells by inhibiting DDP-mediated
ferroptosis through upregulation of FSP1 via m6A
modification mediated by m6A-methylase
methyltransferase-like protein 3 (METLL3) in vitro and
enhancing tumor growth in vivo (112). Furthermore, lncRNA HCP5 promotes
ferroptosis and limits brain metastasis by upregulating homeobox A7
(HOXA7) through competitive binding to miR-17-5p (113). Overexpression of miR-27a-3p,
however, inhibits erastin-induced ferroptosis by suppressing
SLC7A11 (114). miR-6077
prevents cisplatin/pemetrexed (CDDP/PEM)-mediated NSCLC cell death
through cyclin-dependent kinase inhibitor 1A
(CDKN1A)/cyclin-dependent kinase 1 (CDK1)-mediated cell cycle
arrest and ferroptosis suppression by inactivating Kelch-like
ECH-associated protein 1 (KEAP1), which subsequently increases
Nrf2-SLC7A11/NQO1 expression and leads to chemoresistance in
vitro and in vivo (115). Decreased expression of
miR-302a-3p has also been observed in human NSCLC cells and tissue.
miR-302a-3p inhibits cell proliferation and colony formation by
inducing ferroptosis in NSCLC cells (116). Conversely, silencing miR-302a-3p
inhibits erastin- or RSL3-induced ferroptosis and tumor suppression
(116). miR-302a-3p binds
ferroportin and decreases its protein expression; ferroportin
overexpression, in turn, prevents miR-302a-3p-triggered ferroptosis
and tumor suppression (116).
Additionally, miR-302a-3p increases NSCLC cell sensitivity to DDP
and paclitaxel. These results indicate that miR-302a-3p serves as a
tumor suppressor by inducing ferroptosis through ferroportin
targeting in NSCLC (116).
miR-139 overexpression enhances the radiosensitivity of NSCLC cells
in vitro and in vivo by targeting cJUN and
Karyopherin-2 α (KPNA2), which inhibits Nrf2 signaling and promotes
ionizing radiation-induced ferroptosis (117). Ionizing radiation induces
miR-139 expression via early growth response 1, a transcription
factor that binds to the promoter region to activate miR-139. These
results suggest that miR-139 serves as a radiosensitizer by
inhibiting Nrf2 expression in NSCLC (117).
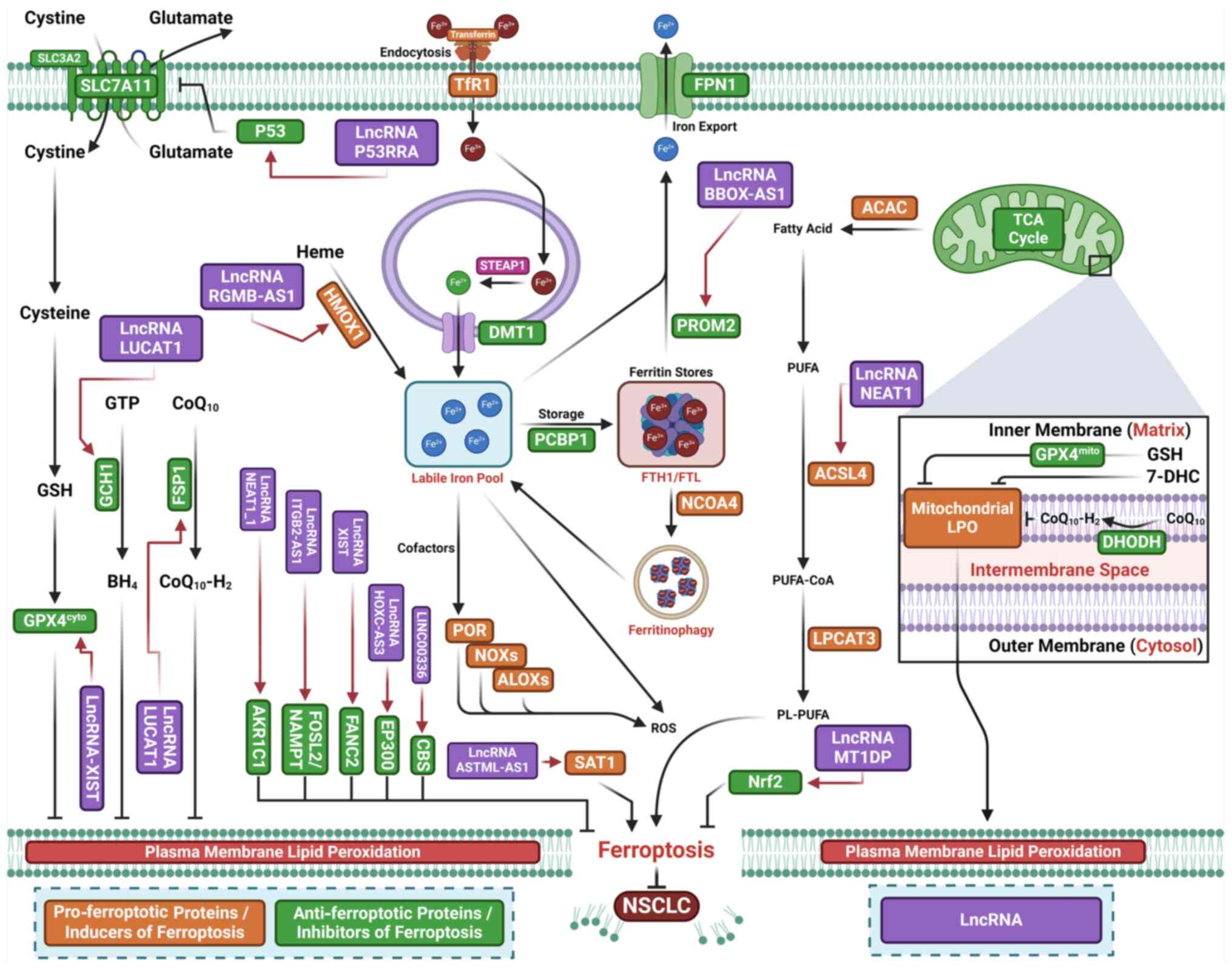
Increased expression of lncRNA p53 upregulated
regulator of P53 levels (PURPL) has been observed in lung cancer
cells and patient samples (118)
(Fig. 4; Table II). Overexpression of lncRNA
PURPL promotes M2 macrophage polarization and suppresses
ferroptosis by stabilizing SLC7A11 mRNA through regulation of
RNA-binding motif 4 (118).
Silencing SLC7A11 reverses the inhibitory effects of lncRNA PURPL
on M2 macrophage polarization by inducing macrophage ferroptosis
(118). Upregulation of lncRNA
tyrosine protein kinase transmembrane receptor 1 antisense RNA 1
(ROR1-AS1), insulin-like growth factor 2 mRNA-binding protein 1
(IGF2BP1) and SLC7A11 is associated with poor prognosis in lung
cancer. ROR1-AS1 stabilizes SLC7A11 mRNA through interaction with
IGF2BP1 (119).
Cancer-associated fibroblast (CAF)-derived exosomal ROR1-AS1
inhibits ferroptosis in lung cancer cells both in vitro and
in vivo (119).
Ferroptosis induced by overexpression of lncRNA ROR1-AS1 or IGF2BP1
is partially reversed by silencing IGF2BP1 or inhibiting SLC7A11 in
lung cancer cells. These results suggest that CAF-secreted exosomal
ROR1-AS1 inhibits ferroptosis by enhancing SLC7A11 expression or
interacting with IGF2BP1 (119).
Elevated levels of lncRNA-X-inactive specific transcript (XIST)
mRNA and protein expression are also observed in clinical tissues
and NSCLC cells (120).
Silencing lncRNA-XIST reduces NSCLC cell viability by inducing
ferroptosis, evidenced by increased ROS, MDA and Fe2+
levels and decreased expression of SLC7A11 and FANCD2.
Additionally. silencing XIST downregulates GPX4 in NSCLC cells
(120). Overexpression of GPX4
reverses ferroptosis and decreases cell viability caused by
lncRNA-XIST knockdown (120).
Increased expression of lncRNA Uc.339 has been observed in patients
with NSCLC (121). Uc.339
inhibits the maturation of miR-339 by competitively binding to
pri-miR-339. As miR-339 targets and downregulates SLC7A11
expression, Uc.339 promotes cell proliferation, migration and
invasion by inhibiting ferroptosis through upregulation of SLC7A11
via miR-339 downregulation (121). Silencing lncRNA nuclear
paraspeckle assembly transcript 1 (NEAT1) inhibits proliferation
and induces ferroptosis by upregulating ACSL4 and downregulating
SLC7A11/GPX4 (122). Silencing
lncRNA H19 promotes curcumenol-induced ferroptosis (123). Increased expression of lncRNA
lung cancer associated transcript 1 (LUCAT1) is observed in LUAD
and LUSC tissue (124). Elevated
lncRNA LUCAT1 expression is found in RSL3-treated A549 cells; this
increase is reversed by the ferroptosis inhibitor Fer-1.
Overexpression of LUCAT1 enhances cell proliferation and suppresses
RSL3- and erastin-induced ferroptosis while silencing LUCAT1
decreases cell proliferation and facilitates ferroptosis (124). Mechanistic study have shown that
silencing LUCAT1 downregulates FSP1 and GCH1 and upregulates
miR-34a-5p, thereby downregulating GCH1 (124). Together, these results suggest
that inhibition of LUCAT1 promotes ferroptosis by downregulating
GCH1 via upregulation of miR-34a-5p (124).
lncRNA γ-butyrobetaine hydroxylase 1-antisense 1
(BBOX1-AS1) promotes cell proliferation, migration and invasion by
inhibiting ferroptosis through post-transcriptional upregulation of
prominin 2 (PROM2) expression by miR-326 sponging (125). lncRNA repulsive guidance
molecule b-antisense 1 (RGMB-AS1) enhances ferroptosis by
interacting with HO-1, preventing its ubiquitination by E3 ligase
TRC8, which leads to increased HO-1 stability. Additionally,
RGMB-AS1 binds to and promotes acetyltransferase activity of
N-alpha-acetyltransferase 10, further contributing to ferroptosis
(126).
The upregulated lncRNA ITGB2-AS1 is found in
DDP-resistant NSCLC cells and cancer tissue from patients with
NSCLC. Silencing ITGB2-AS1 inhibits cell proliferation and enhances
apoptosis and ferroptosis in DDP-resistant NSCLC cells (137). ITGB2-AS1 promotes nicotinamide
phosphoribosyltransferase (NAMPT) expression by binding to FOS-like
antigen 2 (FOSL2), thereby suppressing p53 expression. FOSL2, a
member of the activator protein 1 transcription factor family,
enhances NSCLC malignancy (137). p53 increases cell sensitivity to
ferroptosis by transcriptionally suppressing SLC7A11 expression
(138). The cytosolic lncRNA
P53RRA functions as a tumor suppressor by inducing ferroptosis via
p53 activation (127).
Downregulation of P53RRA inhibits cancer progression by acting as a
tumor suppressor. Chromatin remodeling proteins CxxC finger protein
1 (Cfp1) and lymphoid-specific helicase (LSH) regulate P53RRA
expression. P53RRA binds to Ras GTPase-activating protein-binding
protein 1 (G3BP1) to form a P53RRA-G3BP1 cytosolic complex,
displacing p53 from a G3BP1 complex and promoting p53 retention in
the nucleus, which leads to cell cycle arrest, apoptosis and
ferroptosis (127). The lncRNA
metallothionein 1D pseudogene (MT1DP) increases sensitivity of
NSCLC cells to erastin-induced ferroptosis by downregulating Nrf2
via stabilization of miR-365a-3p (128).
Increased expression of the ferroptosis suppressor
aldo-keto reductase family 1 member C1 (AKR1C1) is found in
gefitinib-resistant LUAD cells and associated with poor prognosis
in patients with LUAD receiving first-generation epidermal growth
factor receptor-tyrosine kinase inhibitor therapy (129). Silencing AKR1C1 increases the
sensitivity of LUAD cells to gefitinib-induced ferroptosis
(129). Reduced miR-338-3p
expression leads to upregulation of AKR1C1 in gefitinib-resistant
LUAD cells (129). lncRNA
NEAT1_1 activates AKR1C1 by sponging miR-338-3p, thereby reducing
miR-338-3p-mediated inhibition of AKR1C1. These results suggest
that lncRNA NEAT1_1 promotes gefitinib resistance by suppressing
ferroptosis via the upregulation of AKR1C1 via miR-338-3p sponging
(129). Silencing lncRNA
ITGB2-AS1 overcomes NSCLC resistance to DDP in vivo by
inhibiting p53-mediated ferroptosis through activation of the
FOSL2-NAMPT axis (130).
Increased lncRNA HOXC-AS3 expression due to methylation promotes
its binding to EP300, thereby suppressing ferroptosis in NSCLC
cells (131). Upregulation of
lncRNA ASMTL-AS1 inhibits tumor growth in vitro and in
vivo by stabilizing spermidine/spermine N1-acetyltransferase 1,
thereby stimulating ferroptosis (132). LINC00336 promotes malignancy by
inhibiting ferroptosis via the absorption of miR6852, acting as a
competing endogenous RNA (ceRNA) to increase the mRNA levels of
cystathionine-β-synthase (133).
The lncRNA syndecan-binding protein 2-antisense RNA 1 (SDCBP2-AS1)
promotes cancer by inhibiting ferroptosis via sponging miR-656-3p,
directly targeting cysteine-rich transmembrane BMP regulator 1
(CRIM1). SDCBP2-AS1 inhibits ferroptosis via miR-656-3p, while
CRIM1 overexpression counteracts the effects of miR-656-3p on
ferroptosis (134). lncRNA
opioid growth factor receptor pseudogene 1 promotes NSCLC cell
proliferation by inhibiting ferroptosis and suppressing miR-299-3p,
leading to upregulation of SLC38A1 expression (135). β-elemene, a primary bioactive
compound derived from Curcuma wenyujin, enhances sensitivity
of cancer cells to erlotinib by inducing ferroptosis via
upregulation of lncRNA H19 (136).
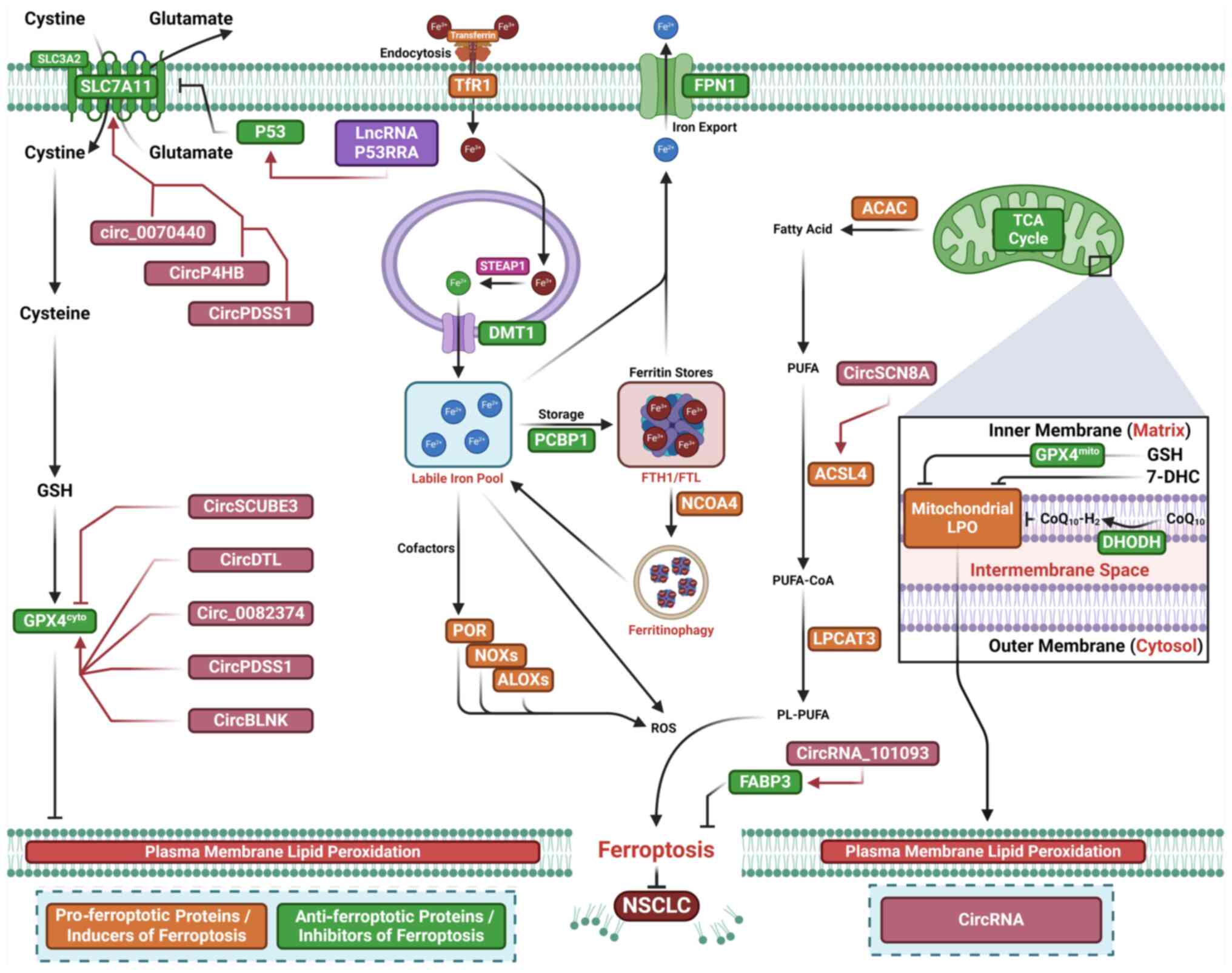
Increased circ_0070440 expression is observed in
LUAD cells. Silencing circ_0070440 inhibits cell proliferation and
enhances apoptosis and ferroptosis in LUAD cells (139). circ_0070440 promotes malignant
progression by suppressing ferroptosis through upregulation of
SLC7A11 expression via miR-485-5p sponging (139) (Fig. 5; Table III). Increased expression of
circP4HB is observed in LUAD. circP4HB inhibits erastin-induced
ferroptosis by promoting GSH synthesis (140). Mechanistic study indicates that
circP4HB upregulates SLC7A11 by serving as a competing endogenous
RNA and modulating miR-1184 expression (140). circPDSS1 expression is elevated
in NSCLC cells (141). Silencing
circPDSS1 inhibits NSCLC cell viability and induces ferroptosis
(141). Mechanistic study
suggest that circPDSS1 functions as a competing endogenous RNA
'sponge' to negatively regulate miR-137 expression (141). Silencing circPDSS1 also
downregulates GPX4 and the glutamate-cysteine ligase catalytic
subunit (141).
circ_0082374 promotes tumorigenesis in NSCLC by
suppressing ferroptosis via upregulation of GPX4 via miR-491-5p
sponging (142). Increased
circ_0082374 levels are observed in NSCLC tissue and cells.
Silencing circ_0082374 inhibits cell proliferation and tumor
metastasis while facilitating ferroptosis by decreasing iron and
LPO levels in NSCLC (142).
Mechanistic study show that circ_0082374 enhances GPX4 expression
by sequestering miR-491-5p (142). In LUAD samples and cell lines,
decreased circRNA signal peptide-CUB-EGF domain-containing protein
3 (circSCUBE3) expression is noted (143). Overexpression of circSCUBE3
decreases GSH levels and GSH/GSSG ratio in LUAD cells, while loss
of circSCUBE3 reverses erastin-induced ferroptosis (143). circSCUBE3 downregulates GPX4
expression in LUAD cells, and silencing GPX4 counteracts the
ferroptosis induced by circSCUBE3 deficiency (143). circSCUBE3 downregulates GPX4
expression by competitively interacting with CREB, which activates
GPX4 transcription. These results suggest that circSCUBE3 inhibits
LUAD development by promoting ferroptosis via the CREB/GPX4/GSH
axis (143). Increased circDTL
expression has been detected in NSCLC cells (144). Silencing circDTL promotes
apoptosis and ferroptosis in NSCLC cells (144) and enhances their sensitivity to
chemotherapy, thus inhibiting tumor growth in vivo. CircDTL
acts as an oncogene by suppressing ferroptosis and increasing GPX4
expression via miR-1287-5p sponging (144).
Downregulated circSCN8A expression has been
observed in NSCLC tissue and cells. Decreased circSCN8A expression
is positively associated with aggressive clinicopathological
characteristics and predicts poor prognosis in patients with NSCLC
(145). circSCN8A suppresses
tumor growth in vivo and inhibits cell proliferation,
migration, invasion and epithelial-mesenchymal transition in
vitro. circSCN8A enhances ferroptosis in NSCLC cells.
Mechanistic studies revealed that circSCN8A functions as a ceRNA by
sponging miR-1290 to upregulate ACSL4 (145). Silencing ACSL4 or overexpressing
miR-1290 reverses circSCN8A-induced ferroptosis and inhibits cell
proliferation and tumor metastasis (145). These results indicate that
circSCN8A suppresses malignant progression by inducing ferroptosis
through miR-1290 sponging to enhance ACSL4 expression (145).
Plasma exosomes from patients with LUAD show
decreased LPO and reduced sensitivity of LUAD cells to ferroptosis
(146). Exosomal circRNA_101093
maintains elevated intracellular circRNA_101093 levels in LUAD
cells to modulate AA (146).
circRNA_101093 binds to and upregulates fatty acid-binding protein
3 (FABP3), which transports AA and enhances its reaction with
taurine (146). These results
indicate that circRNA_101093 inhibits ferroptosis by upregulating
FABP3 and suggest that blocking exosomes may be a novel therapeutic
approach for LUAD (146).
The present review summarizes the role of
ncRNA-mediated epigenetic modulation of ferroptosis in NSCLC and
the roles of miRNAs, lncRNA and circRNA in regulating ferroptosis
in NSCLC cells. However, studies on ncRNA-induced epigenetic
modifications that modulate ferroptosis in NSCLC are in their early
stages and the present study has limitations. First, other ncRNAs
that modulate ferroptosis require further exploration. Second,
while ncRNA-mediated epigenetic modifications of ferroptosis in
NSCLC have been identified, it remains unclear whether
small-molecule compounds effectively target these ncRNA-mediated
mechanisms. Third, ncRNAs modulate the crosstalk between
ferroptosis and regulated cell death mechanisms in cancer (147). However, the role of ncRNAs in
the interplay between ferroptosis and novel RCD mechanisms, such as
cuproptosis, in NSCLC remains largely unknown (148). The aforementioned studies
primarily focused on classical pathways of ferroptosis, such as the
ACSL4-dependent LPO or SLC7A11-GPX4 system. However, the effects of
other ferroptosis inhibitors, including DHODH-CoQH2,
MBOAT1/2-MUFA, GCH1-BH4 and SC5D-7-DHC, are still poorly
understood. Fifth, many ncRNAs directly modulate ferroptosis by
regulating ferroptosis-associated genes involved in antioxidant
defense and iron and lipid metabolism or indirectly target
ferroptosis modulators, such as activated transcription factor 4 in
NSCLC (149). However, whether
ncRNAs modulate other transcription factors, including Nrf2, remain
poorly understood.
The clinical application of RNA-based therapeutics
has made progress, primarily using antisense oligonucleotides and
small interfering RNA (150).
However, no lncRNA-based therapeutics or circRNA-targeted
treatments have entered clinical practice, although miRNA mimics
and anti-miRNA therapeutics are under clinical development
(150-152). Emerging evidence suggests that
small molecule compounds can target ncRNAs for cancer treatment
(153). Small molecules
targeting miRNA (154,155) and lncRNA (136,156) that regulate ferroptosis may
offer new opportunities for NSCLC therapy. However, no small
molecules targeting miRNAs and lncRNAs involved in regulating
ferroptosis have yet been introduced for clinical use in NSCLC
treatment.
Collectively, ncRNAs modulate the tumor burden in
NSCLC by regulating ferroptosis through proteins or genes
associated with pro- and anti-ferroptotic factors. The present
review summarizes advances in understanding the ncRNA-mediated
mechanisms affecting ferroptosis in NSCLC, highlighting a novel
therapeutic avenue for NSCLC through the ncRNA-ferroptosis
axis.
Not applicable.
YW, JC, and MD designed and conceived the review.
ZZ, HW and JSF edited the manuscript. YW, YL and YC wrote the
manuscript. YW and JSF generated the figures. Data authentication
is not applicable. All authors have read and approved the final
manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
No applicable.
The present study was supported by the Wu Jieping Medical
Foundation (320.6750.2024-13-59), Science Foundation of Aerospace
Center hospital (grant nos. YN202402 and YN202423), Science
Foundation of AMHT (grant nos. 2022YK01 and 2022YK27) and Grant of
Chinese Medicine Education Association (grant no. 2022KTZ019).
|
1
|
Adhikari S, Bhattacharya A, Adhikary S,
Singh V, Gadad SS, Roy S and Da C: The paradigm of drug resistance
in cancer: An epigenetic perspective. Biosci Rep.
42:BSR202118122022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bukowski K, Kciuk M and Kontek R:
Mechanisms of multi-drug resistance in cancer chemotherapy. Int J
Mol Sci. 21:32332020. View Article : Google Scholar
|
|
3
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Leiter A, Veluswamy RR and Wisnivesky JP:
The global burden of lung cancer: Current status and future trends.
Nat Rev Clin Oncol. 20:624–639. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rotow J and Bivona TG: Understanding and
targeting resistance mechanisms in NSCLC. Nat Rev Cancer.
17:637–658. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
LoPiccolo J, Gusev A, Christiani DC and
Jänne PA: Lung cancer in patients who have never smoked-an emerging
disease. Nat Rev Clin Oncol. 21:121–146. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Siegel RL, Miller KD, Wagle NS and Jemal
A: Cancer statistics, 2023. CA Cancer J Clin. 73:17–48. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Otano I, Ucero AC, Zugazagoitia J and
Paz-Ares L: At the crossroads of immunotherapy for
oncogene-addicted subsets of NSCLC. Nat Rev Clin Oncol. 20:143–159.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Soerjomataram I and Bray F: Planning for
tomorrow: Global cancer incidence and the role of prevention
2020-2070. Nat Rev Clin Oncol. 18:663–672. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hirsch FR, Scagliotti GV, Mulshine JL,
Kwon R, Curran WJ Jr, Wu YL and Paz-Ares L: Lung cancer: Current
therapies and new targeted treatments. Lancet. 389:299–311. 2017.
View Article : Google Scholar
|
|
11
|
Nagasaka M and Gadgeel SM: Role of
chemotherapy and targeted therapy in early-stage non-small cell
lung cancer. Expert Rev Anticancer Ther. 18:63–70. 2018. View Article : Google Scholar
|
|
12
|
Yin JY, Li X, Zhou HH and Liu ZQ:
Pharmacogenomics of platinum-based chemotherapy sensitivity in
NSCLC: Toward precision medicine. Pharmacogenomics. 17:1365–1378.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Y, Hu J, Fleishman JS, Li Y, Ren Z,
Wang J, Feng Y, Chen J and Wang H: Inducing ferroptosis by
traditional medicines: A novel approach to reverse chemoresistance
in lung cancer. Front Pharmacol. 15:12901832024. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Herbst RS, Morgensztern D and Boshoff C:
The biology and management of non-small cell lung cancer. Nature.
553:446–454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lim ZF and Ma PC: Emerging insights of
tumor heterogeneity and drug resistance mechanisms in lung cancer
targeted therapy. J Hematol Oncol. 12:1342019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lei G, Zhuang L and Gan B: Targeting
ferroptosis as a vulnerability in cancer. Nat Rev Cancer.
22:381–396. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kobayashi H, Yoshimoto C, Matsubara S,
Shigetomi H and Imanaka S: A comprehensive overview of recent
developments on the mechanisms and pathways of ferroptosis in
cancer: The potential implications for therapeutic strategies in
ovarian cancer. Cancer Drug Resist. 6:547–566. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yunchu Y, Miyanaga A and Seike M:
Integrative analysis of ferroptosis-related genes in small cell
lung cancer for the identification of biomarkers and therapeutic
targets. Front Biosci (Landmark Ed). 28:1252023. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo J, Xu B, Han Q, Zhou H, Xia Y, Gong C,
Dai X, Li Z and Wu G: Ferroptosis: A novel anti-tumor action for
cisplatin. Cancer Res Treat. 50:445–460. 2018. View Article : Google Scholar :
|
|
20
|
Lei G, Zhang Y, Koppula P, Liu X, Zhang J,
Lin SH, Ajani JA, Xiao Q, Liao Z, Wang H and Gan B: The role of
ferroptosis in ionizing radiation-induced cell death and tumor
suppression. Cell Res. 30:146–162. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun X, Ou Z, Chen R, Niu X, Chen D, Kang R
and Tang D: Activation of the p62-Keap1-NRF2 pathway protects
against ferroptosis in hepatocellular carcinoma cells. Hepatology.
63:173–184. 2016. View Article : Google Scholar
|
|
22
|
Wang W, Green M, Choi JE, Gijón M, Kennedy
PD, Johnson JK, Liao P, Lang X, Kryczek I, Sell A, et al: CD8(+) T
cells regulate tumour ferroptosis during cancer immunotherapy.
Nature. 569:270–274. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Wu X, Ren Z, Li Y, Zou W, Chen J
and Wang H: Overcoming cancer chemotherapy resistance by the
induction of ferroptosis. Drug Resist Updat. 66:1009162023.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin X, Wu Z, Hu H, Luo ML and Song E:
Non-coding RNAs rewire cancer metabolism networks. Semin Cancer
Biol. 75:116–126. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Balihodzic A, Prinz F, Dengler MA, Calin
GA, Jost PJ and Pichler M: Non-coding RNAs and ferroptosis:
Potential implications for cancer therapy. Cell Death Differ.
29:1094–1106. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ensoy M, Bumin ZS, Jama HA and
Cansaran-Duman D: The regulation role of ferroptosis mechanism of
anti-cancer drugs and noncoding RNAs. Curr Med Chem. 30:1638–1656.
2023. View Article : Google Scholar
|
|
27
|
Luo Y, Huang Q, He B, Liu Y, Huang S and
Xiao J: Regulation of ferroptosis by non-coding RNAs in the
development and treatment of cancer (Review). Oncol Rep. 45:29–48.
2021. View Article : Google Scholar
|
|
28
|
Valashedi MR, Bamshad C, Najafi-Ghalehlou
N, Nikoo A, Tomita K, Kuwahara Y, Sato T, Roushandeh AM and
Roudkenar MH: Non-coding RNAs in ferroptotic cancer cell death
pathway: Meet the new masters. Hum Cell. 35:972–994. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang D, Tang L, Zhang Y, Ge G, Jiang X, Mo
Y, Wu P, Deng X, Li L, Zuo S, et al: Regulatory pathways and drugs
associated with ferroptosis in tumors. Cell Death Dis. 13:5442022.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xie B and Guo Y: Molecular mechanism of
cell ferroptosis and research progress in regulation of ferroptosis
by noncoding RNAs in tumor cells. Cell Death Discov. 7:1012021.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zuo YB, Zhang YF, Zhang R, Tian JW, Lv XB,
Li R, Li SP, Cheng MD, Shan J, Zhao Z and Xin H: Ferroptosis in
cancer progression: Role of noncoding RNAs. Int J Biol Sci.
18:1829–1843. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang H, Fleishman JS, Cheng S, Wang W, Wu
F and Wang Y and Wang Y: Epigenetic modification of ferroptosis by
non-coding RNAs in cancer drug resistance. Mol Cancer. 23:1772024.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sun L, Cao H, Wang Y and Wang H:
Regulating ferroptosis by noncoding RNAs in hepatocellular
carcinoma. Biol Direct. 19:802024. View Article : Google Scholar
|
|
34
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gu Y, Li Y, Wang J, Zhang L, Zhang J and
Wang Y: Targeting ferroptosis: Paving new roads for drug design and
discovery. Eur J Med Chem. 247:1150152023. View Article : Google Scholar
|
|
36
|
Huo L, Liu C, Yuan Y, Liu X and Cao Q:
Pharmacological inhibition of ferroptosis as a therapeutic target
for sepsis-associated organ damage. Eur J Med Chem. 257:1154382023.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Stockwell BR: Ferroptosis turns 10:
Emerging mechanisms, physiological functions, and therapeutic
applications. Cell. 185:2401–2421. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yin L, Liu P, Jin Y, Ning Z, Yang Y and
Gao H: Ferroptosis-related small-molecule compounds in cancer
therapy: Strategies and applications. Eur J Med Chem.
244:1148612022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang Y, Hu J, Wu S, Fleishman JS, Li Y, Xu
Y, Zou W, Wang J, Feng Y, Chen J and Wang H: Targeting epigenetic
and posttranslational modifications regulating ferroptosis for the
treatment of diseases. Signal Transduct Target Ther. 8:4492023.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dixon SJ and Olzmann JA: The cell biology
of ferroptosis. Nat Rev Mol Cell Biol. 25:424–442. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dai E, Chen X, Linkermann A, Jiang X, Kang
R, Kagan VE, Bayir H, Yang WS, Garcia-Saez AJ, Ioannou MS, et al: A
guideline on the molecular ecosystem regulating ferroptosis. Nat
Cell Biol. 26:1447–1457. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen X, Kang R, Kroemer G and Tang D:
Ferroptosis in infection, inflammation, and immunity. J Exp Med.
218:e202105182021. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hadian K and Stockwell BR: SnapShot:
Ferroptosis. Cell. 181:1188–1188.e1. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pope LE and Dixon SJ: Regulation of
ferroptosis by lipid metabolism. Trends Cell Biol. 33:1077–1087.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jiang X, Stockwell BR and Conrad M:
Ferroptosis: Mechanisms, biology and role in disease. Nat Rev Mol
Cell Biol. 22:266–282. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gan B: Mitochondrial regulation of
ferroptosis. J Cell Biol. 220:e2021050432021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gao M, Yi J, Zhu J, Minikes AM, Monian P,
Thompson CB and Jiang X: Role of mitochondria in ferroptosis. Mol
Cell. 73:354–363.e3. 2019. View Article : Google Scholar :
|
|
48
|
Stockwell BR, Angeli JP, Bayir H, Bush AI,
Conrad M, Dixon SJ, Fulda S, Gascón S, Hatzios SK, Kagan VE, et al:
Ferroptosis: A regulated cell death nexus linking metabolism, redox
biology, and disease. Cell. 171:273–285. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang DD: Ironing out the details of
ferroptosis. Nat Cell Biol. 26:1386–1393. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liang D, Minikes AM and Jiang X:
Ferroptosis at the intersection of lipid metabolism and cellular
signaling. Mol Cell. 82:2215–2227. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Doll S and Conrad M: Iron and ferroptosis:
A still ill-defined liaison. IUBMB Life. 69:423–434. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Helberg J and Pratt DA: Autoxidation vs.
antioxidants-the fight for forever. Chem Soc Rev. 50:7343–7358.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Conrad M and Pratt DA: The chemical basis
of ferroptosis. Nat Chem Biol. 15:1137–1147. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kang MJ, Fujino T, Sasano H, Minekura H,
Yabuki N, Nagura H, Iijima H and Yamamoto TT: A novel
arachidonate-preferring acyl-CoA synthetase is present in
steroidogenic cells of the rat adrenal, ovary, and testis. Proc
Natl Acad Sci USA. 94:2880–2884. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kim JW, Lee JY, Oh M and Lee EW: An
integrated view of lipid metabolism in ferroptosis revisited via
lipidomic analysis. Exp Mol Med. 55:1620–1631. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hishikawa D, Shindou H, Kobayashi S,
Nakanishi H, Taguchi R and Shimizu T: Discovery of a
lysophospholipid acyltransferase family essential for membrane
asymmetry and diversity. Proc Natl Acad Sci USA. 105:2830–2835.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Yan B, Ai Y, Sun Q, Ma Y, Cao Y, Wang J,
Zhang Z and Wang X: Membrane damage during ferroptosis is caused by
oxidation of phospholipids catalyzed by the oxidoreductases POR and
CYB5R1. Mol Cell. 81:355–369.e10. 2021. View Article : Google Scholar
|
|
58
|
Zou Y, Li H, Graham ET, Deik AA, Eaton JK,
Wang W, Sandoval-Gomez G, Clish CB, Doench JG and Schreiber SL:
Cytochrome P450 oxidoreductase contributes to phospholipid
peroxidation in ferroptosis. Nat Chem Biol. 16:302–309. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Poursaitidis I, Wang X, Crighton T,
Labuschagne C, Mason D, Cramer SL, Triplett K, Roy R, Pardo OE,
Seckl MJ, et al: Oncogene-selective sensitivity to synchronous cell
death following modulation of the amino acid nutrient cystine. Cell
Rep. 18:2547–2556. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yang WH, Ding CC, Sun T, Rupprecht G, Lin
CC, Hsu D and Chi JT: The hippo pathway effector TAZ regulates
ferroptosis in renal cell carcinoma. Cell Rep. 28:2501–2508.e4.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chu B, Kon N, Chen D, Li T, Liu T, Jiang
L, Song S, Tavana O and Gu W: ALOX12 is required for p53-mediated
tumour suppression through a distinct ferroptosis pathway. Nat Cell
Biol. 21:579–591. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Shah R, Shchepinov MS and Pratt DA:
Resolving the role of lipoxygenases in the initiation and execution
of ferroptosis. ACS Cent Sci. 4:387–396. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Anthonymuthu TS, Tyurina YY, Sun WY,
Mikulska-Ruminska K, Shrivastava IH, Tyurin VA, Cinemre FB, Dar HH,
VanDemark AP, Holman TR, et al: Resolving the paradox of
ferroptotic cell death: Ferrostatin-1 binds to 15LOX/PEBP1 complex,
suppresses generation of peroxidized ETE-PE, and protects against
ferroptosis. Redox Biol. 38:1017442021. View Article : Google Scholar
|
|
64
|
Yang WS, Kim KJ, Gaschler MM, Patel M,
Shchepinov MS and Stockwell BR: Peroxidation of polyunsaturated
fatty acids by lipoxygenases drives ferroptosis. Proc Natl Acad Sci
USA. 113:E4966–E4975. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Haeggström JZ and Funk CD: Lipoxygenase
and leukotriene pathways: Biochemistry, biology, and roles in
disease. Chem Rev. 111:5866–5898. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Alvarez SW, Sviderskiy VO, Terzi EM,
Papagiannakopoulos T, Moreira AL, Adams S, Sabatini DM, Birsoy K
and Possemato R: NFS1 undergoes positive selection in lung tumours
and protects cells from ferroptosis. Nature. 551:639–643. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Gaschler MM, Hu F, Feng H, Linkermann A,
Min W and Stockwell BR: Determination of the subcellular
localization and mechanism of action of ferrostatins in suppressing
ferroptosis. ACS Chem Biol. 13:1013–1020. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chen X, Li J, Kang R, Klionsky DJ and Tang
D: Ferroptosis: Machinery and regulation. Autophagy. 17:2054–2081.
2021. View Article : Google Scholar :
|
|
69
|
Hassannia B, Vandenabeele P and Berghe TV:
Targeting ferroptosis to iron out cancer. Cancer Cell. 35:830–849.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Jacquemyn J, Ralhan I and Ioannou MS:
Driving factors of neuronal ferroptosis. Trends Cell Biol.
34:535–546. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Ayala A, Muñoz MF and Argüelles S: Lipid
peroxidation: Production, metabolism, and signaling mechanisms of
malondialdehyde and 4-hydroxy-2-nonenal. Oxid Med Cell Longev.
2014:3604382014. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Dos Santos AF, Fazeli G, da Silva TN and
Angeli JP: Ferroptosis: Mechanisms and implications for cancer
development and therapy response. Trends Cell Biol. 33:1062–1076.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ryter SW, Kim HP, Hoetzel A, Park JW,
Nakahira K, Wang X and Choi AM: Mechanisms of cell death in
oxidative stress. Antioxid Redox Signal. 9:49–89. 2007. View Article : Google Scholar
|
|
74
|
Bao WD, Pang P, Zhou XT, Hu F, Xiong W,
Chen K, Wang J, Wang F, Xie D, Hu YZ, et al: Loss of ferroportin
induces memory impairment by promoting ferroptosis in Alzheimer's
disease. Cell Death Differ. 28:1548–1562. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Chen PH, Wu J, Ding CC, Lin CC, Pan S,
Bossa N, Xu Y, Yang WH, Mathey-Prevot B and Chi J: Kinome screen of
ferroptosis reveals a novel role of ATM in regulating iron
metabolism. Cell Death Differ. 27:1008–1022. 2020. View Article : Google Scholar :
|
|
76
|
Geng N, Shi BJ, Li SL, Zhong ZY, Li YC,
Xua WL, Zhou H and Cai JH: Knockdown of ferroportin accelerates
erastin-induced ferroptosis in neuroblastoma cells. Eur Rev Med
Pharmacol Sci. 22:3826–3836. 2018.PubMed/NCBI
|
|
77
|
Gao M, Monian P, Quadri N, Ramasamy R and
Jiang X: Glutaminolysis and transferrin regulate ferroptosis. Mol
Cell. 59:298–308. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Gao M, Monian P, Pan Q, Zhang W, Xiang J
and Jiang X: Ferroptosis is an autophagic cell death process. Cell
Res. 26:1021–1032. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Hou W, Xie Y, Song X, Sun X, Lotze MT, Zeh
HJ III, Kang R and Tang D: Autophagy promotes ferroptosis by
degradation of ferritin. Autophagy. 12:1425–1428. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Chen X, Yu C, Kang R and Tang D: Iron
metabolism in ferroptosis. Front Cell Dev Biol. 8:5902262020.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
David S, Jhelum P, Ryan F, Jeong SY and
Kroner A: Dysregulation of iron homeostasis in the central nervous
system and the role of ferroptosis in neurodegenerative disorders.
Antioxid Redox Signal. 37:150–170. 2022. View Article : Google Scholar
|
|
82
|
Stockwell BR and Jiang X: The chemistry
and biology of ferroptosis. Cell Chem Biol. 27:365–375. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Sun Y, Xia X, Basnet D, Zheng JC, Huang J
and Liu J: Mechanisms of ferroptosis and emerging links to the
pathology of neurodegenerative diseases. Front Aging Neurosci.
14:9041522022. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Brigelius-Flohé R and Flohé L: Regulatory
phenomena in the glutathione peroxidase superfamily. Antioxid Redox
Signal. 33:498–516. 2020. View Article : Google Scholar
|
|
85
|
Brigelius-Flohé R and Maiorino M:
Glutathione peroxidases. Biochim Biophys Acta. 1830:3289–3303.
2013. View Article : Google Scholar
|
|
86
|
Seibt TM, Proneth B and Conrad M: Role of
GPX4 in ferroptosis and its pharmacological implication. Free Radic
Biol Med. 133:144–152. 2019. View Article : Google Scholar
|
|
87
|
Ursini F, Maiorino M, Valente M, Ferri L
and Gregolin C: Purification from pig liver of a protein which
protects liposomes and biomembranes from peroxidative degradation
and exhibits glutathione peroxidase activity on phosphatidylcholine
hydroperoxides. Biochim Biophys Acta. 710:197–211. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Forcina GC and Dixon SJ: GPX4 at the
crossroads of lipid homeostasis and ferroptosis. Proteomics.
19:e18003112019. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Angeli JP, Schneider M, Proneth B, Tyurina
YY, Tyurin VA, Hammond VJ, Herbach N, Aichler M, Walch A,
Eggenhofer E, et al: Inactivation of the ferroptosis regulator Gpx4
triggers acute renal failure in mice. Nat Cell Biol. 16:1180–1191.
2014. View Article : Google Scholar
|
|
90
|
Ingold I, Berndt C, Schmitt S, Doll S,
Poschmann G, Buday K, Roveri A, Peng X, Freitas FP, Seibt T, et al:
Selenium utilization by GPX4 is required to prevent
hydroperoxide-induced ferroptosis. Cell. 172:409–422.e21. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yang WS, SriRamaratnam R, Welsch ME,
Shimada K, Skouta R, Viswanathan VS, Cheah JH, Clemons PA, Shamji
AF, Clish C, et al: Regulation of ferroptotic cancer cell death by
GPX4. Cell. 156:317–331. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Koppula P, Zhang Y, Zhuang L and Gan B:
Amino acid transporter SLC7A11/xCT at the crossroads of regulating
redox homeostasis and nutrient dependency of cancer. Cancer Commun
(Lond). 38:122018.PubMed/NCBI
|
|
93
|
Sato H, Tamba M, Ishii T and Bannai S:
Cloning and expression of a plasma membrane cystine/glutamate
exchange transporter composed of two distinct proteins. J Biol
Chem. 274:11455–11458. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Bersuker K, Hendricks JM, Li Z, Magtanong
L, Ford B, Tang PH, Roberts MA, Tong B, Maimone TJ, Zoncu R, et al:
The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit
ferroptosis. Nature. 575:688–692. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Doll S, Freitas FP, Shah R, Aldrovandi M,
da Silva MC, Ingold I, Grocin AG, da Silva TN, Panzilius E, Scheel
CH, et al: FSP1 is a glutathione-independent ferroptosis
suppressor. Nature. 575:693–698. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Nakamura T, Hipp C, Mourão ASD, Borggräfe
J, Aldrovandi M, Henkelmann B, Wanninger J, Mishima E, Lytton E,
Emler D, et al: Phase separation of FSP1 promotes ferroptosis.
Nature. 619:371–377. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Dai E, Zhang W, Cong D, Kang R, Wang J and
Tang D: AIFM2 blocks ferroptosis independent of ubiquinol
metabolism. Biochem Biophys Res Commun. 523:966–971. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Pedrera L, Espiritu RA, Ros U, Weber J,
Schmitt A, Stroh J, Hailfinger S, von Karstedt S and García-Sáez
AJ: Ferroptotic pores induce Ca(2+) fluxes and ESCRT-III activation
to modulate cell death kinetics. Cell Death Differ. 28:1644–1657.
2021. View Article : Google Scholar
|
|
99
|
Kraft V, Bezjian CT, Pfeiffer S,
Ringelstetter L, Müller C, Zandkarimi F, Merl-Pham J, Bao X,
Anastasov N, Kössl J, et al: GTP Cyclohydrolase
1/Tetrahydrobiopterin counteract ferroptosis through lipid
remodeling. ACS Cent Sci. 6:41–53. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Soula M, Weber RA, Zilka O, Alwaseem H, La
K, Yen F, Molina H, Garcia-Bermudez J, Pratt DA and Birso K:
Metabolic determinants of cancer cell sensitivity to canonical
ferroptosis inducers. Nat Chem Biol. 16:1351–1360. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Mao C, Liu X, Zhang Y, Lei G, Yan Y, Lee
H, Koppula P, Wu S, Zhuang L, Fang B, et al: DHODH-mediated
ferroptosis defence is a targetable vulnerability in cancer.
Nature. 593:586–590. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Liang D, Feng Y, Zandkarimi F, Wang H,
Zhang Z, Kim J, Cai Y, Gu W, Stockwell BR and Jiang X: Ferroptosis
surveillance independent of GPX4 and differentially regulated by
sex hormones. Cell. 186:2748–2764. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Doll S, Proneth B, Tyurina YY, Panzilius
E, Kobayashi S, Ingold I, Irmler M, Beckers J, Aichler M, Walch A,
et al: ACSL4 dictates ferroptosis sensitivity by shaping cellular
lipid composition. Nat Chem Biol. 13:91–98. 2017. View Article : Google Scholar :
|
|
104
|
Kagan VE, Mao G, Qu F, Angeli JP, Doll S,
Croix CS, Dar HH, Liu B, Tyurin VA, Ritov VB, et al: Oxidized
arachidonic and adrenic PEs navigate cells to ferroptosis. Nat Chem
Biol. 13:81–90. 2017. View Article : Google Scholar
|
|
105
|
Freitas FP, Alborzinia H, Dos Santos AF,
Nepachalovich P, Pedrera L, Zilka O, Inague A, Klein C, Aroua N,
Kaushal K, et al: 7-Dehydrocholesterol is an endogenous suppressor
of ferroptosis. Nature. 626:401–410. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Li Y, Ran Q, Duan Q, Jin J, Wang Y, Yu L,
Wang C, Zhu Z, Chen X, Weng L, et al: 7-Dehydrocholesterol dictates
ferroptosis sensitivity. Nature. 626:411–418. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Garcia-Martinez L, Zhang Y, Nakata Y, Chan
HL and Morey L: Epigenetic mechanisms in breast cancer therapy and
resistance. Nat Commun. 12:17862021. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Ling C and Rönn T: Epigenetics in human
obesity and type 2 diabetes. Cell Metab. 29:1028–1044. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Shu F, Xiao H, Li QN, Ren XS, Liu ZG, Hu
BW, Wang HS, Wang H and Jiang GM: Epigenetic and post-translational
modifications in autophagy: Biological functions and therapeutic
targets. Signal Transduct Target Ther. 8:322023. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Yang M, Luo H, Yi X, Wei X and Jiang DS:
The epigenetic regulatory mechanisms of ferroptosis and its
implications for biological processes and diseases. MedComm (2020).
4:e2672023. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Deng SH, Wu DM, Li L, Liu T, Zhang T, Li
J, Yu Y, He M, Zhao YY, Han R and Xu Y: miR-324-3p reverses
cisplatin resistance by inducing GPX4-mediated ferroptosis in lung
adenocarcinoma cell line A549. Biochem Biophys Res Commun.
549:54–60. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Song Z, Jia G, Ma P and Cang S: Exosomal
miR-4443 promotes cisplatin resistance in non-small cell lung
carcinoma by regulating FSP1 m6A modification-mediated ferroptosis.
Life Sci. 276:1193992021. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Chen Q, Pan Q, Gao H, Wang Y and Zhong X:
miR-17-5p/HOXA7 is a potential driver for brain metastasis of lung
adenocarcinoma related to ferroptosis revealed by bioinformatic
analysis. Front Neurol. 13:8789472022. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Lu X, Kang N, Ling X, Pan M, Du W and Gao
S: MiR-27a-3p promotes non-small cell lung cancer through
SLC7A11-mediated-ferroptosis. Front Oncol. 11:7593462021.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Bi G, Liang J, Zhao M, Zhang H, Jin X, Lu
T, Zheng Y, Bian Y, Chen Z, Huang Y, et al: miR-6077 promotes
cisplatin/pemetrexed resistance in lung adenocarcinoma via
CDKN1A/cell cycle arrest and KEAP1/ferroptosis pathways. Mol Ther
Nucleic Acids. 28:366–386. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Wei D, Ke YQ, Duan P, Zhou L, Wang CY and
Cao P: MicroRNA-302a-3p induces ferroptosis of non-small cell lung
cancer cells via targeting ferroportin. Free Radic Res. 55:821–830.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Zhang L, Xu Y, Cheng Z, Zhao J, Wang M,
Sun Y, Mi Z, Yuan Z and Wu Z: The EGR1/miR-139/NRF2 axis
orchestrates radiosensitivity of non-small-cell lung cancer via
ferroptosis. Cancer Lett. 595:2170002024. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Guo J, Gong C and Wang H: PURPL promotes
M2 macrophage polarization in lung cancer by regulating RBM4/xCT
signaling. Crit Rev Eukaryot Gene Expr. 34:59–68. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Yao F, Zhao Y, Wang G, Zhao M, Hong X, Ye
Z, Dong F, Li W and Deng Q: Exosomal lncRNA ROR1-AS1 from
cancer-associated fibroblasts inhibits ferroptosis of lung cancer
cells through the IGF2BP1/SLC7A11 signal axis. Cell Signal.
120:1112212024. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Lu CL, Liu J and Yang JF: LncRNA-XIST
promotes lung adenocarcinoma growth and inhibits ferroptosis by
regulating GPX4. Mol Biotechnol. 28: View Article : Google Scholar : 2023.
|
|
121
|
Zhang N, Huang J, Xu M and Wang Y: LncRNA
T-UCR Uc.339/miR-339/SLC7A11 axis regulates the metastasis of
ferroptosis-induced lung adenocarcinoma. J Cancer. 13:1945–1957.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Wu H and Liu A: Long non-coding RNA NEAT1
regulates ferroptosis sensitivity in non-small-cell lung cancer. J
Int Med Res. 49:3000605219961832021. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Zhang R, Pan T, Xiang Y, Zhang M, Xie H,
Liang Z, Chen B, Xu C, Wang J, Huan Q, et al: Curcumenol triggered
ferroptosis in lung cancer cells via lncRNA H19/miR-19b-3p/FTH1
axis. Bioact Mater. 13:23–36. 2022.PubMed/NCBI
|
|
124
|
Tai F, Zhai R, Ding K, Zhang Y, Yang H, Li
H, Wang Q, Cao Z, Ge C, Fu H, et al: Long non-coding RNA lung
cancer-associated transcript 1 regulates ferroptosis via
microRNA-34a-5p-mediated GTP cyclohydrolase 1 downregulation in
lung cancer cells. Int J Oncol. 64: View Article : Google Scholar : 2024.
|
|
125
|
An J, Shi J, Yang C, Luo J, Li Y, Ren J,
Lv Y and Zhang Y: Regulation of tumorigenesis and ferroptosis in
non-small cell lung cancer by a novel BBOX1-AS1/miR-326/PROM2 axis.
Mol Cell Biochem. 479:2143–2155. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Gao GB, Chen L, Pan JF, Lei T, Cai X, Hao
Z, Wang Q, Shan G and Li J: LncRNA RGMB-AS1 inhibits HMOX1
ubiquitination and NAA10 activation to induce ferroptosis in
non-small cell lung cancer. Cancer Lett. 590:2168262024. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Mao C, Wang X, Liu Y, Wang M, Yan B, Jiang
Y, Shi Y, Shen Y, Liu X, Lai W, et al: A G3BP1-interacting lncRNA
promotes ferroptosis and apoptosis in cancer via nuclear
sequestration of p53. Cancer Res. 78:3484–3496. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Gai C, Liu C, Wu X, Yu M, Zheng J, Zhang
W, Lv S and Li W: MT1DP loaded by folate-modified liposomes
sensitizes erastin-induced ferroptosis via regulating
miR-365a-3p/NRF2 axis in non-small cell lung cancer cells. Cell
Death Dis. 11:7512020. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Zhen S, Jia Y, Zhao Y, Wang J, Zheng B,
Liu T, Duan Y, Lv W, Wang J, Xu F, et al: NEAT1_1 confers gefitinib
resistance in lung adenocarcinoma through promoting AKR1C1-mediated
ferroptosis defence. Cell Death Discov. 10:1312024. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Chen H, Wang L, Liu J, Wan Z, Zhou L, Liao
H and Wan R: LncRNA ITGB2-AS1 promotes cisplatin resistance of
non-small cell lung cancer by inhibiting ferroptosis via activating
the FOSL2/NAMPT axis. Cancer Biol Ther. 24:22233772023. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Shi Z, Zhang H, Shen Y, Zhang S, Zhang X,
Xu Y and Sun D: SETD1A-mediated H3K4me3 methylation upregulates
lncRNA HOXC-AS3 and the binding of HOXC-AS3 to EP300 and increases
EP300 stability to suppress the ferroptosis of NSCLC cells. Thorac
Cancer. 14:2579–2590. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Sui X, Hu N, Zhang Z, Wang Y, Wang P and
Xiu G: ASMTL-AS1 impedes the malignant progression of lung
adenocarcinoma by regulating SAT1 to promote ferroptosis. Pathol
Int. 71:741–751. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Wang M, Mao C, Ouyang L, Liu Y, Lai W, Liu
N, Shi Y, Chen L, Xiao D, Yu F, et al: Long noncoding RNA LINC00336
inhibits ferroptosis in lung cancer by functioning as a competing
endogenous RNA. Cell Death Differ. 26:2329–2343. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Dai N, Ma H and Feng Y: Silencing of long
non-coding RNA SDCBP2-AS1/microRNA-656-3p/CRIM1 axis promotes
ferroptosis of lung cancer cells. Cell Mol Biol (Noisy-le-grand).
69:189–194. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Liu L, Su S, Ye D, Yu Z, Lu W and Li X:
Long non-coding RNA OGFRP1 regulates cell proliferation and
ferroptosis by miR-299-3p/SLC38A1 axis in lung cancer. Anticancer
Drugs. 33:826–839. 2022.PubMed/NCBI
|
|
136
|
Xu C, Jiang ZB, Shao L, Zhao ZM, Fan XX,
Sui X, Yu LL, Wang XR, Zhang RN, Wang WJ, et al: β-Elemene enhances
erlotinib sensitivity through induction of ferroptosis by
upregulating lncRNA H19 in EGFR-mutant non-small cell lung cancer.
Pharmacol Res. 191:1067392023. View Article : Google Scholar
|
|
137
|
Xu P, Wang L, Xie X, Hu F, Yang Q, Hu R,
Jiang L, Ding F, Mei J, Liu J and Xiao H: Hsa_circ_0001869 promotes
NSCLC progression via sponging miR-638 and enhancing FOSL2
expression. Aging (Albany NY). 12:23836–23848. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Jiang L, Kon N, Li T, Wang SJ, Su T,
Hibshoosh H, Baer R and Gu W: Ferroptosis as a p53-mediated
activity during tumour suppression. Nature. 520:57–62. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Zhao Y, Cui Q, Shen J, Shen W and Weng Y:
Hsa_ circ_0070440 promotes lung adenocarcinoma progression by
SLC7A11-mediated-ferroptosis. Histol Histopathol. 38:1429–1441.
2023.PubMed/NCBI
|
|
140
|
Pan CF, Wei K, Ma ZJ, He YZ, Huang JJ, Guo
ZZ, Chen ZP, Barr MP, Shackelford RE, Xia Y and Wang J: CircP4HB
regulates ferroptosis via SLC7A11-mediated glutathione synthesis in
lung adenocarcinoma. Transl Lung Cancer Res. 11:366–380. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Wu L, Li N, Zhu L and Shao G: CircPDSS1
(hsa_circ_0017998) silencing induces ferroptosis in non-small-cell
lung cancer cells by modulating the miR-137/SLC7A11/GPX4/GCLC axis.
Toxicol In Vitro. 99:1058872024. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Li Z, Fan M, Zhou Z and Sang X:
Circ_0082374 promotes the tumorigenesis and suppresses ferroptosis
in non-small cell lung cancer by up-regulating GPX4 through
sequestering miR-491-5p. Mol Biotechnol. 4: View Article : Google Scholar : 2024.
|
|
143
|
Fu H and Zhao Q: CircSCUBE3 promoted
ferroptosis to inhibit lung adenocarcinoma progression. Cell Mol
Biol (Noisy-le-grand). 70:161–168. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Shanshan W, Hongying M, Jingjing F, Yiming
Y, Yu R and Rui Y: CircDTL functions as an oncogene and regulates
both apoptosis and ferroptosis in non-small cell lung cancer cells.
Front Genet. 12:7435052021. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Liu B, Ma H, Liu X and Xing W: CircSCN8A
suppresses malignant progression and induces ferroptosis in
non-small cell lung cancer by regulating miR-1290/ACSL4 axis. Cell
Cycle. 22:758–776. 2023. View Article : Google Scholar :
|
|
146
|
Zhang X, Xu Y, Ma L, Yu K, Niu Y, Xu X,
Shi Y, Guo S, Xue X, Wang Y, et al: Essential roles of exosome and
circRNA_101093 on ferroptosis desensitization in lung
adenocarcinoma. Cancer Commun (Lond). 42:287–313. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Zhang Q, Fan X, Zhang X and Ju S:
Ferroptosis in tumors and its relationship to other programmed cell
death: Role of non-coding RNAs. J Transl Med. 21:5142023.
View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Wang Y, Chen Y, Zhang J, Yang Y, Fleishman
JS, Wang Y, Wang J, Chen J, Li Y and Wang H: Cuproptosis: A novel
therapeutic target for overcoming cancer drug resistance. Drug
Resist Updat. 72:1010182024. View Article : Google Scholar
|
|
149
|
Tang D and Kang R: NFE2L2 and ferroptosis
resistance in cancer therapy. Cancer Drug Resist.
7:412024.PubMed/NCBI
|
|
150
|
Winkle M, El-Daly SM, Fabbri M and Calin
GA: Noncoding RNA therapeutics-challenges and potential solutions.
Nat Rev Drug Discov. 20:629–651. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Nappi F: Non-coding RNA-targeted therapy:
A state-of-the-art review. Int J Mol Sci. 25:36302024. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
He AT, Liu J, Li F and Yang BB: Targeting
circular RNAs as a therapeutic approach: Current strategies and
challenges. Signal Transduct Target Ther. 6:1852021. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Zhao R, Fu J, Zhu L, Chen Y and Liu B:
Designing strategies of small-molecule compounds for modulating
non-coding RNAs in cancer therapy. J Hematol Oncol. 15:142022.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Han B, Liu Y, Zhang Q and Liang L:
Propofol decreases cisplatin resistance of non-small cell lung
cancer by inducing GPX4-mediated ferroptosis through the
miR-744-5p/miR-615-3p axis. J Proteomics. 274:1047772023.
View Article : Google Scholar
|
|
155
|
Fu R, You Y, Wang Y, Wang J, Lu Y, Gao R,
Pang M, Yang P and Wang H: Sanggenol L induces ferroptosis in
non-small cell lung cancer cells via regulating the
miR-26a-1-3p/MDM2/p53 signaling pathway. Biochem Pharmacol.
226:1163452024. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Huang J, Deng C, Guo T, Chen X, Chen P, Du
S and Lu M: Cinobufotalin induces ferroptosis to suppress lung
cancer cell growth by lncRNA LINC00597/hsa-miR-367-3p/TFRC pathway
via resibufogenin. Anticancer Agents Med Chem. 23:717–725. 2023.
View Article : Google Scholar
|