Introduction
Vaccination is an effective method of decreasing the
incidence of infectious diseases caused by pathogens to which
individuals are susceptible in early life (1). Unlike live and attenuated organism
vaccines, the inactive or dead organism vaccines and recombinant
protein or subunit vaccines are generally administered with an
adjuvant to aid in or enhance the immune response (2–3).
Adjuvants are compounds that enhance the ability of the
co-inoculated antigens to elicit an early, high and long-lasting
immune response with a reduced amount of antigen (4). Owing to the highly purified
recombinant proteins, which are increasingly used in the new
generation vaccines but are poorly immunogenic, formulations
comprising more than one adjuvant have been widely studied in
various vaccines. To enhance the potency of the first generation
adjuvants, including aluminum salts (also known as alums),
emulsions, polymeric particles and liposomes, additional components
have been added to them, generating the second generation of
vaccine adjuvants. Furthermore, the second generation adjuvants
have been licensed in approved vaccine products in the EU,
including Pandemrix (AS03; o/w emulsion + α-tocopherol) and Fendrix
or Cervarix (AS04; MPL + aluminum hydroxide) (5).
Bacterial DNA and oligodeoxynucleotides containing
unmethylated CpG motifs (CpG ODN) have been studied as vaccine
adjuvants for the induction of protective immunity against
bacterial, viral and parasitic infections in numerous animal models
(6–9). Previous studies have found that CpG
ODN is able to activate a T helper 1 (Th1)-like humoral immune
response with the production of IgG2a antibodies and a
cell-mediated immune response with the secretion of IFN-γ cytokines
(10–11). Furthermore, another study has shown
that plasmid DNA directly injected into the muscle cells of mice
elicited potent humoral and cellular immune responses (12). In addition, it has been shown that
plasmid DNA may be useful as an adjuvant for DNA vaccines (13,14).
Recently, our laboratory revealed that plasmid DNA (pcDNA 3.1)
encapsulated in cationic liposomes delivered an adjuvant effect in
the cancer immunotherapy of murine colon carcinoma (15). Other studies have demonstrated that
the adjuvant properties of plasmid DNA were dependent on its
content of CpG motifs which interacted with TLR9 and directly
induced the production of IL-12 and IFN-α by dendritic cells
(16–19). As the plasmid pcDNA 3.1 contains
the ampicillin resistance gene, it is commonly used for studies in
mice. Considering that the plasmid vector pVAX1, which contains a
kanamycin resistance gene, is permitted to be used in humans
(20), and the high cost of
synthesis of CpG ODN, we constructed a multiple CpG plasmid
(pv-16CpG) containing 16 CpG motifs (CpG 7909) to examine its
adjuvant effect in a hepatitis B surface antigen (HBsAg) vaccine
when combined with aluminum hydroxide (AL) or incomplete Freund’s
adjuvant (IFA).
Aluminium salts, particularly phosphate or hydroxide
salts, have been widely used in human and veterinary vaccines since
1930 with an excellent record of safety (21). Aluminum salts markedly induce the
generation of Th2 cytokines (IL-4 and IL-5) and antibodies of the
IgG1 isotype (22–23). IFA, a water-in-oil emulsion, has
been widely used in veterinary vaccine products and tested in
humans (24–25). The main mechanisms of action of
aluminum salts and IFA are similar, they prolong the duration of
antigen persistence at the injection site (5).
Since the introduction of a hepatitis B vaccine
containing alum over 30 years ago, a decline in the global
incidence of HBV infection has been observed (26). However, 5–10% of healthy
immunocompent individuals fail to produce a protective antibody
response to the HBV vaccine (27).
Improving the immunogenicity of the HBV vaccine is a strategy that
can be employed to overcome this problem. Therefore, new adjuvants
that elicit strong immune responses to HBsAg are required. Previous
studies have shown that certain adjuvant combinations induce a
stronger immune response than either individual component alone
(28). In this study, we used
formulations of pv-16CpG with AL or IFA as adjuvants for a
hepatitis B vaccine. The results showed that pv-16CpG suspended in
IFA triggered a stronger immune response against HBsAg in C57BL/6J
mice than pv-16CpG combined with AL. The plasmid pv-16CpG suspended
in IFA induced a Th1 immune response to HBsAg, characterized by the
generation of specific IgG2a antibodies and IFN-γ in antigen
stimulated T cells. We suggest that pv-16CpG, which may facilitate
the manufacturing process, may be used as an adjuvant for HBsAg
vaccines when mixed with IFA.
Materials and methods
Plasmid pv-16CpG preparation
The plasmid pv-16CpG was constructed as follows:
16CpG motif ODN (16 × 5′-TCGTCGTTTTGTCGTTTTGTCGTT-3′) fragments
were synthesized and inserted into the plasmid pVAX1 (Invitrogen
Life Technologies, San Diego, CA, USA) using the enzymes
HindIII (5′-end of the fragment) and BamHI (3′-end of
the fragment) from Invitrogen. The fragment sequences were
confirmed by DNA sequencing. The plasmid pv-16CpG was prepared
using the Endofree Plasmid Giga kit (Qiagen, Chatsworth, CA, USA).
There was no genomic DNA, small DNA fragments or RNA in the
pv-16CpG DNA (OD260/2280=1.8–2.0).
Reagents
AL gel adjuvant was purchased from Brenntag
Biosector (Frederikssund, Denmark) and IFA was purchased from Sigma
Poole (Dorset, UK). The hepatitis B surface antigen (subtype ad)
protein was purchased from American Research Products, Inc.
(Belmont, MA, USA).
Animals and immunizations
Six to eight-week-old female C57BL/6J mice (n=8 mice
per group) were purchased from the Experimental Animal Center of
Sichuan University and were housed in our animal research facility.
The mice were immunized intramuscularly (hind limbs) three times at
2-week intervals (in weeks 0, 2 and 4) with 0.1 μg HBsAg and
adjuvant in a total volume of 100 μl (Fig. 1). The adjuvant amounts used for
each immunization were: 25 μg Al3+, 50 μg pvAX, 50 μg
pv-16CpG or IFA 50 μl (v/v 1:1), mixed by syringe. The mice were
administered a booster immunization with the same dose in week 31.
Blood samples were collected from the animals in weeks 0, 1, 3, 5,
7, 9, 11, 13, 17, 21, 25, 30, 32, 34 and 50 following the
immunization. The serum was divided into aliquots and was
maintained at −70°C.
 | Figure 1Schema of the immunization protocol.
Mice (n=8) were immunized with HBsAg (0.1 μg) and AL (25 μg
Al3+), IFA (50 μl; v/v=1:1), AL + pvAX (50 μg), IFA +
pvAX (50 μg), AL + pv-16CpG (50 μg) or IFA + pv-16CpG (50 μg) in
weeks 0, 2 and 4. In addition, mice were immunized with the same
dose of vaccine in week 31. Following the third immunization, mice
(n=3) were sacrificed and an ELISPOT assay was performed. Sera were
collected in weeks 1, 3, 5, 7, 9, 13, 17, 21, 25, 30, 32, 34 and
50, and the anti-HBsAg titers were measured by endpoint dilution
ELISA. HBsAg, hepatitis B surface antigen; AL, aluminum hydroxide;
IFA, incomplete Freund’s adjuvant; pvAX, pv-16CpG, plasmid vectors;
ELISA, enzyme-linked immunosorbent assay. |
Enzyme-linked immunosorbent assay
(ELISA)
To measure the HBsAg-specific antibody response, a
96-well plate was coated overnight at 4°C with 100 μl HBsAg
solution (1 μg/ml) in sodium carbonate buffer. The plate was washed
three times with 0.1% Tween in PBS (PBST), and then blocked with 5%
skimmed milk in PBST for 1 h at 37°C. After being washed 5 times
with PBST, the plate was incubated with serially diluted mice sera
in 5% skimmed milk for 1 h at 37°C, followed by further washes.
Bound antibodies were detected using horseradish
peroxidase-conjugated goat anti-mouse IgG, IgG1, IgG2a, IgG2b or
IgG3 (Southern Biotechnologies, Birmingham, AL, USA) diluted at
1:400 for 1 h at 37°C. The plate was washed again and incubated
with 100 μl substrate-chromogen solution (SureBlue™ TMB Microwell
Peroxidase substrate, KPL, Inc., Gaithersburg, MD, USA) for 20 min
at room temperature. A volume of 0.5 M H2SO4
(100 μl) was added to terminate the reaction and the absorbance was
read at 450 nm using a Bio-Rad (Hercules, CA, USA) microtiter plate
reader. ELISA titers were expressed as the reciprocal of the
highest dilution resulting in a value of two standard deviations
above that of non-immunized serum with a cut-off of 0.2.
IFN-γ and IL-4 ELISPOT assays
To measure the numbers of IFN-γ and IL-4 secreting
cells, the dual-color ELISPOT Mouse IFN-γ/IL-4 kit (R&D
systems, Inc., Minneapolis, MN, USA) was used. Following the third
immunization, the mice were sacrificed and their splenocytes were
separated as described in the manual of the EZ-sep™ mouse 1X.
(Dakewe Biotech Co., Ltd., Shenzhen, China). The splenocytes were
added to triplicate wells at a concentration of 5×105
cells/well and cultured in the presence of HBsAg (5 μg/ml) or ConA
(1 μg/ml) for 48 h. Stained spots were counted using a
computer-assisted ELISPOT image analyzer (Immunospot, Cellular
Technology Ltd., Cleveland, OH, USA).
Electrophoretic mobility shift assay
(EMSA) and scanning electron microscopy (SEM) experiments
Binding assays were performed by incubating pv-16CpG
(10 μg) with various amounts of AL in a final volume of 100 μl of
10 mM Tris-HCl and 270 mM sorbitol (pH 7.4). After incubating at
37°C for 10 min, the complexes were centrifuged at 3000 rpm for 5
min. The deposits were subjected to electrophoresis through a 0.8%
agarose gel at 120 V for 15 min. The DNA was visualized by ethidium
bromide staining. The SEM experiments were performed using a
JSM-5900LV SEM instrument to inspect the sizes of the particles.
The samples were prepared according to the prescription of
administration for mice and were then gold-coated and observed
under an accelerating voltage of 20 kV.
Statistical analysis
The studies were performed on groups of 3–5
individual mice. Statistical comparisons were carried out using
one-way analysis of variance (ANOVA) followed by Tukey’s test.
Results were considered statistically significant at p<0.05.
Results
Seroconversion induced by IFA +
pv-16CpG
In week 1 following the immunization, 20% of mice in
the AL + pvAX, IFA + pvAX and IFA + pv-16CpG groups had produced
anti-HBs (titer ≥100) (Fig. 2). In
week 3 post-immunization, the anti-HBs seroconversion rate of the
AL and AL + pvAX groups had also reached 20%. In addition, at this
time, the rates in the IFA + pvAX and IFA + pv-16CpG groups were 40
and 80%, respectively. However, no seroconversion was identified in
the AL + pv-16CpG and IFA groups. The seroconversion rate in the
IFA + pv-16CpG group was clearly improved compared with the other 5
groups. However, 5 weeks after immunization, the seroconversion
rates in the AL + pv-16CpG and IFA groups were 40 and 20%,
respectively. Moreover, while the seroconversion rate of the AL +
pvAX group reached 60%, the rates in the other 3 groups were
100%.
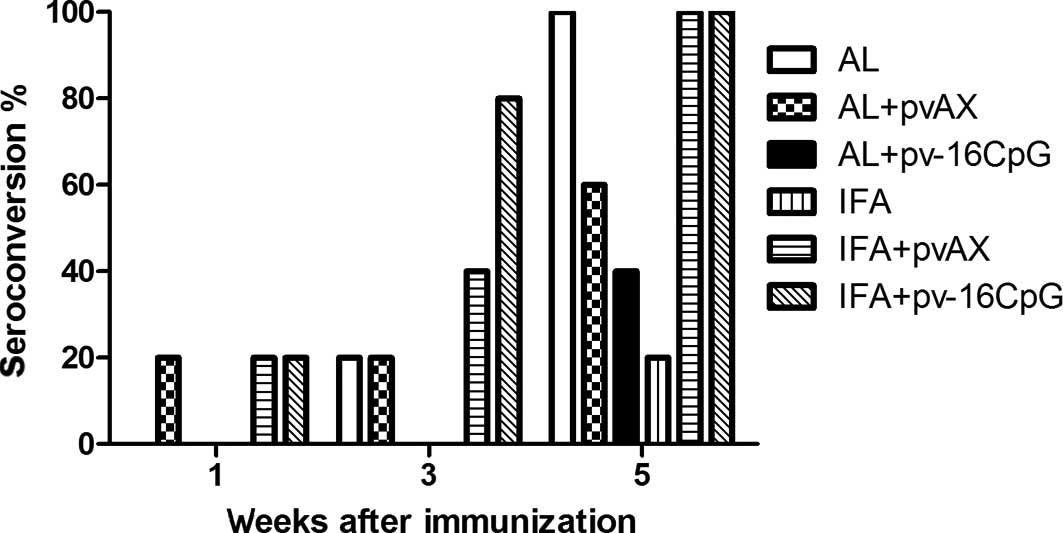 | Figure 2Percentage of seroconversion for
C57BL/6J mice immunized in weeks 0, 2 and 4 using HBsAg with
various adjuvants. C57BL/6J mice (n=5) were immunized 3 times by
intramuscular (i.m.) injection with 0.1 μg HBsAg formulated with 25
μg AL, IFA (50 μl; v/v=1:1), AL + pvAX (50 μg), IFA + pvAX (50 μg),
25 μg AL + 50 μg pv-16CpG, or 50 μg IFA + pv-16CpG (50 μl). Sera
obtained in weeks 1, 3 and 5 were assayed for anti-HBsAg by
endpoint-dilution ELISA assay. Seroconversion was defined as a
dilution titer ≥100 with a cut-off value of 0.2. HBsAg, hepatitis B
surface antigen; AL, aluminum hydroxide; IFA, incomplete Freund’s
adjuvant; pvAX, pv-16CpG, plasmid vectors; ELISA, enzyme-linked
immunosorbent assay. |
Enhancement of anti-HBs titer by IFA +
pv-16CpG
The kinetics of the HBsAg-specific humoral immune
responses were monitored over the 50 weeks following immunization.
C57BL/6J mice were immunized by intramuscular (i.m.) injection of
0.1 μg HBsAg with various adjuvants at different time points
(Fig. 2). When the double adjuvant
combinations were used, the anti-HBs titer of the IFA + pv-16CpG
group in week 13 (first peak) was 50-fold higher than that of the
IFA group and 320-fold higher than that of the AL group (Fig. 3). However, there was no increase in
the anti-HBs titers when the combination of AL and pv-16CpG was
used as the adjuvant. The IFA + pvAX group underwent a significant
increase compared with the remaining groups, with the exception of
the IFA + pv-16CpG group. Throughout the study, the titers of the
IFA + pv-16CpG group were 5–8-fold higher than that of the IFA +
pvAX group at 50 weeks. The antibody titer of the IFA group
underwent a sustained growth following the first immunization, and
this increasing trend stopped in week 17, while in the other
groups, it stopped in week 13. Twenty-one weeks after immunization,
the titers of the 6 groups exhibited a sustained decrease. To
detect the memory response evoked by the vaccine, the mice were
boosted with the same dose of vaccine in week 31. A 2- to 20-fold
increase in anti-HBs titers was observed in all the groups.
However, the anti-HBs titer of the IFA + pv-16CpG group in week 32
was 40- or 300-fold higher than that in the AL and IFA groups,
respectively. Furthermore, the anti-HBs antibody titer in the IFA +
pv-16CpG group in week 32 was 800-fold higher than that in the AL +
pv-16CpG group (Fig. 3).
 | Figure 3IgG antibody titer kinetics against
HBsAg in C57BL/6J mice immunized with AL, IFA, AL + pvAX, IFA +
pvAX, AL + pv-16CpG or IFA + pv-16CpG. Mice were immunized as
described above. ELISA titers are defined as the reciprocal of the
highest dilution resulting in a reading of two standard deviations
above that of non-immunized mice with a cut-off value of 0.2. Sera
obtained from each mouse were used to detect the titer by ELISA in
each group. Data are presented as the group mean for total IgG
titer to HBsAg. Vertical lines are the SEM. Two other experiments
produced similar results; ★p<0.05;
★★p<0.001. HBsAg, hepatitis B surface antigen; AL,
aluminum hydroxide; IFA, incomplete Freund’s adjuvant; pvAX,
pv-16CpG, plasmid vectors; ELISA, enzyme-linked immunosorbent
assay. |
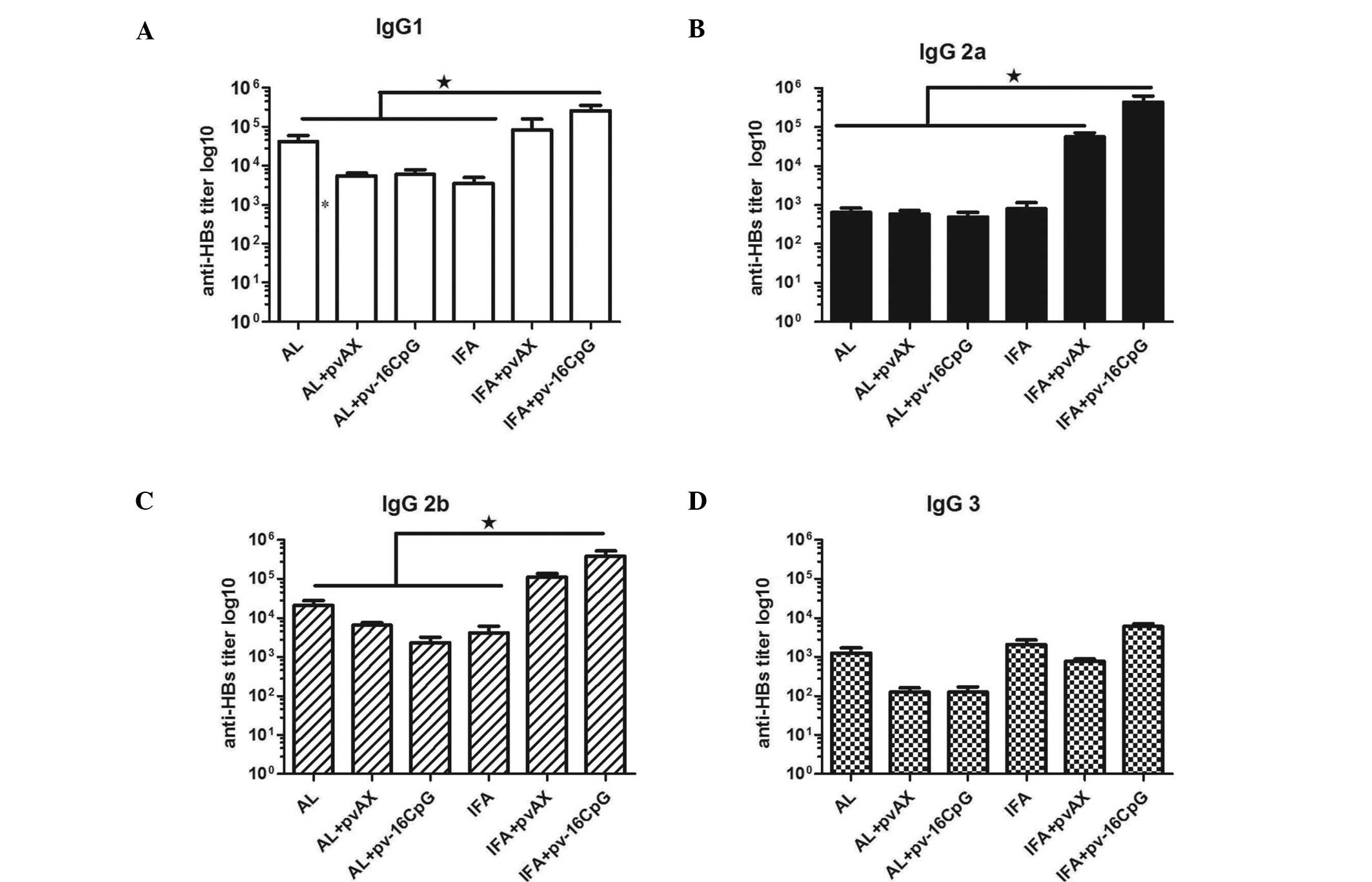
Antibody isotype profiles evoked by IFA +
pv-16CpG
The ratio of IgG2a to IgG1 antibody levels is often
used to evaluate the type of Th response, with a value >1
indicating a predominantly Th1 response and <1 indicating a
predominantly Th2 response. Immunization with AL or IFA alone
generated a Th2 response, with higher levels of anti-HBs antibodies
of the IgG1 isotype than those of the IgG2a isotype (Fig. 4A and B). The values of IgG2a/IgG1
were 0.021 and 0.25 in the AL and IFA groups, respectively, 7 weeks
after immunization. When the plasmid pv-16CpG combined with AL was
added as the adjuvant, the anti-HBs titers of the IgG1 and IgG2a
isotypes were lower than those in the AL group. In addition, the
IgG2a/IgG1 ratio was 0.063, 7 weeks after immunization. When the
plasmid pvAX was combined with AL or IFA, the values of IgG2a/IgG1
in week 7 were 0.078 and 0.5, respectively. This finding suggested
that the combination of plasmid and AL or IFA induced a Th2 immune
response. However, with an increased effect on IgG1 and IgG 2a
antibody titers, the combination of plasmid pv-16CpG and IFA evoked
a Th1 response (Fig. 4A and B)
with a value of IgG2a/IgG1 >1 (2.0). Moreover, the IgG2b and
IgG3 antibody titers in the IFA + pv-16CpG group increased
significantly compared with those in the IFA group, but decreased
in the AL + pv-16CpG group when compared with the AL group
(Fig. 4C and D). These findings
suggest that the plasmid pv-16CpG has great potential for improving
the immune response when combined with IFA but not with AL.
Assay of HBsAg-specific T cell immune
response by ELISPOT
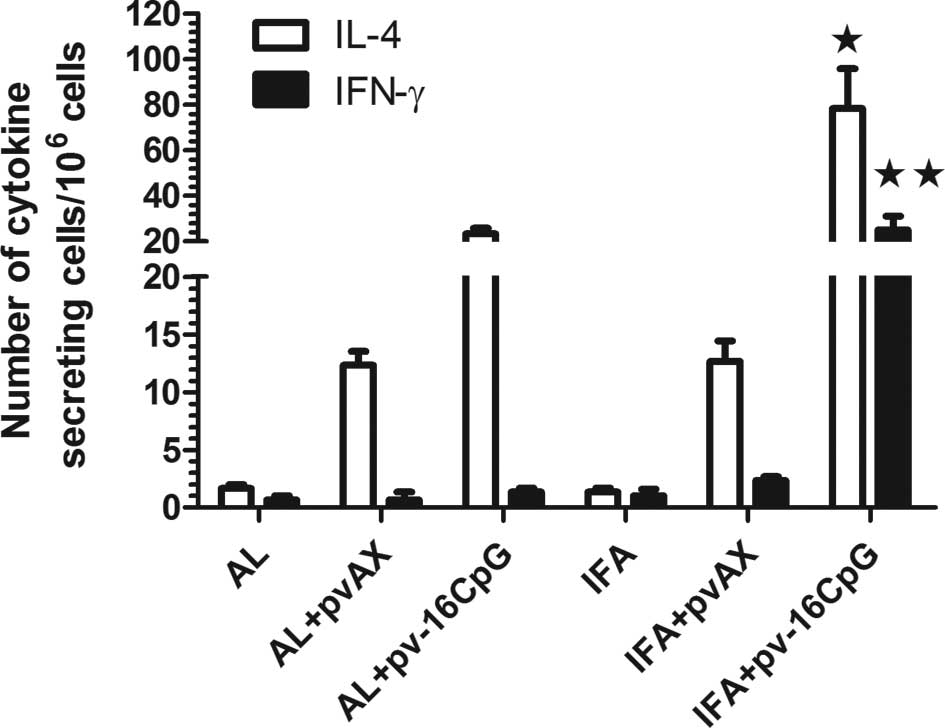
To characterize the immune responses induced by the
various formulations, the production of IL-4 and IFN-γ by
HBsAg-exposed splenocytes from immunized mice were examined by
re-stimulating the splenocytes with HBsAg. The number of
IL-4-secreting T cells in the mice immunized with AL + pv-16CpG or
IFA + pv-16CpG were significantly higher than that in the AL, AL +
pvAX, IFA or IFA + pvAX groups (Fig.
5). The number of IL-4-secreting splenocytes in the mice
immunized with AL + pv-16CpG or IFA + pv-16CpG were approximately
15- and 30-fold higher than in the mice immunized with AL or IFA
alone, respectively (Fig. 5). When
the plasmid pvAX was added to AL or IFA, the number of
IL-4-secreting splenocytes was also lower than that in the IFA +
pv-16CpG group (p<0.05). Furthermore, the highest number of
IFN-γ-secreting T cells was found in the mice receiving IFA +
pv-16CpG. The number of IFN-γ-secreting T cells in the mice
receiving IFA + pv-16CpG was almost 30-fold higher than that in the
mice from the other groups (p<0.001). Moreover, the higher IFN-γ
secretion indicated a potent type 1 cell-mediated immune
response.
 | Figure 5Frequency of HBsAg-specific IFN-γ- and
IL-4-secreting cells in splenocytes of mice 5 weeks after
immunization. C57BL/6J mice (n=3) were immunized with 0.1 μg HBsAg
combined with AL, IFA, AL + pv-16CpG, IFA + pv-16CpG, AL + pvAX or
IFA + pvAX in weeks 0, 2 and 4. The number of cytokine-secreting
cells were determined by a computer-assisted ELISPOT image
analyzer. Bars show the mean ± SEM number of cytokine-specific
cells in splenocytes as determined by the ELISPOT assay.
★p<0.05; ★★p<0.001. HBsAg, hepatitis B
surface antigen; AL, aluminum hydroxide; IFA, incomplete Freund’s
adjuvant; pv-16CpG, pvAX, plasmid DNA. |
Discussion
Vaccines are widely used in the protection and
eradication of a wide range of diseases, including smallpox, polio,
measles, whooping cough and hepatitis. Sub-unit vaccines, including
HBsAg-based vaccines, have a good record of safety but low efficacy
in eliciting an immune response (29). Therefore, adjuvants which enhance
the immunogenicity of these vaccines are urgently required.
Aluminum salts (particularly aluminum hydroxide or
phosphate), which have been licensed as adjuvants for human use,
have been widely and successfully used in a great number of
licensed vaccines although they induce a poor cytotoxic T cell and
Th1 immune response (30–31). As shown in Figs. 4 and 5, AL induces a Th2 immune response, with
predominantly IgG1 antibody and IL-4 secretion from immunized
splenocytes. When the plasmid pv-16CpG was combined with AL, it
induced a weaker immune response than AL alone.
IFA, a water-in-oil emulsion adjuvant, which
produces a high and sustained antibody response, has been used in
experimental vaccines in mice, rabbits, guinea pigs and rhesus
macaques (24). Furthermore, IFA
induces a Th2 type response, which is not effective for a cellular
immune response and limits the adjuvant activity of IFA. Our
results showed that when IFA was used alone, it induced
predominantly IgG1 antibody (Fig. 4A
and B) and IL-4 production, as detected by the IL-4/IFN-γ
ELISPOT assay (Fig. 5). However,
mixing or using IFA in conjunction with other adjuvant-active
compounds has been successful in animal testing and preclinical
trials (24). Furthermore,
findings of another study have shown that insect DNA and CpG
suspended in IFA was a more powerful adjuvant in mice than CFA,
which is considered as the ‘gold standard’ for adjuvant function
(32). In our study, the use of a
combination of IFA and pv-16CpG as the adjuvant for a HBsAg
vaccine, resulted in higher anti-HBs antibody titers than the other
adjuvants tested (at least 10-fold higher than the other groups
throughout the experiment; Fig. 3)
and significantly increased IFN-γ production, even compared with
the AL + pv-16CpG group (p<0.001). The booster immunization in
week 31 resulted in a profoundly increased and sustained memory
response in the IFA + pv-16CpG group. The data suggest that
pv-16CpG suspended in IFA evokes cellular and humoral immune
responses.
The data demonstrated that pv-16CpG exhibited a
strong adjuvant activity when combined with IFA but not with AL.
The reasons which may explain this result include the fact that the
mechanism of action of adjuvant aluminum salts involves the
formation of an antigen depot at the injection site (33), the electrostatic attractive forces
between the aluminum salt (positive charge) and antigen (negative
charge) (34). As pv-16CpG itself
may be considered as a polyelectrolyte with a high negative charge
(35), there is likely to be an
electrostatic interaction between AL and pv-16CpG. As shown in
Fig. 6, pv-16CpG was completely
adsorbed by AL when the ratio was 1:10 by weight. This interaction
may reduce the ability of the aluminum salt to form an antigen
depot from which the antigen is slowly released. However, there may
be not an electrostatic interaction between pv-16CpG and IFA.
Moreover, in the microdroplets of oil formed by mixing, the
electrostatic attractive force between pv-16CpG and the HBsAg
protein may contribute to the slow release of antigen. Due to the
limitations of the technology, EMSA could not be performed for IFA
and plasmid DNA.
 | Figure 6Electrophoretic mobility shift assay
(EMSA) and scanning electron microscopy (SEM) of AL and pvAX or
pv-16CpG complexes. (A) EMSA of AL and pv-16CpG complex: M, marker;
lane 1, pv-16CpG (10 μg); lane 2, pv-16CpG (10 μg) + AL (5 μg);
lane 3, pv-16CpG (10 μg) + AL (10 μg); lane 4, pv-16CpG (10 μg) +
AL (20 μg); lane 5, pv-16CpG (10 μg) + AL (40 μg); lane 6, pv-16CpG
(10 μg) + AL (80 μg). (B) SEM micrograph of AL (25 μg) diluted and
vortexed in a total volume of 100 μl. (C) SEM micrograph of AL and
pvAX complex: AL (25 μg) and pvAX (50 μg) diluted and vortexed in a
total volume of 100 μl. (D) SEM micrograph of AL and pv-16CpG
complex: AL (25 μg) and pv-16CpG (50 μg) diluted and vortexed in a
total volume of 100 μl. Each graph is representative of at least
three experiments. AL, aluminum hydroxide; pvAX, pv-16CpG, plasmid
vectors. |
Another explanation for our results is that the size
of the antigen or the antigen formulated with an adjuvant is a
significant factor for its efficient uptake by antigen-presenting
cells (APCs). The range of particle sizes which allows the most
efficient uptake by APCs is 10–3,000 nm. In addition, the average
particle sizes of emulsions (such as IFA) and mineral salts (such
as AL) are 200–8,000 and 800–12,000 nm, respectively (36). The particle size of AL is smaller
than 1 μm, but becomes larger than 1.5 μm when pv-16CpG is combined
with it (Fig. 6B and C), and this
change may affect the take up of the antigen by APCs. The mechanism
of adjuvant activity in the vaccine comprising pv-16CpG suspended
in IFA requires further study.
In conclusion, the results of our studies have shown
that pv-16CpG may be used as an adjuvant for HBsAg vaccines when
combined with IFA but not with AL, and evoked cellular and humoral
immune responses against HBsAg.
Acknowledgements
This study is supported by the Program for New
Century Excellent Talents in University (NCET) (NCET-09-0575) and
the National Natural Science Foundation of China (grant no.
31070815).
References
|
1
|
Weeratna RD, McCluskie MJ, Xu Y and Davis
HL: CpG DNA induces stronger immune responses with less toxicity
than other adjuvants. Vaccine. 18:1755–1762. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ulmer JB, Valley U and Rappuoli R: Vaccine
manufacturing: challenges and solutions. Nat Biotechnol.
24:1377–1383. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aguilar JC and Rodriguez EG: Vaccine
adjuvants revisited. Vaccine. 25:3752–3762. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gupta RK and Siber GR: Adjuvants for human
vaccines-current status, problems and future prospects. Vaccine.
13:1263–1276. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
O’Hagan DT and De Gregorio E: The path to
a successful vaccine adjuvant - ‘the long and winding road’. Drug
Discov Today. 14:541–551. 2009.
|
|
6
|
Stacey KJ and Blackwell JM:
Immunostimulatory DNA as an adjuvant in vaccination against
Leishmania major. Infect Immun. 67:3719–3726. 1999.PubMed/NCBI
|
|
7
|
Jones TR, Obaldia N III, Gramzinski RA, et
al: Synthetic oligodeoxynucleotides containing CpG motifs enhance
immunogenicity of a peptide malaria vaccine in Aotus monkeys.
Vaccine. 17:3065–3071. 1999. View Article : Google Scholar
|
|
8
|
Elkins KL, Rhinehart-Jones TR, Stibitz S,
Conover JS and Klinman DM: Bacterial DNA containing CpG motifs
stimulates lymphocyte-dependent protection of mice against lethal
infection with intracellular bacteria. J Immunol. 162:2291–2298.
1999.PubMed/NCBI
|
|
9
|
Moldoveanu Z, Love-Homan L, Huang WQ and
Krieg AM: CpG DNA, a novel immune enhancer for systemic and mucosal
immunization with influenza virus. Vaccine. 16:1216–1224. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Klinman DM, Yi AK, Beaucage SL, Conover J
and Krieg AM: CpG motifs present in bacteria DNA rapidly induce
lymphocytes to secrete interleukin 6, interleukin 12, and
interferon gamma. Proc Natl Acad Sci USA. 93:2879–2883. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brazolot Millan CL, Weeratna R, Krieg AM,
Siegrist CA and Davis HL: CpG DNA can induce strong Th1 humoral and
cell-mediated immune responses against hepatitis B surface antigen
in young mice. Proc Natl Acad Sci USA. 95:15553–15558.
1998.PubMed/NCBI
|
|
12
|
Corr M, Lee DJ, Carson DA and Tighe H:
Gene vaccination with naked plasmid DNA: mechanism of CTL priming.
J Exp Med. 184:1555–1560. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kojima Y, Xin KQ, Ooki T, et al: Adjuvant
effect of multi-CpG motifs on an HIV-1 DNA vaccine. Vaccine.
20:2857–2865. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Klinman DM, Barnhart KM and Conover J: CpG
motifs as immune adjuvants. Vaccine. 17:19–25. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang XP, Yang L, Shi HS, et al: An N-,
C-terminally truncated basic fibroblast growth factor and LPD
(liposome-polycation-DNA) complexes elicits a protective immune
response against murine colon carcinoma. Cancer Biol Ther.
10:276–281. 2010. View Article : Google Scholar
|
|
16
|
Wagner H: Bacterial CpG DNA activates
immune cells to signal infectious danger. Adv Immunol. 73:329–368.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Krieg AM: CpG motifs in bacterial DNA and
their immune effects. Annu Rev Immunol. 20:709–760. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tudor D, Dubuquoy C, Gaboriau V, Lefèvre
F, Charley B and Riffault S: TLR9 pathway is involved in adjuvant
effects of plasmid DNA-based vaccines. Vaccine. 23:1258–1264. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Spies B, Hochrein H, Vabulas M, et al:
Vaccination with plasmid DNA activates dendritic cells via
Toll-like receptor 9 (TLR9) but functions in TLR9-deficient mice. J
Immunol. 171:5908–5912. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gurunathan S, Klinman DM and Seder RA: DNA
vaccines: immunology, application, and optimization. Annu Rev
Immunol. 18:927–974. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cox JC and Coulter AR: Adjuvants-a
classification and review of their modes of action. Vaccine.
15:248–256. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Weeratna R, Comanita L and Davis HL: CPG
ODN allows lower dose of antigen against hepatitis B surface
antigen in BALB/c mice. Immunol Cell Biol. 81:59–62. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mancino D and Ovary Z: Adjuvant effects of
amorphous silica and of aluminium hydroxide on IgE and IgG1
antibody production in different inbred mouse strains. Int Arch
Allergy Appl Immunol. 61:253–258. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jensen FC, Savary JR, Diveley JP and Chang
JC: Adjuvant activity of incomplete Freund’s adjuvant. Adv Drug
Deliv Rev. 32:173–186. 1998.
|
|
25
|
Billiau A and Matthys P: Modes of action
of Freund’s adjuvants in experimental models of autoimmune
diseases. J Leukoc Biol. 70:849–860. 2001.
|
|
26
|
Liaw YF and Chu CM: Hepatitis B virus
infection. Lancet. 373:582–592. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zuckerman JN: Protective efficacy,
immunotherapeutic potential, and safety of hepatitis B vaccines. J
Med Virol. 78:169–177. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kovacs-Nolan J, Latimer L, Landi A, et al:
The novel adjuvant combination of CpG ODN, indolicidin and
polyphosphazene induces potent antibody- and cell-mediated immune
responses in mice. Vaccine. 27:2055–2064. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Perrie Y, Mohammed AR, Kirby DJ, McNeil SE
and Bramwell VW: Vaccine adjuvant systems: enhancing the efficacy
of sub-unit protein antigens. Int J Pharm. 364:272–280. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Harandi AM, Medaglini D and Shattock RJ;
Working Group convened by EUROPRIS. Vaccine adjuvants: a priority
for vaccine research. Vaccine. 28:2363–2366. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lindblad EB: Aluminium compounds for use
in vaccines. Immunol Cell Biol. 82:497–505. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun S, Kishimoto H and Sprent J: DNA as an
adjuvant: capacity of insect DNA and synthetic
oligodeoxynucleotides to augment T cell responses to specific
antigen. J Exp Med. 187:1145–1150. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shirodkar S, Hutchinson RL, Perry DL,
White JL and Hem SL: Aluminum compounds used as adjuvants in
vaccines. Pharm Res. 7:1282–1288. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
al-Shakhshir RH, Regnier FE, White JL and
Hem SL: Contribution of electrostatic and hydrophobic interactions
to the adsorption of proteins by aluminium-containing adjuvants.
Vaccine. 13:41–44. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gürsel M, Tunca S, Ozkan M, Ozcengiz G and
Alaeddinoglu G: Immunoadjuvant action of plasmid DNA in liposomes.
Vaccine. 17:1376–1383. 1999.PubMed/NCBI
|
|
36
|
Bachmann MF and Jennings GT: Vaccine
delivery: a matter of size, geometry, kinetics and molecular
patterns. Nat Rev Immunol. 10:787–796. 2010. View Article : Google Scholar : PubMed/NCBI
|