Introduction
Considering the rapid development of comprehensive
treatment for lung cancer, it is regrettable that no breakthrough
has yet been made concerning the long-term efficacy of treatment
and prognosis (1,2). Owing to the development of molecular
biology, gene therapy, following other traditional treatments such
as surgery, radiotherapy and chemotherapy, has become an attractive
therapeutic model. RNA interference, an evolution-conserving cell
defense mechanism mediated by small interferencing RNA (siRNA), can
induce post-transcriptional gene silencing. The advantages of this
method include stability, specificity, efficacy and low
cytotoxicity. Thus it is extensively used on functional genome and
gene therapy (3,4). Survivin, a new member of the
inhibitors of apoptosis (IAP) family, was expressed highly in most
tumor tissue; however, it is not expressed in terminal
undifferentiated mature tissue. Survivin is closely related to the
generation and development of tumors (5,6). We
successfully constructed a siRNA lentiviral vector targeting the
survivin gene and then transfected it to A549 cells. We also
explored the effect of survivin on the proliferation and apoptosis
of human lung cancer cells and investigated the mechanisms
underlying this process.
Materials and methods
Materials
The human lung cancer lines, A549 and 293T, were
purchased from the Shanghai Cell Resource Center of the Chinese
Academy of Sciences. The pGC-LV vector, pHelper 1.0 vector and
pHelper 2.0 vector were purchased from Shanghai GeneChem Co., Ltd.
The Qiagen Plasmid Midi Kit was purchased from Qiagen (Valencia,
CA, USA). Trypsase was purchased from Shanghai Chemical Reagent
Co., Ltd. E.coli DH5α, SYBR Master Mixture, T4 DNA ligase
and TaqDNA polymerase were purchased from Takara, (Shiga, Japan).
Age I restriction enzyme and EcoRI restriction enzyme were
purchased from New England Biolabs Co. Liposome Lipfectamine 2000
was purchased from Invitrogen (Carlsbad, CA, USA). DMSO was
purchased from Shanghai Biological Reagent Co., Ltd. DMEM culture
medium was purchased from Gibco (Carlsbad, CA, USA). FBS was
purchased from Shanghai Weike Biochemical Reagent Co., Ltd.
Oligo(dT) was purchased from Sangon Biotech Co., Ltd (Shanghai,
China). M-MLV reverse transcriptase and dNTP were purchased from
Promega (Madison, WI, USA). A flow cytometer and its software were
purchased from the USA. An inverted microscope was purchased from
Germany. A flow cytometer and its software were purchased from
Beckman-coulter (Fullerton, CA, USA). An inverted microscope was
purchased from Leica (Bensheim, Germany).
Methods
Cell culture
Human lung cancer A549 cells were cultured in DMEM
culture medium containing 10% FBS. The temperature of the
thermostat was 37˚C and CO2 concentration was 5%. The
cells were digested and passaged by 0.25% lipase every 2–3 days.
The cells at log phase were selected for the next step.
Design and screening of siRNA against
survivin
We designed the target sequence according to
survivin mRNA sequence in GenBank and the principles of siRNA
design (7,8). Three pairs of siRNA targeting
survivin and 1 pair of siRNA with the negative control were
designed (Table I). The synthesis
of siRNA was carried out by Shanghai GeneChem Co., Ltd. The siRNA
was then transfected into 293T cells, according to the guidelines
of Lipfectamine 2000 (Invitrogen). The transfecting results were
observed under a fluorescence microscope after 24 h and the
cell-collection and protein-abstraction were carried out 36 h
later. The most efficient siRNA was chosen by western blot
analysis.
 | Table IsiRNA sequence specific to
survivin. |
Table I
siRNA sequence specific to
survivin.
| Target | Source | Locus | Sequence |
|---|
| Survivin-1 | Human | NM_001168 |
GGCTGGCTTCATCCACTGC |
| Survivin-2 | Human | NM_001168 |
GGACCACCGCATCTCTACA |
| Survivin-3 | Human | NM_001168 |
GAAAGTGCGCCGTGCCATC |
| Negative
control | - | - |
TTCTCCGAACGTGTCACGT |
Construction and transfection of
survivin-siRNA lentiviral vector
Western blot analysis proved that the first pair of
siRNA was the most efficient. A double-stranded DNA fragment, with
cohesive termini of Age I restriction enzyme and EcoRI
restriction enzyme, and an internal hairpin sequence of
5′-GGCTGGCTTCATCCACTGCTTCAAGAGAGCAGTGG ATGAAGCCAGCC-3′, was
synthesized in vitro. The fragment was ligated into pGC-LV
and then transfected into E.coli DH5α. Following
amplification and screening, the construction was confirmed by
sequencing. The plasmid was extracted and survivin-siRNA lentiviral
vector was recombined, transfecting the A549 cells into a knockdown
group (KD). The A549 cells transfected with the negative control
and no sequence were labelled negative control (NC) and control
group (CON), respectively.
Isolation of total RNA and RT-qPCR
Total RNA was extracted by TRIzol and then
reverse-transcribed into cDNA, for which real-time quantitative PCR
(RT-qPCR) was then performed. The survivin and actin primers (as
the internal control) were synthesized by Shanghai GeneChem Co.,
Ltd. The sequences are shown in Table
II. The reaction conditions of PCR were: pre-denaturation was
at 95˚C for 15 sec; denaturation was at 95˚C for 5 sec; annealing
was at 60˚C for 30 sec; 45 cycles were completed. The mixture was
denatured for 1 min at the end of PCR and then cooled to 55˚C, at
which the double strands of DNA could combine sufficiently. From 55
to 95˚C the light absorption value was recorded for 4 sec at every
0.5˚C. From this step the melting curve was depicted. The
quantitative analysis was performed with the ratio of the target
gene to actin. The 2−Δ ΔCt method was used for
statistical analysis.
| Table IIPrimer sequences of survivin and
actin. |
Table II
Primer sequences of survivin and
actin.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) | Product size
(bp) |
|---|
| Survivin |
ACCGCATCTCTACATTCAAG |
CAAGTCTGGCTCGTTCTC | 113 |
| Actin |
GTGGACATCCGCAAAGAC |
AAAGGGTGTAACGCAACTA | 302 |
Detection of protein expression by
western blotting
Total protein of A549 cells was isolated 72 h after
transfection. Protein quantification was performed by BCA. The
protein sample was normalized at the same time. The sample load was
30 μg total protein per lane. Protein from 10% SDS-PAGE gel was
transferred to a PVDF membrane following electrophoresis. The
protein was blocked with 5% non-fat dry milk at 4˚C. The primary
antibodies, survivin (1:1000) and GAPDH (1:1000), were then added,
and the mixture was subsequently incubated overnight at 4˚C on a
rocking platform. After washing the membrane, HRP-conjugated
secondary antibody (1:5000) was added to it and it was then
incubated for 2 h. Protein bands were detected (the colored
membranes) with the enhanced chemiluminescence (ECL) system and
exposed to X-ray film. The membranes with no color (gray) were
scanned using the image analytical system.
Cell proliferation by MTT assay
At the log phase of each group, A549 cells were
inoculated into 96-well plates at 100 μl per well. The inoculating
density was 1×104/well. The plates were incubated at
37˚C, 5% CO2 and saturated humidity. MTT assay was
performed on days 1 to 5 following incubation. A value at a
wavelength of 570 nm was detected by a microplate
spectrophotometer. The mean value of 5 wells was the final OD
value. The cell proliferating curve was sketched with the time as
the horizontal axis and OD value as the vertical axis. The
suppression rate of A549 cell proliferation = (1 − OD value of
KD)/OD value of CON ×100%.
Cell cycle and apoptosis by flow
cytometry (FCM)
A549 cells (1×106) of each group were
digested and centrifuged for 5 min. Supernatants were discarded.
Cells were washed with ice-cold PBS, fixated with 70% ethanol,
centrifuged and collected. The sedimentation was washed with PBS.
PI dye (1000 μl of 2 mg/ml solution) was added and mixed. The
mixture was dyed for 30 min at 4˚C lucifugally. Red fluorescence at
a wavelength of 488 nm was tested by FACScalibur and recorded. The
data were analyzed with cell cycle fitting software.
Statistical analysis
Data were processed by SPSS15.0 statistical
software. Quantitative data was expressed as the means ± standard
deviation. One-way ANOVA was performed between different groups.
Dunnett’ t-test (when homogeneity of variances existed) or Dunnett
T3 (when heterogeneity of variances existed) were used to calculate
the values between the different groups. P<0.05 indicated a
statistically significant difference.
Results
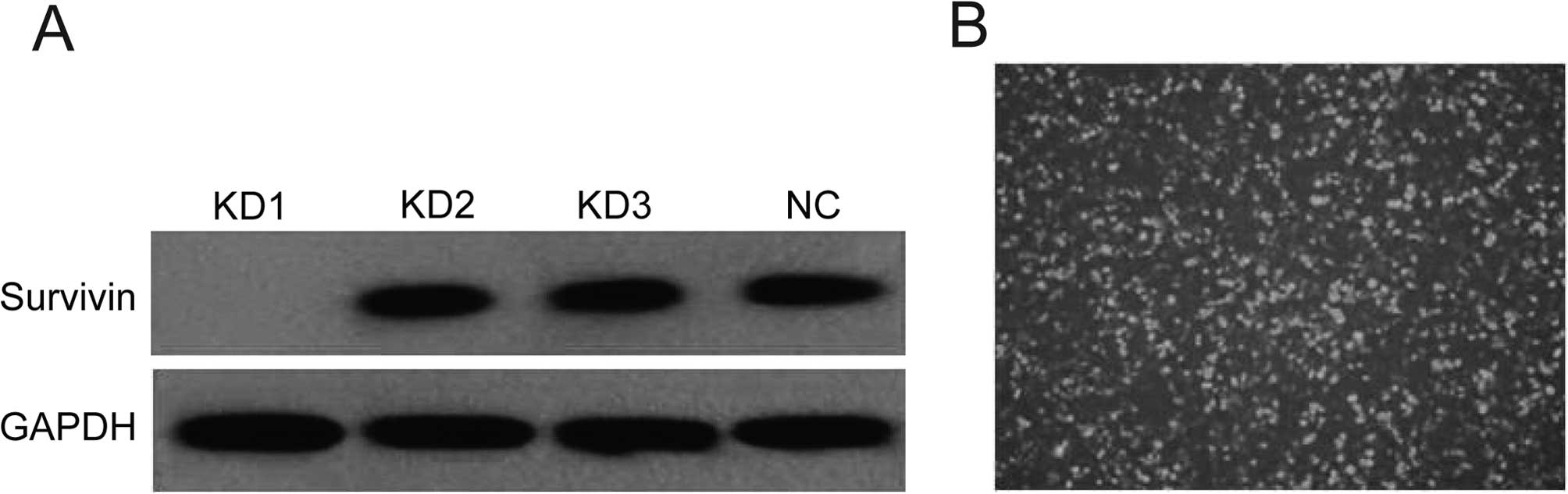
The screening outcome of survivin-siRNA
lentiviral vector
As the results of western blot analysis showed, the
first pair of siRNA was the most efficient to downregulate the
expression (Fig. 1A). Therefore we
selected the first pair for the recombination of the survivin-siRNA
lentiviral vector. After the survivin-siRNA lentiviral vector was
transfected into A549 cells, the cells had green fluorescence
(Fig. 1B). Screening and
amplification with G418 was then carried out.
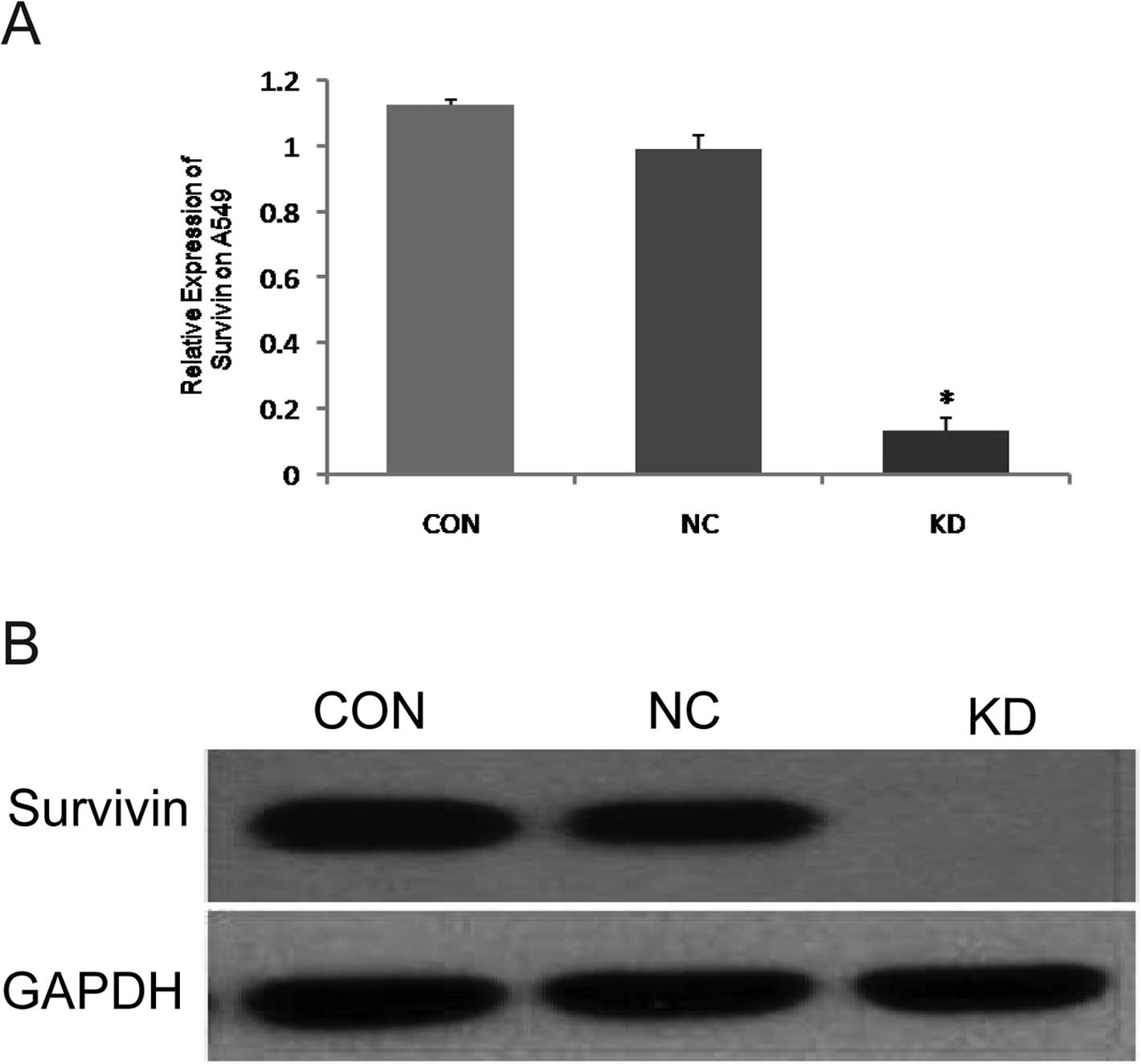
The effect of survivin-siRNA on survival
of lung cancer cells
As the results of RT-qPCR showed, the expression of
survivin mRNA in the KD group was significantly lower than that in
the NC and CON groups (P<0.05) (Fig. 2A), which was consistent with the
results of western blot analysis. Both results signified that the
expression of survivin protein was significantly lower in the KD
than in the NC and CON groups (P<0.05) (Fig. 2B).
The effect of survivin-siRNA on lung
cancer cell proliferation
The cells’ proliferative curves were sketched based
on the OD values from days 1 to 5 of each group. The differences of
the initial OD values among the three groups were not significant,
(CON, 0.18±0.02; NC, 0.17±0.01; KD, 0.16±0.01; P>0.05). However,
the difference in OD values on day 5 between the CON and NC group
was not significant (CON, 1.44±0.01; NC, 1.13±0.44; P>0.05) and
the proliferative activity of the KD group significantly decreased
(KD, 0.80±0.03; P<0.05). The OD values of the KD group on days
3, 4 and 5 were 36.0, 43.1 and 44.6%, respectively (Fig. 3).
The effect of survivin-siRNA on cell
cycle of lung cancer cells and apoptosis
FCM showed that the cell cycle of A549 cells in the
KD group changed significantly: the proportion in the G1 phase
increased and the proportion in the S phase decreased. The
differences were significant (P<0.05). G2/M phase did not change
significantly, though it also showed a reductive tendency (Table III and Fig. 4). The apoptical rate of A549 cells
in the KD group was 10.68±0.28%, which was significantly higher
than that in the NC (6.68±0.44%) and CON (4.97±0.33%) groups
(P<0.05).
| Table IIIEffect of survivin-siRNA on cell
cycle of A549 cells (%, mean ± SD, n=3). |
Table III
Effect of survivin-siRNA on cell
cycle of A549 cells (%, mean ± SD, n=3).
| Group | G0/G1 | S | G2/M |
|---|
| CON | 61.30±0.59a | 30.43±0.68a | 8.28±0.79b |
| NC | 59.19±1.46a | 33.98±1.91a | 6.83±1.63b |
| KD | 66.62±0.59 | 27.34±0.92 | 6.03±1.12 |
The activity of caspase-9 after survivin
interference
The expression of caspase-9 protein in the KD group
was significantly higher than that in the CON and NC groups after
survivin-siRNA transfection (P<0.05). However, the differences
between the CON and NC group were not significant (P>0.05)
(Fig. 5).
Discussion
Currently, lung cancer has the highest rates of
morbidity and mortality of all malignant tumors worldwide. Since
the number of lung cancer patients in China ranked first in the
world, it is an urgent and important task to prevent and treat lung
cancer, which should be the key point in the tumor-control plan of
China. However, the controversial pathogenesis of lung cancer
hinders the methods of prevention and treatment (9,10).
As modern tumor molecular biology shows, the pathogenesis and
development of lung cancer is a complex biological process, in
which multiple genes are involved, multiple factors react and
multiple phases exist. The pathogenesis and development of lung
cancer could be attributed to the activation of an oncogene or the
inactivation of a tumor inhibitor gene, which can induce abnormal
proliferation, differentiation and apoptosis. Therefore, targeted
therapy could be a new method for the prevention and treatment of
lung cancer (11,12).
The IAP protein family (IAP) plays a key role in
inhibiting apoptosis and is, therefore, more important than the BCL
family. Survivin, a new member of the IAP family, has the least
molecular weight, but the strongest capacity. Survivin can not only
regulate the apoptosis, but also control cell cycle. These dual
functions are an exclusive characteristic of the IAP family.
Therefore, survivin plays a significant role in maintaining the
quick proliferation, growth and the malignant functions of cells
(13). Survivin is specifically
expressed in embryo tissue and in developing fetal tissue, but is
not expressed in adult terminal differentiated tissue. It is also
widely expressed in certain cancer tissue, such as non-small cell
lung cancer (NSCLC), gastric and intestinal cancer. Singha et
al (14) concluded that
survivin is one of the most significant independent prognostic
factors of NSCLC. The unique distribution of survivin in the tissue
means that normal tissue is almost not affected by the inhibition
of survivin protein expression. Therefore survivin could be an
ideal target gene for antitumor antisense therapy. To date,
possible mechanisms for inhibiting apoptosis by survivin have been
investigated (15,16). These include: i) the direct or
indirect inhibition of the activities of caspases (caspase-3,
caspase-7, caspase-6, caspase-8, caspase-9 and caspase-10); ii)
competitive interaction with cyclin-dependent kinase (CDK) cdk4/P21
complex, and release of P21; then P21 is conjugated with caspase-3;
conjugated with the second mitochondrial-derived activator/direct
IAP binding protein with low PI (Smac/DIABLO) and isolated from
other IAPs. Studies on survivin in cell mitosis have shown that it
has a mutual function with the inner centromere protein (INCENP)
and Aurora B. These genes have been shown to inhibit the expression
of survivin, destroy microtubule organization and induce the
failure of polyploidy formation and cell mitosis. Yonesaka et
al (17) transfected the lung
cancer cell line PC9 (including mutant p53) with survivin-siRNA,
and found that cell proliferation was retarded. Chao et al
(18) found that the expression of
survivin mRNA was downregulated, the polykaryocyte formed and
apoptosis occured 48 h after transfecting the A549 cells with
survivin-siRNA.
In the present study, RT-qPCR and western blot
analysis proved that the expressions of survivin transcription and
translation were significantly inhibited after transfecting A549
cells with the survivin-siRNA lentiviral vector. These results also
confirmed the successful construction of survivin-siRNA lentiviral
vector. Furthermore, the results also showed that the expressions
of survivin mRNA and protein were downregulated in the KD group,
and the proliferation and growth of lung cancer cells were clearly
inhibited. FCM showed the significant apoptosis of lung cancer
cells. Most cells were blocked in the G1 phase, leading to the
decrease of cells in the S phase. The results of FCM proved that
survivin played an important role in the shift from the G1 to the S
phase; it also demonstrated that after silencing the expression of
survivin, the inhibition of cell proliferation was caused not only
by the increase of apoptosis, but also by the decrease in the
mitotic activity of the cells. These results are consistent with
those from the study by Suzuki et al (19), who proved that the overexpression
of survivin can accelerate the shift from the G1 to the S phase and
resist G1 inhibition. The classical passages of cell apoptosis
included the mitochondrial and death receptor pathways. At present,
the anti-apoptotic mechanisms of survivin have not been fully
elucidated. Some studies have shown that survivin can directly
suppress the activities of caspase-3 and caspase-7, while others
have shown survivin to mainly suppress the caspase-9-dependent
apoptotic pathway. Thr34 phosphorylated survivin can directly bind
with caspase-9 and inactivate it or indirectly block apoptosis by
binding with SMAC (20). Banks
et al and Shi (21,22), observed that the activity of
caspase-3 did not change after suppressing the expression of
survivin. They found that the keratinocytes isolated from
K14-survivin mice highly expressed survivin, and that the
keratinocytes could resist apoptosis mediated by UVB, an activator
of the caspase-9-dependent apoptotic pathway. Apoptosis induced by
a mutant type of survivin (T34A) could be blocked by the antagonist
of caspase-9. All the results proved that survivin should suppress
caspase-9-dependent apoptosis, not caspase-3-dependent. O’Connor
et al (23) showed the
direct interaction between survivin and caspase-9. Our research
revealed that in the KD group, the activity of caspase-9 clearly
increased while the expressions of survivin mRNA and protein
decreased. Therefore, it may be through the activation of caspase-9
that the silencing expression of survivin could decrease the
resistance of lung cancer cells to apoptosis and increase the
susceptibility of the cells to apoptotic factors.
In brief, our study shows that the RNA interference
technique can effectively silence the expression of survivin,
inhibit lung cancer cell growth and induce apoptosis in lung cancer
cells. It may also be related to the activation of caspase-9. The
details of this mechanism require further clarification.
Nevertheless, survivin may be promising for antitumor targeted
therapy (24,25).
Acknowledgements
This study was supported by grants from the Natural
Science Foundation of Fujian Province (No. 2008J0284) and Science
and Technology Project of the Fujian Provincial Department of
Education (No. JA09121).
References
|
1
|
P SeveC DumontetChemoresistance in
non-small cell lung cancerCurr Med Chem Anticancer
Agents57388200510.2174/156801105335260415720263
|
|
2
|
MW LeeDS KimNY MinHT KimAkt1 inhibition by
RNA interference sensitizes human non-small cell lung cancer cells
to cisplatinInt J
Cancer12223802384200810.1002/ijc.2337118224693
|
|
3
|
AQ DongMJ KongZY MaJF QianXH
XuDown-regulation of IGF-IR using small, interfering, hairpin RNA
(siRNA) inhibits growth of human lung cancer cell line A549 in
vitro and in nude miceCell Biol
Int31500507200710.1016/j.cellbi.2006.11.01717196841
|
|
4
|
E AshiharaRNA interference for cancer
therapiesGan To Kagaku Ryoho3720332041201021084799
|
|
5
|
N NakashimaCL HuangD LiuM UenoH
YokomiseIntratumoral Wnt1 expression affects survivin gene
expression in non-small cell lung cancerInt J
Oncol37687694201020664938
|
|
6
|
I PorebskaE SobanskaM KosackaR
JankowskaApoptotic regulators: P53 and survivin expression in
non-small cell lung cancerCancer Genomics
Proteomics7331335201021156966
|
|
7
|
W ZhengX MaD WeiT WangY MaS YangMolecular
cloning and bioinformatics analysis of a novel spliced variant of
survivin from human breast cancer cellsDNA
Seq16321328200516329164
|
|
8
|
ME HornPM WaterhouseRapid match-searching
for gene silencing
assessmentBioinformatics2619321937201010.1093/bioinformatics/btq31820601440
|
|
9
|
N IchiteM ChouguleAR PatelT JacksonS SafeM
SinghInhalation delivery of a novel diindolylmethane derivative for
the treatment of lung cancerMol Cancer
Ther930033014201010.1158/1535-7163.MCT-09-110420978159
|
|
10
|
A BiranM BrownsteinR HaklaiY
KloogDownregulation of survivin and aurora A by histone deacetylase
and RAS inhibitors: a new drug combination for cancer therapyInt J
Cancer128691701201110.1002/ijc.2536720473860
|
|
11
|
DC AltieriValidating survivin as a cancer
therapeutic targetNat Rev Cancer34654200310.1038/nrc96812509766
|
|
12
|
F CappuzzoFR HirschE RossiEpidermal growth
factor receptor gene and protein and gefitinib sensitivity in
non-small cell lung cancerJ Natl Cancer
Inst97643655200510.1093/jnci/dji11215870435
|
|
13
|
F LiG AmbrosiniEY ChuControl of apoptosis
and mitotic spindle checkpoint by
survivinNature396580584199810.1038/251419859993
|
|
14
|
S SinghalA VachaniD Antin-OzerkisLR
KaiserSM AlbeldaPrognostic implications of cell cycle, apoptosis,
and angiogenesis biomarkers in non-small cell lung cancer: a
reviewClin Cancer
Res1139743986200510.1158/1078-0432.CCR-04-266115930332
|
|
15
|
A SuzukiT ItoH KawanoSurvivin initiates
procaspase 3/p21 complex formation as a result of interaction with
Cdk4 to resist Fas-mediated cell
deathOncogene1913461353200010.1038/sj.onc.120342910713676
|
|
16
|
SK ChiouMK JonesAS TarnawskiSurvivin - an
anti-apoptosis protein: its biological roles and implications for
cancer and beyondMed Sci Monit9PI2529200312709681
|
|
17
|
K YonesakaK TamuraT KurataSmall
interfering RNA targeting survivin sensitizes lung cancer cell with
mutant p53 to adriamycinInt J
Cancer118812820200610.1002/ijc.2135016108013
|
|
18
|
JI ChaoHF LiuThe blockage of survivin and
securin expression increases the cytochalasin B-induced cell death
and growth inhibition in human cancer cellsMol
Pharmacol69154164200616219911
|
|
19
|
A SuzukiM HayashidaT ItoSurvivin initiates
cell cycle entry by the competitive interaction with
Cdk4/p16(INK4a) and Cdk2/cyclin E complex
activationOncogene1932253234200010.1038/sj.onc.120366510918579
|
|
20
|
AM VerhagenDL VauxCell death regulation by
the mammalian IAP antagonist
Diablo/SmacApoptosis7163166200210.1023/A:101431861595511865200
|
|
21
|
DP BanksJ PlesciaDC AltieriSurvivin does
not inhibit caspase-3 activityBlood9640024003200011186274
|
|
22
|
Y ShiSurvivin structure: crystal
unclearNat Struct Biol7620623200010.1038/7790410932241
|
|
23
|
DS O’ConnorD GrossmanJ PlesciaRegulation
of apoptosis at cell division by p34cdc2 phosphorylation of
survivinProc Natl Acad Sci USA971310313107200011069302
|
|
24
|
JS JangKM KimKH KangPolymorphisms in the
survivin gene and the risk of lung cancerLung
Cancer603139200810.1016/j.lungcan.2007.09.00817961802
|
|
25
|
NH NabilsiRR BroaddusDS LooseDNA
methylation inhibits p53-mediated survivin
repressionOncogene2820462050200910.1038/onc.2009.6219363521
|