Introduction
Cells become cancerous as a result of mutation of
normal genes, resulting in continuous growth and survival. Human
gastric cancer is linked to irregular eating habits and intake of
fast food (1). In recent years,
studies on cancer cell apoptosis and inhibition of cancer cell
invasion has been conducted (2).
In particular, various seaweeds have been reported to inhibit
cancer cell growth (3–5). The majority of studies on seaweeds
are conducted in Japan and Korea (6,7).
Anti-tumor activity in HT-29 colon cancer cells using Laminaria
japonica(8) and the recovery
of damaged liver cells using Hizikia fusiformis (H.
fusiformis) glycoprotein have been previously reported
(9). In addition, inhibition of
the growth of AGS human gastric cancer cells by a H.
fusiformis extract has been noted (10).
Capsosiphon fulvescens (C. fulvescens)
is a green alga that grows in clean areas off the Korean coast.
C. fulvescens has long been a traditional Korean food and
exhibits anti-cancer effects in addition to improving hangover
symptoms, immune activity and anticoagulant activity. Anti-cancer
effects of C. fulvescens components have been reported
(11,12). Prior to the present study, C.
fulvescens glycoprotein treatment was reported to induce
apoptosis of human gastric cancer AGS cells via Fas signaling
(13). Increased β-catenin levels
increase cell adhesion, leading to invasion and metastasis. These
processes are accelerated in cancer cell metastasis and invasion
mediated by adhesion between adjacent cancer cells; this is
facilitated by increased β-catenin levels. Although cancer cell
apoptosis is induced by Fas, β-catenin levels were decreased.
Therefore, apoptosis was examined via Fas signaling; however, as
invasion was not inhibited, further experiments were conducted.
The inhibitory effect of C. fulvescens on
invasion by human gastric cancer AGS cells was also evaluated.
Tight junction (TJ) and matrix metalloproteinase (MMP) proteins are
overexpressed in a number of cancer cells and are associated with
their invasive properties (14).
Although MMP and TJ protein levels are increased, the
transepithelial electrical resistance (TEER) value is lower. TEER
facilitates material transfer between cells due to disrupted cell
membrane permeability (15,16).
In addition, cancer cell adhesion, metastasis, proliferation and
invasion increased. Therefore, studies of apoptosis signaling in
cancer cells, as well as invasion and metastasis, have been
performed recently. Therefore, in the present study, TJ proteins,
claudin, zo-1 and occludin, and the cell adhesion-related
molecules, β-catenin and E-cadherin, were investigated. C.
fulvescens glycoprotein (Cf-GP) controls protein expression,
which is associated with cancer cell invasion. Results indicate
that Cf-GP treatment inhibits invasion and development of AGS human
gastric cancer cells.
Materials and methods
Preparation of Cf-GP
Cf-GP was purchased in 2006 in Korea. C.
fulvescens powder (40 g) was diluted in 1 liter water and
stirred for 3 h at 80°C using a heating mantle, followed by
centrifugation at 1,500 × g for 15 min at 4°C.
Three volumes of 95% ethanol were added and
precipitate was removed by vacuum filtration. To the supernatants,
80% ammonium sulfate was added, followed by stirring for 24 h.
Next, salt was removed by membrane dialysis (Por Membrane MW 3,500
Da, Spectrum Laboratories Inc., Rancho Dominguez, CA, USA) for 1
day at 4°C. The concentrated solution was aliquoted into 1.5 ml
tubes and stored at −70°C until use. These samples are hereafter
termed Cf-GP.
Cell culture
Human gastric cancer AGS cells (American Type
Culture Collection, Manassas, VA, USA) were maintained at 37°C in a
5% CO2 humidified atmosphere. Cells were cultured in
RPMI-1640 medium with 10% fetal bovine serum (FBS; Hyclone, Logan,
UT, USA), 100 U/l penicillin and 100 mg/l streptomycin. Cells were
cultured to 60–80% confluence in 100-mm diameter dishes. The medium
was replaced every day.
Cell proliferation assays
AGS cell proliferation was measured using a
CellTiter 96® aqueous non-radioactivity cell
proliferation assay (Promega Corporation, Madison, WI, USA). The
assay is based on the cleavage of
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxy-phenyl)-2-(4-sulfonyl)-2H-tetrazolium
(MTS) into a formazan product that is soluble in tissue culture
media. Cells were seeded onto 96-well plates at 2×104
cells/well in 100 μl medium. Cells were maintained for 24 h and the
medium was then replaced with serum-free medium (SFM). Following 24
h, the medium was replaced with SFM containing Cf-GP (0, 5, 10 or
20 μg/ml) for 24 h. Cells were then incubated with MTS solution for
30 min at 37°C. Cell proliferation was measured by means of
absorbance at 490 nm using the Benchmark enzyme-linked
immunosorbent assay (ELISA) plate reader (Bio-Rad, Hercules, CA,
USA).
Cell invasion assays
Cell invasion was measured using an 8.0-μm pore size
insert in Transwell® plates (Corning Costar Inc.,
Corning, NY, USA). AGS cells were seeded into the upper chamber
(Matrigel coated; Corning Costar Inc.) and maintained for 24 h. The
medium was then replaced with SFM containing Cf-GP (0, 5, 10 or 20
μg/ml) for 24 h. The lower chamber was maintained with 10% FBS.
Following 24 h, the cut on the bottom of the filter (Matrigel
coated) was stained with hematoxylin. Stained cells were calculated
by extrapolation from the number counted.
TEER assay
Using a voltohmmeter (EVOM Epithelial Tissue
Voltohmmeter; World Precision Instruments, Sarasota, FL, USA), TEER
was measured. AGS cells were seeded into the upper chamber
(Matrigel coated) of the transwell plates and maintained for 24 h.
Next, SFM was replaced with medium containing Cf-GP (0, 5, 10 or 20
μg/ml) for 24 h. The lower chamber was maintained with 10% FBS.
Following 24 h, the upper chamber was separated and TEER values
were determined using the voltohmmeter.
TEER was calculated using the following formula:
TEER [(resistance (Ω cm2)] = (Ω - background Ω) ×
membrane area (cm2); background resistance was 14 and
the membrane area was 1.54 cm2. The change in TEER
values for each insert was calculated using the following formula:
change in TEER (%) = TEER (Ω cm2)/initial TEER (Ω
cm2) - 100.
mRNA expression
AGS cells were seeded into six-well plates at
2×104 cells/well in 2 ml medium. Cells were incubated
for 24 h and the medium was replaced with SFM. Following 24 h, the
medium was replaced again with SFM containing Cf-GP (0, 5, 10 or 20
μg/ml) for 24 h. Cells were treated with the TRIzol reagent
(Invitrogen Life Technologies, Carlsbad, CA, USA) and the RNA
extracted was quantified using Oligo(dT) primers (Intron
Biotechnology Co. Ltd., Seongnam, Korea); the corresponding cDNA
was then synthesized. cDNA was subjected to amplification using a
PCR kit (dNTP mix, 10X Ex Taq Buffer and Ex Taq;
Takara Bio, Inc., Shiga, Japan) with primers (Table I) in 0.1% diethylpyrocarbonate
water. PCR products were resolved on 1% agarose gels. Gels were
stained with 10 mg/ml ethidium bromide to visualize amplification
products.
 | Table IOligonucleotide sequences of the
primer pairs used for RT-PCR. |
Table I
Oligonucleotide sequences of the
primer pairs used for RT-PCR.
| Sequence of primers
(5′-3′) |
|---|
|
|
|---|
| Name | Sense | Antisense |
|---|
| MMP-2 |
GGC-CCT-GTC-ACT-CCT-GAG-AT |
GGC-ATC-CAG-GTT-ATC-GGG-GA |
| MMP-9 |
CGG-AGC-ACG-GAG-ACG-GGT-AT |
TGA-AGG-GGA-AGA-CGC-ACA-GC |
| TIMP-1 |
TGG-GGA-CAC-CAG-AAG-TCA-AC |
TTT-TCA-GAG-CCT-TGG-AGG-AG |
| β-catenin |
GAA-ACG-GCT-TTC-AGT-TGA-GC |
CTG-GCC-ATA-TCC-ACC-AGA-GT |
| E-cadherin |
GAA-CAG-CAC-GTA-CAC-AGC-CCT |
GCA-GAA-GTG-TCC-CTG-TTC-CAG |
| Claudin-1 |
TCA-GCA-CTG-CCC-TGC-CCC-AGT |
TGG-TGT-TGG-GTA-AGA-GGT-TGT |
| Claudin-2 |
ACA-CAC-AGC-ACA-GGC-ATC-AC |
TCT-CCA-ATC-TCA-AAT-TTC-ATG-C |
| Claudin-3 |
AAG-GCC-AAG-ATC-ACC-ATC-GTG |
AGA-CGT-AGT-CCT-TGC-GGT-CGT |
| Claudin-4 |
TGG-ATG-AAC-TGC-GTG-GTG-CAG |
GCA-GAA-GTG-TCC-CTG-TTC-CAG |
| Occludin |
TCA-GGG-AAT-ATC-CAC-CTA-TCA-CTT-CAG |
CAT-CAG-CAG-CAG-CCA-TGT-ACT-CTT-CAC |
| Zo-1 |
GAA-GCT-TCA-TCT-CCA-GTC-CCT |
TGG-GTA-GGG-CTG-TTT-GTC-ATC-ATA |
| β-actin |
CGT-ACC-ACT-GGC-ATC-GTG |
GTG-TTG-GCG-TAC-AGG-TCT-TTG |
Western blot analysis
AGS cells were cultured in 100-mm diameter dishes.
Cells were cultured to 60–80% confluence and the medium was
replaced with SFM for 4 h. Next, the medium was replaced with fresh
SFM containing Cf-GP (0, 5, 10 or 20 μg/ml) for 24 h. Cells were
washed with phosphate-buffered saline and added to lysis buffer [50
mM Tris-HCl (pH 7.4), 150 mM NaCl, 1 mM EGTA, 1% NP-40, 1 mM NaF, 1
mM Na3VO4, 1 μg/ml aprotinin, 1 μg/ml
leupeptin, 1 μg/ml pepstatin A, 0.25% Na-deoxycholate and 1 mM
PMSF]. Lysates were separated using 10–15% SDS-PAGE and transferred
onto polyvinylidene fluoride membranes (Millipore, Billerica, MA,
USA). The membranes were blocked with 1% bovine serum albumin in
TBS-T [10 mM Tris-HCl (pH 7.5), 150 mM NaCl and 0.1% Tween 20] at
room temperature and incubated with agitation with specific
antibodies: anti-claudin-1, −2, −3 and −4 (1:1,000), anti-β-catenin
(1:1,000), anti-E-cadherin (1:1,000), anti-MMP-2 and −9 (1:1,000)
and anti-tissue inhibitors of metalloproteinases-1 (TIMP-1;
1:1,000; all from Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA). The secondary peroxidase-conjugated goat, mouse and rabbit
antibodies (1:10,000) were purchased from GE Healthcare
Bio-Sciences (Piscataway, NJ, USA). Bands were visualized using
Super Signal West Pico Stable Peroxide solution and the Super
Signal West Pico Luminol/Enhancer solution (Pierce Biotechnology,
Inc., Rockford, IL, USA) and developed using Kodak X-ray film
(Eastman Kodak Company, Rochester, NY, USA).
Gelatin zymography
AGS cells were cultured in six-well plates to 60–80%
confluence and the medium was replaced with SFM for 4 h. Next, the
medium was replaced with fresh SFM containing Cf-GP (0, 5, 10 or 20
μg/ml) for 24 h. The obtained conditioned medium was loaded in 10%
SDS-free acrylamide gels with 0.1% gelatin. The completed loading
gel was treated with 2.5% Triton X-100 and incubated under
agitation for 30 min, followed by incubation with developing buffer
(50 mM Tris-HCl, 150 mM NaCl, 5 mM CaCl2 and 1 μM
ZnCl2; pH 7.5) at 37°C for 2 days. The gel was fixed (7%
acetic acid) and stained (0.5% Coomassie Brilliant Blue 250 in
dilute fixing solution).
Statistical analysis
Data are presented as the mean ± SD and were
calculated using SPSS version 10.0 (SPSS, Inc., Chicago, IL, USA).
Data were validated by analysis of variance (ANOVA). P<0.05 was
considered to indicate a statistically significant difference and
was determined by Duncan's multiple range test for group
comparisons.
Results
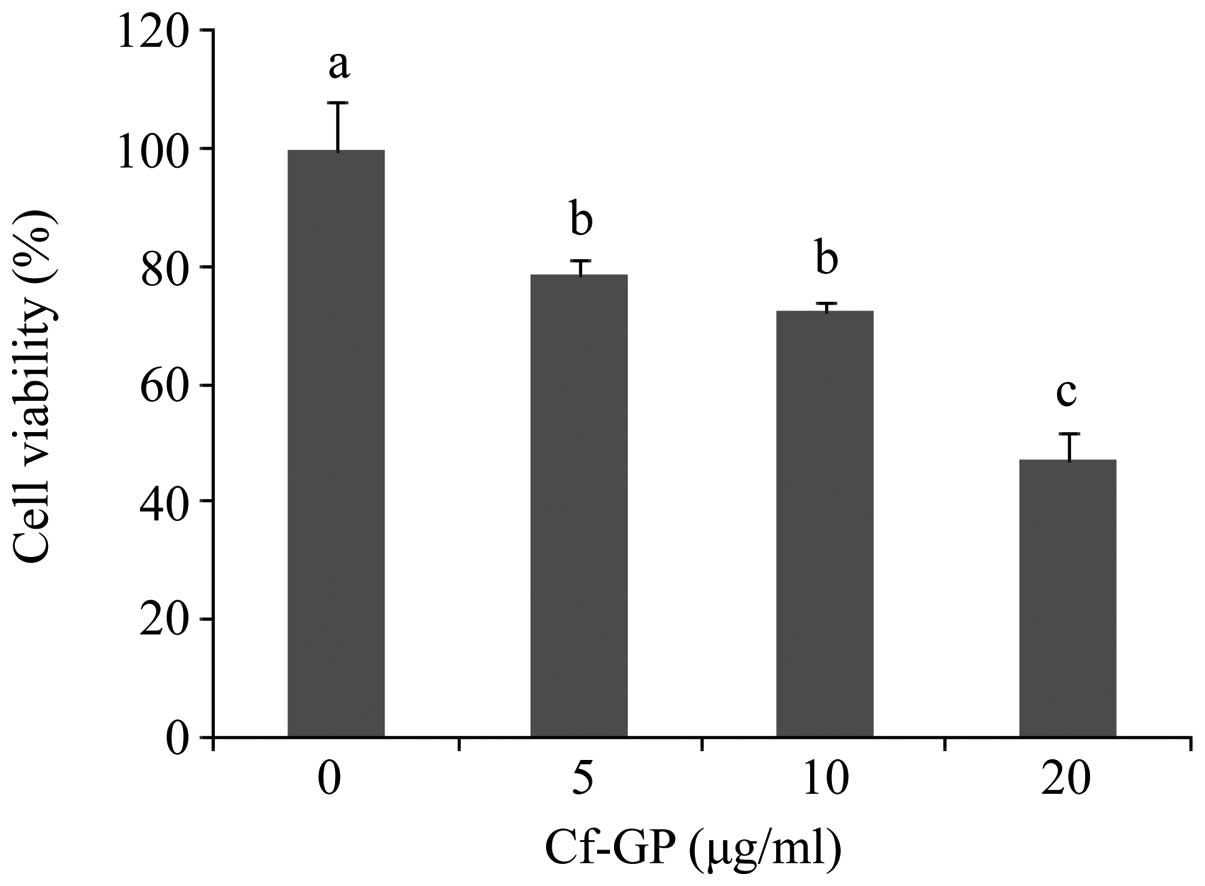
Cf-GP inhibits AGS cell
proliferation
MTS assays were used to investigate the effect of
Cf-GP (0, 5, 10 or 20 μg/ml) on AGS cell proliferation. Cf-GP at 20
μg/ml resulted in a 50% decrease in proliferation (Fig. 1). In addition, Cf-GP inhibited AGS
cell growth in a dose-dependent manner. We previously reported that
Cf-GP induced proliferation of human intestinal epithelial IEC-6
cells (17).
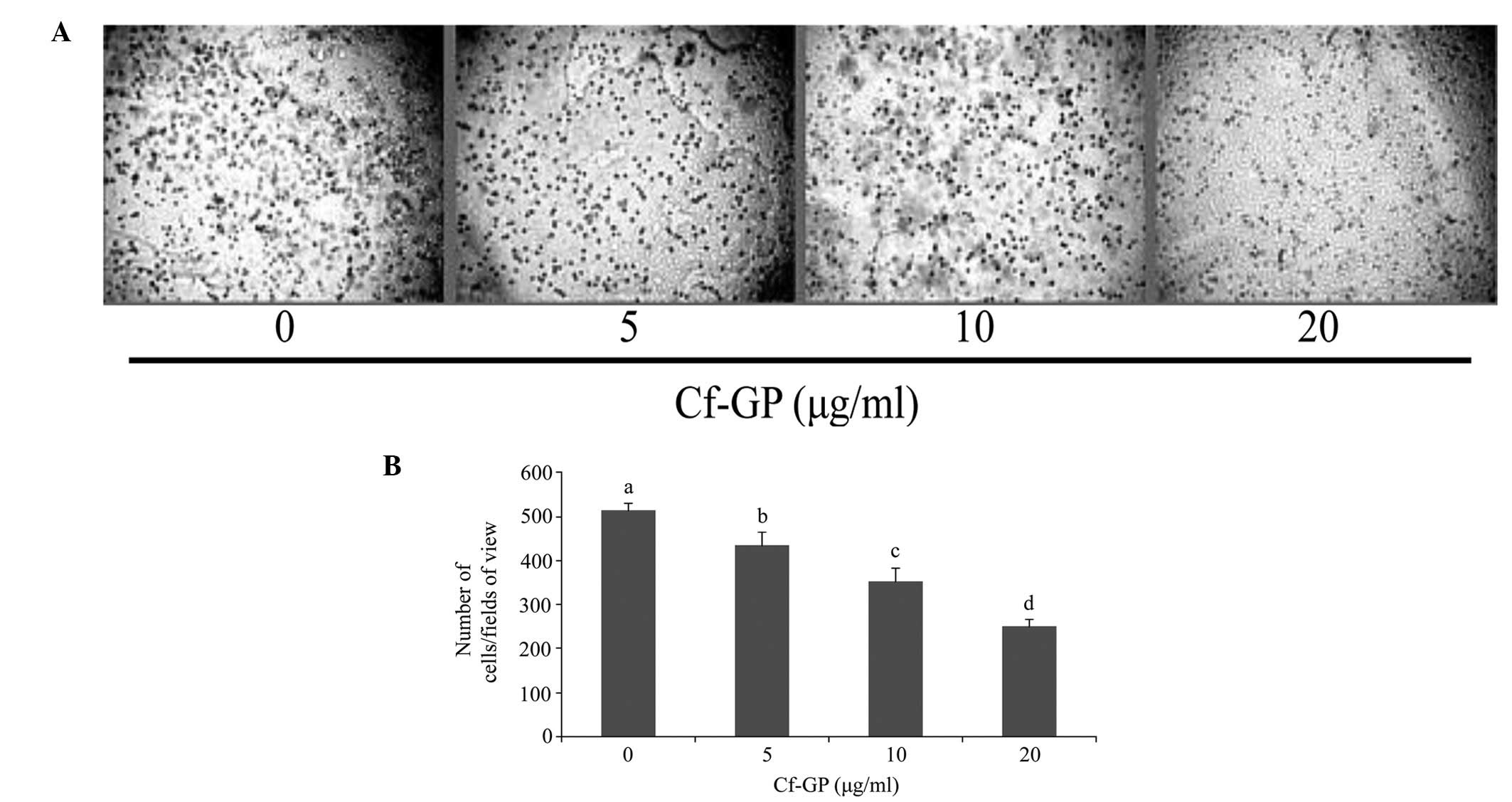
Cf-GP inhibits AGS cell invasion
In general, increased proliferation of cancer cells
to other tissues is a result of accelerated invasion. Since Cf-GP
inhibited AGS cell growth, the effect of Cf-GP on invasion was
investigated using hematoxylin staining (Fig. 2A). As demonstrated in Fig. 2B, the Cf-GP treatment group
exhibited decreased cell invasion compared with control cells.
Treatment with 20 μg/ml Cf-GP led to a 50% reduction compared with
the control group.
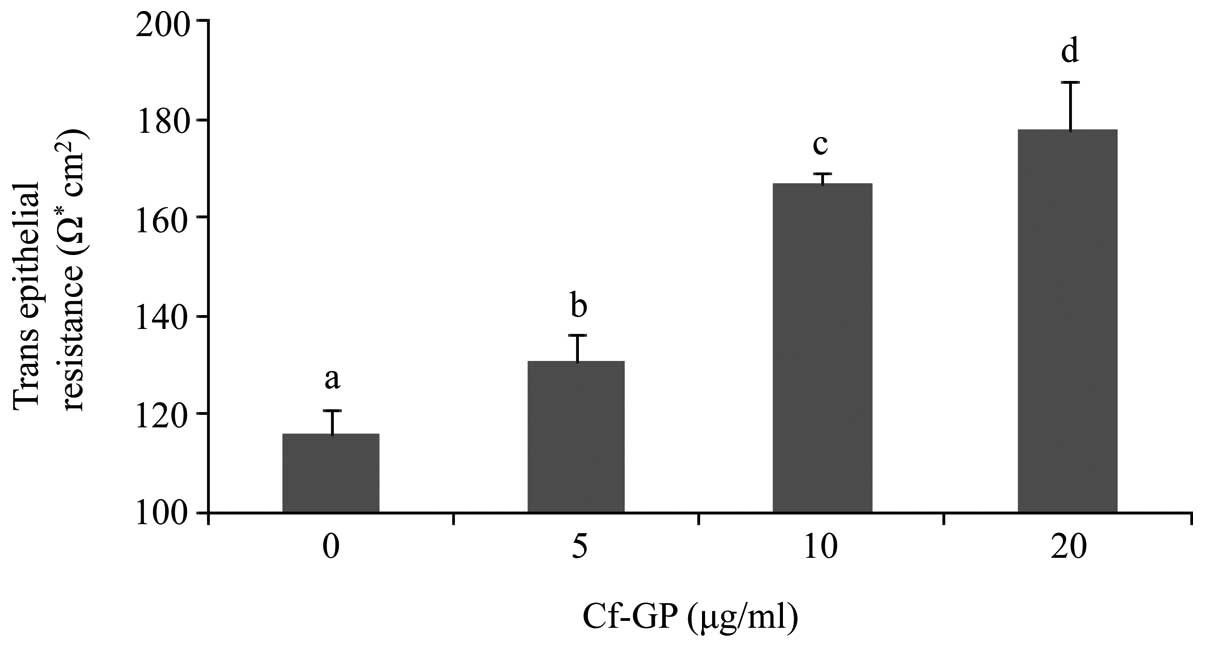
Cf-GP increases TEER values in AGS
cells
Increased cell membrane permeability is mediated by
weakening of TEER. We thus evaluated the effect of Cf-GP on AGS
cell invasion by measuring TEER. Decreases of TEER values allowed
easier penetration of intracellular material. Normal cells indicate
higher TEER values by the strong electrical resistance of the cell
membrane. However, MMP activity induced degradation of cell
membranes and permeability in cancer cells. Therefore, cancer cells
have lower TEER values and this increases invasion. In the present
study, TEER values in AGS cells were increased upon Cf-GP treatment
in a dose-dependent manner (Fig.
3).
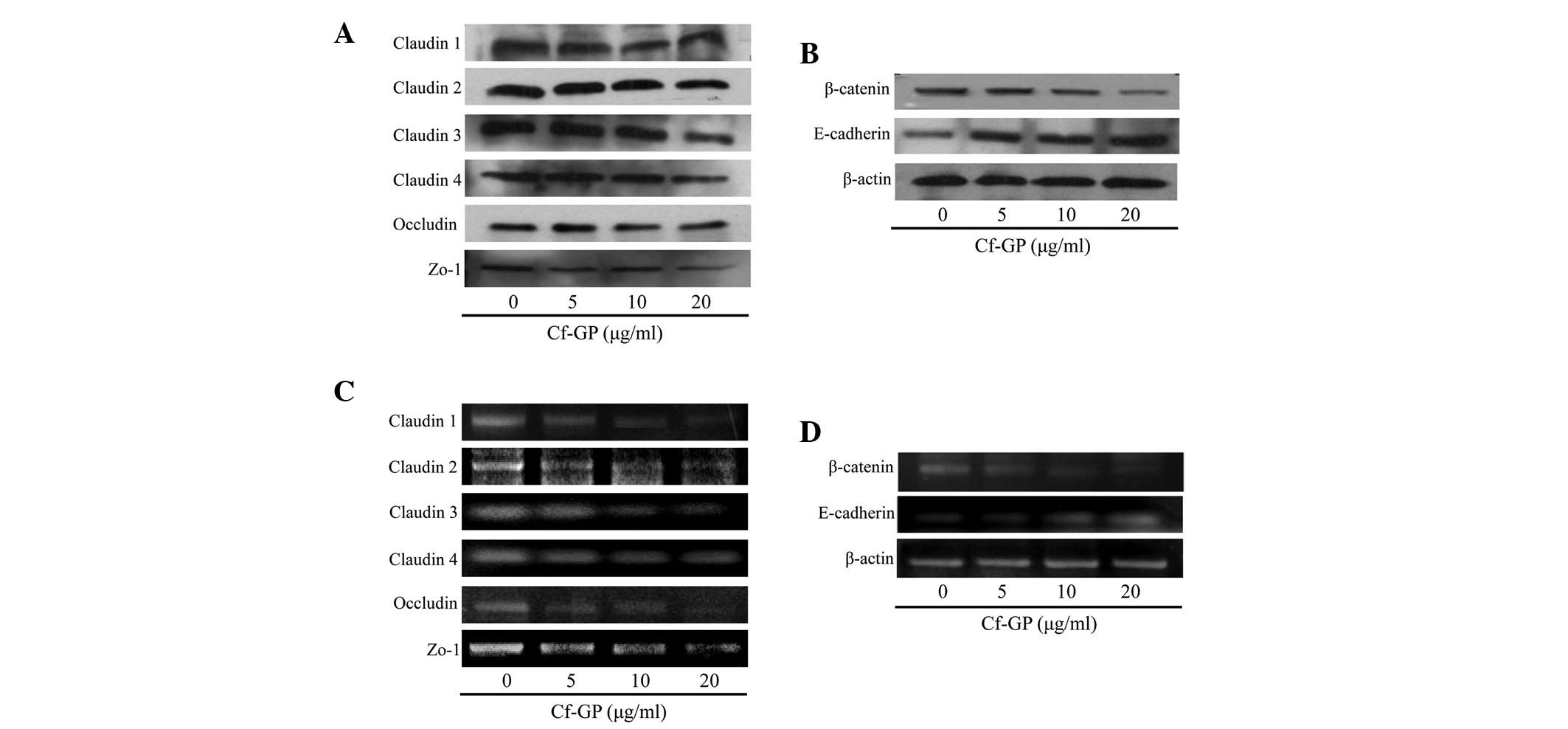
Effect of Cf-GP on TJ- and
metastasis-associated protein expression
The increased membrane permeability in cancer cells
is due to mutations in a variety of genes associated with TJ
proteins. Typically, overexpression of TJ proteins (claudin, zo-1
and occludin) and TJ-related proteins (β-catenin) is observed in
cancer cells. In addition, β-catenin levels are increased due to
loss of E-cadherin. Therefore, β-catenin increases cancer cell
adhesion. In the present study, protein and mRNA levels were
determined by western blot analysis and RT-PCR, respectively. As
demonstrated in Fig. 4A and B, in
control cells, expression levels of the TJ proteins, claudin, zo-1
and occludin increased, but were reduced in a dose-dependent manner
upon Cf-GP treatment. In addition, β-catenin levels were reduced
while that of the inhibitor of β-catenin, E-cadherin, was increased
by Cf-GP treatment at the protein and mRNA levels (Fig. 4C and D).
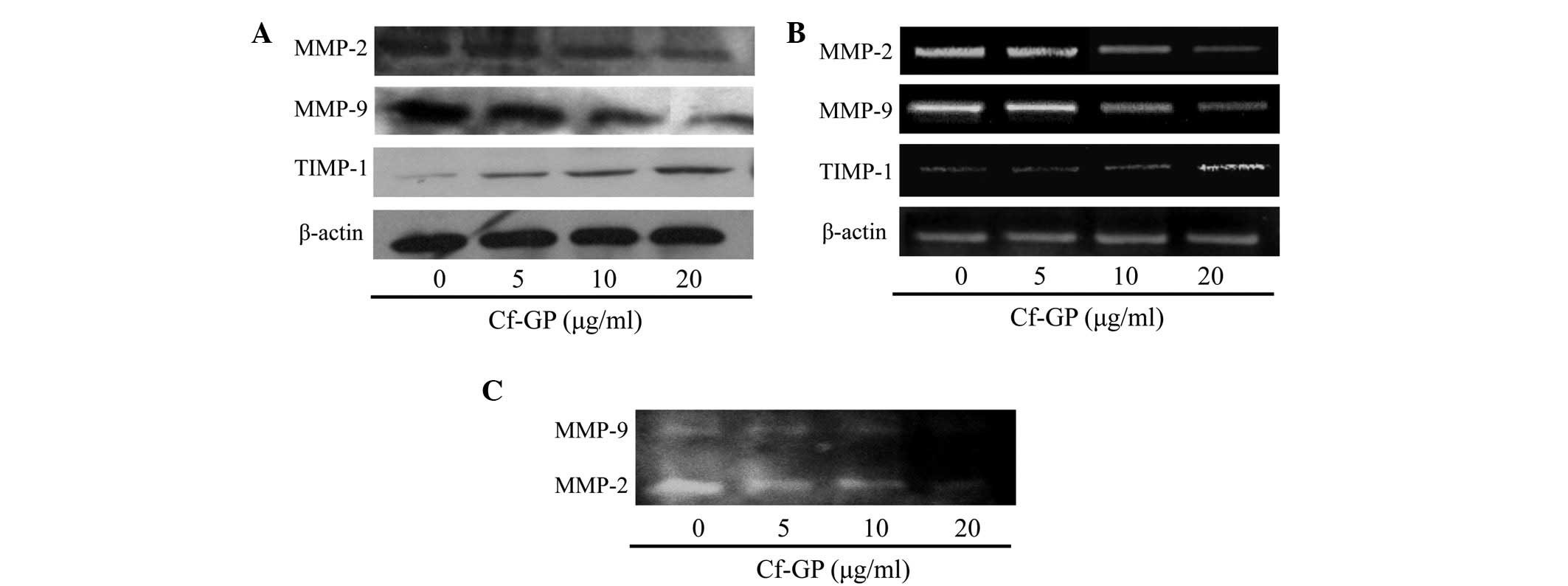
Cf-GP inhibits MMP activity and
expression
The interaction between cancer cells and the
basement membrane is a major step in metastasis and invasion
(18). The basement membrane is
composed of substances, including collagen and lamin. Cell membrane
degradation by MMP proteins and cancer cells is an important
process for invasion and metastasis. In particular, MMP-2 and MMP-9
are important mediators of basement membrane degradation of gelatin
and factors involved in angiogenesis and cancer cell invasion are
known to induce MMP-2 and −9 (19). Thus, inhibition of MMP-2 and −9 is
essential for preventing basement membrane destruction. MMP
proteins were associated with TJ proteins and upregulation of TJ
proteins induced MMP activation in cancer cells. MMP activity leads
to cell invasion and metastasis. In general, normal cells have
tissue inhibitors of metalloproteinases (TIMPs). Therefore MMP
activation and degradation of cell basement membrane are inhibited.
However, cancer cells have constant proliferation and invasion by
upregulation of TJ proteins and do not have TIMPs (20). In our previous study, expression of
TJ proteins was confirmed. Therefore, MMP and TIMP levels were
evaluated in AGS cells following treatment with Cf-GP. MMP-2 and −9
levels were decreased due to increased TIMP-1 as determined by
western blot analysis and RT-PCR (Fig.
5A and B). In addition, gelatin zymography assays indicated
that MMP protein levels decreased with increasing Cf-GP
concentrations (Fig. 5C).
Discussion
Vividiffusion, division and gene expression are
maintained by homeostasis in normal cells. However, in cancer
cells, homeostasis is not maintained, enabling mutations to develop
and invasion to continue. Therefore, cancer cell metastasis and
invasion must be inhibited. MMP and TJ proteins are important for
cancer cell invasion and metastasis, and are closely associated
with cell membrane permeability (21). Invasion and metastasis of cancer
cells occurs through degradation of the cell basement membrane by
upregulation of MMP proteins (22). Cancer cell adhesion is affected by
overexpression of proteins associated with cell membrane
permeability. Increased MMP expression correlates with cancer cell
metastasis and invasion (23,24).
In addition, expression of adhesion proteins is important for loss
of invasion and metastatic capacities. TJ proteins are key for the
passage through epithelial and endothelial barriers as well as in
osmoregulation and maintenance of cell polarity (25–27).
Therefore, cancer cell membrane permeability is weakened due to
overexpression of TJ and MMP proteins, resulting in enhanced
metastasis and invasion. Expression levels of the TJ proteins,
claudin, zo-1 and occludin, are increased, resulting in the
inhibition of invasion and metastasis by cancer cells (28,29).
Changes in cell membrane permeability are induced by increased
expression of claudin, zo-1 and occludin. In addition, TEER values
are decreased by overexpression of TJ proteins (30). Therefore, suppression of claudin,
zo-1 and occludin is important for inhibition of metastasis and
invasion. In addition, increased expression of β-catenin
facilitates invasion and metastasis of cancer cells, which is
mediated by the simultaneous actions of MMP and TJ proteins. Thus,
expression of β-catenin and E-cadherin is important (31,32).
In normal cells, E-cadherin inhibits β-catenin, which is associated
with cell adhesion, inhibiting its nuclear accumulation. Since
E-cadherin expression is decreased in cancer cells, β-catenin
accumulates in the nucleus, resulting in enhanced adhesion. Cell
membrane permeability is impeded by increased MMP and TJ
expression, while adhesion is enhanced, resulting in the rapid
proliferation of cancer cells. Previous studies using Cf-GP
revealed that apoptosis of AGS cells was associated with Fas
signaling (17) and that β-catenin
expression was reduced during apoptosis. Therefore, the effect of
Cf-GP on AGS human gastric cancer cell invasion and metastasis was
investigated.
Cf-GP treatment (0, 5, 10 or 20 μg/ml) for 24 h
inhibited the growth of AGS cells. The Cf-GP-treated group
exhibited decreased cell invasion and increased TEER. The latter is
based on the electrical resistance of the cell membrane and is
closely associated with cell invasion due to its association with
cell membrane permeability. Next, the expression levels of
associated proteins, MMPs, TIMP-1, claudin, zo-1, occludin,
β-catenin and E-cadherin, were evaluated. Dose-dependent decreases
in MMP expression and increases in TIMP-1 expression, an MMP
inhibitor, were identified following Cf-GP treatment for 24 h at
the protein and mRNA levels. In addition, MMP-2 and −9 levels were
observed by gelatin zymography, confirming inhibition at the
protein level. As a result, compared with the control group,
Cf-GF-treated cells exhibited significantly decreased claudin, zo-1
and occludin protein and mRNA levels. This result confirmed that
suppression of cancer cell invasion is mediated by inhibition of TJ
protein expression.
In summary, Cf-GP decreased MMP expression in a
dose-dependent manner. MMP expression is associated with cell
basement membrane degradation factor and inhibition of the TJ
proteins involved in membrane permeability. Inhibition of AGS
growth and invasion was confirmed. Anti-tumor activity in various
cell lines using marine algae has been previously reported and
results of the present study indicate the promise of functional
therapeutics using Cf-GP.
Acknowledgements
This study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
funded by the Ministry of Education, Science and Technology (no.
2012R1A6A1028677).
References
|
1
|
Bishop JM: The molecular genetics of
cancer. Science. 235:305–311. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim SO, Choi YH and Choe WK:
Indol-3-carbinol regulated tight junction permeability and
associated-protein level and suppressed cell invasion in human
colon cancer cell line, HT-29. Korean J Nutr. 41:13–21. 2008.
|
|
3
|
Higashi-Okaj K, Otani S and Okai Y: Potent
suppressive effect of a Japanese edible seaweed, Enteromorpha
prolifera (Sujiao-nori) on initiation and promotion phases of
chemically induced mouse skin tumorigenesis. Cancer Lett.
140:21–25. 1999.PubMed/NCBI
|
|
4
|
Okai Y, Higashi-Okai K, Nakamura S, Yano Y
and Otani S: Suppressive effects of the extracts of Japanese edible
seaweeds on mutagen-induced umu C gene expression in Salmonella
typhimurium (TA 1535/pSK 1002) and tumor promoter-dependent
ornithine decarboxylase induction in BALB/c 3T3 fibroblast cells.
Cancer Lett. 87:25–32. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamamoto I, Maruyama H and Moriguchi M:
The effect of dietary seaweeds on
7,12-dimethylbenz[a]anthracene-induced mammary tumorigenesis in
rats. Cancer Lett. 35:109–118. 1987.
|
|
6
|
Noda H, Amano H, Arashima K and Nisizawa
K: Antitumor activity of marine algae. Hydrobiologia. 204–205.
577–584. 1990.
|
|
7
|
Kwon MJ and Nam TJ: Porphyran induces
apoptosis related signal pathway in AGS gastric cancer cell lines.
Life Sci. 79:1956–1962. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Go H, Hwang HJ and Nam TJ: A glycoprotein
from Laminaria japonica induces apoptosis in HT-29 colon
cancer cells. Toxicol in Vitro. 24:1546–1553. 2010.
|
|
9
|
Hwang HJ, Kim IH and Nam TJ: Effect of a
glycoprotein from Hizikia fusiformis on
acetaminophen-induced liver injury. Food Chem Toxicol.
46:3475–3481. 2008.PubMed/NCBI
|
|
10
|
Choi YH: Inhibition of cell invasion by
ethyl alcohol extracts of Hizikia fusiforme in AGS human
gastric adenocarcinoma cells. Korea J Life Sci. 20:1784–1791. 2010.
View Article : Google Scholar
|
|
11
|
Cho EK, Yoo SK and Choi YJ: Inhibitory
effects of maesaengi (Capsosiphon fulvescens) extracts on
angiotensin converting enzyme and α-glucosidase. Korea J Life Sci.
21:811–818. 2011.
|
|
12
|
Park HY, Lim CW, Kim YK, Yoon HD and Lee
KJ: Immunostimulating and anti-cancer activities of hot water
extract from Capsosiphon fulvescens. J Korean Soc Appl Biol
Chem. 49:343–348. 2006.
|
|
13
|
Kim YM, Kim IH and Nam TJ: Induction of
apoptosis signaling by a glycoprotein of Capsosiphon
fulvescens in AGS cell. Kor J Fish Aquat Sci. 44:216–224.
2011.
|
|
14
|
Peralta SA, Mullin JM, Knudsen KA and
Marano CW: Tissue remodeling during tumor necrosis factor-induced
apoptosis in LLC-PK1 renal epithelial cells. Am J Physiol.
270:F869–F879. 1996.PubMed/NCBI
|
|
15
|
Swift JG, Mukherjee TM and Rowland R:
Intercellular junctions in hepatocellular carcinoma. J Submicrosc
Cytol. 15:799–810. 1983.PubMed/NCBI
|
|
16
|
Van Itallie CM and Anderson JM: The
molecular physiology of tight junction pores. Physiology
(Bethesda). 19:331–338. 2004.PubMed/NCBI
|
|
17
|
Kim YM, Kim IH and Nam TJ: Induction of
apoptosis signaling by glycoprotein of Capsosiphon
fulvescens in human gastric cancer (AGS) cells. Nutr cancer.
64:761–769. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lamszus K, Kunkel P and Westphal M:
Invasion as limitation to anti-angiogenic glioma therapy. Acta
Neurochir Suppl. 88:169–177. 2003.PubMed/NCBI
|
|
19
|
Duffy MJ, Maguire TM, Hill A, McDermott E
and O'Higgins N: Metalloproteinases: role in breast carcinogenesis,
invasion and metastasis. Breast Cancer Res. 2:252–257. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Curry JD, Glaser MC and Smith MT:
Real-time reverse transcription polymerase chain reaction detection
and quantification of t(1;19)(E2A-PBX1) fusion genes associated
with leukemia. Br J Haematol. 115:826–830. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park HS, Kim GY, Choi IW, Kim ND, Hwang
HJ, Choi YW and Choi YH: Inhibition of matrix metalloproteinase
activities and tightening of tight junctions by diallyl disulfide
in AGS human gastric carcinoma cells. J Food Sci. 76:T105–T111.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Coussens LM and Werb Z: Matrix
metalloproteinases and the development of cancer. Chem Biol.
3:895–904. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bennett JH, Morgan MJ, Whawell SA, Atkin
P, Roblin P, Furness J and Speight PM: Metalloproteinase expression
in normal and malignant oral keratinocytes: stimulation of MMP-2
and MMP-9 by scatter factor. Eur J Oral Sci. 108:281–291. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ylisirniö S, Höyhtyä M and
Turpeenniemi-Hujanen T: Serum matrix metalloproteinases −2, −9 and
tissue inhibitors of metalloproteinases −1, −2 in lung
cancer-TIMP-1 as a prognostic marker. Anticancer Res. 20:1311–1316.
2000.
|
|
25
|
Song ES, Lee BI, Kim JM, Lee KY, An KS,
Sung SM, Kwon HJ, Park JH, Han JY and Choi SJ: Increase expressions
of claudin-1 and claudin-7 in cervical squamous intraepithelial
neoplasias and invasive squamous cell carcinomas. Korean J Obstet
Gynecol. 49:1065–1072. 2006.
|
|
26
|
Langbein L, Grund C, Kuhn C, Praetzel S,
Kartenbeck J, Brandner JM, Moll I and Franke WW: Tight junctions
and compositionally related junctional structures in mammalian
stratified epithelia and cell cultures derived therefrom. Eur J
Cell Biol. 81:419–435. 2002. View Article : Google Scholar
|
|
27
|
Tsukita S and Furuse M: Pores in the wall:
claudins constitute tight junction strands containing aqueous
pores. J Cell Biol. 149:13–16. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Agarwal R, D'Souza T and Morin PJ:
Claudin-3 and claudin-4 expression in ovarian epithelial cells
enhances invasion and is associated with increased matrix
metalloproteinase-2 activity. Cancer Res. 65:7378–7385. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Balda MS, Garrett MD and Matter K: The
ZO-1-associated Y-box factor ZONAB regulates epithelial cell
proliferation and cell density. J Cell Biol. 160:423–432. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee H, Kim D, Sohn D, Jeong B, Choi H, Sim
K, Lee K, Cho H, Kim S, Lee J, Jeong Y, Kim S, Lee W and Kim K: The
changes of occludin in tight junction of blood-brain barrier by
ROS. Korean J Electron Microscopy. 34:231–239. 2004.
|
|
31
|
Tunggal JA, Helfrich I, Schmitz A, Schwarz
H, Günzel D, Fromm M, Kemler R, Krieg T and Niessen CM: E-cadherin
is essential for in vivo epidermal barrier function by regulating
tight junctions. EMBO J. 24:1146–1156. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mess ST, Mennigen R, Spieker T, Rijcken E,
Senninger N, Haier J and Bruewer M: Expression of tight and
adherens junction proteins in ulcerative colitis associated
colorectal carcinoma: upregulation of claudin-1, claudin-3,
claudin-4 and β-catenin. Int J Colorectal Dis. 24:361–368.
2009.PubMed/NCBI
|