Introduction
Human inflammatory bowel disease (IBD), a type of
serious gastrointestinal tract disease that includes ulcerative
colitis (UC) and Crohn’s disease (CD), is characterized by
recurrent chronic inflammation and mucosal tissue damage of the
gastrointestinal tract (1).
However, the etiology of IBD remains unknown. Generally, IBDs are
the result of various factors including environmental, genetic and
lifestyle effects, as well as immunological disorders (1,2). In
particular, immunological disorders have resulted in an imbalance
between pro-inflammatory cytokines, such as tumor necrosis factor-α
(TNF-α), interferon-γ (IFN-γ), interleukin (IL)-1β, -6 and -12, and
anti-inflammatory cytokines such as IL-4 and -10. Therefore,
cytokines may be important in the pathological process of IBD
(3). The inflammatory response
begins with an infiltration of neutrophils and macrophages.
Activated macrophages produce a potent mixture of broadly active
inflammatory cytokines, such as TNF-α, IL-1β and -6 which
subsequently causes damage to the colonic tissues (4,5).
Downregulation of the aberrant immune response and inhibition of
the pro-inflammatory cytokines that induce inflammatory cascades,
has been recognized as a major therapeutic target in IBD treatment
(6).
Traditional therapeutic agents, including
5-aminosalicylaes (5-ASA) and corticosteroids, which are used in
clinical IBD treatment, are also associated with serious
complications and undesirable side effects (7). 5-ASA is well tolerated but diarrhea,
cramps and abdominal pain are occasional side effects and these may
be accompanied by fever, rash or kidney problems. Corticosteroids
also result in systemic immunosuppression and have several
well-known side effects, such as rounding of the face, acne,
increased body hair, diabetes, weight gain and high blood pressure
(8). For these reasons, the
development of effective and safe therapeutic agents to treat IBD,
has become increasingly important.
Kudingcha is a bitter tea that is prepared from
Ilex kudingcha C.J. Tseng. It has been consumed
traditionally as a herbal tea in China, Vietnam and various regions
of Southwest Asia (9). Kudingcha
has been demonstrated to exhibit numerous beneficial functions,
including antioxidant, -obesity, -diabetic, -inflammatory,
cardiovascular, hepatoprotective and neuroprotective activities
(10–18). Previous studies have identified
that kudingcha is rich in polyphenolic compounds, such as
caffeoylquinic acid (CQA) and its derivatives (11,12,19).
In the present study, we investigated the anti-inflammatory effects
of Ilex kudingcha C.J. Tseng methanol extract using an in
vivo animal model of IBD, induced by DSS.
Materials and methods
Chemicals and reagents
DSS (molecular weight: 36,00–50,000) was obtained
from MP Biomedicals (Solon, OH, USA). TRIzol reagent,
OligodT18 primer, murine maloney leukemia virus (MMLV)
reverse transcriptase, RNase inhibitor, ethidium bromide (EtBr) and
agarose, were purchased from Invitrogen Life Technologies
(Carlsbad, CA, USA). All other reagents were of analytical
grade.
Plant extract preparation
Fresh kudingcha (Ilex kudingcha C.J. Tseng.)
leaves were purchased from a local market in Chongqing, China in
October, 2012. The fresh kudingcha leaves were freeze-dried and
then ground into a fine powder. A 12-fold volume of methanol (80%,
vol/vol) was added to the powdered samples and extracted three
times by stirring overnight. Kudingcha methanol extracts (KME) were
concentrated by heat evaporation, cryodessication and stored at 4°C
until further study.
Animal studies
Male mice (C57BL/6J strain; age, 6 weeks) were
purchased from the Experimental Animal Center of Chongqing Medical
University (Chongqing, China). The mice were housed in a standard
12-h light/dark cycle at room temperature, and had ad
libitum access to food and water. Colitis was induced in mice
by administration of 3% (wt/vol) DSS in the drinking water for 7
days. Mice were randomly divided into four groups with 6 mice per
group: Group 1, the normal controls were treated with 0.9% normal
saline; group 2, DSS-treated mice and groups 3 and 4 received DSS
and were administered with KME (50 and 200 mg/kg) daily via an
intragastric route (0.2 ml/mouse) for 7 days, until sacrifice. The
animal protocol used in this study was reviewed by the Animal
Ethics Committee of Chongqing Medical University.
Evaluation of disease activity index
(DAI)
The DAI was used to evaluate the grade and extent of
intestinal inflammation. Body weight, stool consistency and blood
in the stools were monitored daily for determination of DAI. Each
score was provided as follows: Body weight loss (0, none; 1, 1–5%;
2, 5–10%; 3, 10–20%; 4, >20%), diarrhea (0, normal; 2, loose
stools; 4, watery diarrhea) and blood (0, normal; 2, slight
bleeding; 4, gross bleeding). The DAI score ranged from 0 to 12
(total score) (20). The mice were
sacrificed on day 7, and the weight and length of the colon were
measured.
Histological observations
The distal colons from each animal were subjected to
histological examination. The colon tissues were fixed in 10%
(vol/vol) neutral-buffered formalin, dehydrated in ethanol and
embedded in paraffin. Colon tissue sections (4 μm) were then cut
and stained with hematoxylin and eosin (H&E).
Myeloperoxidase (MPO) activity
MPO activity was assessed as described previously
(21), but with modifications.
Colon tissues (50 mg) were washed, homogenized in cooled
phosphate-buffered saline (PBS, 80 mM, pH 5.4) containing 0.5%
hexadecyltrimethylammonium bromide (HTAB) and centrifuged at 12,000
× g, for 20 min at 4°C. The supernatant was added to a mixture of
150 μl 3,3′,5,5′-tetramethylbenzidine (2 mM), 50 μl
H2O2 (300 mM), 250 μl PBS (pH 5.4, 80 mM) and
incubated for 30 min at 25°C. The reaction was quenched by 2.5 ml
H2SO4 (200 mM) and the absorbance of the
resulting mixture was measured at 450 nm with a UV-2401PC
spectrophotometer (Shimadzu Corporation, Kyoto, Japan).
Lipid peroxidation levels
Lipid peroxidation was measured by the
thiobarbituric acid (TBA)-reactive substance (TBARS) assays for
malondialdehyde (MDA) following a previously described method
(22), but with modifications.
Colon tissue (100 mg) was washed and homogenized in cooled PBS.
Total protein was determined with a bicinchoninic acid (BCA) assay.
The suspension was mixed with 1 ml TBA (0.67%, w/v) and 1 ml
trichloroacetic acid (TCA; 25%, w/v), heated for 45 min at 95°C and
centrifuged at 12,000 × g for 20 min at 4°C. TBA reacted with the
oxidative degradation products of lipids, yielding red complexes
that are absorbed at 535 nm. The volume of MDA was determined using
a spectrophotometer (UV-2401PC).
Glutathione (GSH) levels
GSH levels were assessed as described previously by
Ellman (23). Colon tissue (100
mg) was washed and homogenized in cooled PBS. The homogenate (0.5
ml) was well mixed with 10% TCA (0.5 ml) and centrifuged at 3,000 ×
g for 5 min. An aliquot of supernatant (0.1 ml) was mixed with 1.7
ml of potassium phosphate buffer (0.1 M, pH 8.0) and 0.1 ml of
Ellman’s reagent. After 5 min, the optical density was measured at
412 nm against a blank using a spectrophotometer (UV-2401PC).
Measurement of colonic pro-inflammation
cytokine levels
Colon samples were washed and homogenized for 5 min
in 3 ml PBS (0.1 M, pH 7.4) at 4°C. The tissue homogenates were
centrifuged at 12,000 × g for 5 min at 4°C. Colonic levels of
TNF-α, IL-1β and -6 were measured with a commercial enzyme-linked
immunosorbent assay kit (ELISA MAX, BioLegend Inc., San Diego, CA,
USA) according to the manufacturer’s instructions.
RT-PCR assay
mRNA expression of TNF-α, IL-1β, -6, inducible
nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) in the
colon tissue, was measured with RT-PCR. Total RNA was isolated with
TRIzol reagent and centrifuged at 12,000 × g, for 15 min at 25°C,
following the addition of chloroform. Isopropanol was added to the
supernatant at a 1:1 ratio and the RNA was pelleted by
centrifugation at 12,000 × g for 15 min. After washing with
ethanol, the RNA was solubilized in diethyl pyrocarbonate-treated
RNase-free water and quantified by measuring the absorbance at 260
nm using a spectrophotometer (UV-2401PC). Equal amounts of RNA (1
μg) were reverse transcribed in a master mix containing 1X reverse
transcriptase buffer, 1 mM dNTPs, 500 ng of oligodT18
primers, 140 units of MMLV reverse transcriptase and 40 units of
RNase inhibitor for 45 min at 42°C. PCR was then carried out in an
automatic thermocycler (Bioneer, Daejeon, South Korea) for 25
cycles (94°C for 30 sec, 55°C for 30 sec and 72°C for 40 sec)
followed by an 8 min extension at 72°C. The PCR products were
separated in 2% agarose gels and visualized by EtBr staining.
β-actin was used for normalization.
Statistical analysis
Data were presented as the mean ± standard
deviation. Differences between the mean values for individual
groups were assessed by a one-way analysis of variance with
Duncan’s multiple range tests. P<0.05 was considered to indicate
a statistically significant difference. The SAS v9.1 statistical
software package (SAS Institute Inc., Cary, NC, USA) was used for
analysis.
Results
KME attenuated DSS-induced colitis
symptoms
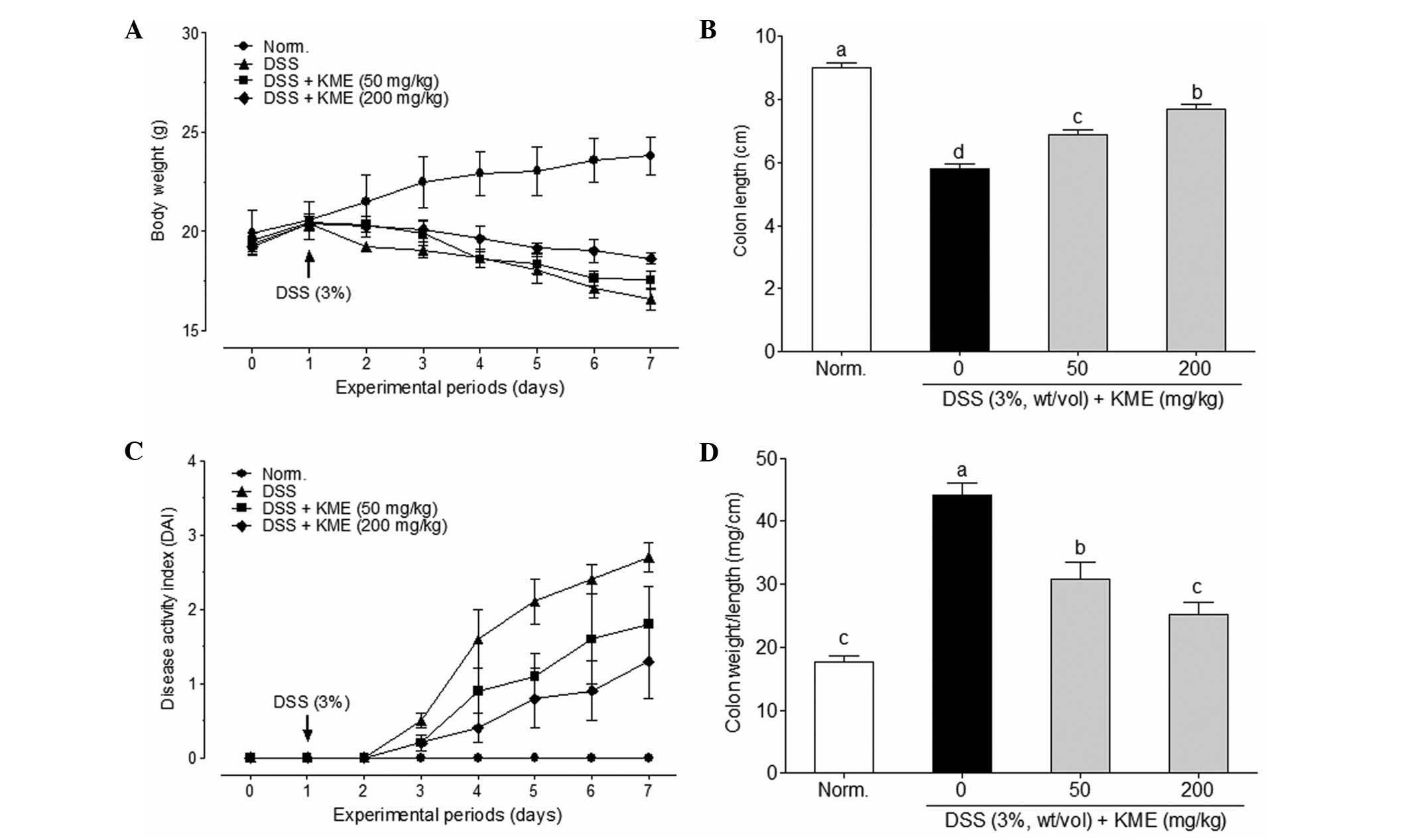
No animals died during the experimental period. As
shown in Fig. 1A, the body weight
gain of mice in the DSS group was significantly lower than that in
the normal control group. KME-treated groups prevented the
DSS-induced body weight loss. The symptoms of DSS-induced colitis
in mice were similar to those observed in humans, such as body
weight loss, diarrhea and gross bleeding (24). We quantitatively scored these
symptoms according to the DAI. The DAI score indicated that KME
alleviated the severity of DSS-induced colitis (Fig. 1C). DSS-induced colitis is
associated with a marked decrease in colon length (25). As shown in Fig. 1B, DSS significantly decreased the
colon length in the DSS treatment group (5.7±0.5 cm), compared with
that in the normal control group (9.0±0.4 cm). However, KME was
able to reduce the DSS-induced colon shortening in mice with
colitis. In addition, colon weight to length ratio was used as an
indicator of disease-associated intestinal wall thickening and
intensity of inflammation. KME significantly inhibited DSS-induced
intestinal wall thickening compared with that of the normal control
group (Fig. 1D).
Effects of KME on histological changes
and MPO activity in DSS-induced colitis mice
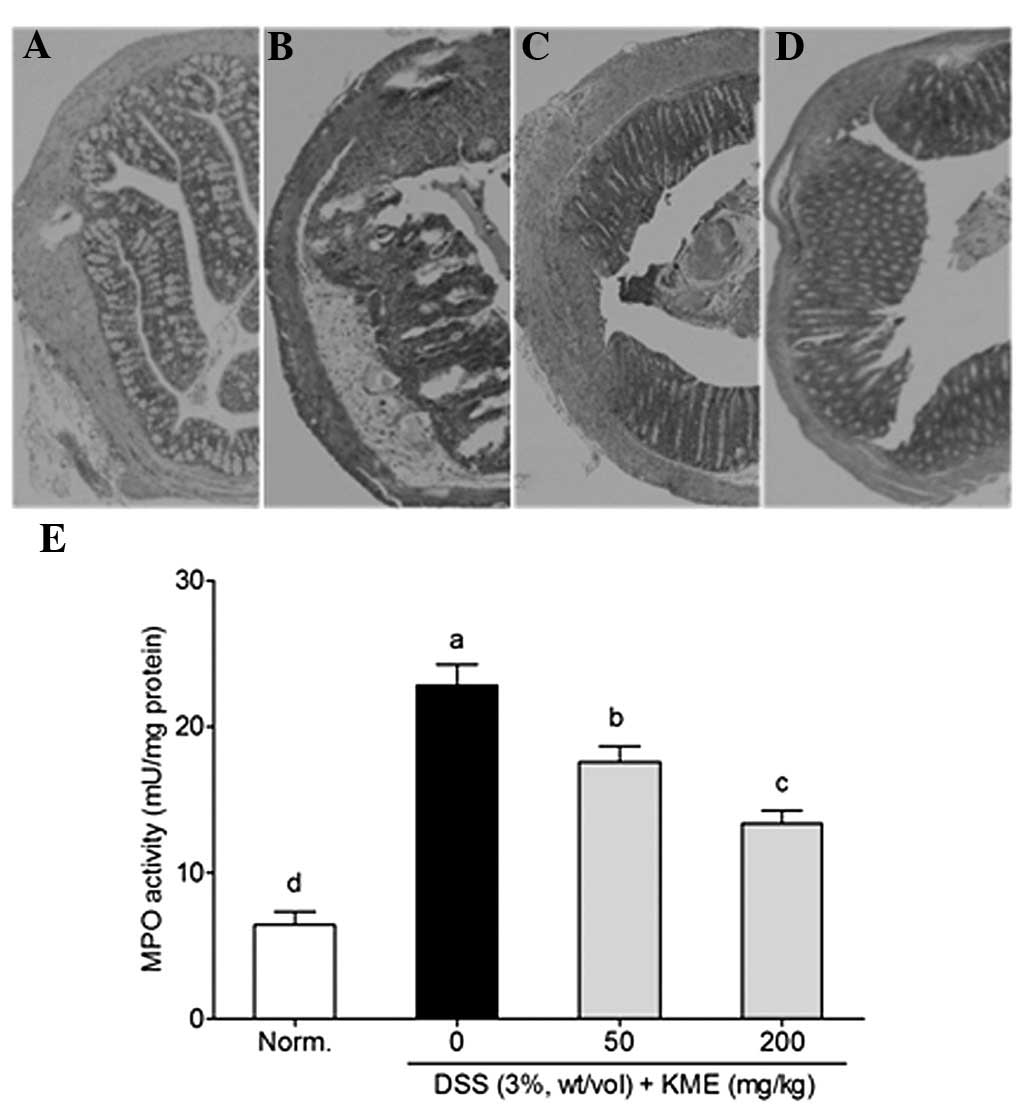
The H&E staining assay was used to evaluate the
therapeutic effects of KME in DSS-induced colonic inflammation and
mucosal injury in colitis mice. As is evident in Fig. 2A, the tissue sections from normal
mice showed intact surface epithelium, cryptal gland, stroma and
submucosa, while the tissue sections from the DSS-induced colitis
mice showed distorted crypt epithelium and extensive mucosal damage
with a large number of inflammatory cells (Fig. 2B). However, tissue sections from
KME-treated DSS-colitis mice had more intact surface epithelium,
crypt glands and less inflammatory reactions than those in the
DSS-colitis mice (Fig. 2C and
D).
MPO activity, which is an indicator of acute
inflammation, reflects the volume of neutrophil infiltration
(26). DSS significantly increased
the colonic MPO activity in colitis mice. However, following a
treatment of 50 and 200 mg/kg KME significantly suppressed MPO
accumulation in the colonic tissues of DSS-induced colitis mice
(Fig. 2E).
Effects of KME on GSH and MDA levels in
DSS-induced colitis mice
DSS significantly increased the colonic MDA levels
(to 0.98±0.10 nmol/mg protein) compared with that of the normal
control group (0.47±0.02 nmol/mg protein) (Table I). KME significantly reduced the
colonic MDA levels of 0.79±0.07 and 0.63±0.07 nmol/mg protein at 50
and 200 mg/kg KME, respectively. In addition, KME also attenuated
the DSS-induced reduction in colonic GSH levels in the colitis
mice. The colonic GSH levels of the DSS-colitis mice significantly
increased following treatment with KME, with the increased levels
ranging from 4.58±0.44 to 5.84±0.49 μmol/mg protein at 50 and 200
mg/kg KME, respectively.
 | Table IEffects of kudingcha methanol extract
on the levels of malondialdehyde and glutathione in the colon
tissue of dextran sulfate sodium-treated mice. |
Table I
Effects of kudingcha methanol extract
on the levels of malondialdehyde and glutathione in the colon
tissue of dextran sulfate sodium-treated mice.
| Groups | GSH (μmol/mg
protein) | MDA (nmol/mg
protein) |
|---|
| Normal control | 8.70±0.40a | 0.47±0.02d |
| DSS | 2.62±0.34d | 0.98±0.10a |
| DSS + KME (50
mg/kg) | 4.58±0.44c | 0.79±0.07b |
| DSS + KME (200
mg/kg) | 5.84±0.49b | 0.63±0.07c |
Effects of KME on the colonic and mRNA
levels of pro-inflammatory cytokines in DSS-induced colitis
mice
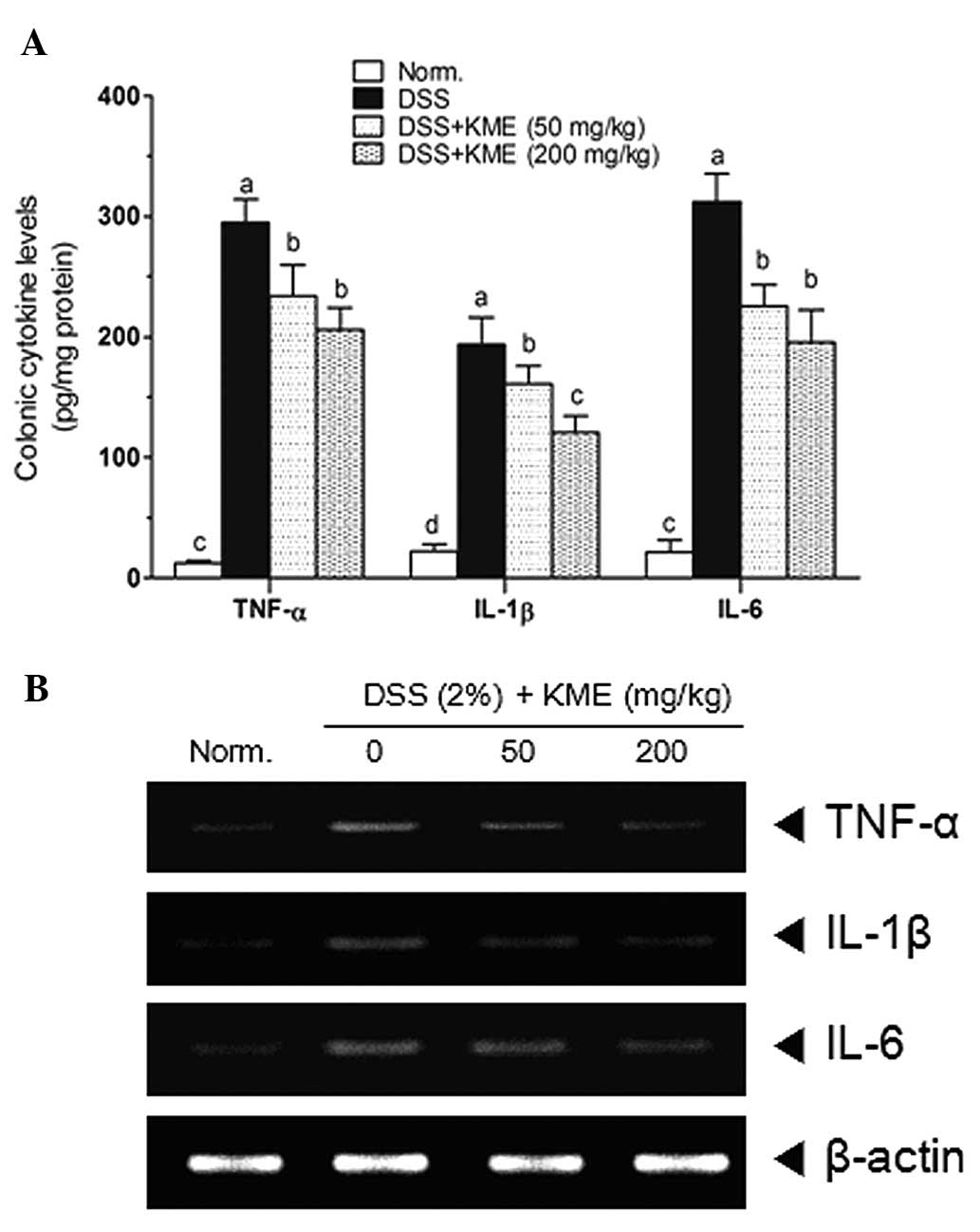
Increased pro-inflammatory cytokine levels are
associated with the UC pathological process. As shown in Fig. 3A, DSS significantly increased the
colonic levels of TNF-α, IL-1β and -6. KME significantly reduced
the levels of TNF-α, IL-1β and -6 compared with that in the
DSS-induced mice with colitis. To investigate the anti-inflammatory
effects of KME on DSS-induced colitis in mice, mRNA expression of
TNF-α, IL-1β and -6 in colonic tissue was analyzed by RT-PCR. As
shown in Fig. 3B, colonic
inflammation induced by DSS. resulted in an elevated expression of
all pro-inflammatory cytokines. Our findings demonstrate that
administration of KME effectively reduced the mRNA expression of
TNF-α, IL-1β and -6 in the colon tissue of mice with DSS-induced
colitis.
KME inhibited the iNOS and COX-2 gene
expression in DSS-induced colitis mice
iNOS and COX-2 are two types of inflammation-related
enzymes and are important in the pathological process of UC.
Therefore, we evaluated the effects of KME on iNOS and COX-2 mRNA
expression in the colonic tissue of DSS-colitis mice compared with
that of the untreated DSS-colitis mice. As shown in Fig. 4, DSS significantly increased the
mRNA levels of iNOS and COX-2 in the colonic tissue of DSS-induced
colitis mice. In addition, KME significantly and dose-dependently
reduced the iNOS and COX-2 mRNA levels.
Discussion
In western countries, the incidence and prevalence
of IBD has increased in the past 50 years from 8–14/100,000 to
120–200/100,000 in individuals with UC (27). Recently, the prevalence of UC has
been on the increase in the Asian-Pacific region (28). In general, therapeutic treatment
for UC includes anti-inflammation and immunosuppression. However,
these treatments have also been associated with undesirable side
effects. Therefore, natural medicine has become an alternative
therapy in addition to the conventional therapies that are used to
treat UC (29). In the present
study, we investigated the anti-inflammatory activity of kudingcha
methanol extracts (KME) using DSS-induced mice colitis model. KME
(50 and 200 mg/kg) and DSS (3%) in mice were orally coadministered
and then clinical colitis was assessed by examining body weight
loss, shortening of the colon length, increasing colon weight to
length ratio and the DAI.
KME administration attenuated body weight loss,
colonic shortening and intestinal wall thickening that was induced
by DSS (Fig. 1). DSS significantly
induced inflammatory cell infiltration, mucosal erosion, distortion
and loss of crypts (Fig. 2B), as
well as elevated MPO accumulation in the colon tissue of
DSS-colitis mice (Fig. 2E). MPO, a
member of the hemeperoxidase-cyclooxygenase superfamily, is
abundantly expressed in neutrophils, and to a lesser extent in
monocytes and certain types of macrophages (31). It is a specific marker that may be
used to determine neutrophil influx into the colon tissue. The
decrease in MPO activity may be explained through the reduction of
neutrophil accumulation in inflamed tissues (32). In the present study, KME markedly
reduced leukocyte (neutrophil and macrophage) infiltration and also
decreased the colonic MPO levels to ameliorate the inflammatory
conditions in the colonic tissues of DSS-induced colitis mice.
A typical development of DSS-induced colitis is
leukocyte infiltration into the colonic tissues. Leukocyte
infiltration is an important source of reactive oxygen species
(ROS) and reactive nitrogen species (RNS), which act as cytotoxic
agents by cross-linking proteins, lipids and nucleic acids, thus
causing cell damage (33). Excess
ROS markedly disrupts the oxidant/antioxidant balance as shown by
the increased lipid peroxidation and reduction in colonic GSH
content (34,35). Oxidative stress or cellular damage
with its dual of free radicals, generates profound lipid
peroxidation and are the hallmarks of UC (36). As a good in vitro free
radical scavenger (12,19), KME markedly increased the colonic
GSH levels and reduced the generation of MDA to attenuate the
DSS-induced colitis in mice.
It is well known that the increased pro-inflammatory
cytokines (TNF-α, IL-1β and -6) amplify the inflammatory cascade
and result in intestinal tissue damage in patients with UC, as well
as in animal models of DSS-induced colitis (5,37,38).
The downregulation and/or blockade of pro-inflammatory cytokine
activity was useful in the treatment of IBD (39). For example, anti-TNF-α antibody
(Infilximab, a mouse monoclonal antibody) had effective therapeutic
effects on UC in a clinical case (40,41).
In addition, IL-1β and -6 are two key mediators of the progression
of UC. IL-1β is known to stimulate diarrhea and the reduction of
its function subsequently suppressed the infiltration of
inflammatory cells into the intestinal tissue and inhibited
intestinal necrosis in animals with UC (42–44).
Furthermore, anti-IL-1β antibody attenuated not only the symptoms
of DSS-induced colitis, but also IL-6 gene expression (45). In the present study, we observed
that the colonic levels of TNF-α, IL-1β and -6 in the DSS-induced
colitis mice were markedly decreased by KME administration, as
identified by ELISA (Fig. 3A). The
RT-PCR assay also confirmed that the mRNA levels of these
pro-inflammatory cytokines were reduced by KME in the colonic
tissue of DSS-induced colitis mice (Fig. 3B). These results indicate that KME
may exert anti-inflammatory effects on DSS-induced colitis by
reducing the activity of TNF-α, IL-1β and -6.
Previous studies have indicated that iNOS and COX-2
are considered to be vital in the inflammatory process of UC
(5,46,47).
Pro-inflammatory cytokines induced the mRNA expression of iNOS and
increased the generation of nitric oxide (NO), which is an
inflammatory mediator associated with the pathological process of
UC. Suppression of iNOS attenuated UC in human and animal cases
(47,48). Moreover, the inhibition of
excessive COX-2 activity induced the generation of prostaglandins
E2 (PGE2), which may result in the suppression of DSS-induced
colitis in mice (49,50). In the
present study, we demonstrated that KME decreased the mRNA
expression of iNOS and COX-2 in the colonic tissue of DSS-induced
colitis mice (Fig. 4).
In conclusion, results of the present study have
demonstrated the potential anti-inflammatory effects of KME in
DSS-induced colitis mice. These results suggest that KME
administration prevents DSS-induced body weight loss, colonic
shortening and modulates MPO activity, as well as reducing
intestinal wall thickening. In addition, KME administration
increased the colonic GSH levels, decreased colonic lipid
peroxidation, and reduced the production and mRNA levels of TNF-α,
IL-1β and -6. Results of the present study suggest that the
potential mechanism of KME involves suppressing the production of
TNF-α, IL-1β, -6, iNOS and COX-2, and may be considered an
important anti-inflammatory treatment agent against colonic
inflammation.
References
|
1
|
Macdonald TT and Monteleone G: Immunity,
inflammation, and allergy in the gut. Science. 307:1920–1925. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bouma G and Strober W: The immunological
and genetic basis of inflammatory bowel disease. Nat Rev Immunol.
3:521–533. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ardizzone S and Bianchi Porro G: Biologic
therapy for inflammatory bowel disease. Drugs. 65:2253–2286. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hanauer SB: Inflammatory bowel disease:
epidemiology, pathogenesis, and therapeutic opportunities. Inflamm
Bowel Dis. 12:S3–S9. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Podolsky DK: Inflammatory bowel disease. N
Engl J Med. 347:417–429. 2002. View Article : Google Scholar
|
|
6
|
Rogler G and Andus T: Cytokines in
inflammatory bowel disease. World J Surg. 22:382–389. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van Dieren JM, Kuipers EJ, Samsom JN,
Nieuwenhuis EE and van der Woude CJ: Revisiting the
immunomodulators tacrolimus, methotrexate, and mycophenolate
mofetil: their mechanisms of action and role in the treatment of
IBD. Inflamm Bowel Dis. 12:311–327. 2006.PubMed/NCBI
|
|
8
|
Xu CT, Meng SY and Pan BR: Drug therapy
for ulcerative colitis. World J Gastroenterol. 10:2311–2317.
2004.PubMed/NCBI
|
|
9
|
Sun Y, Xu W, Zhang W, Hu Q and Zeng X:
Optimizing the extraction of phenolic antioxidants from kudingcha
made from Ilex kudingcha C.J. Tseng by using response
surface methodology. Sep Sci Technol. 78:311–320. 2011.
|
|
10
|
Nishimura K, Fukuda T, Miyase T, Noguchi H
and Chen XM: Activity-guided isolation of triterpenoid acyl CoA
cholesteryl acyl transferase (ACAT) inhibitors from Ilex kudincha.
J Nat Prod. 62:1061–1064. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu F, Cai YZ, Sun M, Ke J, Lu D and Corke
H: Comparison of major phenolic constituents and in vitro
antioxidant activity of diverse kudingcha genotypes from Ilex
kudingcha, Ilex cornuta, and Ligustrum robustum.
J Agric Food Chem. 57:6082–6089. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Thuong PT, Su ND, Ngoc TM, Hung TM, Dang
NH, Thuan ND, Bae K and Oh WK: Antioxidant activity and principles
of Vietnam bitter tea Ilex kudingcha. Food Chem.
113:139–145. 2009. View Article : Google Scholar
|
|
13
|
Chen ZY, Wong IY, Leung MW, He ZD and
Huang Y: Characterization of antioxidants present in bitter tea
(Ligustrum pedunculare). J Agric Food Chem. 50:7530–7535.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wong IY, He ZD, Huang Y and Chen ZY:
Antioxidative activities of phenylethanoid glycosides from
Ligustrum purpurascens. J Agric Food Chem. 49:3113–3119.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
She GM, Wang D, Zeng SF, Yang CR and Zhang
YJ: New phenylethanoid glycosides and sugar esters from
ku-ding-cha, a herbal tea produced from Ligustrum
purpurascens. J Food Sci. 73:C476–C481. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lau KM, He ZD, Dong H, Fung KP and But PP:
Anti-oxidative, anti-inflammatory and hepato-protective effects of
Ligustrum robustum. J Ethnopharmacol. 83:63–71. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pu F, Mishima K, Irie K, Egashira N,
Ishibashi D, Matsumoto Y, Ikeda T, Iwasaki K, Fujii H, Kosuna K and
Fujiwara M: Differential effects of buckwheat and kudingcha extract
on neuronal damage in cultured hippocampal neurons and spatial
memory impairment induced by scopolamine in an eight-arm radial
maze. J Health Sci. 51:636–644. 2005. View Article : Google Scholar
|
|
18
|
Kim JY, Jeong HY, Lee HK, Yoo JK, Bae K
and Seong YH: Protective effect of Ilex latifolia, a major
component of ‘kudingcha’, against transient focal ischemia-induced
neuronal damage in rats. J Ethnopharmacol. 133:558–564. 2011.
|
|
19
|
Liu L, Sun Y, Laura T, Liang X, Ye H and
Zeng X: Determination of polyphenolic content and antioxidant
activity of Kudingcha made from Ilex kudingcha C.J. Tseng.
Food Chem. 112:35–41. 2009. View Article : Google Scholar
|
|
20
|
Azuma YT, Nishiyama K, Matsuo Y, Kuwamura
M, Morioka A, Nakajima H and Takeuchi T: PPARα contributes to
colonic protection in mice with DSS-induced colitis. Int
Immunopharmacol. 10:1261–1267. 2010.
|
|
21
|
Shin VY, Liu ES, Koo MW, Wang JY, Matsui H
and Cho CH: Cigarette smoke extracts delay wound healing in the
stomach: involvement of polyamine synthesis. Exp Biol Med
(Maywood). 227:114–124. 2002.PubMed/NCBI
|
|
22
|
Gan XL, Hei ZQ, Huang HQ, Chen LX, Li SR
and Cai J: Effect of Astragalus membranaceus injection on
the activity of the intestinal mucosal mast cells after hemorrhagic
shock-reperfusion in rats. Chin Med J (Engl). 119:1892–1898.
2006.
|
|
23
|
Ellman GL: Tissue sulfhydryl groups. Arch
Biochem Biophys. 82:70–77. 1959. View Article : Google Scholar
|
|
24
|
Strober W, Fuss IJ and Blumberg RS: The
immunology of mucosal models of inflammation. Annu Rev Immunol.
20:495–549. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hendrickson BA, Gokhale R and Cho JH:
Clinical aspects and pathophysiology of inflammatory bowel disease.
Clin Microbiol Rev. 15:79–94. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamamki K, Kim DH, Ryu N, Kim YP, Shin KH
and Ohuchi K: Effects of naturally occurring isoflavones on
prostaglandin E2 production. Planta Med. 68:97–100. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cosnes J, Gower-Rousseau C, Seksik P and
Cortot A: Epidemiology and natural history of inflammatory bowel
diseases. Gastroenterology. 140:1785–1794. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chung HL, Yue GG, To KF, Su YL, Huang Y
and Ko WH: Effect of Scutellariae Radix extract on experimental
dextran-sulfate sodium-induced colitis in rats. World J
Gastroenterol. 13:5605–5611. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Langmead L, Dawson C, Hawkins C, Banna N,
Loo S and Rampton DS: Antioxidant effects of herbal therapies used
by patients with inflammatory bowel disease: an in vitro study.
Aliment Pharmacol Ther. 16:197–205. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Malle E, Furtmüller P, Sattler W and
Obinger C: Myeloperoxidase: a target for new drug development? Br J
Pharmacol. 152:838–854. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Babbs CF: Oxygen radicals in ulcerative
colitis. Free Radic Biol Med. 13:169–181. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Holma R, Salmenperä P, Riutta A, Virtanen
I, Korpela R and Vapaatalo H: Acute effects of the
cys-leukotriene-1 receptor antagonist, montelukast, on experimental
colitis in rats. Eur J Pharmacol. 429:309–318. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mustafa A, EI-Medany A, Hagar HH and
EI-Medany G: Ginkgo biloba attenuates mucosal damage in a
rat model of ulcerative colitis. Pharmacol Res. 53:324–330. 2006.
View Article : Google Scholar
|
|
34
|
Osman N, Adawi D, Ahrné S, Jeppsson B and
Molin G: Probiotics and blueberry attenuate the severity of dextran
sulfate sodium (DSS)-induced colitis. Dig Dis Sci. 53:2464–2473.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fiocchi C: Inflammatory bowel disease: new
insights into mechanisms of inflammation and increasingly
customized approaches to diagnosis and therapy. Curr Opin
Gastroenterol. 20:309–310. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dieleman LA, Palmen MJ, Akol H, Bloemena
E, Peña AS, Meuwissen SG and Van Rees EP: Chronic experimental
colitis induced by dextran sulphate sodium (DSS) is characterized
by Th1 and Th2 cytokines. Clin Exp Immunol. 114:385–391. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bai A and Peng Z: Biological therapies of
inflammatory bowel disease. Immunotherapy. 2:727–742. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Perrier C and Rutgeerts P: Cytokine
blockade in inflammatory bowel diseases. Immunotherapy.
3:1341–1352. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rutgeerts P, Sandborn WJ, Feagan BG,
Reinisch W, Olson A, Johanns J, Travers S, Rachmilewitz D, Hanauer
SB, Lichtenstein GR, et al: Infliximab for induction and
maintenance therapy for ulcerative colitis. N Engl J Med.
353:2462–2476. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Duca I, Ramírez de la Piscina P, Estrada
S, Calderón R, Spicakova K, Urtasun L, Marra-López C, Zabaleta S,
Bengoa R, Marcaide MA and García-Campos F: Steroid-refractory
ulcerative colitis and associated primary sclerosing cholangitis
treated with infliximab. World J Gastroenterol. 19:590–593. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Siegmund B, Lehr HA, Fantuzzi G and
Dinarello CA: IL-1 beta-converting enzyme (caspase-1) in intestinal
inflammation. Proc Natl Acad Sci USA. 98:13249–13254. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tountas NA, Casini-Raggi V, Yang H, Di
Giovine FS, Vecchi M, Kam L, Melani L, Pizarro TT, Rotter JI and
Cominelli F: Functional and ethnic association of allele 2 of the
interleukin-1 receptor antagonist gene in ulcerative colitis.
Gastroenterology. 117:806–813. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dionne S, D’Agata ID, Hiscott J, Vanounou
T and Seidman EG: Colonic explant production of IL-1 and its
receptor antagonist is imbalanced in inflammatory bowel disease
(IBD). Clin Exp Immunol. 112:435–442. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kwon KH, Murakami A, Hayashi R and
Ohigashi H: Interleukin-1beta targets interleukin-6 in progressing
dextran sulfate sodium-induced experimental colitis. Biochem
Biophys Res Commun. 337:647–654. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Papadakis KA and Targan SR: Role of
cytokines in the pathogenesis of inflammatory bowel disease. Annu
Rev Med. 51:289–298. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Dudhgaonkar S, Tandan SK, Kumar D,
Raviprakash V and Kataria M: Influence of simultaneous inhibition
of cyclooxygenase-2 and inducible nitric oxide synthase in
experimental colitis in rats. Inflammopharmacology. 15:188–195.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cross RK and Wilson KT: Nitric oxide in
inflammatory bowel disease. Inflamm Bowel Dis. 9:179–189. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Martín A, Villegas I and Alarcón de la
Lastra C: The COX-2 inhibitor, rofecoxib, ameliorates dextran
sulphate sodium induced colitis in mice. Inflamm Res. 54:145–151.
2005.PubMed/NCBI
|
|
49
|
Tanaka K, Suemasu S, Ishihara T, Tasaka Y,
Arai Y and Mizushima T: Inhibition of both COX-1 and COX-2 and
resulting decrease in the level of prostaglandins E2 is responsible
for non-steroidal anti-inflammatory drug (NSAID)-dependent
exacerbation of colitis. Eur J Pharmacol. 603:120–132. 2009.
View Article : Google Scholar : PubMed/NCBI
|