Introduction
The circadian gene network sets and regulates the
circadian rhythms in organisms as oscillations with an approximate
24 h cycle (1). At the molecular
level, circadian rhythms are controlled by the interaction between
positive and negative feedback loops consisting of several key
clock regulators (2,3). A model encompassing a feedback system
involving heterodimer transcriptional factors (Clock and Bmal1),
two cryptochromes (Cry1 and Cry2), and three Period (Per1, Per2,
and Per3) regulators has been widely described. Among these clock
genes, hClock is at the core of the circadian gene
family.
Previous studies have revealed that the disruption
of circadian rhythms is one of the endogenous factors that
contributes to the initiation and development of colorectal
carcinoma (CRC). Krugluger et al reported that the
expression of hPer1 mRNA in CRCs was lower than that found in
normal cells (4). Wang et
al detected that the expression of the hPer2 protein in CRC
cells was lower than that in non-cancerous cells (5). Correlations between the decreased
hPer2 level and the patient’s age, histological grade and TNM stage
were also detected. The human Clock (hClock) gene
functions upstream of hPer2 to regulate its expression (5). Therefore, the circadian hClock
gene may play a vital role in the development of CRC.
Hoffman et al observed that the hCLOCK
protein expression level was approximately two times higher in
breast tumor cells compared with that observed in the adjacent
normal tissues and three times higher than that in normal breast
cells (6). Alhopuro et al
reported that ~53% of hClock genes in CRC cells had point
mutations in the DNA sequence (7).
Oshima et al demonstrated that the expression level of
hClock mRNA in CRC cells was significantly higher compared with
that found in normal tissues (8).
CRC is one of the most commonly observed
malignancies and is the leading cause of cancer-associated
mortality worldwide (9). In 2011,
~141,210 new CRC cases and 49,380 mortalities were predicted in the
USA (10). In China, CRC is the
fourth leading cause of cancer mortality in large cities and the
fifth in more rural areas. However, in Shanghai, the incidence of
CRC and its mortality rate ranked second and third, respectively,
for females (11). As surgical
approaches and conventional therapeutics have been incapable of
fully controlling the outcomes of CRC, there is an urgent need to
develop more effective treatments.
The circadian rhythm is interconnected with many
aspects of cellular function, such as cell proliferation, migration
and differentiation, and it also helps regulate the digestive
system (12,13). The molecular mechanism governing
the development of CRC is a multiple factor and multistep process,
yet the molecular changes underlying it remain unexplored. The core
circadian gene, hClock, may participate in the development
of CRC, but the correlation between them has not been reported.
In this study, immunofluorescent histochemistry and
quantitative polymerase chain reaction (qPCR) were used to
characterize the role of hClock in the development of human CRC.
The correlation between the hClock gene and tumor-related
genes Bak, Bax, Bid, HIF-1α,
ARNT, tumor necrosis factor receptor I (TNFR
I), TNFR II and VEGF were also examined. Based on
our data, the expression of hClock in human CRCs and its
possible effects were discussed. Our investigation may provide a
new angle for understanding tumorigenesis and a new target for CRC
treatment development.
Materials and methods
Tissue samples
In total, 30 specimens were obtained from the Tumor
Tissue Bank of the Center of Pathological Research in the
Institutes of Biomedical Sciences at Fudan University (Shanghai,
China). The specimens were obtained from patients who had undergone
a radical CRC resection between January 2007 and September 2008
from the General Surgery Department of Huashan Hospital Affiliated
to Fudan University (Shanghai, China). The specimens were all
obtained between 10:00 am–14:00 pm. The cancerous tissue was
dissected from the cancerous mucosa while the paired non-cancerous
tissue was obtained from the normal section of the colorectal
mucosa of the same patient. All tumor tissues were pathologically
confirmed as CRC and the patients did not receive any presurgical
treatment nor exhibited any metastasis. Following surgical removal,
samples were immediately preserved in liquid nitrogen and stored at
−80°C. Complete medical records including the patient’s age, tumor
site, tumor type, histological grade and Dukes’ stage were also
obtained. This study was approved by the Medical Ethics Committee
of Huashan Hospital, Fudan University, Shanghai, China. Written
informed consent was obtained from the patients.
Immunofluorescent histochemistry
Frozen tissues were cut into 6 μm sections and
stored at −80°C. Sections were fixed at 4°C with 70% ethanol
solution and blocked by 10% normal goat serum solution at 37°C for
60 min. Subsequently, sections were incubated with goat anti-human
hClock antibody (1:50, Santa Cruz Biotechnology Inc., Santa Cruz,
CA, USA) as the primary antibody at 4°C overnight. Following a
brief wash with PBS, secondary donkey anti-goat IgG fluorescent
antibody (1:200; Invitrogen Life Technologies, Carlsbad, CA, USA)
was added for 2 h at room temperature. Following several washes
with PBS, the slides were mounted. Sections which were only
incubated with 10% normal goat serum solution, served as negative
controls. The image analysis software Image-Pro Plus (Media
Cybernetics, Rockville, MD, USA) was used to measure the Integrated
Optical Density (IOD), which is the cumulative OD of the positive
immunofluorescent expression of a selected area and represents the
total protein expression of that area.
RNA extraction and first-strand cDNA
synthesis
Preparation of total RNA from tissue samples was
performed using the TRIzol reagent (Invitrogen Life Technologies).
The quantity of total RNA was determined using UV spectrophotometry
and RNA integrity was assessed using agarose gel electrophoresis.
First-strand cDNA was prepared with oligo-dT primers using a
commercial cDNA synthesis kit (ReverTra Ace qPCR RT kit, Toyobo,
Osaka, Japan). The cDNA was subsequently amplified for 33 cycles
with specific primers for Bak, Bax, Bid, HIF-1α, ARNT, TNFR I, TNFR
II and VEGF. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH)
served as an internal reference (Table
I). Sequence data were analyzed using the basic local alignment
search tool (BLAST) on the National Centre for Biotechnology
Information (NCBI) website (http://www.ncbi.nlm.nih.gov).
 | Table IPrimer pairs used for qPCR. |
Table I
Primer pairs used for qPCR.
| Gene | Primer (5′-3′) | Product length
(bp) |
|---|
| hClock | Forward:
GGCTGAAAGACGACGAGAAC
Reverse: GGTGTTGAGGAAGGGTCTGA | 210 |
| ARNT | Forward:
CAAGCCCCTTGAGAAGTCAG
Reverse: GGGGTAGGAGGGAATGTGTT | 165 |
| Bak | Forward:
GGGTCTATGTTCCCCAGGAT
Reverse: AATGGGAGCAGGGGTAGAGT | 165 |
| Bax | Forward:
TCTGACGGCAACTTCAACTG
Reverse: TTGAGGAGTCTCACCCAACC | 188 |
| Bid | Forward:
GAGGATTGTGGCCTTCTTTG
Reverse: CTCAGCCCAGACTCACATCA | 185 |
| HIF-1α | Forward:
GAAAGCGCAAGTCCTCAAAG
Reverse: TGGGTAGGAGATGGAGATGC | 167 |
| TNFR I | Forward:
GTGCCTACCCCAGATTGAGA
Reverse: TGTCGATTTCCCACAAACAA | 175 |
| TNFR II | Forward:
GAGTTGTGGAAAGCCTCTGC
Reverse: CAGGGGGAGAAACAAACAAA | 193 |
| VEGF | Forward:
AAGGAGGAGGGCAGAATCAT
Reverse: ATCTGCATGGTGATGTTGGA | 226 |
| GAPDH | Forward:
ACAGCCGCATCTTCTTGTGCAGTA
Reverse: GGCCTTGACTGTGCCGTGAATTTT | 226 |
PCR reactions containing SYBR-green were amplified
on a Corbett Real Time PCR machine (Bio-Rad, Hercules, CA, USA).
Following reverse transcription, the cDNA was diluted to 10 ng/μl.
The PCR reaction mixture contained 4 μl of diluted cDNA, 1 μl of 10
μM primers (0.5 μl of upstream primers, 0.5 μl of downstream
primers), 10 μl SYBR Green Realtime PCR Master mix (Bio-Rad) and 5
μl of H2O, into a final volume of 20 μl. The samples
were subjected to 40 cycles of amplification (denaturation at 95°C
for 15 sec, primer annealing for 30 sec and extension at 72°C for
30 sec).
Following each qPCR, melting profiles were performed
in order to rule out the possibilities of nonspecific PCR products
and primer dimers. Data from the qPCR was represented by
fluorescent intensity, ΔRn, and number of cycles.
ΔRn=Rn+ − Rn−, Rn+ refers to the
fluorescent intensity of products at any time, Rn−
refers to the systematic basic fluorescent intensity (the
fluorescent intensity at the 6–15th cycles). The threshold was set
to the intermediate value of logΔRn vs. cycle function.
The CT value stands for the number of cycles at the cross point of
ΔRn vs. threshold. The higher the CT value, the greater the number
of steady cycles, thus the lower the gene expression concentration.
As the amplification efficiencies of the target genes and the
internal control were equal, the relative changes to the target
gene expression in the tumor cells compared with that in normal
colorectal mucosa (ΔCT calibrator value) were calculated using the
equation 2−ΔΔCT, where ΔΔCT = ΔCT(non-cancer)
− ΔCT(cancer). The ΔCT values were determined by
subtracting the average GAPDH CT value from the average target gene
CT value.
Statistical analysis
Data are reported as the means ± SD. Comparison
between the groups was performed using the paired Student’s t-test.
Relative analysis was conducted using Pearson’s χ2 test.
IBM SPSS Statistics 19.0 was used for all data analysis. P<0.01
was considered to indicate a statistically significant
difference.
Results
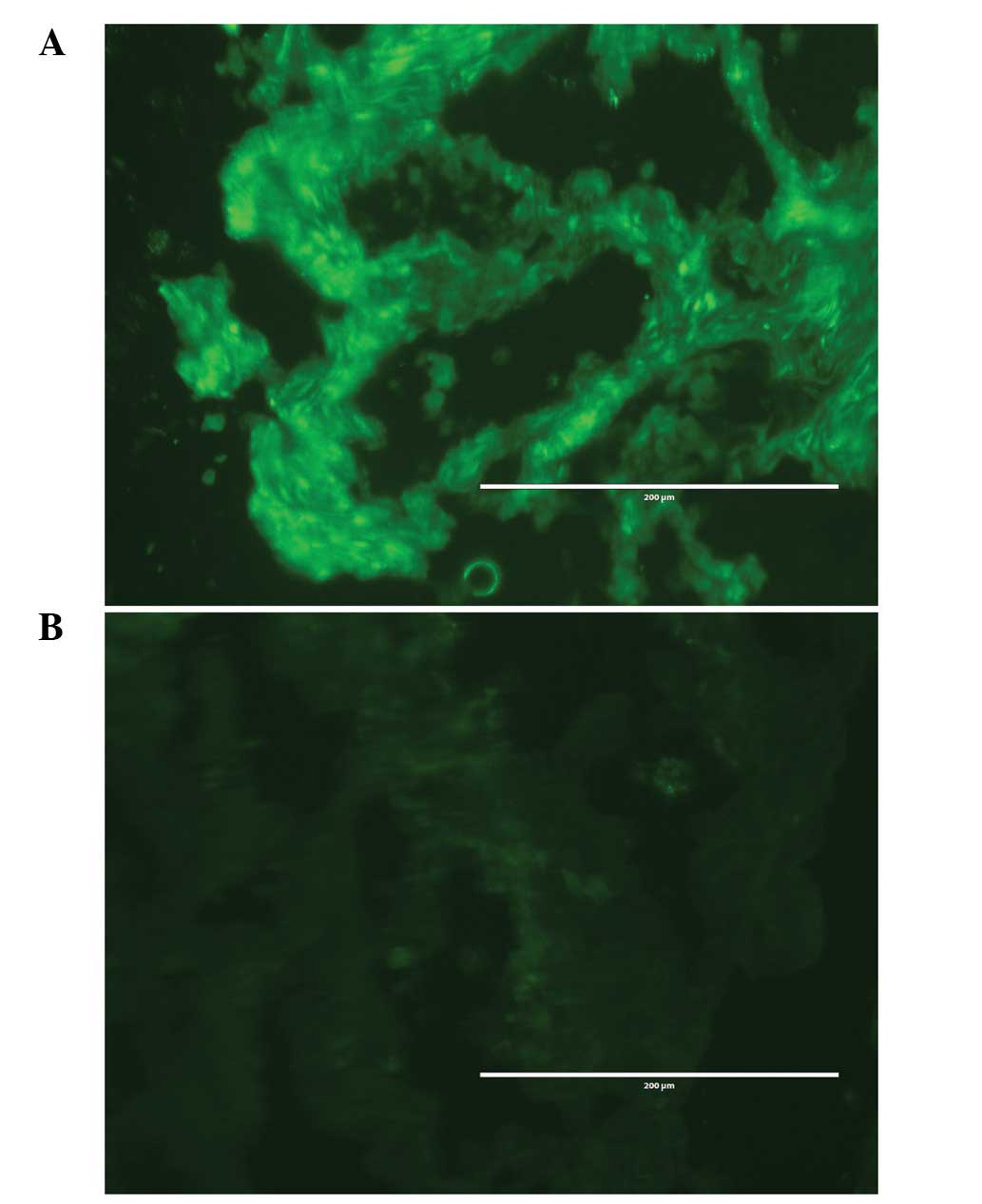
Immunofluorescent histochemical analysis
of hCLOCK protein expression
To analyze expression of the hClock gene in
CRC, we first examined the expression of hCLOCK protein in 30
paired colorectal cancerous and non-cancerous tissues using
immunofluorescent histochemistry. For these 30 cases, hCLOCK
protein expression in colorectal tissues exhibited three typical
patterns. Type I (11 cases): the immunofluorescence was markedly
stronger in the cancerous cells compared with that in the paired
non-cancerous cells, particularly in the nucleolus and cytoplasm.
Type II (17 cases): there were no significant differences in the
immunofluorescence between the cancerous and non-cancerous cells.
Type III (2 cases): the immunofluorescence was stronger in the
non-cancerous cells compared with that in the paired cancerous
cells (Fig. 1).
In these 30 specimens, the average cumulative OD of
the hCLOCK protein in cancerous tissues was 5389.59±2385.20 and
that in paired non-cancerous tissues was 4558.34±1385.87, P<0.05
(P=0.034). These results revealed that the hCLOCK protein was
expressed in all specimens and a higher expression level of hCLOCK
protein was found in human CRCs compared with that in the paired
non-cancerous tissues.
Correlation between hCLOCK protein
expression and clinical-pathological features in CRC patients
In order to analyze the clinical and pathological
relevance of high hCLOCK protein expression in human CRC, we
assessed the correlation between hCLOCK expression and the
respective clinical-pathological parameters in the 30 cases of CRC.
As shown in Table II, an
increased expression of hCLOCK protein was found more in male than
female patients (P=0.039). We observed a higher expression of
hCLOCK in poorly differentiated (P=0.042), or late-stage, Dukes’
grade (P=0.032) tumors. Furthermore, 64.3% of tumors with lymph
node metastasis expressed higher hCLOCK levels. However, no
correlations were found between high levels of hCLOCK expression
and age, tumor site or pathology type.
| Table IICorrelation between increased hCLOCK
protein expression levels in CRC and clinical-pathological
features. |
Table II
Correlation between increased hCLOCK
protein expression levels in CRC and clinical-pathological
features.
|
Clinical-pathological features | Increased hCLOCK
expression in tumor | Non-increased
hCLOCK expression in tumor | P-value |
|---|
| Gender | | | 0.039 |
| Male | 8 | 11 | |
| Female | 3 | 8 | |
| Tumor site | | | >0.05 |
| Colon | 7 | 11 | |
| Rectum | 4 | 8 | |
| Pathology type | | | >0.05 |
| Ulcer | 6 | 11 | |
| Protrude | 5 | 6 | |
| Infiltrating | 0 | 2 | |
| Histological
grade | | | 0.042 |
| I | | 3 | |
| I–II, II | 7 | 13 | |
| II–III, III | 4 | 3 | |
| Dukes’ stage | | | 0.032 |
| 0 | 0 | 1 | |
| A | 0 | 4 | |
| B | 4 | 7 | |
| C | 7 | 7 | |
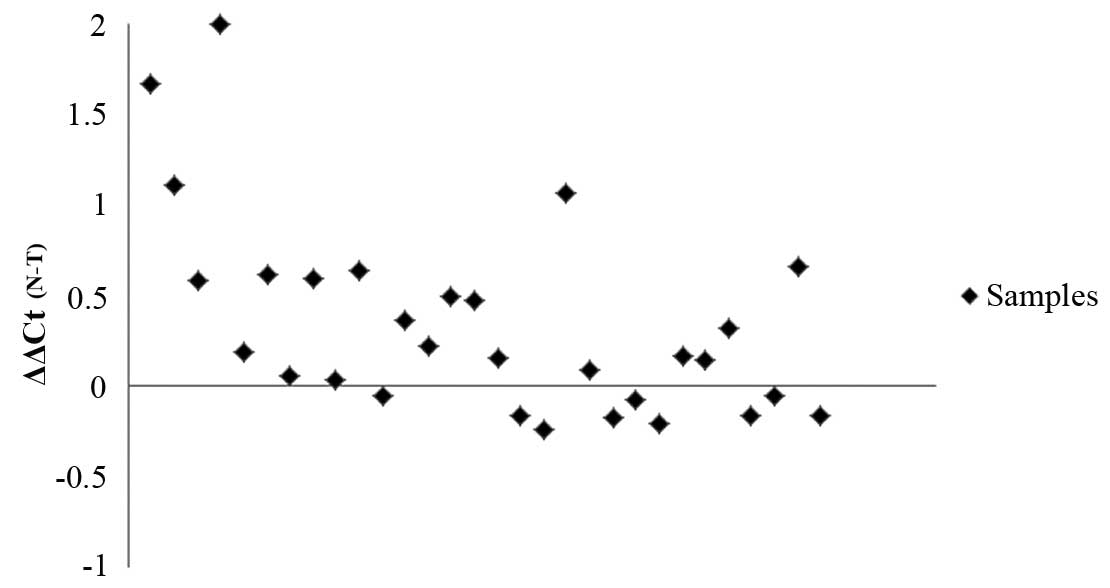
Determination of hCLOCK mRNA level by
qPCR
We further analyzed hClock mRNA levels in the 30
paired cancerous and non-cancerous tissues by qPCR. The quantity of
hClock mRNA was normalized to the endogenous reference gene, GAPDH.
The normalized hClock mRNA expression (ΔCt) of tumorous tissues was
subsequently compared with the ΔCt of the paired non-tumorous
tissues from the same CRC patient in order to determine their
relative expression levels (ΔΔCt) (Fig. 2). The ΔCt of hClock in tumorous
tissues (5.91±1.36) was significantly lower than that found in
paired non-tumorous tissues (6.26±1.47; P=0.002), indicating a
significantly higher level of hClock expression in human
CRCs than in paired non-cancerous tissues.
Amongst the 30 specimens, 12 tumor tissues
demonstrated a high expression of the hClock gene.
Furthermore, in these 12 tumor tissues, 10 specimens also
demonstrated a higher expression of the hCLOCK protein. The
correlation coefficient (r) between hClock mRNA and the hCLOCK
protein was 0.474, P<0.01 (P=0.008). The expression of the
hClock gene and the hCLOCK protein revealed a strong
positive linear correlation in human CRCs.
Correlation between the hClock gene and
tumor-related genes
To further investigate the potential role of hClock
at the molecular level, the correlation between the hClock
gene and a number of tumor-related genes were determined by qPCR.
As shown in Table III, there was
a strong positive linear correlation between the hClock gene
and the ARNT, HIF-1α and VEGF genes in human
CRCs. There was no significant correlation between hClock
with Bak, Bax, Bid, TNFR I and TNFR
II. The higher expression of ARNT, HIF-1α and
VEGF in poorly differentiated, or late-stage, Dukes’ grade
tumors was observed. Furthermore, high VEGF expression was
found in 14 tumor cases with lymph node metastasis.
| Table IIICorrelation between the hClock
gene and other tumor-related genes. |
Table III
Correlation between the hClock
gene and other tumor-related genes.
| Statistical
significance | Bak | Bax | Bid | HIF-1α | ARNT | TNFR I | TNFR II | VEGF |
|---|
| r | 0.114 | 0.005 | −0.272 | 0.543 | 0.608 | 0.370 | −0.280 | 0.714 |
| P-value | 0.548 | 0.980 | 0.415 | 0.046 | 0.027 | 0.068 | 0.083 | 0.008 |
Discussion
It has been demonstrated that the circadian clock
has important roles in orchestrating temporal integrations of
physiology and behavior with the environment. This master clock,
which resides in the hypothalamic suprachiasmatic nucleus in
mammals, synchronizes multiple peripheral oscillators to ensure
temporally coordinated physiology (1,14,15).
Clocks located outside the brain, peripheral clocks, are important
in regulating physiological functions and pathological
situations.
Previous studies have suggested that the disruption
of circadian rhythms, which are regulated by circadian genes, was
one of the endogenous factors that contribute to tumor initiation
and progression. Studies have reported evidence regarding the
beneficial effects of chronotherapy, which refers to the delivery
of chemotherapy according to the circadian rhythms (16,17).
The hClock gene is at the core of the circadian gene family
and directly, or indirectly, regulates certain clock-controlled
genes (18,19). The role of hClock in human
CRCs is unclear. In this study, we aimed to investigate this
question directly. We first examined the protein expression of
hCLOCK in CRC patients using immunofluorescent histochemistry. The
results revealed that hCLOCK was expressed in all CRC specimens and
there was a higher expression of hCLOCK in human CRC tissues
compared with that found in the paired non-cancerous tissues
(P=0.034).
We further analyzed hCLOCK expression patterns with
the respective clinical-pathological features of these colorectal
tumor tissues. We found that cancerous tissues with poorly
differentiated (grade II or above), or late-stage, Dukes’ grade
tumors (stage B or C) demonstrated increased expression of hCLOCK
protein. The greater expression of hCLOCK in the CRC tissues of
male patients compared with that in paired non-cancerous tissues
was observed, but not in female patients. This may be further
validated by using a larger sample set in the future.
qPCR analyses revealed that hClock was
expressed in all of the specimens. A significantly higher level of
hClock expression was found in human CRC compared with that
in the paired non-cancerous tissue (P=0.002). hClock had a
positive linear correlation with the hCLOCK protein in human CRCs
(r=0.474, P=0.008). A higher expression of hClock was
prevalent in tumor malignancies of a higher degree. These results
suggested that the hClock is closely related to the
development and prognosis of human CRCs.
We hypothesized that the circadian gene
hClock may be a proto-oncogene. A proto-oncogene is usually
a normal gene with rhythm, which is overexpressed and subsequently
loses its rhythmic character in tumor cells. Hoffman et al
observed deregulation of tumor-related genes in the hClock-silent
cell strain, and that the protein hCLOCK expression in breast tumor
cells was significantly higher compared with that in the adjacent
normal tissues (6). Fu et
al reported that c-myc in the tumorous cells of mice was
regulated by circadian genes (20). The hClock gene correlated
with cell cycles and had the characteristics of rhythmicity and
specificity (21). This hypothesis
requires further validation by examination with a larger sample
set.
Circadian genes are important in regulating certain
downstream clock-controlled genes (CCGs), including a variety of
tumor-related genes (22). In our
study, there was a significant positive linear correlation between
the hClock gene and the ARNT, HIF-1α and
VEGF genes in human CRCs. There was no significant
correlation between hClock and Bak, Bax,
Bid, TNFR I or TNFR II. Malignant tumors
promote the secretion of various proangiogenic factors, of which
the most important is VEGF. VEGF has a marked proangiogenic
function and is closely related to tumor differentiation, Dukes’
stage tumors and tumors with lymph node metastasis.
When tumors were under environmental hypoxic
conditions, the heterodimer HIF-1α/ARNT would be overexpressed,
resulting in a higher VEGF expression. hClock may interact
with HIF-1α/ARNT and activate VEGF to stimulate tumor angiogenesis
and metastasis (23,24).
In conclusion, our data suggest that aberrant
expression of hClock may correlate with the development and
progression of CRCs. However, the mechanisms leading to a high
expression of hClock in CRCs and how this promotes
tumorigenesis remains unclear. Further studies using a larger
sample set are required in order to elucidate the role of
hClock in human CRCs.
Acknowledgements
This study was supported by the Natural Science
Foundation of China (81070234 and 81000355).
References
|
1
|
Panda S, Hogenesch JB and Kay SA:
Circadian rhythms from flies to human. Nature. 417:329–335. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen-Goodspeed M and Lee CC: Tumor
suppression and circadian function. J Biol Rhythms. 22:291–298.
2007. View Article : Google Scholar
|
|
3
|
Ko CH and Takahashi JS: Molecular
components of the mammalian circadian clock. Hum Mol Genet.
15:R271–277. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Krugluger W, Brandstaetter A, Kállay E, et
al: Regulation of genes of the circadian clock in human colon
cancer: reduced period-1 and dihydropyrimidine dehydrogenase
transcription correlates in high-grade tumors. Cancer Res.
67:7917–7922. 2007. View Article : Google Scholar
|
|
5
|
Wang Y, Hua L, Lu C and Chen Z: Expression
of circadian clock gene human Period2 (hPer2) in human colorectal
carcinoma. World J Surg Oncol. 9:1662011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hoffman AE, Yi CH, Zheng T, et al: CLOCK
in breast tumorigenesis: genetic, epigenetic, and transcriptional
profiling analyses. Cancer Res. 70:1459–1468. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Alhopuro P, Björklund M, Sammalkorpi H, et
al: Mutations in the circadian gene CLOCK in colorectal cancer. Mol
Cancer Res. 8:952–960. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oshima T, Takenoshita S, Akaike M, et al:
Expression of circadian genes correlates with liver metastasis and
outcomes in colorectal cancer. Oncology reports. 25:1439–1446.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zheng S and Cai SR: Colorectal cancer
epidemiology and prevention study in China. Chinese-German J Clin
Oncol. 2:72–75. 2003. View Article : Google Scholar
|
|
10
|
Siegel R, Ward E, Brawley O and Jemal A:
Cancer statistics, 2011: the impact of eliminating socioeconomic
and racial disparities on premature cancer deaths. CA Cancer J
Clin. 61:212–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zheng S and Cai SR: Colorectal cancer
epidemiology and prevention study in China. Chinese-German J Clin
Oncol. 2:72–75. 2003. View Article : Google Scholar
|
|
12
|
Zhang EE, Liu AC, Hirota T, et al: A
genome-wide RNAi screen for modifiers of the circadian clock in
human cells. Cell. 139:199–210. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Scheving LA: Biological clocks and the
digestive system. Gastroenterology. 119:536–549. 2000. View Article : Google Scholar
|
|
14
|
Reppert SM and Weaver DR: Coordination of
circadian timing in mammals. Nature. 418:935–41. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hirota T and Fukada Y: Resetting mechanism
of central and peripheral circadian clocks in mammals. Zoolog Sci.
21:359–68. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Innominato PF, Focan C, Gorlia T, et al:
Circadian rhythm in rest and activity: a biological correlate of
quality of life and a predictor of survival in patients with
metastatic colorectal cancer. Cancer Res. 69:4700–4707. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Block KI, Block PB, Fox SR, et al: Making
circadian cancer therapy practical. Integr Cancer Ther. 8:371–386.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gekakis N, Stakins D, Nguyen HB, et al:
Role of the CLOCK protein in the mammalian circadian mechanism.
Science. 280:1564–1569. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Matsuo T, Yamaguchi S, Mitsui S, Emi A,
Shimoda F and Okamura H: Control mechanism of the circadian clock
for timing of cell division in vivo. Science. 302:255–259. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fu L, Pelicano H, Liu J, Huang P and Lee
C: The circadian gene Period2 plays an important role in tumor
suppression and DNA damage response in vivo. Cell. 111:41–50. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Polidarová L, Soták M, Sládek M, Pacha J
and Sumová A: Temporal gradient in the clock gene and cell-cycle
checkpoint kinase Weel expression along the gut. Chronobiol Int.
26:607–620. 2009.PubMed/NCBI
|
|
22
|
Gery S and Koeffler HP: The role of
circadian regulation in cancer. Cold Spring Harb Symp Quant Biol.
72:459–464. 2007. View Article : Google Scholar
|
|
23
|
Park JE, Lee DH, Lee JA, Park SG, Kim NS,
Park BC and Cho S: Annexin A3 is a potential angiogenic mediator.
Biochem Biophys Res Commun. 337:1283–1287. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chau NM, Rogers P, Aherne W, et al:
Identification of novel small molecule inhibitors of
hypoxia-inducible factor-1 that differentially block
hypoxia-inducible factor-1 activity and hypoxia-inducible
factor-1alpha induction in response to hypoxic stress and growth
factors. Cancer Res. 65:4918–4928. 2005. View Article : Google Scholar
|