Introduction
Hydrogen sulfide (H2S) gas exhibits
numerous physiological and pathological effects. H2S is
endogenously produced from L-cysteine by cystathionine γ-lyase
(CSE) and cystathionine β-synthase in mammalian tissues, and
endogenous hydrogen sulfide exists in the sodium hydrosulfide
(NaHS, 2/3) and H2S (1/3) forms (1). H2S has been demonstrated
to exert a therapeutic effect in a wide range of diseases,
including neuronal injury (2),
hypertension (3), myocardial
infarction (4) and hypoxic
pulmonary hypertension (5). In
addition, H2S exhibits antigrowth potential against a
wide variety of human cancer cells (6). In breast cancer cells, H2S
decreased the in vivo tumor mass through the inhibition of
cellular proliferation, induction of apoptosis and decrease in
nuclear factor-κB (NF-κB) levels (7). Furthermore, H2S has been
found to inhibit breast cancer-induced osteoclast formation and
activity, suppress osteoclastogenesis and prevent osteolysis
(8). As bone metastases are common
in breast cancer, H2S may have inhibitory effects on
breast cancer cell invasion and metastasis, thereby suppressing
osteoclastogenesis and osteolysis. However, there are few studies
concerning the anti-invasive effect of H2S on cancer
cells, including breast cancer cells.
The invasion of breast cancer is a multi-stage
process that involves abnormal signaling by transforming growth
factor-β (TGF-β). TGF-β acts as a tumor suppressor in the early
stages of carcinogenesis, but in late-stage breast cancer it
promotes invasion and metastatic dissemination (9). Moreover, TGF-β acquires a proinvasive
effect in the advanced stages of breast cancer through a complex
process known as epithelial-mesenchymal transition (EMT) (10). H2S has been found to
attenuate EMT in human alveolar epithelial cells (11). Moreover, H2S has been
reported to inhibit the pathogenesis of pulmonary and hepatic
fibrosis, and suppress the migration of human lung fibroblasts, all
of which involve the EMT process (12–14).
Therefore, it was hypothesized that H2S may act as a
suppressor of EMT in breast cancer and exhibit antitumor and
anti-invasive effects.
In the present study, breast cancer cells were
incubated with TGF-β1 to induce an EMT phenotype. The effects of
NaHS, an H2S-releasing molecule, on cell viability, cell
cycle, apoptosis, invasion and EMT were investigated. The protein
expression of CSE and phospho-p38 in breast cancer cells treated
with TGF-β1 was also analyzed.
Materials and methods
Cell culture
MCF-7 human breast cancer cells were purchased from
the cell bank of the Institute of Biochemistry and Cell Biology,
Shanghai Institutes for Biological Sciences, Chinese Academy of
Sciences (Shanghai, China). The cells were cultured with high
glucose Dulbecco’s modified Eagle’s medium (Invitrogen Life
Technologies, Carlsbad, CA, USA) supplemented with 10% fetal bovine
serum (Sijichun Bioengineering Materials Inc., Hangzhou, China),
100 U/ml penicillin and 100 mg/ml streptomycin at 37°C in a
humidified 5% CO2 incubator. After cultured cells
reached 70% confluence, the cells underwent trypsinization and were
subcultured with a 1:3 split ratio in new culture flasks.
DL-propargylglycine (PPG, Sigma, St. Louis, MO, USA), an inhibitor
of CSE, was used to inhibit endogenous H2S in MCF-7
cells.
Cell viability assay
An MTT (Sigma Chemical Co., St. Louis, MO, USA)
assay was performed to determine cell viability. Briefly, cells in
suspension at the logarithmic growth phase were added to each well
of 96-well culture plates at a density of 1×103
cells/ml, with 100 μl cell suspension in each well. The cells were
incubated for 24 h at 37°C in a humidified atmosphere with 5%
CO2. They were then incubated with TGF-β1 (100 ng/ml) or
recombinant human TGF-β1 (100 ng/ml, R&D Systems Inc.,
Minneapolis, MN, USA) with different concentrations of NaHS (0,
100, 200 and 500 μmol/l; Sigma Chemical Co.). Following treatment
for 12, 24, 48 and 72 h, 10 μl of 5 mg/ml MTT solution was added to
each well and the plates were incubated at 37°C for 4 h. Following
centrifugation at 1,409 × g for 10 min, the formazan pellets were
isolated by discarding the supernatant and then dissolved
completely in 100 μl dimethylsulfoxide (DMSO), agitating the plates
for 10 min. The optical density (OD) at 570 nm wavelength was
measured using an ELISA plate reader (Ricso RK201; Shenzhen Ricso
Technology Co., Ltd., Shenzhen, China) to determine the quantity of
pellet.
Cell cycle analysis
MCF-7 cells in the logarithmic growth phase were
cultured in serum-free medium for 24 h. They were then incubated
with TGF-β (100 ng/ml) and/or 500 μmol/l NaHS for 24 h. The cells
were harvested by trypsinization and following washes with cold
phosphate-buffered saline (PBS), the cells were fixed in cold 70%
ethanol. Finally, 1 μl propidium iodide (PI) staining solution
(containing 20 mg/ml PI and 1 mg/ml RNAse) was added to the
samples. The distribution of cells in each phase of the cell cycle
was measured and analyzed by flow cytometry (FACScan;
Becton-Dickinson, San Francisco, CA, USA). The percentage of cells
in the G0/G1, S and G2/M phases were calculated. Results were
acquired from 10,000 cells.
Apoptosis assay
MCF-7 cells were randomly divided into four groups:
Control, NaHS (500 μmol/l), TGF-β (100 ng/ml) and TGF-β+NaHS.
Following incubation for 24 h, at least 2×105 cells were
harvested from each group for the apoptosis assay. Subsequent to
centrifugation at 626 × g for 5 min and washing with PBS buffer,
the pellet was resuspended in 100 μl of 1X binding buffer and
incubated with 2.5 μl Annexin V and 5 μl PI (at a final
concentration of 10 μg/ml). After incubation in the dark for 30
min, apoptosis was immediately determined by FACScan flow cytometry
and the associated data were analyzed using Lysis II software
(Becton Dickinson). At least 10,000 events were analyzed for each
sample.
Cell migration assay
The in vitro invasion capability of MCF-7
cells was measured by Boyden chamber assay with matrigel (BD
Bioscience, Bedford, MA, USA) in 24-well tissue culture plates with
Transwell® filter membranes (5 μm pore; Costar, Boston,
MA, USA). The lower sides of the filters were coated with type I
collagen (0.5 mg/ml) and the lower section of the filter contained
low-serum media. In each well, 5×104 cells were
resuspended in 100 μl DMEM media and seeded in the upper part of a
Transwell® plate. The cells were then incubated with
NaHS (500 μmol/l) and/or TGF-β (100 ng/ml) for 24 h. Subsequent to
the removal of the cells on the upper surface of the filter, the
cells that had migrated to the lower part were stained with
hematoxylin and eosin (Sigma Chemical Co.) and counted under an
inverted light microscope (Olympus IX70; Olympus Optical Co., Ltd.,
Tokyo, Japan; magnification, ×200) as the number of migrated cells
(invasion index). Each sample was analyzed in triplicate and
repeated twice.
Western blot analysis
MCF-7 cells were cultured with NaHS (500 μmol/l)
and/or TGF-β (100 ng/ml) for 24 h. The cell proteins were then
extracted and a bicinchoninic acid protein concentration assay kit
(Beijing Biosea Biotechnology Co., Ltd., Beijing, China) was used
to determine their concentrations. The cell lysates (50 μg) were
resolved in 15% sodium dodecyl sulfate-polyacrylamide gels and
electrophoretically transferred to a polyvinylidene difluoride
membrane (Bio-Rad, Hercules, CA, USA). The membrane was blocked
with 5% skimmed milk powder in Tris-buffered saline containing
0.05% Tween-20 at room temperature for 2 h. It was then incubated
with primary mouse antibodies against SNAI1 (Snail) protein, CSE or
phospho-p38 (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA),
followed by horseradish peroxidase-conjugated rabbit anti-mouse
secondary antibody (Santa Cruz Biotechnology, Inc.) at 1:1,000
dilutions for 2 h at room temperature. Enhanced chemiluminescence
(Pierce® ECL Plus Western Blotting Substrate; Pierce
Biotechnology, Inc., Rockford, IL, USA) was used to visualize the
protein blots and β-actin served as an internal control.
Measurement of H2S
concentration in cell culture media
To measure the concentration of H2S, 500
ml culture media from each group was mixed with 425 ml distilled
water in a microtube containing zinc acetate (1% w/v; 250 ml).
N,N-dimethyl-p-phenylenediamine sulphate (20 mM; 133 ml) in 7.2 M
HCl was subsequently added, followed by FeCl3 (30 mM;
133 ml) in 1.2 M HCl. Trichloroacetic acid (10% w/v; 250 ml) was
then used to precipitate any protein. The OD value of the resulting
solution was measured using a 96-well microplate reader at 670 nm
wavelength (Tecan Group Ltd., Männedorf, Switzerland).
Statistical analysis
All quantitative data are presented as the mean ±
standard deviation. SPSS version 14.0 (SPSS, Inc., Chicago, IL,
USA) was used for statistical analysis. The statistical
significance of difference between two groups was determined by
Student’s t-test (unpaired, two tailed) and P<0.05 was
considered to indicate a statistically significant difference.
Results
NaHS inhibits MCF-7 cell growth induced
by TGF-β1
The MCF-7 cells were incubated with TGF-β (100
ng/ml), then treated with various concentrations of NaHS (0, 100,
200 and 500 μmol/l) for 12, 24, 48 and 72 h. MCF-7 cells treated
with PBS served as a control. The MTT assay revealed that compared
with the control, TGF-β treatment promoted cell proliferation and
increased cell viability in a time-dependent manner. However, NaHS
inhibited cell viability in the MCF-7 cells treated with TGF-β in a
dose- and time-dependent manner. NaHS exhibited the most potent
effect on cell viability at a 500 μmol/l concentration at all
time-points (Fig. 1A). Therefore,
in subsequent experiments, a concentration of 500 μmol/l was used
for NaHS treatment of the cells.
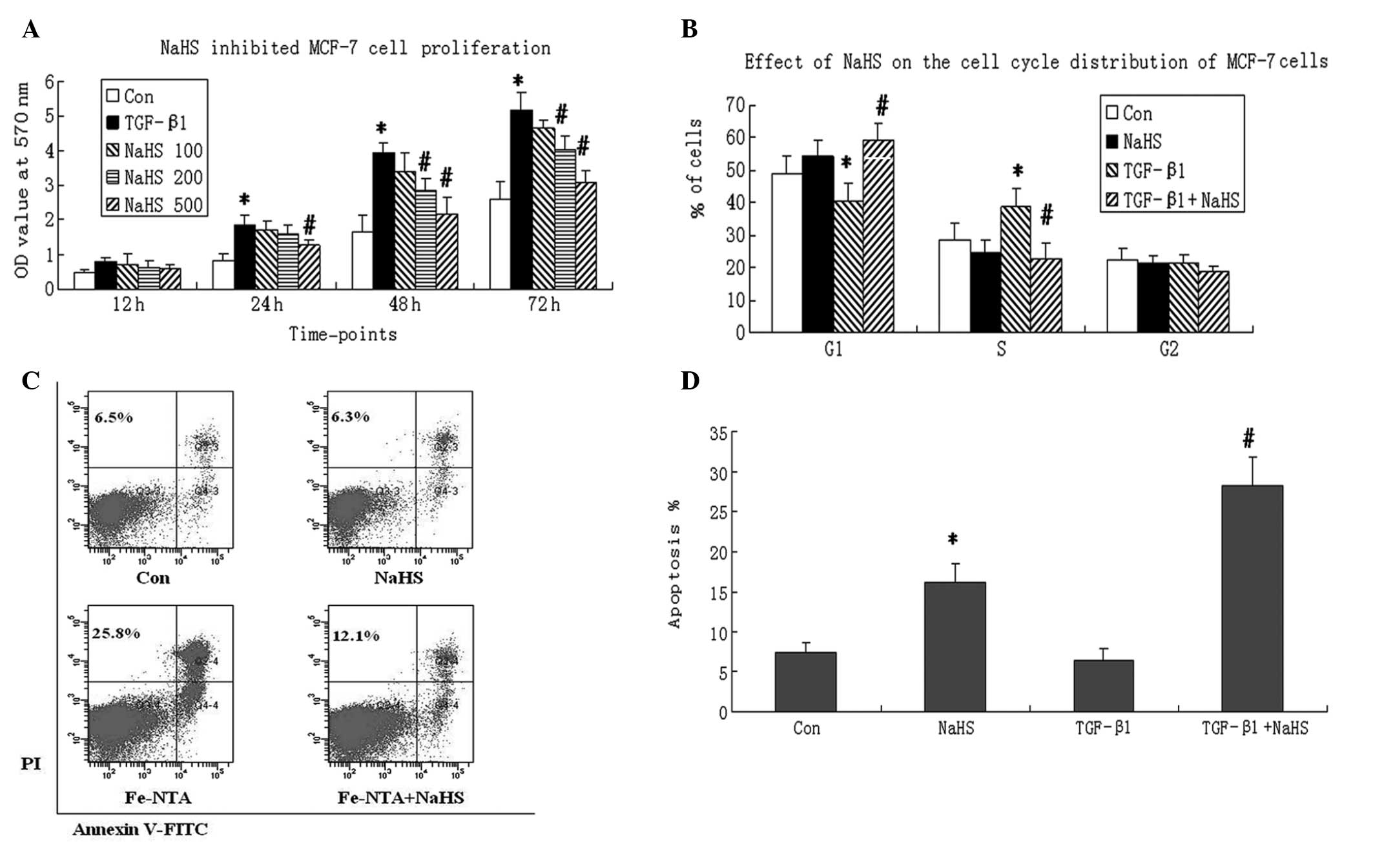 | Figure 1Effect of sodium hydrosulfide (NaHS)
on cell viability, cell cycle and apoptosis in MCF-7 breast cancer
cells induced by transforming growth factor-β1 (TGF-β1). (A) NaHS
inhibited the proliferation of TGF-β1-treated MCF-7 cells. MCF-7
cells were seeded in 96-well culture plates at a density of
1×103 cells/ml. After adherence, cells were incubated
with PBS (Control group, Con), TGF-β1 (100 ng/ml, TGF-β1 group),
TGF-β1 plus various concentrations of NaHS (0, 100, 200 or 500
μmol/l). Following incubation for 12, 24, 48 and 72 h, an MTT assay
was performed to determine cell viability. At least three
independent experiments were conducted. Data are presented as
optical density (OD) values at 570 nm wavelength. (B) MCF-7 cells
were cultured in serum-free medium for 24 h, and then were
incubated with PBS (Con), NaHS (500 μmol/l), TGF-β1 (100 ng/ml),
and TGF-β1+NaHS for 24 h. Propidium iodide (PI, 20 μg/ml) staining
was performed to determine the percentages of cells in the G0/G1, S
and G2/M phases. (C) MCF-7 cells were randomly divided into
control, NaHS, TGF-β1 and TGF-β1+NaHS groups. Annexin V-fluorescein
isothiocyanate (FITC) and PI double staining was performed to
detect cell apoptosis using flow cytometry 24 h following
treatment. NaHS treatment increased the apoptotic rate in MCF-7
cells regardless of whether they were treated with TGF-β1.
Representative images from three experiments are shown. (D)
Apoptotic rates from the control, NaHS, TGF-β1 and TGF-β1+NaHS
groups. The Annexin V+/PI− and Annexin
V+/PI+ populations were considered to
represent apoptotic cells. A two-tailed, unpaired t-test was
performed to compare the differences between two groups. Data are
presented as the mean ± standard deviation. *P<0.05,
vs. the control group (Con). #P<0.05, vs. the TGF-β1
group. |
Cell cycle distribution was analyzed by flow
cytometry to investigate the detailed mechanism of the
antiproliferative activity of NaHS. The MCF-7 cells were treated
with NaHS (500 μmol/l) and/or TGF-β (100 ng/ml) for 24 h. The TGF-β
treatment decreased the percentage of cells in the G0/G1 phase and
increased the percentage of cells in the S phase. NaHS increased
the percentage of cells in the G1 phase significantly, while
decreasing the percentage of cells in the S phase in TGF-β-treated
MCF-7 cells (P<0.05). However, no change was found in the
percentage of cells in the G2 phase following TGF-β1 or NaHS
treatment (Fig. 1B). This assay
indicated that NaHS inhibited cell proliferation by inducing G0/G1
phase arrest in MCF-7 cells treated with TGF-β1.
To investigate whether a decrease in cell viability
was a result of the proapoptotic effect of NaHS, an apoptosis assay
was performed in MCF-7 cells by double staining with Annexin
V-fluorescein isothiocyanate and PI. It was found that TGF-β1
treatment alone did not change the apoptotic rate of MCF-7 cells.
However, treatment with NaHS increased the apoptotic rate
regardless of whether the cells had been treated with TGF-β1
(P<0.05, Fig. 1C and D).
Furthermore, MCF-7 cells treated with TGF-β1 and NaHS demonstrated
a higher apoptotic rate than control cells, which indicates that
induction with TGF-β1 may enhance sensitivity to NaHS.
NaHS decreases cell invasion and EMT
induced by TGF-β1
To investigate whether NaHS can inhibit the
migration of breast cancer cells, the invasive capability of MCF-7
cells was determined by a Boyden chamber invasion assay. The cells
were incubated with TGF-β1 to induce an invasive state, and the
results revealed that TGF-β1 significantly increased the invasion
index of MCF-7 cells (P<0.05). NaHS treatment following TGF-β
administration significantly decreased the invasion index compared
with that of cells treated with TGF-β only (P<0.05, Fig. 2A). However, compared with control
cells, NaHS treatment alone made no significant difference to the
invasion index.
To investigate whether the EMT process is involved
in the anti-invasive effect of NaHS, western blot analysis was
performed to determine the expression of an EMT marker, Snail
protein. TGF-β1 treatment significantly induced EMT in MCF-7 cells,
as evidenced by increased expression of Snail protein. Following
NaHS treatment, TGF-β1-induced Snail protein expression was
significantly decreased. However, compared with control MCF-7
cells, Snail protein levels remained unchanged in cells treated
with NaHS alone (Fig. 2B and
C).
NaHS increases CSE protein expression and
supernatant H2S levels
The expression of CSE protein in TGF-β1-induced
MCF-7 cells was significantly increased by NaHS treatment.
Moreover, the CSE protein level was decreased following TGF-β1
treatment alone (Fig. 3A and B).
To investigate whether an enhanced CSE protein level produces a
greater level of endogenous H2S, the H2S
levels in the cell culture media were measured. The H2S
level in the cell culture media in cells treated with 100 ng/ml
TGF-β1 was significantly decreased in comparison with the control
cells (P<0.05). When compared with the TGF-β1-treated cells, the
cells treated with NaHS had a significantly higher H2S
level at all concentrations of NaHS (P<0.05, Fig. 3C). These results indicate that the
anticancer effect of NaHS may be mediated by activation of the
CSE/H2S pathway.
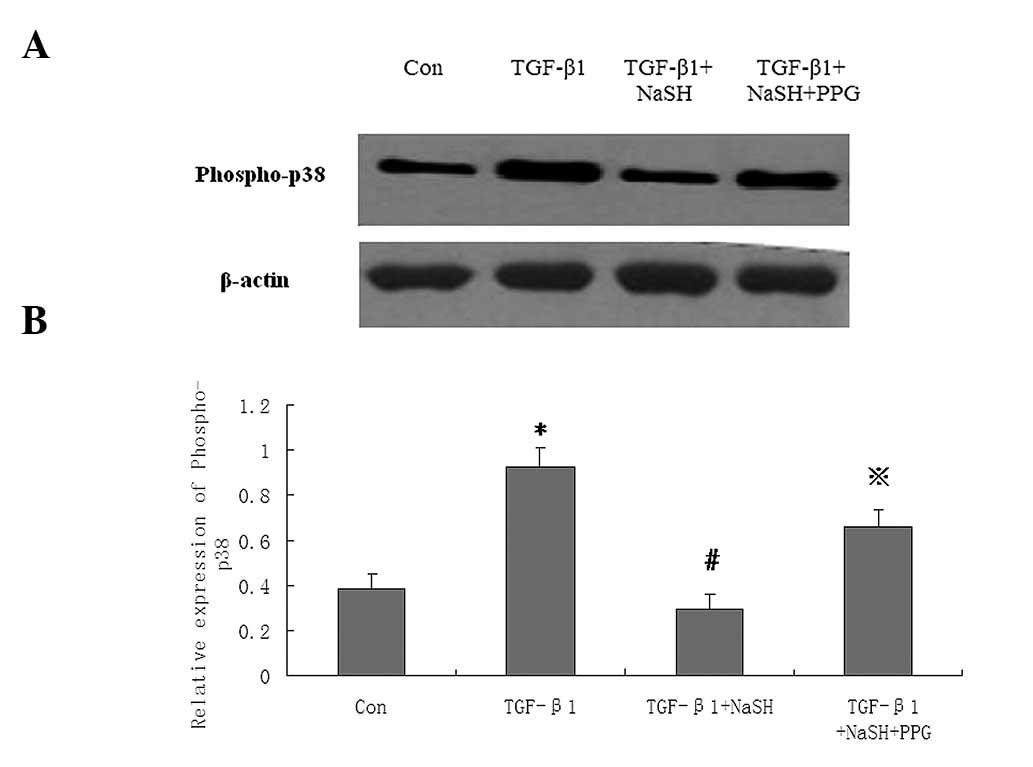
NaHS decreases p38 mitogen-activated
protein kinase (MAPK) phosphorylation in MCF-7 cells stimulated by
TGF-β1
To investigate the underlying signaling pathways in
NaHS-treated MCF-7 cells, the expression of phospho-p38 MAPK, a
signaling protein associated with apoptosis and EMT, was
investigated. The MCF-7 cells were pretreated with PPG (an
inhibitor of CSE, 100 μmol/l) for 1 h, then administered either
TGF-β1 (100 ng/ml), TGF-β1+NaHS (500 μmol/l) or TGF-β1+NaHS+PPG,
and incubated for 24 h. Western blot analysis revealed that TGF-β1
significantly increased phospho-p38 protein expression. However,
NaHS significantly decreased the phospho-p38 protein levels in
MCF-7 cells treated with TGF-β1, which was attenuated by PPG
pretreatment (Fig. 4).
Discussion
In the present study, NaHS (a bioactive compound
releasing H2S) exhibited anticancer effects in
TGF-β1-treated MCF-7 breast cancer cells, demonstrated by the
inhibition of cell proliferation, cell cycle arrest in G0/G1 phase
and induction of apoptosis. NaHS treatment also inhibited tumor
invasion and decreased protein expression of an EMT marker, Snail.
The underlying mechanisms may be associated with increased
endogenous CSE protein expression and decreased p38 MAPK
phosphorylation in MCF-7 cells stimulated by TGF-β1 following NaHS
administration.
The anticancer effects of exogenous H2S
on breast cancer cells indicate a novel therapeutic strategy for
breast cancer. NaHS was revealed to reduce cell viability in a
dose- and time-dependent manner, and the detailed mechanism lies in
G0/G1 cell cycle arrest and induction of apoptosis. These results
are in accordance with another study, which revealed that
H2S-releasing aspirin decreased tumor mass through
inhibition of cell proliferation and induction of G0/G1 arrest in
estrogen receptor-negative breast cancer cells (7). In the present study, NaHS, which is
an exogenous H2S-releasing molecule and one form of
H2S in mammalian tissues, was used, therefore providing
additional evidence regarding the anticancer effects of
H2S. In a variety of pathological conditions,
H2S has been found to exhibit potent antiapoptotic
effects including in hypoxia-induced mouse hippocampal neurons
(15), high-glucose-induced rat
cardiomyocytes (16) and hepatic
ischemia/reperfusion injury (17).
However, in the present study, NaHS was found to exhibit a potent
apoptotic effect on breast cancer cells, particularly in cells
treated with TGF-β1. This contradiction may be a result of
differences in the reaction to H2S between non-tumor
cells and tumor cells as H2S has also been demonstrated
to mediate the antisurvival effect of sulforaphane in human
prostate cancer cells (18).
Therefore, there may be benefits in introducing
H2S-releasing therapeutic agents in the treatment of
cancer.
In the present study, H2S was found to
have an inhibitory effect on invasion and EMT in breast cancer
cells. EMT is a complex, multi-step process that involves
epithelial cells developing a malignant phenotype, including
invasive, migratory and metastatic capabilities (19). MCF-7 cells were incubated with
TGF-β1 to induce the EMT state, significantly increasing the
invasion index of the cells and increasing the protein expression
of an EMT marker, Snail. NaHS was revealed to significantly
decrease this invasion index and the expression of Snail protein in
the MCF-7 cells induced by TGF-β. This finding is in accordance
with another study, which observed that H2S inhibited
EMT in human alveolar epithelial cells, demonstrated by decreased
vimentin expression and increased E-cadherin expression (11). EMT is a vital process in driving
epithelial cells to acquire a malignant phenotype and invasive
properties (20). The results of
the present study indicate that NaHS suppressed invasion through
inhibition of the EMT process in MCF-7 cells. It was also found
that tumor invasion and Snail protein expression remained unchanged
following NaHS treatment in control MCF-7 cells without TGF-β1.
This indicates that compared with breast cancer cells without
metastasis, NaHS may demonstrate an enhanced effect on the
metastasis and EMT of breast cancer cells, thereby providing
further evidence for the causal link between EMT and invasion in
TGF-β1-treated MCF-7 cells.
In the present study, NaHS increased CSE protein
expression and supernatant H2S levels. H2S
can be generated by CSE with L-cysteine as its substrate. Increased
expression of CSE protein following NaHS administration indicates
that the anticancer effect of NaHS not only depend on direct change
from NaHS to H2S in extracellular fluid, but also may be
mediated by activation of the CSE/H2S pathway, thereby
acquiring a more durable effect. The elevated H2S level
in the culture media provides further evidence that the
CSE/H2S pathway instigates the anticancer effects of
NaHS. In fact, a number of H2S-releasing drugs may
produce their effects through activation of the CSE/H2S
pathway. For example, one H2S donor,
S-propargyl-cysteine, exhibits its anticancer effect by increasing
CSE protein expression in gastric cancer cells (21).
In order to investigate the detailed mechanisms
underlying the inhibitory effect of H2S on invasion and
EMT in TGF-β1-treated MCF-7 cells, the expression of a signal
protein associated with apoptosis and invasion was measured. TGF-β1
treatment alone increased phospho-p38 protein expression, but the
addition of NaHS decreased the expression of phospho-p38 in
TGF-β1-treated cells. A previous study found that p38 MAPK promoted
successful invasion and metastasis in tumor cells (22). p38 MAPK activation has also been
demonstrated to mediate TGF-β1-induced EMT in A549 alveolar
epithelial cells (23), which is
in accordance with the results of the present study. The inhibition
of p38 MAPK has been found to reverse the EMT process and may be a
potential therapeutic strategy to decrease cancer invasion
(24). Therefore, decreased
phospho-p38 may be an underlying mechanism for the inhibition of
EMT by NaHS. To further confirm the inhibition of phospho-p38 by
the CSE/H2S pathway, MCF-7 cells were pretreated with
PPG (an inhibitor of CSE) then incubated with TGF-β1 and NaHS. The
decrease in phospho-p38 expression in TGF-β1-treated MCF-7 cells
following administration of NaHS was significantly attenuated by
PPG. These results suggest a novel mechanism for the anticancer
effects of exogenous H2S via CSE/H2S-induced
inhibition of cell growth, induction of apoptosis, and the
inhibition of invasion and EMT.
The detailed association between p38 MAPK and EMT
remains unknown. One study demonstrated that the p38/NF-κB/Snail
pathway was involved in the caffeic acid-induced inhibition of the
migratory capacity of malignant human keratinocytes (25). This suggests that inhibition of
NF-κB may mediate the causal association between decreased
phospho-p38 expression and reduced EMT by NaHS, which warrants
further investigation. The anticancer effect of NaHS and the
potential CSE/H2S pathway also require further
verification in animal models.
In conclusion, the results of the present study
demonstrate that an H2S donor, NaHS, exhibits anticancer
effects on breast cancer cells, as evidenced by inhibition of
proliferation, induction of apoptosis, and the inhibition of
invasion and EMT. The underlying mechanisms of the NaHS anticancer
effect may be through activation of the CSE/H2S pathway
and decreased phospho-p38. These results suggest that exogenous
H2S may be a potential therapeutic strategy for breast
cancer.
References
|
1
|
Renga B: Hydrogen sulfide generation in
mammals: the molecular biology of cystathionine-β-synthase (CBS)
and cystathionine-γ-lyase (CSE). Inflamm Allergy Drug Targets.
10:85–91. 2011.
|
|
2
|
Zhang LM, Jiang CX and Liu DW: Hydrogen
sulfide attenuates neuronal injury induced by vascular dementia via
inhibiting apoptosis in rats. Neurochem Res. 34:1984–1992. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ahmad FU, Sattar MA, Rathore HA, et al:
Exogenous hydrogen sulfide (H2S) reduces blood pressure
and prevents the progression of diabetic nephropathy in
spontaneously hypertensive rats. Ren Fail. 34:203–210.
2012.PubMed/NCBI
|
|
4
|
Qipshidze N, Metreveli N, Mishra PK,
Lominadze D and Tyagi SC: Hydrogen sulfide mitigates cardiac
remodeling during myocardial infarction via improvement of
angiogenesis. Int J Biol Sci. 8:430–441. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Olson KR, Whitfield NL, Bearden SE, et al:
Hypoxic pulmonary vasodilation: a paradigm shift with a hydrogen
sulfide mechanism. Am J Physiol Regul Integr Comp Physiol.
298:R51–R60. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chattopadhyay M, Kodela R, Nath N, et al:
Hydrogen sulfide-releasing NSAIDs inhibit the growth of human
cancer cells: a general property and evidence of a tissue
type-independent effect. Biochem Pharmacol. 83:715–722. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chattopadhyay M, Kodela R, Nath N,
Barsegian A, Boring D and Kashfi K: Hydrogen sulfide-releasing
aspirin suppresses NF-κB signaling in estrogen receptor negative
breast cancer cells in vitro and in vivo. Biochem Pharmacol.
83:723–732. 2012.PubMed/NCBI
|
|
8
|
Frantzias J, Logan JG, Mollat P, et al:
Hydrogen sulphide-releasing diclofenac derivatives inhibit breast
cancer-induced osteoclastogenesis in vitro and prevent osteolysis
ex vivo. Br J Pharmacol. 165:1914–1925. 2012. View Article : Google Scholar
|
|
9
|
Daroqui MC, Vazquez P, Bal de Kier Joffé
E, Bakin AV and Puricelli LI: TGF-β autocrine pathway and MAPK
signaling promote cell invasiveness and in vivo mammary
adenocarcinoma tumor progression. Oncol Rep. 28:567–575. 2012.
|
|
10
|
Wendt MK, Smith JA and Schiemann WP:
Transforming growth factor-β-induced epithelial-mesenchymal
transition facilitates epidermal growth factor-dependent breast
cancer progression. Oncogene. 29:6485–6498. 2010.
|
|
11
|
Fang LP, Lin Q, Tang CS and Liu XM:
Hydrogen sulfide attenuates epithelial-mesenchymal transition of
human alveolar epithelial cells. Pharmacol Res. 61:298–305. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fang L, Li H, Tang C, Geng B, Qi Y and Liu
X: Hydrogen sulfide attenuates the pathogenesis of pulmonary
fibrosis induced by bleomycin in rats. Can J Physiol Pharmacol.
87:531–538. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fan HN, Wang HJ, Ren L, et al: Decreased
expression of p38 MAPK mediates protective effects of hydrogen
sulfide on hepatic fibrosis. Eur Rev Med Pharmacol Sci. 17:644–652.
2013.PubMed/NCBI
|
|
14
|
Fang LP, Lin Q, Tang CS and Liu XM:
Hydrogen sulfide suppresses migration, proliferation and
myofibroblast transdifferentiation of human lung fibroblasts. Pulm
Pharmacol Ther. 22:554–561. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Luo Y, Liu X, Zheng Q, et al: Hydrogen
sulfide prevents hypoxia-induced apoptosis via inhibition of an
H2O2-activated calcium signaling pathway in
mouse hippocampal neurons. Biochem Biophys Res Commun. 425:473–477.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou X and Lu X: Hydrogen sulfide inhibits
high-glucose-induced apoptosis in neonatal rat cardiomyocytes. Exp
Biol Med (Maywood). 238:370–374. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bos EM, Snijder PM, Jekel H, et al:
Beneficial effects of gaseous hydrogen sulfide in hepatic
ischemia/reperfusion injury. Transpl Int. 25:897–908. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pei Y, Wu B, Cao Q, Wu L and Yang G:
Hydrogen sulfide mediates the anti-survival effect of sulforaphane
on human prostate cancer cells. Toxicol Appl Pharmacol.
257:420–428. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huber MA, Kraut N and Beug H: Molecular
requirements for epithelial-mesenchymal transition during tumor
progression. Curr Opin Cell Biol. 17:548–558. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sánchez-Tilló E, Liu Y, de Barrios O, et
al: EMT-activating transcription factors in cancer: beyond EMT and
tumor invasiveness. Cell Mol Life Sci. 69:3429–3456.
2012.PubMed/NCBI
|
|
21
|
Ma K, Liu Y, Zhu Q, et al: H2S
donor, S-propargyl-cysteine, increases CSE in SGC-7901 and
cancer-induced mice: evidence for a novel anti-cancer effect of
endogenous H2S? PLoS One. 6:e205252011.
|
|
22
|
del Barco Barrantes I and Nebreda AR:
Roles of p38 MAPKs in invasion and metastasis. Biochem Soc Trans.
40:79–84. 2012.PubMed/NCBI
|
|
23
|
Chen HH, Zhou XL, Shi YL and Yang J: Roles
of p38 MAPK and JNK in TGF-β1-induced human alveolar epithelial to
mesenchymal transition. Arch Med Res. 44:93–98. 2013.
|
|
24
|
Antoon JW, Nitzchke AM, Martin EC, et al:
Inhibition of p38 mitogen-activated protein kinase alters microRNA
expression and reverses epithelial-to-mesenchymal transition. Int J
Oncol. 42:1139–1150. 2013.PubMed/NCBI
|
|
25
|
Yang Y, Li Y, Wang K, Wang Y, Yin W and Li
L: P38/NF-κB/snail pathway is involved in caffeic acid-induced
inhibition of cancer stem cells-like properties and migratory
capacity in malignant human keratinocyte. PLoS One.
8:e589152013.PubMed/NCBI
|