Introduction
MicroRNAs (miRNAs), that are abundantly expressed in
the brain, have emerged as key mediators of post-transcriptional
gene silencing in ischemic stroke biology (1). Several studies investigating
successfully implemented miRNA-based therapeutics have provided new
treatment strategies for ischemic stroke, by means of regulating
large sets of genes in numerous associated pathways (1).
Tissue-specific miRNAs have been demonstrated to
serve as diagnostically sensitive plasma biomarkers in tissue
injury (2). A number of miRNAs,
including miR-124 and miR-210, have been investigated for their
role as potential diagnostic markers in cerebral ischemia (3,4). It
has been reported that miR-21 and miR-24 are upregulated in stroke
patients (5), and that they can
also be detected in the brain of rats subjected to middle cerebral
artery occlusion (MCAO), a common animal model in cerebral ischemia
(CI) research (6)
Overexpression of miR-21 protects against ischemic
neuronal death by downregulating FASLG (7). miR-24 suppresses cardiomyocyte
apoptosis, in part by directly repressing Bim, a novel member of
the Bcl-2 family, which positively regulates apoptosis (8). miR-23a is part of the same cluster as
miR-24 and by regulating X-linked inhibitor of apoptosis protein
(XIAP), contributes to sex differences in responses to cerebral
ischemia (9). Furthermore, miR-497
promotes ischemic neuronal death by negatively regulating
anti-apoptotic proteins, Bcl-2 and Bcl-w (10), and Bcl-2 is also a potential target
of miR-21 (11,12)
In the present study, it was hypothesized that
miR-21 and miR-24 have important roles in ischemic stroke, but the
mechanism underlying this effect, whether it involves targeting
Bcl-2, Bim and XIAP, remains unclear. Furthermore, whether these
miRNAs may serve as non-invasive plasma biomarkers has not been
investigated. In the present study, the difference of plasma miR-21
and miR-24 in acute cerebral infarction (ACI) and healthy controls
was investigated, and the correlations between plasma miR-21,
miR-24 and NIHSS scores were analyzed. Furthermore, we investigated
the expression of miR-21, miR-24 and apoptosis/autophage-associated
proteins in N2A cells following oxygen-glucose deprivation (OGD)
and reoxygenation, and the effects on protein Bcl-2, XIAP following
the induction of a gain of function in miR-21 and miR-24.
Materials and methods
Study population and ethics
A total of 68 patients with ACI participated in the
study, with 21 healthy individuals used as the controls. Patients
with ACI were recruited from the Department of Neurology at Peking
University Third Hospital (Beijing, China). The methods were
approved by the local ethics committee of Peking University Third
Hospital and all study participants signed informed consent forms.
The diagnosis of ACI was conducted based on patient history, lab
examination, neurological deficit, magnetic resonance imaging (MRI)
and magnetic resonance angiography (MRA) results. Patients with a
history of tumor, immune disease, blood disease, acute infectious
disease, renal or liver failure were excluded. The severity was
further evaluated by NIHSS. Plasma samples were extracted from
ethylene diamine tetraacetic acid (EDTA)-containing tubes (BD
Biosciences, Franklin Lakes, NJ, USA) and stored at −80°C.
Hemolysis may affect the result, so the sample hemolysis can not be
used. The first samples were collected within 24 h following the
patients’ admission to the hospital and the control plasma samples
were obtained from healthy volunteers. The mean age of the control
is matched with that of ACI patients.
N2A cell culture and OGD
N2A cells were purchased from the cell resource
center of Peking Union Medical College (Beijing, China). An OGD
model was utilized to mimic ischemic-like conditions in
vitro (10) and a Bio-Bag was
used to produce anaerobic environment.
Gain of miR-21 and miR-24 function
N2A cells were plated in 6-well plates and treated
with miR-21 or miR-24 mimics, inhibitor and no template control
(NTC). The cells were harvested following 24 or 48 h to detect
miRNA or protein expression levels.
Western blotting
Total protein was isolated from N2A cells. Samples
of 30–60 μg protein were subjected to 10–12% SDS/polyacrylamide gel
(SDS/PAGE) and transferred onto polyvinylidene fluoride (PVDF)
membranes (0.45 μm). The membranes were blocked in 5% skimmed milk
(dissolved in 1X TBS Tween-20 buffer) for 1 h at room temperature.
Following this, the membrane was incubated with the primary
antibody overnight at 4°C then washed with TBST three times at 10
min intervals, followed by incubation with secondary antibody at
room temperature for 2 h and another three washes with TBST. The
membrane was scanned with LI-COR Odyssey (800 nm; LI-COR, Inc.,
Lincoln, NE, USA). All primary antibodies were purchased from Cell
Signaling Technology, Inc. (CST, Boston, MA, USA; 1:1,000 in 5%
BSA). Mouse anti-GAPDH antibodies were purchased from CoWin Biotech
Co., Ltd. (Beijing, China; 1:5,000 in 5% BSA). The secondary
fluorescein-conjugated antibody was purchased from LI-COR, Inc.
(1:10,000, 5% skimmed milk).
RNA isolation
Total RNA was isolated from N2A cell culture and
plasma by TRIzol and TRIzol LS (Invitrogen Life Technologies, Inc.,
Carlsbad, CA, USA; no. 15596-026, no. 10296-028) respectively,
according to the manufacturer’s instructions. In particular, the
EDTA anticoagulant plasma was separated by centrifugation for 10
min at 3,625 × g, then 750 μl TRIzol LS was added to 250 μl plasma
followed by 5 min incubation at room temperature and 10 μl
cel-mir-39 (1 μM) was added to each sample as an internal
calibrator (13). The miRNA
sequences are summarized in Table
I.
 | Table ImiRNA sequences. |
Table I
miRNA sequences.
| miRNA | Sequence |
|---|
| hsa-mir-21-5p |
UAGCUUAUCAGACUGAUGUUGA |
| hsa-mir-24-3p |
UGGCUCAGUUCAGCAGGAACAG |
| cel-miR-39 |
UCACCGGGUGUAAAUCAGCUUG |
Reverse transcription (RT) and
quantitative (q)PCR
Total RNA extracted from N2A cell cultures was
reversely transcribed using TaqMan miRNA RT kit (Applied
Biosystems, Carlsbad, CA, USA). PCRs were then conducted using the
TaqMan® miRNA assay kit (Applied Biosystems). The
relative miRNA levels were normalized to endogenous U6 expression
for each sample (10). RT primers
were dissolved in nuclease-free water and mixed in equal
concentration to generate a RT primer mix. RT of plasma RNA was
conducted according to the manual for the reverse transcription
system (RevertAid First Strand cDNA Synthesis kit, no. 1621).
qRT-PCR was performed using the SYBR Green PCR Master Mix kit
(CWBIO, no. CW0956) and was processed in 96-well plates on a 7500HT
analyzer (Applied Biosystems). Expression values were normalized
using the mean threshold cycle (Ct) obtained from the spiked-in
controls cel-mir-39 (calculation formula: 2 exp (mean Ct spiked-in
controls - Ct target miRNA) and log transformed (14).
The TaqMan® miRNA assay (Applied
Biosystems) uses gene-specific stem loop reverse-transcription
primers and TaqMan® probes to detect mature miRNA
transcripts. This method is highly specific but very expensive. So,
we detected plasma miRNA using SYBR qRT-PCR (15–17),
which produced a good amplification and melting curve (data not
shown). Table II summarizes the
sequences of RT and PCR primer used.
| Table IISequences of RT primers and PCR
primers. |
Table II
Sequences of RT primers and PCR
primers.
| miRNA | RT primer | F/R | PCR primer |
|---|
| has-mir-21-5p | ***TCAACA | F |
GCCGCTAGCTTATCAGACTGA |
| | R | GTGCAGGGTCCGAGGT |
| has-mir-24-3p | ***CTGTTC | F |
GGTGGCTCAGTTCAGCAG |
| | R | GTGCAGGGTCCGAGGT |
| cel-miR-39 | ***CAAGCT | F |
GCGTCACCGGGTGTAAATC |
| | R | GTGCAGGGTCCGAGGT |
| U6 | CGCTTCACGAAT | F |
GCTTCGGCAGCACATATACTAAAAT |
| TTGCGTGTCAT | R |
CGCTTCACGAATTTGCGTGTCAT |
Statistical data analysis
Statistical analysis was performed by SPSS 16.0
software (SPSS, Inc., Chicago, IL, USA). For the data with a
non-normal distribution, the comparison of two independent groups
was analyzed by non-parametric Mann-Whitney U test. Spearman
correlation between the two variables was performed. A value of
P<0.05 was considered to indicate a statistically significant
result. The data with a non-normal distribution was presented as
the median value (lower quartile; upper quartile). The relative
plasma miRNA was presented as box-and-whisker plots. The normal
distribution data was analyzed using the Student’s t-test.
Quantitative data for the cells were expressed as the mean ± SD
based on at least three separate experiments of triplicate samples.
ImageJ software was used to analyze selected bands in the western
blot analysis. Differences among groups were statistically analyzed
by the one-way analysis of variance (ANOVA) test.
Results
Patient clinical characteristics
Table III
summarizes the demographic and baseline clinical characteristics of
the patients that participated in the study. The median age was not
different (P=0.095) between the stroke patients and the controls.
However, the stroke patients exhibited an increased percentage of
the risk factors, compared with the controls, including
hypertension 54.4%, diabetes 17%, hyperlipidemia 11.7% and history
of cardiovascular and cerebrovascular disease 24%.
| Table IIIDemographic and clinical
characteristics in acute ischemic stroke patients and healthy
controls. |
Table III
Demographic and clinical
characteristics in acute ischemic stroke patients and healthy
controls.
| Characteristic | Control | ACI |
|---|
| Sample size | 21 | 68 |
| Age | 58 (54,67) | 64 (55,76) |
| Gender (M/F) | 10/9 | 45/23 |
| Hypertension | - | 37 (54.4%) |
| Diabetes | - | 17 (25%) |
| Hyperlipidaemia | - | 8 (11.7%) |
| History of CVD and
CVA | - | 24 (35.3%) |
| Smoking | - | 18 (26.5%) |
| Alcohol | - | 8 (11.8%) |
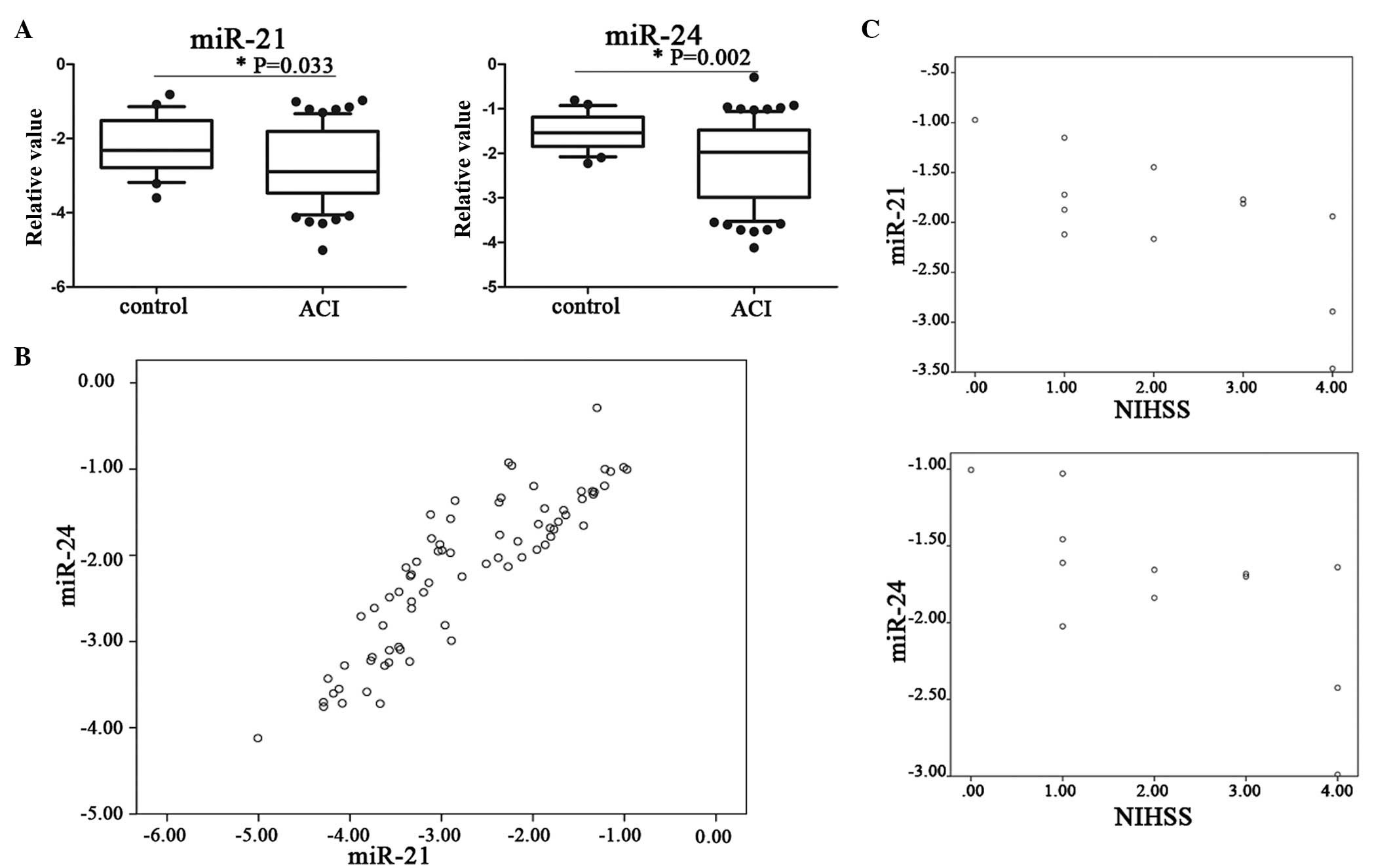
Expression of miR-21 and miR-24 in plasma
of stroke patients
In the present study, it was not possible to use any
specific miRNA or set of miRNAs as endogenous controls in
plasma/serum (18), so synthesized
cel-miR-39 (40 pmol/l) was added, as internal calibrators, as
described previously (13).
To investigate whether miR-21 and miR-24 are
involved in acute ischemic stroke, plasma miR-21 and miR-24 levels
were examined in the patients and controls. It was identified that
plasma miR-21 and miR-24 levels were decreased in acute ischemic
stroke compared with the controls (P<0.05; Fig 1A).
It was identified that plasma miR-21 and miR-24
levels were positively correlated (r=0.884, P<0.01;Fig. 1B). To examine the associations
between plasma miR-21 and miR-24 and the outcome of ACI, the
correlations between the expression of miR-21, miR-24 and NIHSS
score were examined. We identified a significant negative
correlation within the first day after onset (r=−0.703, P<0.05;
r=−0.694, P<0.05, respectively; Fig
1C).
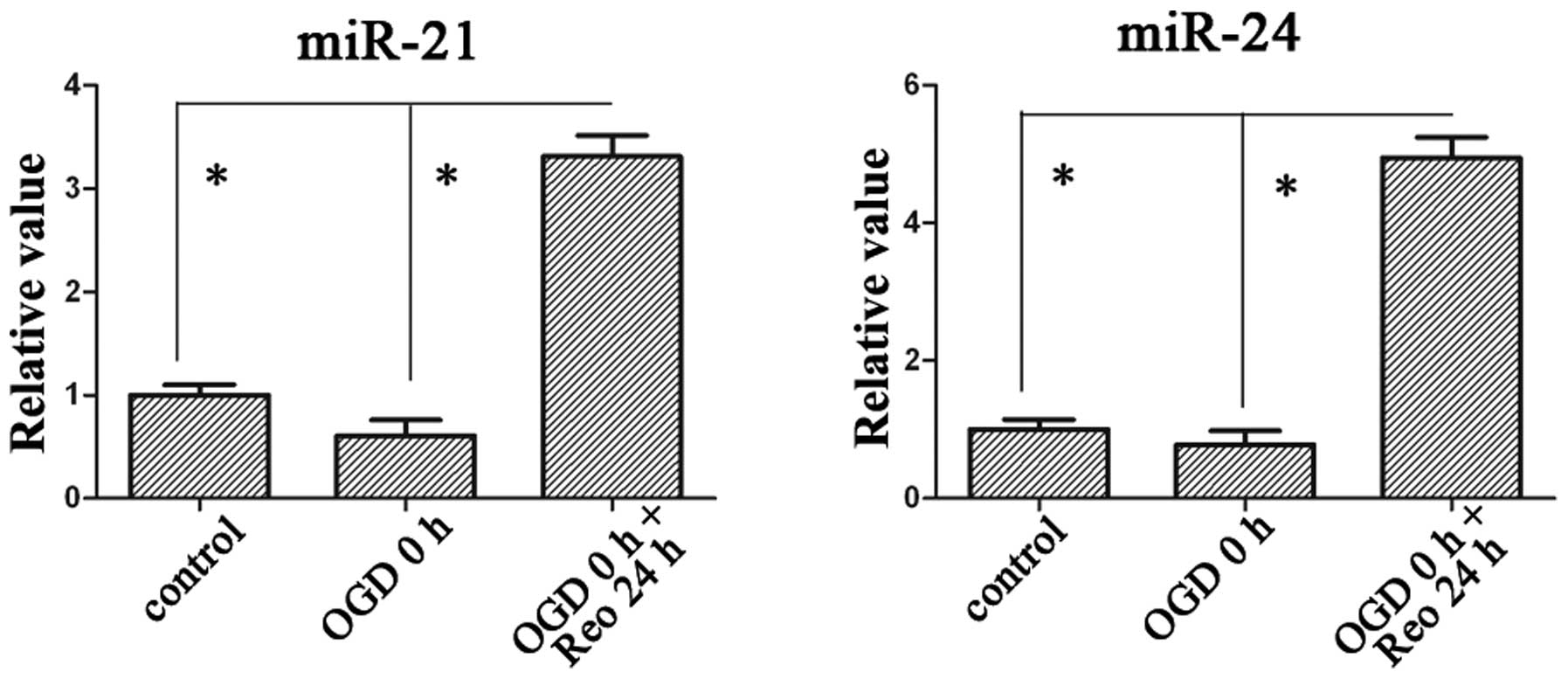
miR-21 and miR-24 are upregulated in N2A
following OGD and reoxygenation
In the present study, the change in plasma levels of
miR-21 and miR-24 were observed, we then investigated their change
in cells. The expression levels of miR-21 and miR-24 were
upregulated ~3.3- and 4.9-fold, respectively, when the recovery
time persisted up to 24 h following 3 h of OGD (Fig. 2).
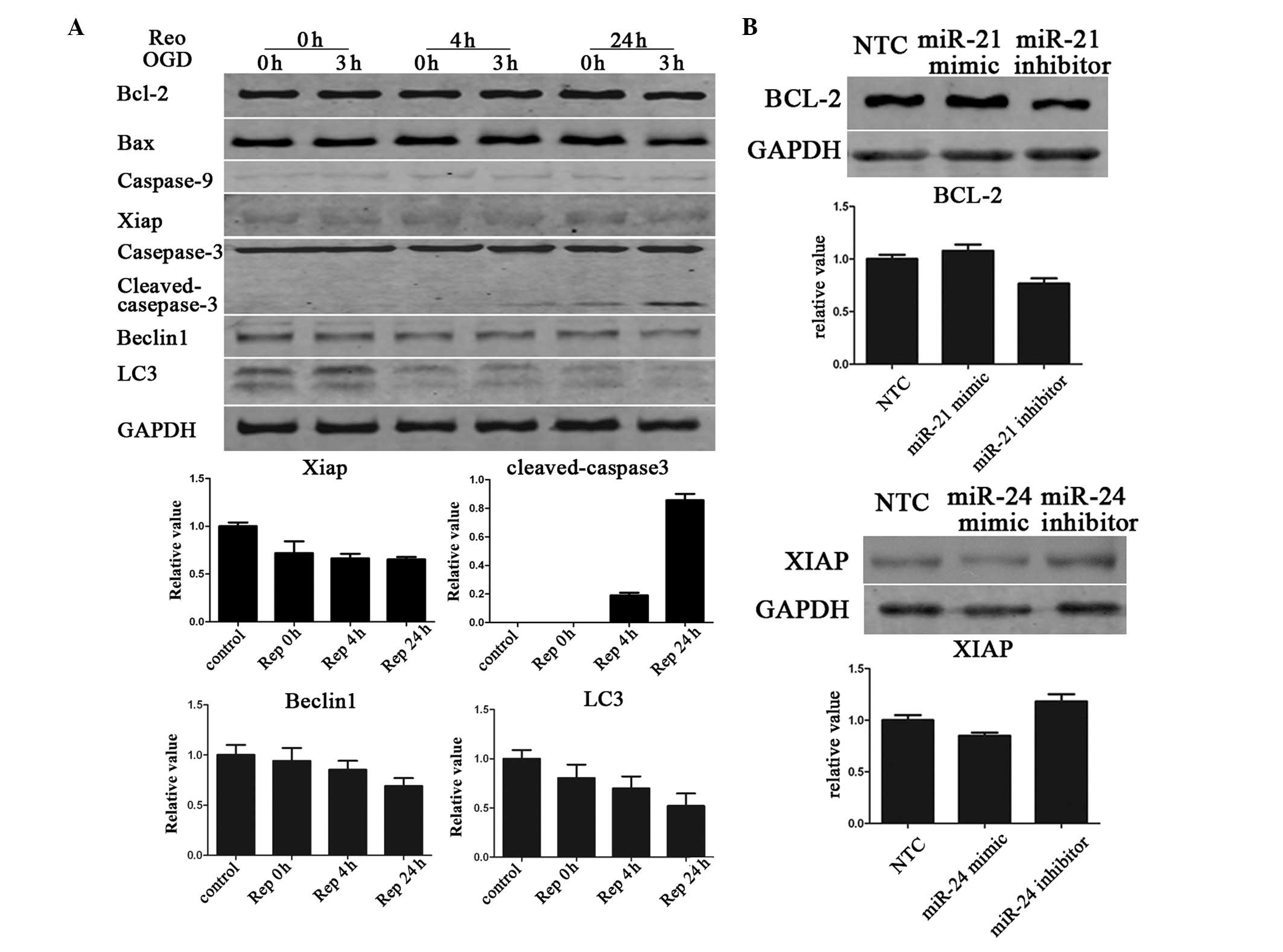
Apoptosis of N2A culture cell is enhanced
while autophagy is downregulated following OGD
Caspase-3 cleavage is an indicator of apoptosis. OGD
exposure for 3 h does not result in visible caspase-3 cleavage, but
the cleaved product became visible following 24 h of reoxygenation.
The expression of antiapoptotic protein XIAP was downregulated as
the recovery time prolonged, and the expression of LC3-II and
Beclin1 was downregulated, implying a decrease in autophagy
(Fig. 3A.)
miR-21 enhances Bcl-2 expression, while
miR-24 suppresses XIAP expression
miR-21 targets the Bcl-2 gene, an important
anti-apoptotic protein (11,12).
In the present study, it was observed that ectopic expression of
miR-21 moderately increases the Bcl-2 protein level in N2A cells.
Ectopic expression of miR-24 also moderately decreases the
expression levels of XIAP (Fig.
3B).
Discussion
The complex and intricate signaling pathways
activated by cerebral ischemia are not completely understood, but
recent studies have implicated miRNAs as important in the
pathological processes that cause ischemic injury (4,7,9,10).
miRNA has been identified as a circulating biomarker in numerous
diseases (2,19–21),
including ischemic stroke (3,4), but
whether miR-21 and miR-24 may be utilized as diagnostic biomarkers
in cerebral ischemia, has not been investigated.
The results from the present study demonstrated that
the differences in plasma miR-21 and miR-24 between ACI and control
were statistically significant (P<0.05; Fig. 1A). It was identified that miR-21
and miR-24 were positively correlated, which suggests their release
may be stimulated at the time of ischemic insult, or occur
following the resultant cerebral injury (Fig. 1B).
It is well established that the integrity of the
blood-brain barrier (BBB) is disrupted in stroke (22), which provides an opportunity for
miRNA to be released into the plasma. Stroke therapeutics is
currently limited by a lack of accurate and reliable blood
biomarkers, the identification of which would facilitate early
diagnosis and risk prediction. The results from the present study
and others alike, have identified that miR-21 and miR-24 may have
opposing effects. So, we hypothesized that the ratio of
miR-21/miR-24 may be more useful as an indicator of ACI, than just
one miRNA alone. However, we identified a positive correlation
between miR-21 and miR-24 (Fig.
1B) and a negative correlation between miR-21 and miR-24 with
the NIHSS score on first day onset (Fig. 1C), but not the ratio of
miR-21/miR-24 (data not shown).
Following this, we continued by investigating the
role of these two miRNAs in ischemic stroke. The results
demonstrated that miR-21 and miR-24 in the N2A culture cell were
upregulated 3.3- and 4.9-fold, respectively, at the 24 h recovery
time point following 3 h of OGD. Their change may have important
role in the OGD and reoxygenation injury of N2A cells.
Since miR-24 inhibits apoptosis in mouse
cardiomyocytes, in part, by direct repression of Bim (8), we investigated whether elevated
miR-24 may repress Bim and have a protective role in CI. However,
the results identified that the expression of Bim in N2A culture
cells was too low to be detected by western blot analysis (data not
shown). This result suggests that Bim may have limited involvement
in OGD and reoxygenation injury of N2A cells.
miR-23a, encoded by the same cluster as miR-24,
which regulates XIAP, contributes to sex differences in response to
cerebral ischemia (9). XIAP is
also a target of miR-24 (23), so
the present study aimed to investigate the role of XIAP in our OGD
model and whether miR-24 may be important in CI by directly
targeting XIAP. It was identified that the expression of XIAP was
reduced during the recovery time extension following 3 h OGD. The
expression of XIAP was downregulated following the addition of an
miR-24 mimic to the N2A cells, compared with application of an
miR-24 inhibitor and NTC. These data indicated that miR-24 may be a
pro-apoptotic factor during cerebral ischemia.
Bcl-2, an important anti-apoptotic protein within
the intrinsic apoptotic pathway, is one of the potential targets of
miR-21. However, it has been identified that miR-21 positively
regulated the expression of Bcl-2, which contradicted our existing
understanding of the miRNA (11,12).
miR-21 is known to be a strong antiapoptotic factor (24, 25)
and is neuroprotective against ischemic neuronal death by
downregulating FASLG (7). Our
results revealed that miR-21 expression was elevated in N2A
following 24 h of reoxygenation. Previously, Yin et al
identified that miR-497 promoted ischemic neuronal death by
negatively regulating antiapoptotic proteins, Bcl-2 and Bcl-w, and
the protein levels of Bcl-2 and Bcl-w were significantly decreased
following OGD (10). By contrast,
we did not observe any significant changes in Bcl-2 following OGD.
However, the expression of Bcl-2 was slightly increased after
adding the miR-21 mimic to the N2A cells, compared with the miR-21
inhibitor and NTC. These data indicate that miR-21 may be a
protective and anti-apoptotic factor in cerebral ischemia. The
results from the present study are consistent with the data from a
previous study (7). These findings
enrich our understanding of the impact of miR-21 and miR-24
expression in ACI, and the mechanisms underlying this effect.
Therefore, miR-21 and miR-24 may be an attractive therapeutic
molecule for the treatment of stroke
In conclusion, the present study identified that
miR-21 and miR-24 have different effects in the pathogenesis of CI,
and may be possible plasma markers in early stage of ACI. These
miRNAs are therefore potential therapeutic targets for the
development of novel diagnostic and treatment strategies in CI.
Acknowledgements
This study was supported by the National Key
Technology R&D Program (2012BAI37B01).
References
|
1
|
Rink C and Khanna S: MicroRNA in ischemic
stroke etiology and pathology. Physiol Genomics. 43:521–528. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Laterza OF, Lim L, Garrett-Engele PW, et
al: Plasma MicroRNAs as sensitive and specific biomarkers of tissue
injury. Clin Chem. 55:1977–1983. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weng H, Shen C, Hirokawa G, et al: Plasma
miR-124 as a biomarker for cerebral infarction. Biomed Res.
32:135–141. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zeng L, Liu J, Wang Y, et al: MicroRNA-210
as a novel blood biomarker in acute cerebral ischemia. Front Biosci
(Elite Ed). 3:1265–1272. 2011.PubMed/NCBI
|
|
5
|
Tan KS, Armugam A, Sepramaniam S, et al:
Expression profile of MicroRNAs in young stroke patients. PLoS One.
4:e76892009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jeyaseelan K, Lim KY and Armugam A:
MicroRNA expression in the blood and brain of rats subjected to
transient focal ischemia by middle cerebral artery occlusion.
Stroke. 39:959–966. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Buller B, Liu X, Wang X, et al:
MicroRNA-21 protects neurons from ischemic death. FEBS J.
277:4299–4307. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Qian L, Van Laake LW, Huang Y, Liu S,
Wendland MF and Srivastava D: miR-24 inhibits apoptosis and
represses Bim in mouse cardiomyocytes. J Exp Med. 208:549–560.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Siegel C, Li J, Liu F, Benashski SE and
McCullough LD: miR-23a regulation of X-linked inhibitor of
apoptosis (XIAP) contributes to sex differences in the response to
cerebral ischemia. Proc Natl Acad Sci USA. 108:11662–11667. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yin KJ, Deng Z, Huang H, et al: miR-497
regulates neuronal death in mouse brain after transient focal
cerebral ischemia. Neurobiol Dis. 38:17–26. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shi L, Chen J, Yang J, Pan T, Zhang S and
Wang Z: MiR-21 protected human glioblastoma U87MG cells from
chemotherapeutic drug temozolomide induced apoptosis by decreasing
Bax/Bcl-2 ratio and caspase-3 activity. Brain Res. 1352:255–264.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dong J, Zhao YP, Zhou L, Zhang TP and Chen
G: Bcl-2 upregulation induced by miR-21 via a direct interaction is
associated with apoptosis and chemoresistance in MIA PaCa-2
pancreatic cancer cells. Arch Med Res. 42:8–14. 2011. View Article : Google Scholar
|
|
13
|
McDonald JS, Milosevic D, Reddi HV, Greb
SK and Algeciras-Schimnich A: Analysis of circulating microRNA:
preanalytical and analytical challenges. Clin Chem. 57:833–840.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Devaux Y, Vausort M, Goretti E, et al: Use
of circulating microRNAs to diagnose acute myocardial infarction.
Clin Chem. 58:559–567. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen C, Ridzon DA, Broomer AJ, et al:
Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic
Acids Res. 33:e1792005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen X, Ba Y, Ma L, et al:
Characterization of microRNAs in serum: a novel class of biomarkers
for diagnosis of cancer and other diseases. Cell Res. 18:997–1006.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yao R, Ma Y, Du Y, et al: The altered
expression of inflammation-related microRNAs with microRNA-155
expression correlates with Th17 differentiation in patients with
acute coronary syndrome. Cell Mol Immunol. 8:486–495. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kroh EM, Parkin RK, Mitchell PS and Tewari
M: Analysis of circulating microRNA biomarkers in plasma and serum
using quantitative reverse transcription-PCR (qRT-PCR). Methods.
50:298–301. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fichtlscherer S, Zeiher AM and Dimmeler S:
Circulating microRNAs: biomarkers or mediators of cardiovascular
diseases? Arterioscler Thromb Vasc Biol. 31:2383–2390. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lorenzen JM and Thum T: Circulating and
urinary microRNAs in kidney disease. Clin J Am Soc Nephrol.
7:1528–1533. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schwarzenbach H, Hoon DS and Pantel K:
Cell-free nucleic acids as biomarkers in cancer patients. Nat Rev
Cancer. 11:426–437. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Doyle KP, Simon RP and Stenzel-Poore MP:
Mechanisms of ischemic brain damage. Neuropharmacology. 55:310–318.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xie Y, Tobin LA, Camps J, et al:
MicroRNA-24 regulates XIAP to reduce the apoptosis threshold in
cancer cells. Oncogene. 32:2442–2451. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Frankel LB, Christoffersen NR, Jacobsen A,
Lindow M, Krogh A and Lund AH: Programmed cell death 4 (PDCD4) is
an important functional target of the microRNA miR-21 in breast
cancer cells. J Biol Chem. 283:1026–1033. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Papagiannakopoulos T, Shapiro A and Kosik
KS: MicroRNA-21 targets a network of key tumor-suppressive pathways
in glioblastoma cells. Cancer Res. 68:8164–8172. 2008. View Article : Google Scholar : PubMed/NCBI
|