1. Introduction
The components of the insulin-like growth factor
(IGF) system, include IGFs (IGF-I and IGF-II), type 1 and type 2
IGF receptors, a family of six secreted IGF-binding proteins
(IGFBPs) and IGFBP proteases (1).
The IGFs are single-chain mitogenic polypeptides, structurally
similar to proinsulin, that function in an autocrine/paracrine
manner and also as classical hormones. The two IGF receptors are
structurally and functionally unrelated. IGF ligand signaling is
mediated by IGF-1R, which is a transmembrane glycoprotein with
tyrosine kinase activity (2).
IGF-2R is a single-chain protein without kinase activity (3). IGF-1R binds IGF-I with up to 20-fold
higher affinity than for IGF-II, while IGF-2R strongly binds
IGF-II, but barely recognizes IGF-I (2,3). In
biological fluids, IGFs are usually bound by members of the
secreted IGFBP family, of which the exact role remains unknown.
IGFBPs may serve as transport serum proteins, as IGF presentation
molecules to the IGF receptors, as molecules prolonging IGF
half-life or as a means for tissue-specific IGF localization. In
addition, IGFBPs are considered to mediate IGF-independent actions
via their own receptors (1). The
Igf2 and Igf2r genes are imprinted, expressed in a
monoallelic manner depending on parental legacy. In the murine
embryo, only the paternal Igf2 allele is expressed, while
only the maternal Igf2r allele is expressed (4). However, subsequent to birth,
Igf2 expression becomes biallelic in certain tissues, for
example, in the liver (5). The
present review focuses on the role of IGF-I in fetal growth and
development, paying particular attention to skeletal
development.
2. Role of IGFs in fetal growth
In initial IGF studies, the predominant roles of
IGF-I and -II in fetal growth were elucidated by abundant but
largely indirect evidence. IGFs were shown to act as proliferation
and differentiation factors in cultured fetal cells (6–8) and
preimplantation embryos (9), and
were demonstrated to be secreted by cultured fetal cells and
explants in vitro (10–11).
Direct evidence of the importance of IGFs and IGF
receptors in the regulation of embryonic and early postnatal growth
was provided by a series of studies using gene knockout, analyzing
the phenotypes manifested by mutations, alone or in combination
(4). Igf2(−/−) nullizygotes
and heterozygous mice carrying a paternally derived mutated
Igf2 gene were phenotypically indistinguishable (12). The mice were viable dwarfs with a
birth weight 60% that of normal. Ablation of the Igf1 gene
(Igf1 nullizygotes) resulted in a similar reduction of fetal
growth (13) contradicting the
prevailing hypothesis that IGF-II was the predominant mediator of
fetal growth. Furthermore, the growth deficiency of the Igf1
mutants became evident at mouse embryonic day E13.5, when the size
of the mutant embryos was ~90% that of normal size, subsequent to
which the Igf1(−/−) embryos continued to grow at a slower
rate, thus the mice were ~60% of normal size at the end of
gestation (14). Surviving mutants
continued to grow postnatally at a retarded rate, resulting in
gaining only 30% of normal body weight as adults (14). This is in contrast to the normal
birth weights observed in mice with GH-deficiency or GH-resistance
(15,16) and suggests that in prenatal mice,
IGF-I is secreted independently of GH. Nevertheless, evidence
indicates that GH acts as a local growth, differentiation and cell
survival factor in the embryo, independent of IGF-I (17). Igf1r nullizygotes exhibited
an even greater reduction in birthweight (45% of normal) and died
immediately following birth (13).
The proposed underlying mechanism for growth retardation of
Igf1 knock-out mice is that IGF-I and -II are not mitogenic
per se. Deletion of Igf1 is suggested to lead to
elongation of cell cycle time, resulting in fewer proliferation
events during the same period and the generation of fewer cells
than those required for the completion of embryonic development. In
addition, evidence provided by Walenkamp et al (18) and Bhakta et al (19) show that the influence of IGF-1 on
fetal growth is dose-related.
3. Expression levels of IGF genes and
proteins in fetal serum and tissues
The two IGFs have been detected in the fetal plasma
early in gestation in the majority of animal species investigated
thus far (20–22), with plasma concentrations of IGF-II
found to be several fold higher than those of IGF-I (20,22).
Notably, high IGF-II concentrations in fetal serum were
demonstrated to decline within days following birth (20,23),
while serum concentrations of IGF-I appeared to be low in the fetus
and rise in the immediate postnatal period, primarily as a result
of the onset of GH-stimulated IGF-I production by the liver
(20,24).
In accordance with the findings regarding plasma
concentrations of IGF-II, the majority of studies reported higher
abundance of Igf2 mRNA in fetal tissues compared with adult
tissues (25). This raised the
suggestion that IGF-II is the IGF that mediates growth and
differentiation in developing fetal tissues. However, while IGF-II
was revealed to be more abundant than IGF-I within the conceptus
(serum and tissues), IGF-I was most closely associated with fetal
growth in the majority of species. Thus, the plasma concentration
of IGF-I, but not IGF-II, was found to correlate positively with
fetal size and length, as well as birth and placental weight in
humans (26–29). Alterations in the plasma or serum
concentrations of IGF-I and IGFBP-1 and -3 have been identified in
pregnancies complicated by preeclampsia and intrauterine growth
restriction, where placental function is inadequate and fetal
growth reduced (30–33). In such complicated pregnancies, the
placental expression levels of IGF-I and IGFBP-1 are also decreased
(34,35).
Since serum concentrations may not reflect the
production of peptides in specific tissues, several studies have
attempted to detect the expression levels of Igf genes
and/or peptides in vivo. Using reverse
transcription-polymerase chain reaction (RT-PCR), transcripts of
Igf and Igf receptor genes were detected in the fetal
tissues of various species between the earliest stage of
pre-implantation and the final phase of tissue maturation (36–39),
while sensitive hybridization methods have shown that Igf
gene expression was present in almost all human and rodent fetal
tissues (40), including the
liver, pancreas and osteochondrous tissue.
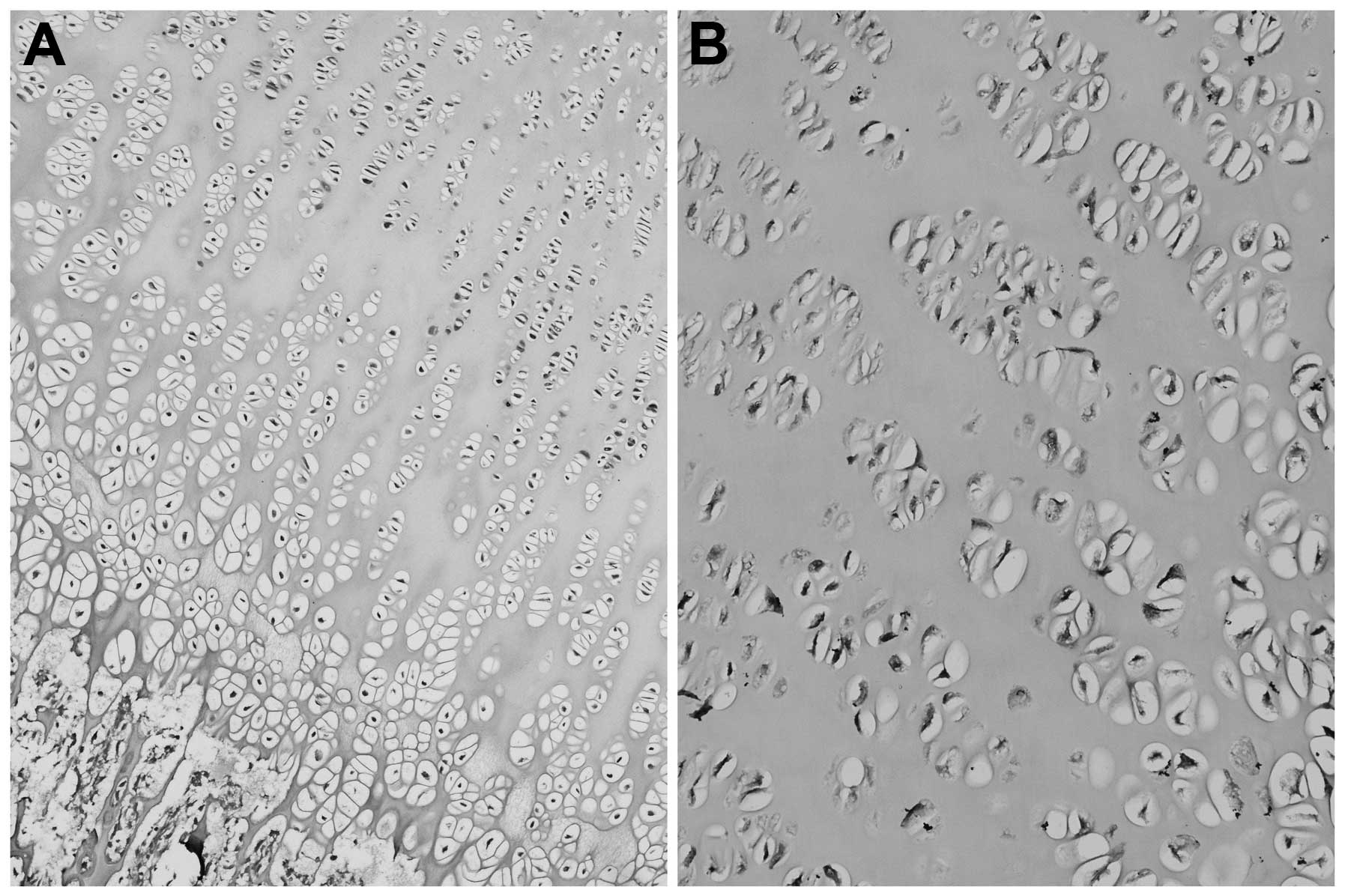
Previous studies regarding the distribution of IGFs
in the bones of piglets and mice, revealed localization within the
growth plate (41,42). Igf1 and Igf2 mRNA was
expressed throughout all zones, albeit Igf1 less
extensively. Immunohistochemical techniques also revealed the
expression of IGFs within the resting zone, the hypertrophic zone
and the proliferative zone of the growth plate (41) (Fig.
1). Additionally, with the use of RT-PCR, IGFs were also
detected within the perichondrium and metaphyseal bone in rats
(43).
4. IGF-I in limb morphogenesis
During mammalian embryogenesis, growth factors are
important not only in cellular proliferation and differentiation
but also in morphogenesis. The developing limb constitutes an
attractive model of tissue morphogenesis. At the end of week 4 of
gestation, the developing limb buds become visible as outpocketings
from the ventrolateral body wall. Initially, the limb buds consist
of a mesenchymal core derived from the lateral plate mesoderm that
forms the bones and connective tissues of the limb, covered by a
layer of ectoderm. The ectoderm at the distal border of the limb
thickens and forms the apical ectodermal ridge (AER) (44). This ridge exerts an inductive
influence on the adjacent mesenchyme, causing the mesenchyme to
remain as a population of undifferentiated, rapidly proliferating
cells, the progressing zone. As the limbs grow, cells farther from
the influence of the AER begin to differentiate into cartilage and
muscle. In this manner, development of limb proceeds
proximodistally (44). Fingers and
toes are formed when cell death in the AER separates the ridge into
five parts. The zone of polarizing activity (ZPA) is an additional
signaling region at the posterior margin of the limb mesenchyme
that controls the antero-posterior patterning of the limb (45).
While the external shape is being established, the
mesenchyme in the buds begins to condense and differentiate into
chondrocytes. By week 6 of development, the first hyaline cartilage
models, foreshadowing the bones of the extremities, are formed by
these chondrocytes. Ossification of the bones of the extremities,
endochondral ossification, begins by the end of the embryonic
period. Primary ossification centers are present in all long bones
of the limb by week 12 of development (44).
Several studies have demonstrated the predominant
role of the IGFs in limb development. IGF-I has been demonstrated
to stimulate proliferation of dissociated limb mesenchymal cells
(46), isolated human fetal
chondocytes (6), and explanted
limb buds of rat and chicken embryos in vitro (47,48).
Other studies, using in situ hybridization and
immunohistochemistry, have demonstrated that IGF-I and its receptor
(IGF-1R) are expressed in vivo by the sub ridge mesodermal
cells of the developing rat and chicken limb buds (49–52),
while in mouse embryos IGF-I has also been detected in the progress
zone (53), suggesting that IGF-I
may be involved in promoting the proliferation and outgrowth of the
limb mesoderm in response to the AER or ZPA regions. Furthermore,
several studies have revealed Igf1 transcripts in the
condensing central core of mouse and chicken limbs (52,53),
which implicates IGF-I in the regulation of chondrogenic
differentiation. However, other studies did not detect Igf1
transcripts, and reported only Igf2 and Igf1r
transcripts in the undifferentiated mesenchymal condensations and
differentiated chondrocyte precursors in murine fetus
chondrogenesis (54), verifying
the results of in vitro experiments in limb organ cultures
(47,48,52).
Igf1 transcripts have been reported to be present in the
osteoblast, osteo- and chondoclasts and nascent matrix of the long
bones of developing chicken and mouse limbs, a location consistent
with a potential role for IGF-I in endochondral bone formation
(49). Notably, during the
outgrowth and patterning of the limbs, IGF-I has been identified in
mesoderm regions that undergo programmed cell death, including the
interdigital zone in mouse and chicken embryos (52,53,55).
Therefore, IGF-1 is implicated in all activities (proliferation,
differentiation and apoptosis) essential for proper limb
morphogenesis.
Notably, no knock-outs of any IGF-axis member to
date have been reported to result in defects in limb initiation,
outgrowth or patterning. Thus, although Igf1(−/−) and
Igf2(−/−) mutants exhibited growth impairment, only marginal
ossification retardation occurred, and this did not exceed one
embryonic day (12,13). However, postnatal comparisons of
wild-type and surviving Igf1(−/−) mutants revealed the rate
of long bone ossification to be greatly reduced in the mutants
(14). Ablation of the
Igf1r gene resulted in a greater delay in the appearance of
the ossification centers in facial and cranial bones (lag of ~2
embryonic days), and ossification of the interparietal bone
exhibited an even longer delay (~4 days) (13). In support of these findings, the
intravascular infusion of recombinant IGF-I in late gestation of
fetal sheep resulted in no change in the lengths of the fetus and
long bones. However, a rise in skeletal maturation was observed, as
assessed by the acceleration of the appearance of epiphysial
centers and the increase in cross-sectional areas of the bones
(56). Furthermore, overexpression
of IGF-I in mice resulted in disproportionate overgrowth of certain
organs but no increase in the length of long bones (57). These observations indicate the
existence of redundant mechanisms for the developmental processes
of limb morphogenesis, including chondro- and osteogenesis, and/or
compensatory actions of IGF-axis members. In this regard, Dealy and
Kosher (58) observed that insulin
mimics the effects of IGF-I in promoting AER induction and limb
outgrowth in vitro, and Messiano et al (59) demonstrated that hypophysectomized
lamb fetuses with normal plasma concentrations of IGF-I and IGF-II
exhibited delayed osseous maturation, which was restored by
thyroxine administration. These observations suggest that IGFs are
not the sole trophic factors involved in fetal skeletal
development. More recent studies, using novel methods to visualize
and quantify differences in the structure and mineral density of
fetal bones in Igf1(−/−) knock-out mice compared with bones
in wild-type mice, report hypomineralization and differences in
bone microstructure, possibly representing impaired remodeling
activity in the absence of IGF-I (60,61).
5. IGF-I genetic disorders in humans
In the last few years, reports of patients with
genetic defects in various components of the IGF-axis have
broadened knowledge regarding the role of IGFs in intra-uterine
growth and development. The first human case concerning a patient
with a homozygous partial deletion (exons 4 and 5) of the
Igf1 gene was described by Woods et al (62). This mutation was manifested by
severe intrauterine growth retardation [birth weight −3.9 standard
deviations (SD), birth length −5.4 SD and microcephalia] and
dysmorphic features (micrognathia and bilateral clinodactyly), in
addition to postnatal growth failure, including delayed bone age
and severe osteopenia. Although the growth impairment of the
Igf1 null patient was relatively more severe than that of
the Igf1 knock-out mice (13), the overall phenotypic features were
similar.
Since then, a variety of molecular defects in the
Igf1 gene have been reported, including homozygosity for a
missense mutation (18), another
missense mutation with a milder phenotype (63) and a nucleotide substitution
(polymorphism) (64), which was
later found to also occur in healthy controls (65). In 2010, van Duyvenvoorde et
al (66), described a case of
heterozygosity for a frameshift mutation characterized by short
stature and microcephaly. Later, using molecular methods, the same
group concluded that the short stature of the patients cannot be
attributed exclusively to the Igf1 gene defect but to the
combination of the Igf1 gene with other factors, including
placental IGF-I insufficiency and other genetic factors (67). In any case, severe pre- and
postnatal growth impairment, microcephaly, dysmorphic features,
retarded skeletal maturation, deaf-mutism and mental retardation,
though variable in density, appear to be common characteristics of
patients with Igf1 gene molecular defects.
In humans, the potential lethal effect from total
loss of IGF-1R may explain why only heterozygous mutations in the
Igf1r gene have been reported to date. Thus, several cases
of either heterozygosity for Igf1r gene mutations (68–75)
or Igf1r gene haploinsufficiency (loss of the distal long
arm of chromosome 15q26) have been reported (76–79).
All patients exhibit a similar phenotype to patients with mutations
in the Igf1 gene. However, compared with patients exhibiting
heterozygosity for Igfr1 mutations (Igfr1
haploinsufficiency), patients with loss of the Igf1r gene
tend to have more prominent phenotypic abnormalities, with greater
dysmorphic features, an increased delay in motor development and
impaired psychosocial skills. The extent to which these features
reflect the loss of contiguous genes on chromosome 15 is uncertain.
The heterogeneity in the clinical phenotypes of patients with
molecular defects in the Igf1 or Igf1r genes may
suggest variability in the degree of functional loss of the
IGF-I/IGF-1R interaction and/or the involvement of other genetic or
environmental factors.
6. Conclusion
The two IGFs and the main IGF receptor IGF-1R are
indisputably important in embryonic and fetal growth and
development, as indicated by in vitro findings, in
vivo experiments with knock-out mice and case reports of
patients with molecular defects in the IGF-axis members. Although
IGF-II is more abundantly expressed in the serum and tissues of the
conceptus than IGF-I, IGF-I appears to be more closely associated
with fetal growth in the majority of species. IGF-I is generally
considered to affect fetal growth in a dose-related manner,
independently of GH. However, controversy remains surrounding the
data from in vitro and in vivo observations, and the
exact role of IGFs, as pertains to the prenatal development of the
skeleton, remains uncertain. Further investigation is required in
fetuses with impaired skeletal development, in the context of fetal
growth restriction or skeletal dysplasia, in order to elucidate the
role of the IGFs in fetal growth and skeletal development.
References
|
1
|
Jones JI and Clemmons DR: Insulin-like
growth factors and their binding proteins: biological actions.
Endocr Rev. 16:3–34. 1995.PubMed/NCBI
|
|
2
|
LeRoith D, Werner H, Beitner-Johnson and
Roberts CT Jr: Molecular and cellular aspects of the insulin-like
growth factor I receptor. Endocr Rev. 16:143–163. 1995. View Article : Google Scholar
|
|
3
|
Kornfeld S: Structure and function of the
mannose-6-phosphate/insulin-like growth factor II receptors. Annu
Rev Biochem. 61:307–330. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Efstratiadis A: Genetics of mouse growth.
Int J Dev Biol. 42:955–976. 1998.
|
|
5
|
Davies SM: Developmental regulation of
genomic imprinting of the Igf2 gene in human liver. Cancer Res.
54:2560–2562. 1994.PubMed/NCBI
|
|
6
|
Vetter U, Zapf J, Heit W, et al: Human
fetal and adult chondrocytes. Effect of insulin-like growth factors
I and II, insulin, and growth hormone on clonal growth. J Clin
Invest. 77:1903–1908. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bhaumick B and Bala RM: Differential
effects of insulin-like growth factors I and II on growth,
differentiation and glucoregulation in differentiating chondrocyte
cells in culture. Acta Endocrinol (Copenh). 125:201–211.
1991.PubMed/NCBI
|
|
8
|
Lorenzo M, Valverde AM, Ternel T and
Benito M: IGF-I is a mitogen involved in differentiation-related
gene expression in fetal rat brown adipocytes. J Cell Biol.
123:1567–1575. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Harvey MB and Kaye PL: Insulin-like growth
factor-1 stimulates growth of mouse preimplantation embryos in
vitro. Mol Reprod Dev. 31:195–199. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Canalis E, McCarthy T and Centrella M:
Isolation and characterization of insulin-like growth factor I
(somatomedin-C) from cultures of fetal rat calvariae.
Endocrinology. 122:22–27. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
D’Ercole AJ, Applewhite GT and Underwood
LE: Evidence that somatomedin is synthesized by multiple tissues in
the fetus. Dev Biol. 75:315–328. 1980.PubMed/NCBI
|
|
12
|
DeChiara TM, Efstratiadis A and Robertson
EJ: A growth-deficiency phenotype in heterozygous mice carrying an
insulin-like growth factor II gene disrupted by targeting. Nature.
345:78–80. 1990. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu J-P, Baker J, Perkins AS, Roberson EJ
and Efstratiadis A: Mice carrying null mutations of the genes
encoding insulin-like growth factor I (Igf-1) and type 1 IGF
receptor (Igf1r). Cell. 75:59–72. 1993.
|
|
14
|
Baker J, Liu J-P, Robertson EJ and
Efstratiadis A: Role of insulin-like growth factors in embryonic
and postnatal growth. Cell. 75:73–82. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wajnrajch MP, Gertner JM, Harbison MD,
Chua SC Jr and Leibel RL: Nonsense mutation in the human growth
hormone-releasing hormone receptor causes growth failure analogous
to the little (lit) mouse. Nat Genet. 12:88–90. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Savage MO, Blum WF, Ranke MB, et al:
Clinical features and endocrine status in patients with growth
hormone insensitivity (Laron syndrome). J Clin Endocrinol Metab.
77:1465–1471. 1993.PubMed/NCBI
|
|
17
|
Sanders EJ and Harvey S: Growth hormone as
an early embryonic growth and differentiation factor. Anat Embryol
(Berl). 209:1–9. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Walenkamp MJ, Karperien M, Pereira AM, et
al: Homozygous and heterozygous expression of a novel insulin-like
growth factor-I mutation. J Clin Endocrinol Metab. 90:2855–2864.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bhakta KY, Marlin SJ, Shen JJ and
Fernandes CJ: Terminal deletion of chromosome 15q26.1: case report
and brief literature review. J Perinatol. 25:429–432. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gluckman PD and Butler JH:
Parturition-related changes in insulin-like growth factors-I and
-II in the perinatal lamb. J Endocrinol. 99:223–232. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Holland MD, Hossner KL, Williams SE,
Wallace CR, Niswender GD and Odde KG: Serum concentrations of
insulin-like growth factors and placental lactogen during gestation
in cattle. Domest Anim Endocrinol. 14:231–239. 1997. View Article : Google Scholar
|
|
22
|
Lee CY, Chung CS and Simmen FA: Ontogeny
of the porcine insulin-like growth factor system. Mol Cell
Endocrinol. 93:71–80. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Moses AC, Nissley SP, Short PA, et al:
Increased levels of multiplication-stimulating activity, an
insulin-like growth factor, in fetal rat serum. Proc Natl Acad Sci
USA. 77:3649–3653. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Daughaday WH, Parker KA, Borowsky S,
Trivedi B and Kapadia M: Measurement of somatomedin-related
peptides in fetal, neonatal, and maternal rat serum by insulin-like
growth factor (IGF) I radioimmunoassay, IGF-II radioreceptor assay
(RRA), and multiplication-stimulating activity RRA after
acid-ethanol extraction. Endocrinology. 110:575–581. 1982.
View Article : Google Scholar
|
|
25
|
Lund PK, Moats-Staats BM, Hynes MA, et al:
Somatomedin-C/insulin-like growth factor-I and insulin-like growth
factor-II mRNAs in rat fetal and adult tissues. J Biol Chem.
261:14539–14544. 1986.PubMed/NCBI
|
|
26
|
Gluckman PD, Johnson-Barrett JJ, Butler
JH, Edgar BW and Gunn TR: Studies of insulin-like growth factor-I
and -II by specific radioligand assays in umbilical cord blood.
Clin Endocrinol (Oxf). 19:405–413. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lassarre C, Hardouin S, Daffos F,
Forestier F, Frankenne F and Binoux M: Serum insulin-like growth
factors and insulin-like growth factor binding proteins in the
human fetus. Relationships with growth in normal subjects and in
subjects with intrauterine growth retardation. Pediatr Res.
29:219–225. 1991. View Article : Google Scholar
|
|
28
|
Verhaeghe J, Van Bree R, Van Herck E,
Laureys J, Bouillon R and van Assche FA: C-peptide, the
insulin-like growth factors-I and -II, and IGF-binding protein I in
umbilical cord serum: correlations with birthweight. Am J Obstet
Gynecol. 169:89–97. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Giudice LC, de Zegher F, Gargosky SE, et
al: Insulin-like growth factors and their binding proteins in the
term and preterm human fetus and neonate with normal and extremes
of intrauterine growth. J Clin Endocrinol Metab. 80:1548–1555.
1995.PubMed/NCBI
|
|
30
|
Sifakis S, Akolekar R, Kappou D, Mantas N
and Nicolaides KH: Maternal serum insulin like growth factor-I
(IGF-I) at 11–13 weeks in preeclampsia. Prenat Diagn. 30:1026–1031.
2010.
|
|
31
|
Sifakis S, Akolekar R, Kappou D, Mantas N
and Nicolaides KH: Maternal serum insulin-like growth factor
binding protein-1 (IGFBP-1) at 11–13 weeks in preeclampsia. Prenat
Diagn. 31:196–201. 2011.PubMed/NCBI
|
|
32
|
Sifakis S, Akolekar R, Kappou D, Mantas N
and Nicolaides KH: Maternal serum insulin-like growth factor
binding protein-3 (IGFBP-3) at 11–13 weeks in preeclampsia. J Hum
Hypertens. 26:253–258. 2012.PubMed/NCBI
|
|
33
|
Sifakis S, Akolekar R, Kappou D, Mantas N
and Nicolaides KH: Maternal serum insulin like growth factor
(IGF-I) and binding proteins IGFBP-1 and IGFBP-3 at 11–13 weeks’
gestation in pregnancies delivering small for gestational age
neonates. Eur J Obstet Gynecol Reprod Biol. 161:30–33. 2012.
|
|
34
|
Koutsaki M, Sifakis S, Zaravinos A,
Koutroulakis D, Koukoura O and Spandidos DA: Decreased placental
expression of hPGH, IGF-I and IGFBP-1 in pregnancies complicated by
fetal growth restriction. Growth Horm IGF Res. 21:31–36. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Koukoura O, Sifakis S, Zaravinos A, et al:
Aberrant methylation of IGF2 is associated with loss of imprinting
in placentas from pregnancies complicated with fetal growth
restriction. Int J Mol Med. 28:481–487. 2011.PubMed/NCBI
|
|
36
|
Watson AJ, Hogan A, Hahnel A, Wiemer KE
and Schultz GA: Expression of growth factor ligand and receptor
genes in the preimplantation bovine embryo. Mol Reprod Dev.
31:87–95. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Watson AJ, Watson PH, Arcellana-Panlilio
M, et al: A growth factor phenotype map for ovine preimplantation
development. Biol Reprod. 50:725–733. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang X, Kidder GM, Watson AJ, Schultz GA
and Armstrong DT: Possible roles of insulin and insulin-like growth
factors in rat preimplantation development: investigation of gene
expression by reverse transcription-polymerase chain reaction. J
Reprod Fertil. 100:375–380. 1994. View Article : Google Scholar
|
|
39
|
Lighten AD, Hardy K, Winston RM and Moore
GE: Expression of mRNA for the insulin-like growth factors and
their receptors in human preimplantation embryos. Mol Reprod Dev.
47:134–139. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rotwein P, Pollock KM, Watson M and
Milbrandt JD: Insulin-like growth factor gene expression during rat
embryonic development. Endocrinology. 121:2141–2144. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Schlegel W, Halbauer D, Raimann A, et al:
IGF expression patterns and regulation in growth plate
chondrocytes. Mol Cell Endocrinol. 327:65–71. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang Y, Cheng Z, Elalieh HZ, et al: IGF1R
signaling in chondrocytes modulates growth plate development by
interacting with the PTHrP/Ihhpathway. J Bone Miner Res.
26:1437–46. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Parker EA, Hegde A, Buckley M, Barnes KM,
Baron J and Nilsson O: Spatial and temporal regulation of
GH-IGF-related gene expression in growth plate cartilage. J
Endocrinol. 194:31–40. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sadler TW: Limbs. Langman’s Medical
Embryology. Lippincott Williams and Wilkins; Philadephia, PA: 12th
edition. pp. 151–152
|
|
45
|
Sifakis S, Basel D, Ianakiev P, Kilpatrick
M and Tsipouras P: Distal limb malformations: underlying mechanisms
and clinical associations. Clin Genet. 60:165–172. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kaplowitz PB, D’Ercole AJ and Underwood
LE: Stimulation of embryonic mouse limb bud mesenchymal cell growth
by peptide growth factors. J Cell Physiol. 112:353–359. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bhaumick B and Bala RM: Receptors for
insulin-like growth factors I and II in developing embryonic mouse
limb bud. Bioch Bioph Acta. 927:117–128. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Geduspan JS and Solursh M: Effects of the
mesonephros and insulin-like growth factor I on chondrogenesis of
limb explants. Dev Biol. 156:500–508. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Streck RD, Wood TL, Hsu MS and Pintar JE:
Insulin-like growth factor I and II and insulin-like growth factor
binding protein 2 RNAs are expressed in adjacent tissues within rat
embryonic and fetal limbs. Dev Biol. 151:586–596. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ralphs JR, Wylie L and Hill DJ:
Distribution of insulin-like growth factor peptides in the
developing chick embryo. Development. 109:51–58. 1990.PubMed/NCBI
|
|
51
|
Geduspan JS, Padanilam BJ and Solursh M:
Coordinate expression of IGF-I and its receptor during limb
outgrowth. Dev Dyn. 195:67–73. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mc Queeney K and Dealy CN: Roles of
insulin-like growth factor-I (IGF-I) and IGF-I binding protein-2
(IGFBP2) and -5 (IGFBP5) in developing chick limbs. Growth Horm IGF
Res. 11:346–363. 2001.PubMed/NCBI
|
|
53
|
van Kleffens M, Groffen C, Rosato RR, et
al: mRNA expression patterns of the IGF system during mouse limb
bud development determined by whole mount in situ hybridization.
Mol Cell Endocrinol. 138:151–161. 1998.PubMed/NCBI
|
|
54
|
Wang E, Wang J, Chin E, Zhou J and Bondy
CA: Cellular patterns of insulin-like growth factor system gene
expression in murine chondrogenesis and osteogenesis.
Endocrinology. 136:2741–2751. 1995.PubMed/NCBI
|
|
55
|
Allan GJ, Flint DJ, Darling SM, Geh J and
Patel K: Altered expression of insulin-like growth factor 1 and
insulin-like growth factor binding proteins 2 and 5 in the mouse
mutant Hypodactyly (Hd) correlates with sites of apoptotic
activity. Anat Embryol (Berl). 202:1–11. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lok F, Owens JA, Mundy L, Robinson JS and
Owens PC: Insulin-like growth factor I promotes growth selectively
in fetal sheep in late gestation. Am J Physiol. 270:R1148–R1155.
1996.PubMed/NCBI
|
|
57
|
Mathews LS, Hammer RE, Behringer RR, et
al: Growth enhancement of transgenic mice expressing human
insulin-like growth factor I. Endocrinology. 123:2827–2833. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Dealy CN and Kosher RA: Studies on
insulin-like growth factor-I and insulin in chick limb
morphogenesis. Dev Dyn. 202:67–79. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Mesiano S, Young IR, Baxter RC, Hintz RL,
Browne CA and Thorburn GD: Effect of hypophysectomy with and
without thyroxine replacement on growth and circulating
concentrations of insulin-like growth factors I and II in the fetal
lamb. Endocrinology. 120:1821–1830. 1987. View Article : Google Scholar
|
|
60
|
Bikle D, Majumdar S, Laib A, et al: The
skeletal structure of insulin-like growth factor I-deficient mice.
J Bone Miner Res. 16:2320–2329. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Burghardt AJ, Wang Y, Elalieh H, et al:
Evaluation of fetal bone structure and mineralization in IGF-I
deficient mice using synchrotron radiation microtomography and
Fourier transform infrared spectroscopy. Bone. 40:160–168. 2007.
View Article : Google Scholar
|
|
62
|
Woods KA, Camacho-Hübner C, Barter D,
Clark AJL and Savage MO: Insulin-like growth factor I gene deletion
causing intrauterine growth retardation an severe short stature.
Acta Paediatr Suppl. 423:39–45. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Netchine I, Azzi S, Housang M, et al:
Partial primary deficiency of insulin-like growth factor (IGF)-I
activity associated with IGF1 mutation demonstrates its critical
role in growth and brain development. J Clin Endocrinol Metab.
94:3913–3921. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Bonapace G, Concolino D, Formicola S and
Strisciuglio P: A novel mutation in a patient with insulin-like
growth factor 1 (IGF1) deficiency. J Med Genet. 40:913–917. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Coutinho DC, Coletta RR, Costa EM, et al:
Polymorphisms identified in the upstream core polyadenylation
signal of IGF1 gene exon 6 do not cause pre- and postnatal growth
impairment. J Clin Endocrinol Metab. 92:4889–4892. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
van Duyvenvoorde HA, van Setten PA,
Walenkamp MJE, et al: Short stature associated with a novel
heterozygous mutation in the insulin-like growth factor 1 gene. J
Clin Endocrinol Metab. 95:E363–E367. 2010.PubMed/NCBI
|
|
67
|
van Duyvenvoorde HA, van Doorn J, Koenig
J, et al: The severe short stature in two siblings with a
heterozygous IGF1 mutation is not caused by a dominant negative
effect of the putative truncated protein. Growth Hormone IGF Res.
21:44–50. 2011.PubMed/NCBI
|
|
68
|
Abuzzahab MJ, Schneider A, Goddard A, et
al: IGF-I receptor mutations resulting in intrauterine and
postnatal growth retardation. N Engl J Med. 349:2211–2222. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kawashima Y, Kanzaki S, Yang F, et al:
Mutation at cleavage site of insulin-like growth factor receptor in
a short-stature child born with intrauterine growth retardation. J
Clin Endocrinol Metab. 90:4679–4687. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Raile K, Klammt J, Schneider A, et al:
Clinical and functional characteristics of the human Arg59Ter
insulin-like growth factor I receptor (IGF1R) mutation: implication
for a gene dosage effect of the human IGF1R. J Clin Endocrinol
Metab. 91:2264–2271. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Walenkamp MJ, van der Kamp HJ, Pereira AM,
et al: A variable degree of intrauterine and postnatal growth
retardation in a family with a missense mutation in the
insulin-like growth factor I receptor. J Clin Endocrinol Metab.
91:3062–3070. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Inagaki K, Tiulpakov A, Rubtsov P, et al:
A familial insulin-like growth factor I receptor mutant leads to
short stature: clinical and biochemical characterization. J Clin
Endocrinol Metab. 92:1542–1548. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Kruis T, Klammt J, Galli-Tsinopoulou A, et
al: Heterozygous mutation within a kinase-conserved motif of the
insulin-like growth factor I receptor causes intrauterine and
postnatal growth retardation. J Clin Endocrinol Metab.
95:1137–1142. 2010. View Article : Google Scholar
|
|
74
|
Wallborn T, Wüller S, Klammt J, et al: A
heterozygous mutation of the insulin-like growth factor-I receptor
causes retention of the nascent protein in the endoplasmic
reticulum and results in intrauterine and postnatal growth
retardation. J Clin Endocrinol Metab. 95:2316–2324. 2010.
View Article : Google Scholar
|
|
75
|
Choi JH, Kang M, Kim GH, et al: Clinical
and functional characteristics of a novel heterozygous mutation of
the IGF1R gene and IGF1R haploinsufficiency due to terminal
15q26.2->qter deletion in patients with intrauterine growth
retardation and postnatal catch-up growth failure. J Clin
Endocrinol Metab. 96:E130–E134. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Roback EW, Barakat AJ, Dev VG, Mbikay M,
Chrétien M and Butler MG: An infant with deletion of the distal
long arm of chromosome 15 (q26.1 - qter) and loss of insulin-like
growth factor I receptor gene. Am J Med Genet. 38:74–79. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Walemkamp MJ, de Muinck Keizer-Schrama SM,
de Mos M, et al: Successful long-term growth hormone therapy in a
girl with haploinsufficiency in the insulin-like growth factor-I
receptor due to a terminal 15q26.2->qter deletion detected by
multiplex ligation probe amplification. J Clin Endocrinol Metab.
93:2421–2425. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Fang P, Schwartz ID, Johnson BD, et al:
Familial short stature caused by haploinsufficiency of the
insulin-like growth factor I receptor due to nonsense-mediated
messenger ribonucleic acid decay. J Clin Endocrinol Metab.
94:1740–1747. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Ester WA, van Duyvenvoorde HA, de Wit CC,
et al: Two short children born small for gestational age with
insulin-like growth factor 1 receptor haploinsufficiency illustrate
the heterogeneity of its phenotype. J Clin Endocrinol Metab.
94:4717–4727. 2009. View Article : Google Scholar : PubMed/NCBI
|