Introduction
Hepatocellular carcinoma (HCC) is the sixth most
prevalent neoplasm in the world, with the number of reported cases
increasing annually. The highest incidence areas are Asia and
Africa, of which 80% of cases occur in Asian countries (1). For these countries, HCC malignant
tumors are the third most common type of cancer (2). At present, surgical resection, local
ablation therapy and liver transplantation are considered
fundamental and effective treatments for HCC. However, only ~20% of
diagnosed patients benefit from surgical treatment options as the
majority of cases HCC are diagnosed in the late stages of tumor
progression or the patients have underlying symptoms of cirrhosis;
therefore, for numerous patients it is too late for surgery to be
effective. In addition, for these cases, radiotherapy and
chemotherapy are not curative and can only provide relief of
symptoms (3). It has been
speculated that HCC occurs due to the synergy of multiple gene
abnormalities. Therefore, the discovery of a key regulatory gene
for HCC carcinogenesis would provide a new target for HCC
therapies.
In 1996, death receptor 3 (DR3), a member of the
tumor necrosis factor receptor (TNFR) superfamily, was discovered.
DR3 is a type II transmembrane protein containing 417 amino acids
and its gene was found to be localized to 1p36.3. DR3, similarly to
other TNFR family proteins, has a proteolytic function of an amino
acid residue segment on a protein called the death domain in the
cytoplasm. The DR3 protein also has a high degree of homology with
TNFR-1 and Fas, allowing it to pass on apoptotic signals to
downstream pathways, initiating apoptosis (4–6). In
addition, DR3 can also activate the expression of nuclear factor
kappa-light-chain-enhancer of activated B cells (NF-κB), allowing
NF-κB to translocate to the nucleus and activate transcription
(7). DR3 was found to be primarily
distributed in lymphoid tissues, including the spleen, thymus and
peripheral blood lymphocytes (lymphocytes, natural killer cells and
macrophages) (8,9). Therefore, previous studies have
mainly focused on the involvement of DR3 in immune adjustment,
where it was reported to promote the occurrence and development of
numerous inflammatory diseases, including inflammatory bowel
disease and arthritis (10–14).
In addition, DR3 was found to be located in numerous tumor types
(15–17). Previously, it was reported that
lipeol, at the appropriate dosage, inhibited hepatocarcinoma cell
proliferation and apoptosis, as well as reduced the high expression
of DR3 (18). Therefore, it has
been hypothesized that DR3 may have an important role in the
development and progression of HCC. The aim of the present study
was to observe the effect of silencing DR3 expression on
hepatocarcinoma cell growth, apoptosis and invasion in order to
elucidate the role of DR3 in tumor development, and therefore
provide a theoretical target for HCC therapies.
Materials and methods
Cell culture
The human hepatocarcinoma cell lines Bel-7402,
SMMC-7721, Huh7 and HepG2, and normal human liver HL-7702 cells,
were purchased from the cell bank of the Chinese Academy of Science
(Shanghai, China). The cells were cultured in RPMI-1640 medium
(Gibco-BRL, Carlsbad, CA, USA) and DMEM medium (Gibco-BRL)
containing 10% fetal bovine serum (FBS; Invitrogen Life
Technologies, Carlsbad, CA, USA) in a humidified atmosphere of 5%
CO2 at 37°C.
Design and synthesis of Stealth RNA
interference (RNAi) small interfering RNA (siRNA) targeting the DR3
gene sequence
Sequences of the three synthesized oligonucleotides
were: AF026070.1_stealth_880 sense 5′-UUCUCACUGCUGUCAGGAGGUGCUA-3′
and anti-sense 5′-UAGCACCUCCUGACAGCAGUGAGAA-3′;
AF026070.1_stealth_883 sense, 5′-AUCUUCUCACUGCUG UCAGGAGGUG-3′ and
anti-sense, 5′-CACCUCCCUGAC AGCAGUGAGAAGAU-3′;
AF026070.1_stealth_888 sense, 5′-UGCAGAUCUUCUCAACUGCUGUCAGG-3′ and
anti-sense, 5′-CCUGACAGCAGUGAGAAGAUCUGCA-3′. These target sequences
were synthesized by Invitrogen Life Technologies and subjected to a
Basic Local Alignment Search Tool (www.ncbi.nlm.nih.gov/BLAST.cgi) analysis to ensure
that only the DR3 gene was targeted.
Transfection
To transfect the Stealth RNAi siRNA against DR3 into
Bel-7402 and SMMC-7721, Lipofectamine™ RNAiMAX (Invitrogen Life
Technologies) was used, and the negative control, the Stealth™ RNAi
Negative Control Duplexes (Invitrogen Life Technologies) were used.
BLOCK-It™ Alexa Fluor® Red Fluorescent Oligo (Invitrogen
Inc, USA) was used to facilitate assessment and optimize the
delivery of double-stranded RNA oligonucleotides into Bel-7402 and
SMMC-7721 cells, according to the manufacturer’s instructions.
Reverse transfection was used to deliver Stealth RNAi siRNA, Red
Fluorescent Oligo or negative control duplexes into two cell lines
as follows: Lipofectamine™ RNAiMAX complexes were prepared
according to the manufacturer’s instructions, cells were seeded at
appropriate dilutions and incubated for 24 h to reach 30–50%
confluence. The complexes were added to the cells
(5×105) and incubated for 48 and 72 h at 37°C in a
CO2 incubator until ready for gene knockdown assays.
Nucleic acid assessment
Total RNA was extracted using RNAiso™ Plus (Takara
Co., Ltd., Otsu, Japan), following transfection. First strand
complementary DNA (cDNA) synthesis and amplification were performed
using a two-step reverse transcription quantitative polymerase
chain reaction (RT-qPCR) kit (Takara Co., Ltd.) with the Rotor-Gene
6,000 (Qiagen, Hilden, Germany). The reverse transcription reaction
was performed at 37°C for 15 min followed by 85°C for 5 min in
order to inactivate reverse transcriptase. The reaction volume was
25 μl, with 12.5 μl SYBR® Premix Ex Taq™ II (2X) (Takara
Co., Ltd.).
The sequences of primers were: DR3 sense,
5′-GTGTGTCCCCAAGACACCTT-3′ and anti-sense,
5′-GTCTAGGCATGGTTGGCAGT-3′ (GenBank accession no. AF026070.1);
Casepase3 sense, 5′-GGTTCATCC AGTCGCTTTGT-3′ and anti-sense,
5′-CGGTTAACCCGGGT AAGAAT-3′ (GenBank accession no. NM_004346.3);
Caspase8 sense, 5′-CCAAATGCAAACTGGATGATGAC-3′ and anti-sense,
5′-CTCTTGTTGATTTGGGCACAG AC-3′ (GenBank accession no. NM-001228.4);
NF-κB sense, 5′-TTGTGGCCGCCTAAGTGGA-3′ and anti-sense,
5′-ACCACCTTGATCTGGGTAGCACATA-3′ (GenBank accession no. AF018253.1);
P53 sense, 5′-GGCCCACTTCACCGTACTAA-3′ and anti-sense,
5′-GTGGTTTCAAGGCCAGATGT-3′ (GenBank accession no. NM_000546.4);
apoptosis antigen-3 ligand (Apo-3L) sense,
5′-GAGGAATTCTCAGCCACTGC-3′ and anti-sense,
5′-CCCTCAGTGAACCTGGAAGA-3′ (GenBank accession no. NM_003809.2); and
β-actin sense, 5′-AGAGATGGCCACGGCTGCTT-3′ and anti-sense,
5′-ATTTGCGGTGGACGATGGAG (GenBank accession no. NM_001101.3) (Takara
Co., Ltd.).
Protein assessment
Cultured cells were lysed in lysis buffer
phenylmethylsulfonyl fluoride (Sangon Biotech, Shanghai, China),
and a protein standard curve was used to calculate the density of
total protein. Proteins were separated using 10% SDS-PAGE (Sangon
Biotech) under denaturing conditions and then transferred to
nitrocellulose membranes (Applygen Technologies, Inc., Beijing,
China). Membranes were incubated with rat anti-DR3 polyclonal
antibody (pAb), rabbit anti-TNF-related weak inducer of apoptosis
(TWEAK) pAb, rabbit anti-NF-κB pAb, rabbit Caspase3 monoclonal
antibody (mAb), rabbit Caspase8 mAb, rat anti-P53 pAb, rabbit
anti-Fas pAb or rat anti-β-actin mAb primary antibodies (all
1:1,000; Abcam, Cambridge, MA, USA), followed by incubation with
anti-mouse secondary antibody conjugated to horseradish peroxidase
(1:5,000; Amersham Biosciences, Chalfont St. Giles, UK).
Immunoreactive proteins were visualized using Poncuar S staining
solution (Sangon Biotech), developing liquid and fixing solution
(Beyotime Institute of Biotechnology, Shanghai, China), Super ECL
Plus (Applygen Technologies Inc. Beijing, China) and a Bio-Rad gel
imaging system (Bio-Rad, Hercules, CA, USA).
Cell proliferation assessment
Following reverse transfection of Bel-7402 cells
with Stealth RNAi siRNA targeting the DR3 gene or negative control
duplexes in 96-well plates, MTT (Sigma-Aldrich, St. Louis, MO, USA)
was added at 24, 48 and 72 h, in order to determine the rates of
cell proliferation. The optical density was measured using a
UR-4100 plate reader (Fisher Thermo Scientific, Waltham, MA,
USA).
Assessment of apoptosis
To determine the occurrence of apoptosis within 72 h
of transfection, cells were harvested using trypsinization
(Sigma-Aldrich) and rinsed twice with phosphate-buffered saline
(Sangon Biotech). Cells were then centrifuged (4°C, 1,000 ×g) for
10 min. Cells were resuspended in 200 μl binding buffer (Sangon
Biotech) and treated with 10 μl Annexin V-fluorescein
isothiocyanate (FITC) and 5 μl propidium iodide (PI; both
Sigma-Aldrich) for 15 min at room temperature. The rate of
apoptosis was then determined using flow cytometry (Epics-XL;
Beckman-Coulter, Shanghai, China).
Invasion assessment
Invasion assessments were performed using a 24-well
Transwell chamber (Corning Inc., Corning, NY, USA). Each Transwell
chamber was coated with 200 μl Matrigel® (Corning,
Inc.), which was pre-diluted into 100 μg/ml with 0.1% FBS). A total
of 5×105/ml cells were seeded into the pre-coated wells.
The lower parts of the chambers were filled with 200 μl RPMI 1640
medium containing 10% FBS. Following incubation for 24 h, the cells
on the upper surface were gently removed using a cotton swab, and
the filters were fixed with 95% ethanol for 30 min and stained with
0.1% hexamethylpararosaniline (both Sangon Biotech) for 15 min. The
number of cells on the lower surface of the membranes was
quantified using a microscope (LX71; Olympus Corp., Tokyo,
Japan).
Statistical analysis
Statistical analysis was performed using the SPSS
16.0 statistical software package (IBM, Armonk, NY, USA). P<0.05
was considered to indicate a statistically significant difference
between values.
Results
Content, purity and integrity of total
RNA in different cell lines
An ultraviolet spectrophotometer was used to detect
the total RNA at the optical density (OD)260/OD280 (R) from the
cell lines Huh7, SMMC7721, HepG2, Bel-7402 and HL-7702. The
R-values were 1.85, 1.86, 1.95, 1.91, and 2.01, reflecting high
levels of purity. The integrity of total RNA is shown in Fig. 1.
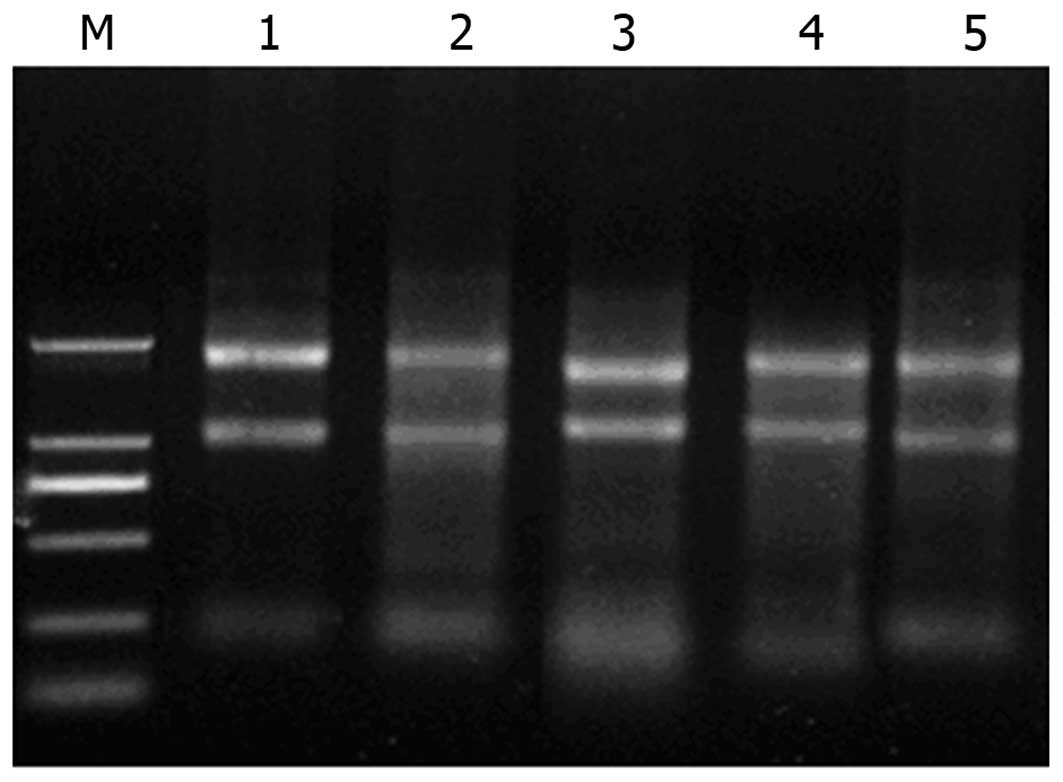 | Figure 1SDS-PAGE for total RNA analysis.
Lanes from left to right: M, DL2000 Marker; 1, Huh7; 2, SMMC7721;
3, HepG2; 4, Bel-7402; and 5, HL-7702 cells. In each
electrophoresis lane, three bands are observed, with sedimentation
coefficients of ribosomal RNA of s=28, 18 and 5, demostrating the
high integrity of total RNA. |
The expression of DR3 messenger RNA
(mRNA) in different cell lines
Following the extraction of total RNA from each cell
line, they were reverse-transcribed into cDNA. To ensure the
reliability and accuracy of the results, the target gene-specific
primers were amplified at the same time as β-actin (Figs. 2 and 3). The ΔΔCt method was used to calculate
the relative expression levels of each DR3 sample (Table I). DR3 expression at the nucleic
acid level was detected in multiple strains of hepatocarcinoma cell
lines. DR3 expression was significantly higher in hepatocarcinoma
cell lines compared to that of normal liver cells. Therefore, it
was speculated that DR3 expression may be associated with the
occurrence of liver cancer.
 | Table IExpression of DR3 in different cell
lines. |
Table I
Expression of DR3 in different cell
lines.
| Cell lines | DR3(Ct) | β-actin(Ct) | ΔCt | ΔΔCt |
2−ΔΔCt |
|---|
| HL-7702 | 19.5 | 10.54 | 8.71 | 0 | 1 |
| Hhh7 | 16.71 | 10.36 | 6.35 | −2.36 | 5.134 |
| Bel-7402 | 16.40 | 9.74 | 6.66 | −2.05 | 4.141 |
| SMMC-7721 | 16.05 | 8.97 | 7.08 | −1.63 | 3.095 |
| HepG2 | 16.03 | 9.13 | 6.90 | −1.81 | 3.506 |
Transfection efficiency and
conditions
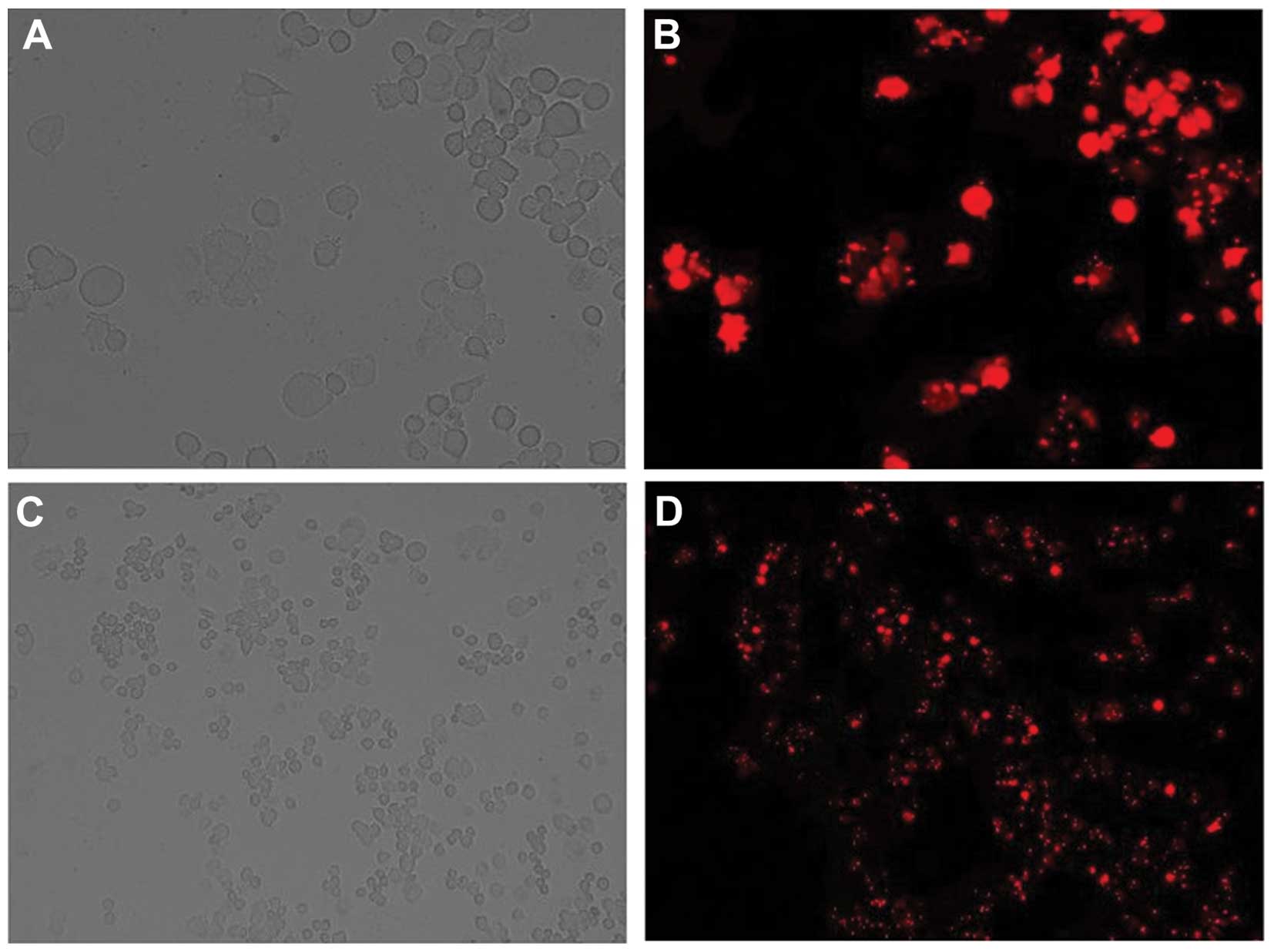
Following the determination of transfection
efficiency in Bel-7402 cells, it was revealed that when using 30 nM
red fluorescent oligo and 5×105/ml Bel-7402 cells, the
efficiency reached >85% (Fig.
4). These were therefore the conditions used to perform the
subsequent experiments.
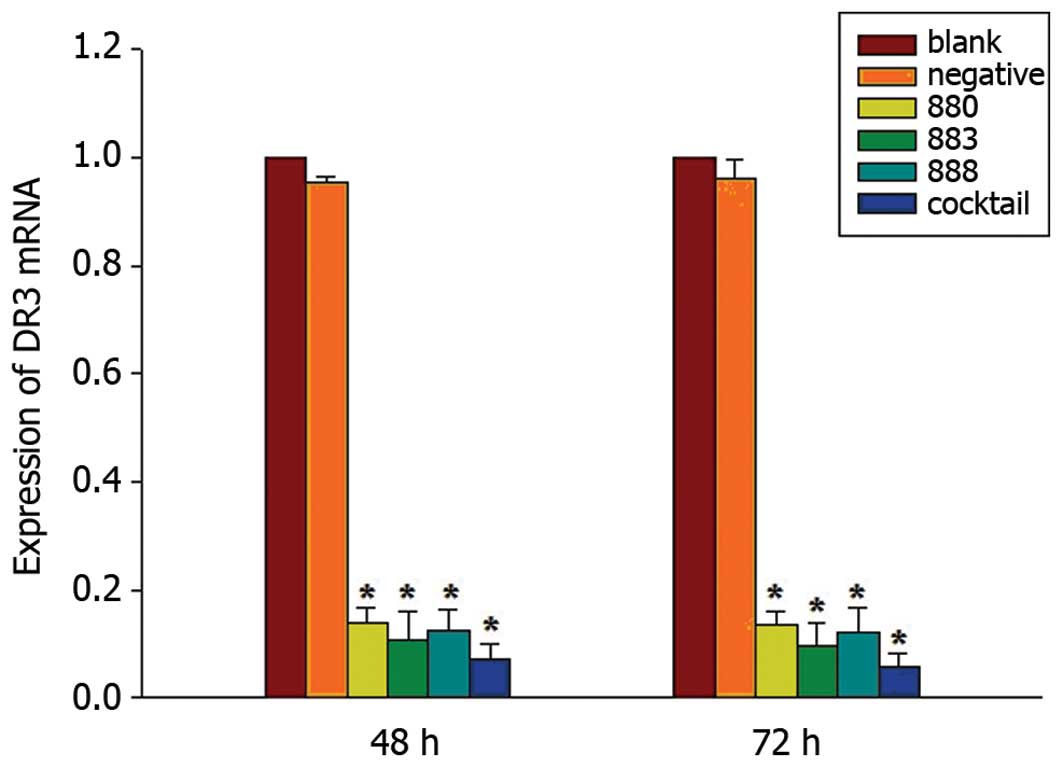
Screening of the most efficient stealth
RNAi sequence targeting DR3
Following transfection of Bel-7402 cells with the
stealth siRNA targeting DR3, the RT-qPCR method was used to measure
mRNA levels of DR3. A significant decrease in DR3 mRNA levels
following DR3 RNAi was observed, while the levels of β-actin
remained unchanged. AF02670.1_stealth_883 demonstrated a more
potent suppression of DR3 mRNA than AF026070.1_stealth_880 or
AF02670.1_stealth_888; however, a cocktail of the three stealth
RNAi siRNAs caused the most potent decrease in DR3 mRNA expression.
Quantification revealed that AF02670.1_stealth_883 reduced DR3 mRNA
by 89.46 and 90.53% of the blank control at 48 and 72 h following
transfection, respectively. The cocktail reduced DR3 mRNA
expression by 92.75 and 94.25% compared with that of the blank
control at 48 and 72 h following transfection, respectively, while
the AF026070.1_stealth_880, AF02670.1_stealth_888 and negative
control group caused decreases of 86.03, 87.52 and 4.70% at 48h, as
well as 86.48, 88.01 and 3.94% at 72 h following transfection,
respectively (Fig. 5).
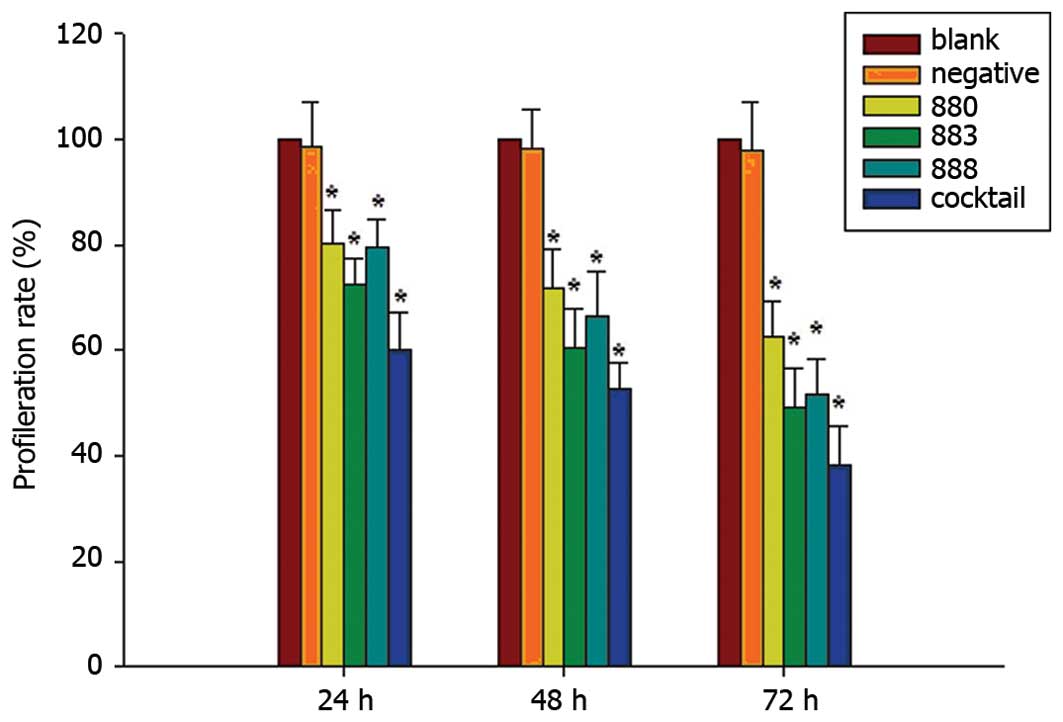
Cell proliferation assessment
MTT assays were used to determine the effect of
transfection at 24, 48 and 72 h on the proliferation in Bel-7402
cells. Cell viability was reduced significantly following treatment
with the individual stealth siRNAs as well as the cocktail against
DR3, compared to that of the negative and blank controls
(P<0.05) (Fig. 6). The cocktail
caused the most potent suppression of proliferation with inhibition
rates of 39.86, 47.51 and 61.76% at 24, 48 and 72 h following
transfection, with the AF026070.1_stealth_883 siRNA causing the
most efficient decrease, with proliferation rates of 27.59, 39.47
and 50.76% of that of the control at 24, 48 and 72 h, respectively
(Fig. 6).
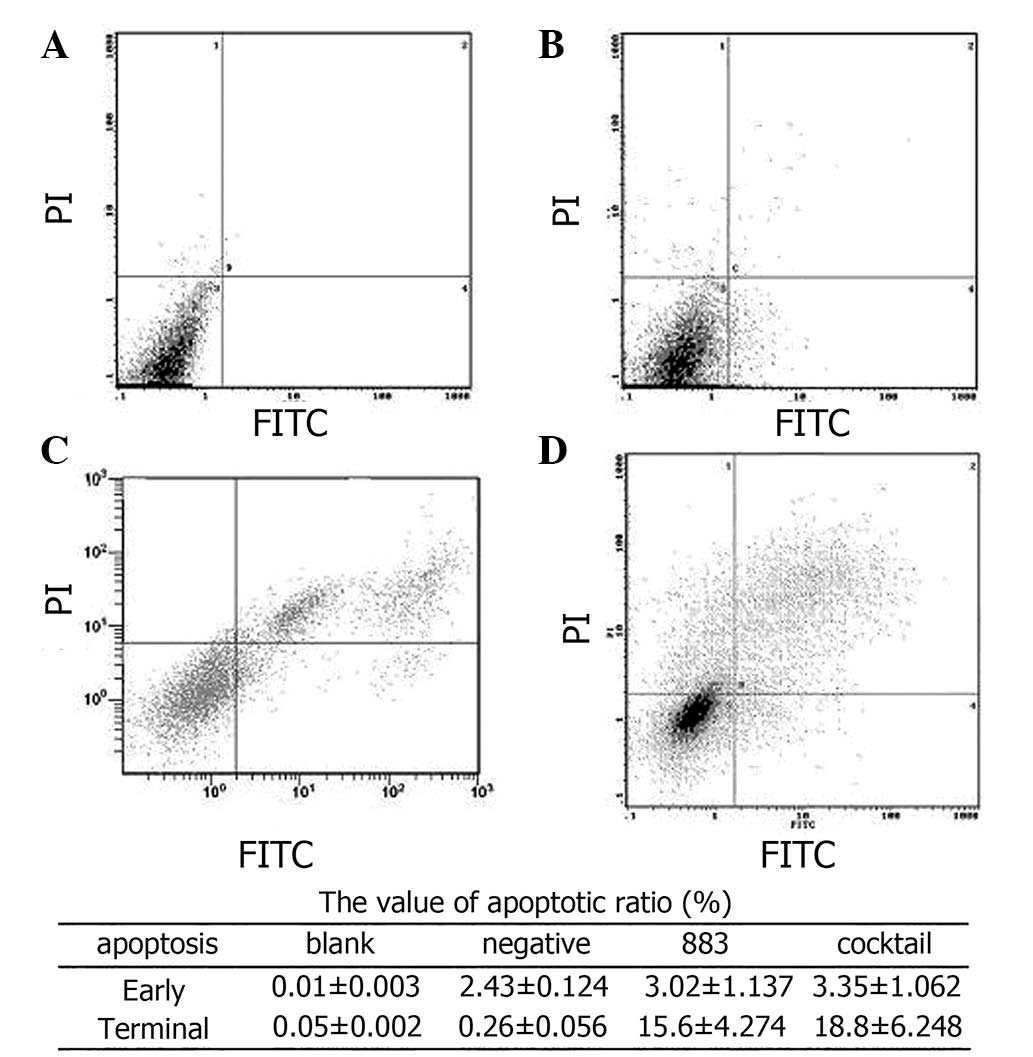
Assessment of apoptosis
To determine the effects of DR3-silencing on
apoptosis, a flow cytometric PI single-staining assay was used on
Bel-7402 cells following 48 h transfection. PI staining revealed
that following AF026070.1_stealth_883 or cocktail treatment, an
apoptotic sub-G1 cell population was present, and the apoptotic
rates were 8.663 and 10.425% (P<0.05) compared with those of the
normal Bel-7402 cells at 0.203% (Fig.
7). In order to further investigate the occurrence of
apoptosis, double staining with FITC/PI was used, the results of
which demonstrated that following 72 h of transfection with
AF026070.1_stealth_883 or cocktail, the proportion of apoptotic
cells at the terminal stage was significantly increased compared to
those in the negative and blank control groups (Fig. 8).
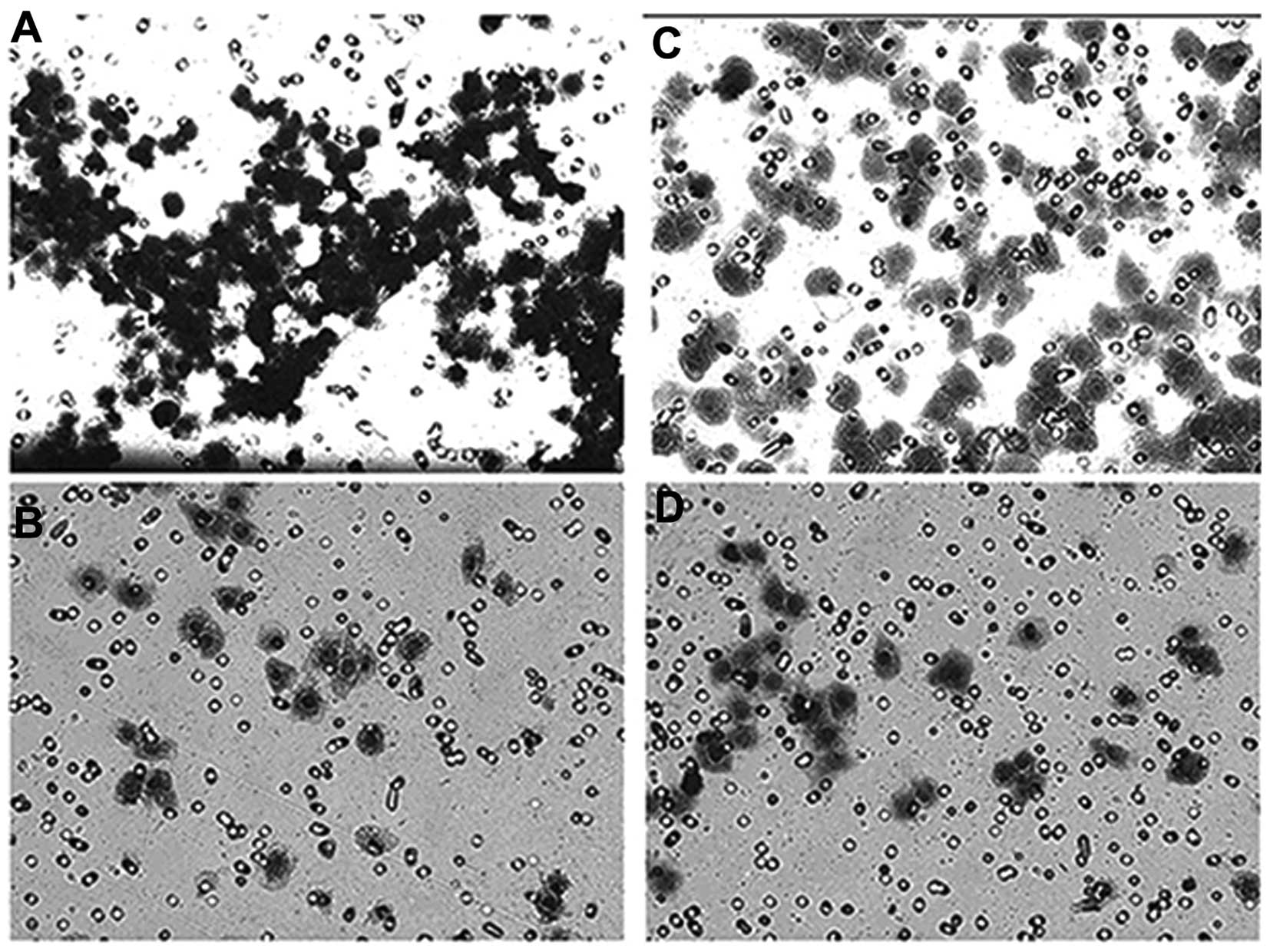
Invasion assay
The results of the Transwell cell invasion assay are
presented in Fig. 9. Following 72
h of transfection by AF026070.1_stealth_883 or cocktail, the
invasion ability of Bel-7402 cells was significantly reduced
compared with that of the blank and negative controls.
Protein assessment
Western blot analysis was used to determine the
expression levels of several proteins in order to elucidate the
mechanisms of DR3 gene silencing-induced apoptosis. Following 72 h
of transfection with AF026070.1_stealth_883 or cocktail into
Bel-7402 cells, protein levels of P53, Caspase3, Fas, Caspase8,
NF-κB, and DR3/TWEAK were detected (Table II). The results revealed
significantly reduced expression of P53 and NF-κB (P<0.05),
whereas Caspase3, Fas, and Caspase8 were increased in the
AF026070.1_stealth_883 and cocktail groups (P<0.05). Levels of
DR3/TWEAK showed no significant change.
| Table IIExpression of proteins at 72 h after
transfection. |
Table II
Expression of proteins at 72 h after
transfection.
| Protein | Blank | Negative | 883 | Cocktail |
|---|
| DR3 | 0.472±0.045 | 0.428±0.037 | 0.072±0.026 | 0 |
| Apo-3L | 0.884±0.054 | 0.867±0.062 | 0.853±0.075 | 0.850±0.047 |
| NF-κb | 0.571±0.051 | 0.528±0.065 | 0.293±0.047 | 0.144±0.038 |
| P53 | 0.973±0.081 | 0.892±0.076 | 0.774±0.068 | 0.683±0.084 |
| Fas | 0.036±0.039 | 0.041±0.053 | 0.479±0.062 | 0.492±0.048 |
| Caspase8 | 0.029±0.007 | 0.043±0.012 | 0.480±0.027 | 0.503±0.037 |
| Caspase3 | 0.370±0.035 | 0.400±0.026 | 0.538±0.044 | 0.582±0.030 |
Discussion
Due to the rapid progression in the fields of
cellular and molecular biology, molecular oncology and other
disciplines, the mechanisms of numerous cell signaling pathways and
target molecules have been elucidated, therefore allowing for the
discovery of targeted oncology drugs such as Sorafenib, which was
approved as a first-line drug for the treatment of advanced renal
cell carcinoma with dual anti-tumor effects by the US Food and Drug
Administration in November 2005. Sorafenib was reported to be able
to directly inhibit tumor growth via inhibition of the rapidly
accelerated fibrosarcoma/mitogen-activated protein kinase
kinase/extracellular signal-regulated kinase signaling pathway.
Conversely, Sorafenib also inhibited the activity of several
tyrosine kinase receptors associated with angiogenesis, and tumor
development, preventing tumor angiogenesis, thereby indirectly
inhibiting tumor growth (19–21).
Bevacizumab [Avastin, recombinant humanized monoclonal
antibody-vascular endothelial growth factor (rhuMAb-VEGF)] was the
first approved drug to inhibit tumor revascularization via VEGF. As
a synthetic recombinant humanized immunoglobulin G1 monoclonal
antibody, bevacizumab can specifically bind to VEGF and form an
inhibitory combination of VEGF and endothelial cell surface
receptors fms-related tyrosine kinase 1 and kinase insert domain
receptor, thereby preventing endothelial cell proliferation and
tumor angiogenesis and subsequently inhibiting tumor growth
(22–24). Another signal transmitter and
activator of the transcription factor 3 [signal transducer and
activator of transcription 3, (STAT3)] signaling pathway of tumor
necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) has
also been studied for its involvement in tumor-associated signal
transduction pathways (25–27).
The results of these studies determined the necessity of
understanding the roles of target molecules in cancer cell survival
and provide a broad spectrum of molecules and pathways for further
study. Although the mechanisms of numerous signaling pathways and
their target molecules have been elucidated, resulting in numerous
effective therapeutic strategies (28–30),
gene therapy for hepatocellular carcinoma has remained ineffective.
Therefore, research into finding the key molecules that regulate
the growth of hepatocarcinoma cells has become an important task
for elucidating the mechanisms of HCC and enhancing the prospects
of therapeutic gene therapy.
DR3, a type of death receptor of the TNF superfamily
(6), was discovered by the
expressed sequence tag database (http://www.ncbi.nlm.nih.gov/nuccore/AF026070.1) from
the umbilical vein endothelial cell cDNA library, following
screening with TNF in 1996, for having similar applications to a
clone of the family members, also known as Apo-3, Trf4/Air2/Mtr4p
polyadenylation complex (TRAMP, lymphocyte-associated receptor of
death), WSL-1 (28,31–33).
DR3 is usually found in organs rich in lymphoid tissue, including
the spleen, thymus, small intestine and peripheral blood
lymphocytes (6). Apoptosis is the
physiological programmed death of cells; a key feature of
apoptosis, caspase enzyme cascade reaction, can occur via internal
and external triggers, ultimately resulting in the breakdown of
DNA, thereby causing karyopyknosis or karyorrhexis (34–37).
Apoptosis consists of three main pathways, namely the mitochondrial
pathway, the death receptor pathway and the endoplasmic reticulum
pathway (38–40). The occurrence of apoptosis via the
death receptor pathway occurs when specific cell surface death
receptors are triggered by extracellular death signals, which then
activate the intracellular mechanism of apoptosis induction.
Previous studies on DR3 reported that under pathological
conditions, DR3 may cause apoptosis via a downstream death domain
(41,42). Studies have demonstrated that the
expression of DR3 in the hepatocarcinoma cell lines SMMC-7721 and
HepG2 was increased compared to that of normal liver cells;
furthermore, the anti-cancer drug lupeol, was shown to have
potential in decreasing DR3 expression levels in hepatocellular
carcinoma cells (43). In 2006,
Gout et al (15) reported
that DR3 expression in colon cancer tissue was higher than that in
adjacent and normal colon tissues and that silencing the gene
expression of DR3 reduced colon cancer HT29 cell adhesion and
migration capacity, as well as weakened the metastatic potential of
HT29 cells (16). In addition,
there have been several studies investigating the association
between the DR3 and HCC; Jiang et al (16) found that DR3 was highly expressed
in hepatocarcinoma H3B cells. However, the role of DR3 in the
progression of HCC remains to be elucidated; furthermore, it
remains to be explained why the high expression of DR3 in tumor
cells fails to induce apoptosis.
The aim of the present study was to clarify the role
of DR3 in human hepatocarcinoma cells. RNAi siRNAs were used to
silence DR3 expression in the hepatocarcinoma cell line Bel-7402.
RT-qPCR experiments demonstrated that the three targeted DR3 siRNAs
(Stealth siRNA AF026070.1_stealth_880, AF02670.1_stealth_883 and
AF02670.1_stealth_888) and cocktail mixtures following transfection
for 48 and 72 h effectively inhibited the expression of DR3 mRNA,
with a silencing efficiency of >85%; the highest silencing
efficiencies, expressed as the inhibitory rate of DR3 mRNA levels,
were achieved by AF02670.1_stealth_883 and a cocktail of the three
sequences following transfection for 48 and 72 h at 89.46 and
92.75%, and 90.53 and 94.25% (P<0.01), respectively.
MTT assays confirmed that silencing DR3 gene
expression significantly inhibited cell proliferation in Bel-7402
cells following transfection with AF026070.1_stealth_880,
AF02670.1_stealth_883, AF02670.1_stealth_888 and a cocktail of the
three the Stealth™ RNAi siRNAs at 24, 48 and 72 h (P<0.05). The
most potent inhibition of cell proliferation was observed with
AF02670.1_stealth_883 and cocktail 72 h following transfection, at
50.76 and 61.76% (P<0.05) compared to that of the negative
control siRNA, which showed no inhibitory effect on the growth of
Bel-7402 cells. Flow cytometry following PI staining and PI/FITC
double staining confirmed that DR3-silencing induced apoptosis, and
Transwell experiments demonstrated significantly reduced
hepatocarcinoma cell invasion. These results indicated that high
expression of DR3 in hepatocarcinoma Bel-7402 cells may promote
proliferation and inhibit apoptosis.
Western blot analysis revealed that following DR3
silencing, the expression levels of apoptosis-associated proteins,
including Fas, Caspase8 and Caspase3, were increased, while the
expression of NF-κB was significantly reduced, which was consistent
with the MTT results. Protein levels of the mitochondrial
transcription factor P53 were also significantly decreased,
indicating DR3 may also interact with the mitochondrial pathway to
regulate apoptosis. Expression levels of Apo-3L were not
significantly altered; therefore, it was hypothesized that Apo-3L
does not only bind DR3 in the regulation of tumor cell apoptosis,
but acts as a ligand for other regulators of apoptosis in tumor
cells that remain to be elucidated.
In conclusion, the results of the present study
demonstrated that silencing the expression of DR3 significantly
inhibited hepatocarcinoma cell proliferation and invasion,
therefore indicating that DR3 promotes proliferation and invasion
of HCC, as well as downregulates apoptosis. Overall, the inhibition
of DR3 may be a potential target for the treatment of HCC.
Based on the results of the present and previous
studies, the next progression of research into DR3-targeted
therapies is to construct a targeted small hairpin RNA (shRNA)
plasmid vector of the DR3 gene and cultivate stable DR3 silenced
hepatocarcinoma cell lines for nude mice experiments in
vitro. These experiments will aim to compare DR3 −/− and high
expressing DR3 liver cancer cell tumorigenicity and metastatic
ability. Furthermore, yeast two-hybrid technology, with DR3 as a
bait can be used to screen a human liver cDNA library for cDNA that
can interact with the protein. Bioinformatics may then be employed
to identify positive clones; immunoprecipitation technology may be
used to identify the positive clones expressing specific proteins
with a specific DNA fragment of combination and precipitation,
collected for the purpose of analyzing fragments to identify the
DR3-specific binding molecules in liver cancer cells and which
initiate the apoptotic program. This may aid in accurately
clarifying the molecular biological mechanism of DR3 in the process
of development of HCC, and provide theoretical support for
molecular targeted therapy of HCC.
Acknowledgements
This study was supported by the Gansu Provincial
Natural Science grant (no. 1107RJZA103), 2010’s Central
Universities Fundamental Research grant from Lanzhou
University-Lzujbky-2010-206 and Lanzhou University Second Hospital
research project (no. YJ2010-01).
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Estimating the world cancer burden: Globocan 2000. Int J Cancer.
94:153–156. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Llovet JM and Bruix J: Novel advancements
in the management of hepatocellular carcinoma in 2008. J Hepatol.
48(Suppl 1): S20–S37. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bergé M, Bonnin P, Sulpice E, et al: Small
interfering RNAs induce target-independent inhibition of tumor
growth and vasculature remodeling in a mouse model of
hepatocellular carcinoma. Am J Pathol. 177:3192–3201. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schuster MJ and Wu GY: Gene therapy for
hepatocellular carcinoma: progress but many stones yet unturned.
Gastroenterology. 112:656–659. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chinnaiyan AM, O’Rourke K, Yu GL, et al:
Signal transduction by DR3, a death domain-containing receptor
related to TNFR-1 and CD95. Science. 8:990–992. 1996. View Article : Google Scholar
|
|
6
|
Ashkenazi A and Dixit VM: Death receptor:
signaling and modulation. Science. 281:1305–1308. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Croft M: The role of TNF superfamily
members in T-cell function and diseases. Nat Rev Immunol.
9:271–285. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kang YJ, Kim WJ, Bae HU, et al:
Involvement of TL1A and DR3 in induction of proinflammatory
cytokines and matrix metalloproteinase-9 in atherogenesis.
Cytokine. 29:229–235. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fang L, Adkins B, Deyev V and Podack ER:
Essential role of TNF receptor superfamily 25 (TNFRSF25) in the
development of allergic lung inflammation. J Exp Med.
205:1037–1048. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Migone TS, Zhang J, Luo X, et al: TL1A is
a TNF-like ligand for DR3 and TR6/DcR3 and functions as a T cell
costimulator. Immunity. 16:479–492. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pappu BP, Borodovsky A, Zheng TS, et al:
TL1A-DR3 interaction regulates Th17 cell function and Th17-mediated
autoimmune disease. J Exp Med. 205:1049–1062. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Meylan F, Davidson TS, Kahle E, et al: The
TNF-family receptor DR3 is essential for diverse T cell-mediated
inflammatory diseases. Immunity. 29:79–89. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bamias G, Martin C III, Marini M, et al:
Expression, localization, and functional activity of TL1A, a novel
Th1-polarizing cytokine in inflammatory bowel disease. J Immunol.
171:4868–4874. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Warzocha K, Ribeiro P, Charlot C, Renard
N, Coiffier B and Salles G: A new death receptor 3 isoform:
expression in human lymphoid cell lines and non-Hodgkin’s
lymphomas. Biochem Biophys Res Commun. 242:376–379. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gout S, Morin C, Houle F and Huot J: Death
receptor-3, a new E-Selectin counter-receptor that confers
migration and survival advantages to colon carcinoma cells by
triggering p38 and ERK MAPK activation. Cancer Res. 66:9117–9124.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang S, Song MJ, Shin EC, Lee MO, Kim SJ
and Park JH: Apoptosis in human hepatocarcinoma cell lines by
chemotherapeutic drugs via Fas-dependent and Fas-independent
pathways. Hepatology. 29:101–110. 1999. View Article : Google Scholar
|
|
17
|
Zhang L, Zhang Y, Zhang L, Yang X and Lv
Z: Lupeol, a dietary triterpene, inhibited growth and induced
apoptosis through down-regulation of DR3 in SMMC7721 cells. Cancer
Invest. 27:163–170. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Strumberg D: Preclinical and clinical
development of the oral multikinase inhibitor sorafenib in cancer
treatment. Drugs Today (Barc). 41:773–784. 2005. View Article : Google Scholar
|
|
19
|
Adnane L, Trail PA, Taylor I and Wilhelm
SM: Sorafenib (BAY 43-9006, Nexavar), a dual-action inhibitor that
targets RAF/MEK/ERK pathway in tumor cells and tyrosine kinases
VEGFR/PDGFR in tumor vasculature. Methods Enzymol. 407:597–612.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rini BI: Sorafenib. Expert Opin
Pharmacother. 7:453–461. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Willett CG, Boucher Y, di Tomaso E, et al:
Direct evidence that the VEGF-specific antibody bevacizumab has
antivascular effects in human rectal cancer. Nat Med. 10:145–147.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Goodman L: Persistence - luck - Avastin. J
Clin Invest. 113:9342004. View
Article : Google Scholar
|
|
23
|
Bergsland E and Dickler MN: Maximizing the
potential of bevacizumab in cancer treatment. Oncologist. 9(Suppl
1): 36–42. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu AX, Blaszkowsky LS, Ryan DP, et al:
Phase II study of gemcitabine and oxaliplatin in combination with
bevacizumab in patients with advanced hepatocellular carcinoma. J
Clin Oncol. 4:1898–1903. 2006. View Article : Google Scholar
|
|
25
|
David D, Rajappan L, Balachandran KK, et
al: Prognostic significance of STAT3 and phosphorylated STAT3 in
human soft tissue tumors-a clinicopathological analysis. J Exp Clin
Res. 30:562011. View Article : Google Scholar
|
|
26
|
Roth W, Grund K, Wiestler OD and
Schirmacher P: The anti-diabetic drug troglitazone sensitizes colon
cancer cells to TRAIL-induced apoptosis by down-regulating FLIP.
Verh Dtsch Ges Pathol. 91:294–301. 2007.(In German).
|
|
27
|
Zhao B, Li L, Cui K, Wang AL, et al:
Mechanisms of TRAIL and gemcitabine induction of pancreatic cancer
cell apoptosis. Asian Pac J Cancer Prev. 12:2675–2678. 2011.
|
|
28
|
Wilhelm SM, Carter C, Tang L, et al: BAY
43-9006 exhibits broad spectrum oral antitumor activity and targets
the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in
tumor progression and angiogenesis. Cancer Res. 64:7099–7109. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Herold-Mende C, Steiner HH, Andl T, et al:
Expression and functional significance of vascular endothelial
growth factor receptors in human tumor cells. Lab Invest.
79:1573–1582. 1999.
|
|
30
|
Kitson J, Raven T, Jiang YP, et al: A
death-domain-containing receptor that mediates apoptosis. Nature.
384:372–375. 1996. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Marsters SA, Sheridan JP, Donahue CJ, et
al: Apo-3, a new member of the tumor necrosis factor receptor
family, contains a death domain and activates apoptosis and
NF-kappa B. Curr Biol. 6:1669–1676. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bodmer JL, Burns K, Schneider P, et al:
TRAMP, a novel apoptosis-mediating receptor with sequence homology
to tumor necrosis factor receptor 1 and Fas (Apo-1/CD95). Immunity.
6:79–88. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Screaton GR, Xu XN, Olsen AL, Cowper AE,
Tan R, McMichael AJ and Bell JI: LARD: A new lymphoid-specific
death domain containing receptor regulated by alternative pre-mRNA
splicing. Proc Natl Acad Sci USA. 94:4615–4619. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Abou-Alfa GK, Schwartz L, Ricci S, et al:
Phase II study of sorafenib in patients with advanced
hepatocellular carcinoma. J Clin Oncol. 24:4293–4300. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Butterfield LH: Immunotherapeutic
strategies for hepatocellular carcinoma. Gastroenterology. 127(5
Suppl 1): S232–S241. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Butterfield LH: Recent advances in
immunotherapy for hepatocellular cancer. Swiss Med Wkly. 137:83–90.
2007.PubMed/NCBI
|
|
37
|
Song G, Luo Q, Qin J, Wang L, Shi Y and
Sun C: Effects of oxymatrine on proliferation and apoptosis in
human hepatoma cells. Colloids Surf B Biointerfaces. 48:1–5. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ashkenazi A and Dixit VM: Death receptors:
signaling and modulation. Science. 281:1305–1308. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nakagawa T, Zhu H, Morishima N, et al:
Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and
cytotoxicity by amyloid-beta. Nature. 403:98–103. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pittoni P, Tripodo C, Piconese S, Mauri G,
Parenza M, Rigoni A, Sangaletti S and Colombo MP: Mast cell
targeting hampers prostate adenocarcinoma development but promotes
the occurrence of highly malignant neuroendocrine cancers. Cancer
Res. 71:5987–5997. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dechant MJ, Fellenberg J, Scheuerpflug CG,
Ewerbeck V and Debatin KM: Mutation analysis of the apoptotic
‘death-receptors’ and the adaptors TRADD and FADD/MORT-1 in
osteosarcoma tumor samples and osteosarcoma cell lines. Int J
Cancer. 109:661–667. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
He Y, Liu F, Zhang L, et al: Growth
inhibition and apoptosis induced by lupeol, a dietary triterpene,
in human hepatocellular carcinoma cells. Biol Pharm Bull.
34:517–522. 2011. View Article : Google Scholar : PubMed/NCBI
|