Introduction
Annually, >3,500,000 patients worldwide suffer
from peripheral nerve injuries, accounting for 3% of all trauma
patients. Among the various types of peripheral nerve injury,
transection injuries, in which the nerve trunk is completely
separated, particularly those that result in large defects, may
produce a marked impact on patients′ quality of life, and in these
cases, reconstructive surgery is required as a therapeutic strategy
to achieve nerve regeneration. Several studies have attempted to
accelerate the regeneration of traumatized nerves via the
application of microsurgical techniques (1–6).
However, despite advancements in these techniques, complete
recovery is rarely achieved (7,8).
Schwann cells (SCs) are considered to be the primary
structural and functional cells of the peripheral nervous system,
and these cells exert a critical supportive role in peripheral
nerve regeneration (9,10). The transplantation of cultured SCs
into various peripheral nerve grafts scaffolds has been
demonstrated to promote axonal regeneration across nerve gaps
(11). However, the clinical
application of SCs remains controversial as obtaining sufficiently
large numbers of SCs is difficult. In the past few decades, bone
marrow stromal cells (BMSCs) and adipose-derived stem cells (ADSCs)
have been considered as alternative cell sources for tissue
regeneration following peripheral nerve injury, as these cells are
capable of selectively differentiating into SCs and enhancing
axonal regeneration (12,13). Each of these cell types has
advantages; however, numerous problems have been associated with
the use of each of the currently available cell sources, including
complications regarding cell survival, immune rejection,
tumorigenesis and ethical issues. Therefore, the development of an
ideal cell source is important to address these problems. Recently,
human amniotic epithelial cells (hAECs) have been reported to
exhibit stem cell characteristics, and these cells have been shown
to differentiate into liver cells, fat cells, heart cells and nerve
cells in vitro (14–19).
hAECs are easily accessible and highly abundant since the amnion is
normally discarded following birth. Furthermore, hAECs exhibit low
immunogenicity and have the potential to differentiate into various
types of functional cell (20).
These characteristics render hAECs an alternative source of adult
stem cells for SC transplantation for peripheral nerve
regeneration.
In the present study, whether hAECs possess the
potential to differentiate into Schwann-like cells was investigated
via indirect co-culture of hAECs with SCs in vitro.
Immunocytochemical staining was performed to determine the
biological features of the differentiated cells. Co-culture of the
differentiated hAECs with PC12 cells was conducted to assess the
effect of differentiated hAECs on axon regeneration.
Materials and methods
Isolation and culture of hAECs
In brief, discarded placentas from uncomplicated
cesarean resections at 37–40 weeks gestational age were obtained
from The First Affiliated Hospital of Sun Yat-sen University
(Guangzhou, China) with the approval of the University of Sun
Yat-sen Institutional Review Board. The amnion layer was
mechanically peeled away from the chorion and washed multiple times
using phosphate-buffered saline (PBS; Invitrogen, Carlsbad, CA,
USA) to remove the blood. The amnion layer was then cut into 2×2 cm
sections and processed according to previously described methods
(21). To release the hAECs, the
amnion sections were incubated at 37°C in 0.05% trypsin containing
0.2 g/l ethylenediaminetetraacetic acid (Invitrogen/Gibco-BRL;
Carlsbad, CA, USA). Subsequently, the trypsin was inactivated by
adding 10 ml Dulbecco’s modified Eagle’s medium/F-12 Ham nutrient
mixture (DMEM/F-12; 1:1; Gibco-BRL) supplemented with 10%
heat-inactivated fetal bovine serum (FBS; Invitrogen Life
Technologies,). The cells from the initial 10 min of digestion were
discarded to remove any debris. The cells from the second and third
30-min digestions were pooled and washed three times with PBS. The
hAECs were then plated on six-well dishes at a seeding density of
20,000 cells/cm2 in DMEM/F-12 (1:1) culture medium
supplemented with 10% FBS, 1% penicillin/streptomycin (Invitrogen),
1% GlutaMAX™ (Invitrogen) and 10 ng/ml epidermal growth factor
(EGF; PeproTech, Rocky Hill, NJ, USA). The hAECs from the initial
culture were subcultured to passage 1 using a split ratio of 1:3.
The above-mentioned culture medium was replaced every three
days.
Isolation and culture of SCs
Rat SCs were harvested as previously described, with
minor modifications (22).
Briefly, for collection of the sciatic nerves, the male
Sprague-Dawley rats (1–3 days old, weighing 8–15 g; Laboratory
Animal Center of Sun Yat-sen University, Guangzhou, China) were
anesthetized by cooling on ice one at a time. Once the animal had
stopped moving it was decapitated, and the head placed into a
collection bag. The body was spayed with 70% ethanol. The sciatic
nerves were exposed, dissected out under a binocular microscope
(Leica S6E, Wetzlar, Germany) and placed in a sterile culture dish
(3-cm diameter; Costar, Corning Inc., Corning, NY, USA) containing
ice-cold PBS with 1% penicillin/streptomycin, where they were
maintained until the nerves had been harvested from all of the rat.
Then, the sciatic nerves were enzymatically dissociated via
sequential incubation at 37°C in 0.3% collagenase type II
(Sigma-Aldrich, St. Louis, Missouri, USA) and 0.125% trypsin
(Invitrogen/Gibco-BRL) for 30 and 10 min, respectively. The mixture
was triturated, centrifuged and resuspended in DMEM/F-12
supplemented with 10% FBS and 1% penicillin/streptomycin. On the
following day, 10 μM cytosine arabinoside (Sigma-Aldrich) was
added, and the mixture was allowed to incubate for an additional 48
h to remove any fibroblasts. Subsequently, the culture was
maintained in DMEM/F-12 supplemented with 10% FBS, 1%
penicillin/streptomycin and 2 ng/ml heregulin (Sigma-Aldrich, St.
Louis, MO, USA) to stimulate SC proliferation. All media and
supplements were purchased from Invitrogen/Gibco-BRL. All
experiments were performed in accordance with the guidelines for
the use of experimental animals from the National Institutes of
Health and ethical approval for this study was obtained from the
Ethics Committee of Sun Yat-sen University.
Viability and proliferation assays for
hAECs
The hAECs were harvested during the logarithmic
growth phase and digested using trypsin. To examine cell
proliferation and viability, the hAECs (5×103
cells/well) were plated in 96-well dishes and incubated in
DMEM/F-12 supplemented with 10% FBS. Plates were incubated at 37°C
in 5% CO2 for 24 or 48 hours. Four hours prior to the
end of the incubation time, 20 μl 3-(4, 5-dimethylthiazol-2-yl)
2,5diphenyltetrazolium bromide solution (MTT, Sigma-Aldrich) was
added to each well. Following a 4 h of incubation, plates were
centrifuged and culture medium was removed. Precipitated formazan
crystals were dissolved by adding 150 μL DMSO to each well. The
microplates were shaken at room temperature for 10 min. A
microplate reader (OPTImax; Molecular Dynamics, Sunnyvale, CA, USA)
was used to measure the absorbance (A) of each well at 540 nm, and
the average values were obtained. Experiments were repeated ≥3
times, and the data are presented as the means ± standard deviation
(SD).
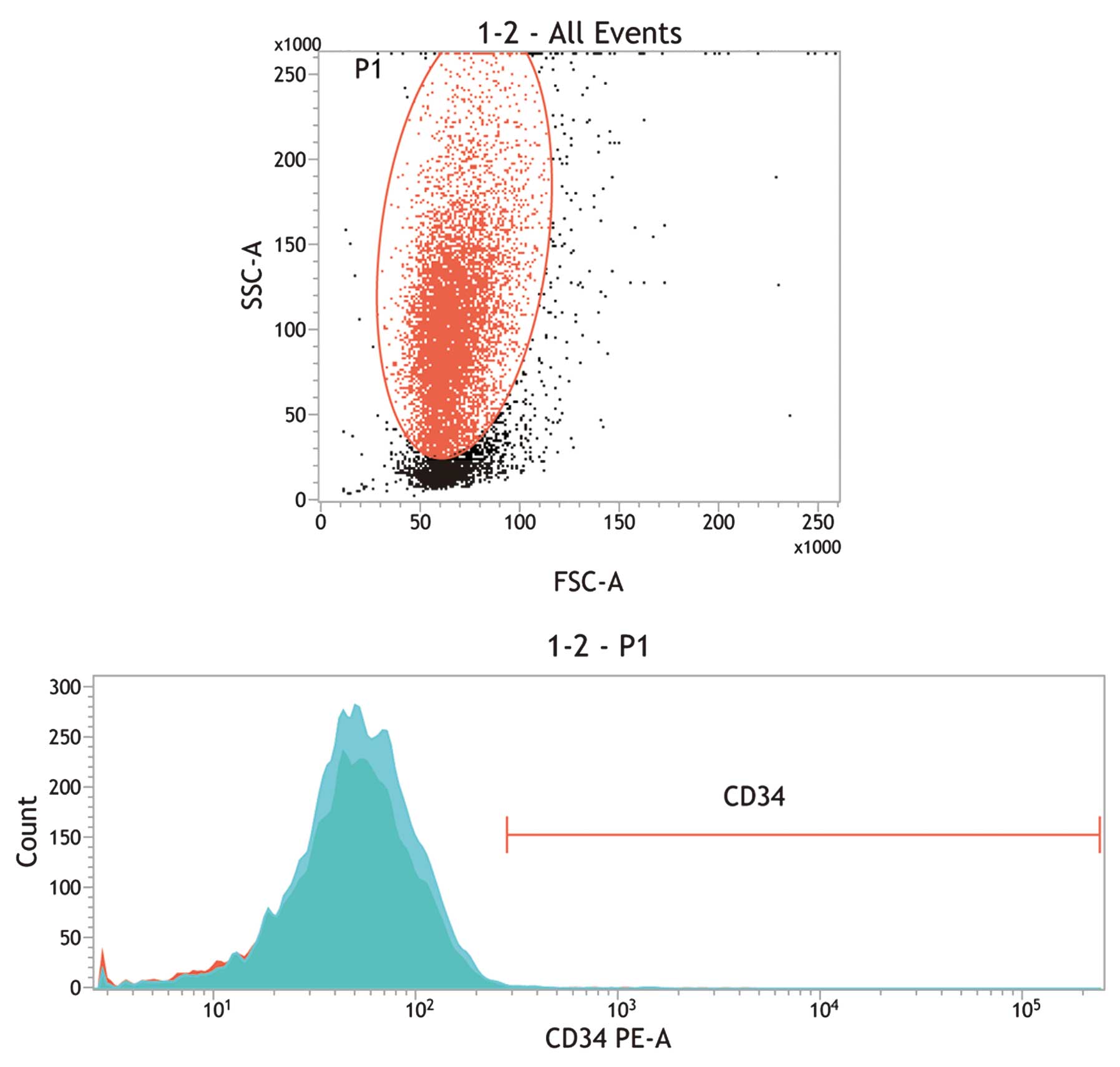
Flow cytometric analysis
Isolated hAECs were harvested and washed using 0.5%
bovine serum albumin (BSA)/0.01 M PBS solution. Following fixation
using 4% paraformaldehyde (Invitrogen), the cells were resuspended
in 0.5% BSA/PBS solution and incubated for 1 h at room temperature
with fluorescein isothiocyanate (FITC)-conjugated primary mouse
monoclonal antibody against CD34 (MA1-12103; Thermo Fisher
Scientific, Waltham, MA, USA; 1:200). Negative controls were
produced using mouse immunoglobulin G 1-isotyped antibodies
conjugated to FITC (Beckman Coulter, Marseille, France). The
samples were analyzed using a BD FACSCanto™ II system (BD
Biosciences, San Jose, CA, USA). A total of 3×104 events
were captured using the BD FACSDiva™ software (BD Biosciences)
histogram feature. The data were further analyzed using CellQuest
software (BD Biosciences) and WinMDI software version 2.9 (Scripps
Research Institute; La Jolla, CA, USA).
Differentiation of hAECs via co-culture
with SCs
hAECs were co-cultured with SCs in a double-chamber
system with SCs on the upper level and hAECs on the lower level of
the culture plate. The cultures were suspended in DMEM/F-12 culture
medium at a density of 2×104 cells/well in a 6-well dish
(Costar; Corning Inc., Corning, NY, USA). Culture inserts (Costar;
Corning Inc.), each containing 8×104 SCs, were placed in
four of the six wells using one insert/well. An insert membrane
with a pore size of 0.4 μm diameter was used to permit only the
transmission of soluble factors and to prevent direct interaction
between the hAECs and the SCs. Half the volume of the co-culture
medium was replaced every three days. hAECs under identical
conditions but without co-culture with SCs served as negative
controls and SCs alone served as positive controls.
Immunocytochemistry
The characteristics of differentiated hAECs at the
protein expression level were assessed via immunocytochemistry. The
cells were fixed at room temperature with 4% paraformaldehyde for
10 min. Subsequently, the cells were washed with PBS, followed by
permeabilization for 30 min using 0.1% Triton X-100. Non-specific
binding sites were blocked by incubation with 5% goat serum for 1
h. The following primary antibodies were employed for
immunocytochemistry: monoclonal mouse against Pan-cytokeratin
(4545; 1:200) and monoclonal mouse against S-100 (5529; 1:200)
obtained from Cell Signaling Technology (Danvers, MA, USA). The
cells were incubated in the primary antibodies overnight at 4°C.
The cells were washed three times with PBS and incubated with the
corresponding goat secondary antibodies (4408; 1:500) for 2 h at
room temperature. Subsequent to washing twice with PBS, the cells
were incubated in 4′,6′-diamidino-2-phenylindole dihydrochloride
(1:1,000; Sigma-Aldrich) for 5 min at room temperature to
counterstain the cell nuclei. The specimens were washed three times
with PBS, each for 5 min, and were then mounted using an antifade
solution. Immunolabeled cells were examined via fluorescence
microscopy (Zeiss Axiovision Z1, Oberkochen, Germany). Images were
digitally recorded and processed using Image-Pro Plus (Media
Cybernetics, Rockville, MD, USA).
RNA isolation
To collect total RNA, 2×106 cells were
seeded on 10 cm diameter dishes in 8 ml growth medium. The cultures
were maintained at 37°C in a humidified atmosphere of 5%
CO2/95% air. The media were replaced, including the drug
of interest. After six days, total RNA was isolated using an Isogen
kit (Nippon Gene, Tokyo, Japan) according to the manufacturer’s
instructions. RNA quantity and purity were determined via
spectrophotometry (DU-65; Beckman Coulter, Fullerton, CA, USA).
Reverse trancription quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed using an RNA PCR kit [avian
myeloblastosis virus (AMV)] version 2.1 (Takara Bio, Inc., Shiga,
Japan) according to the manufacturer’s instructions and with a PCR
Thermal Cycler (Takara Bio, Inc.). First-strand cDNA was
synthesized from total RNA (1 mg) using AMV reverse transcriptase
XL primed using 50 pmol random 9 mers (Takara Bio, Inc.). The
first-strand reaction was conducted as follows: 30°C for 10 min,
50°C for 30 min, 99°C for 5 min and 58°C for 5 min. The
reverse-transcription reaction products (10 ml) were utilized as
templates for PCR using 0.2 mM of each primer. Primers specific to
rat nerve growth factor (NGF), glial cell-derived neurotrophic
factor (GDNF), brain-derived neurotrophic factor (BDNF) and
glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were designed
using Primer Express 2.0 software (Applied Biosystems, Foster City,
CA, USA) and were obtained from Shanghai HuiRui Biotechnology Co.,
Ltd. (Shanghai, China). The PCR products were separated via precast
2% agarose gel (Daiichi Pure Chemicals Co., Ltd., Tokyo, Japan)
electrophoresis and visualized via SYBR-Green 2 (FMC Bioproducts,
Rockland, ME, USA) staining using a ultraviolet trans-illuminator
(UVP Inc., San Gabriel, CA, USA). The signal intensity of the PCR
products was determined using Image J software. The expression
levels of the GDNF, BDNF and NGF PCR products were quantified as
determined by the intensity of the PCR products compared with that
of the corresponding GAPDH reactions.
Functional co-culture assay with PC12
cells
PC12 cells were plated in six-well chamber slides
pre-coated with collagen (Sigma-Aldrich) at a density of 1,000
cells/well. The PC12 cells were incubated for 24 h in DMEM
supplemented with 5% horse serum, 10% FBS, 100 U/ml penicillin and
100 μg/ml streptomycin. At 24 h prior to plating the PC12 cells,
293T cells and hAECs differentiated towards a SC phenotype were
seeded on 0.4 μm pore size culture inserts at a seeding density of
2×104 cells/insert and incubated at 37°C, 95% humidity
and 5% CO2. After 48 h, the inserts were checked for
cell adherence and were added to the PC12 chamber slides such that
molecules secreted by the cells were able to permeate to the PC12
cells without direct cell contact. Neuron-like outgrowth was
measured using an Olympus IX81 microscope (Olympus Corporation,
Tokyo, Japan) and a U-CMAD 3 camera (Olympus Corporation). Cell
body size and axon length were analyzed using SIS image analysis
software version 3.2 (Olympus Soft Imaging Solutions GmbH, Münster,
Germany). PC12 cells with axons longer than the average cell
diameter were used for data analysis. Cell axon length was analyzed
using image-pro-plus (SIS, Münster, Germany).
Statistical analysis
All data were analyzed using SPSS 11.0 statistical
software (SPSS, Inc., Chicago, IL, USA). The experimental data are
expressed as the mean ± SD. The Student’s t-test was performed to
compare the values. P<0.05 was considered to indicate a
statistically significant difference.
Results
Identification of hAECs
The viability of the isolated cells was >95%, as
determined via trypan blue staining. In the presence of EGF, hAECs
proliferated robustly and formed a confluent monolayer of
cobblestone-shaped epithelial cells (Fig. 1A). Approximately all isolated cells
reacted with antibodies against pan-cytokeratin, which revealed the
epithelial nature of the cells and the lack of contamination with
human amniotic mesenchymal stromal cells (Fig. 1B). The absence of CD34-positive
cells in this population indicates that the isolates were not
contaminated with hematopoietic stem cells, such as umbilical cord
blood cells or embryonic fibroblasts (Fig. 2).
Identification of differentiated
hAECs
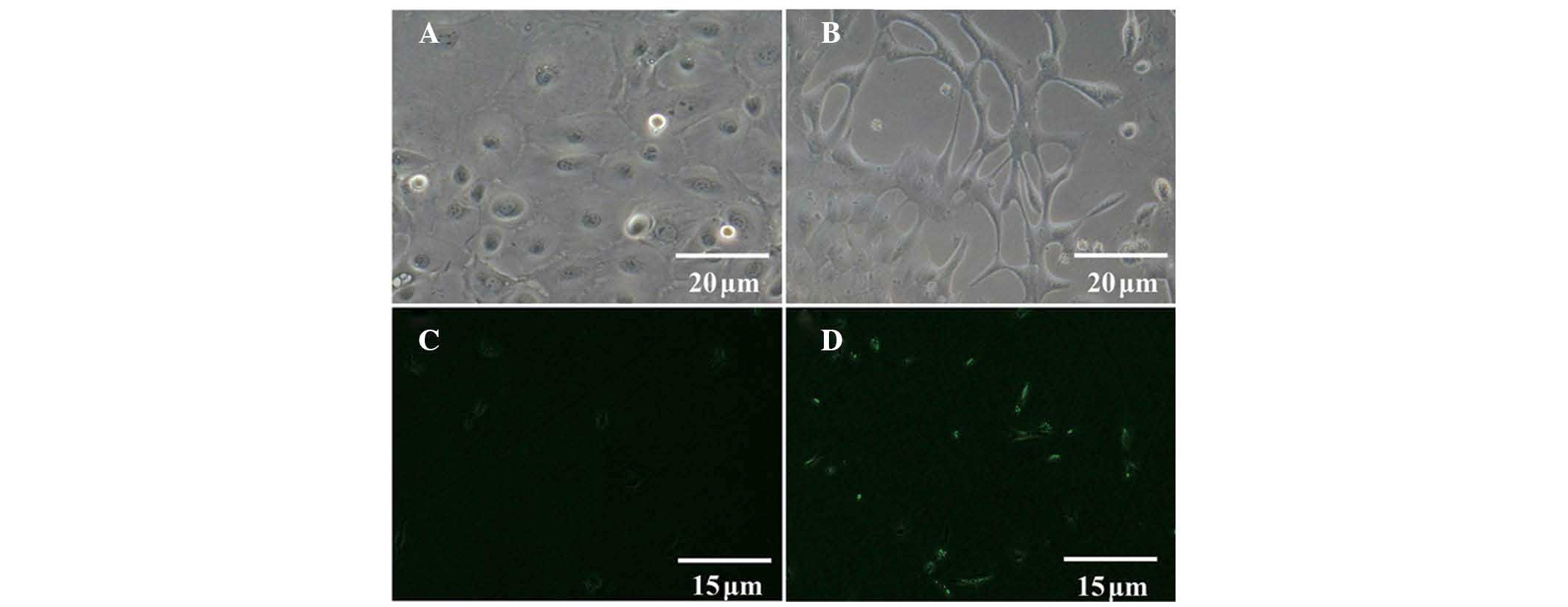
hAECs cultured alone appeared as a monolayer of
cobblestone-shaped epithelial cells (Fig. 3A). Following co-culture of the
cells with SCs for 21 days, certain hAECs exhibited a typical
bipolar or tripolar morphology (Fig.
3B). As compared with undifferentiated hAECs (Fig. 3C), differentiated hAECs exhibited
positive expression of S-100 (Fig.
3D).
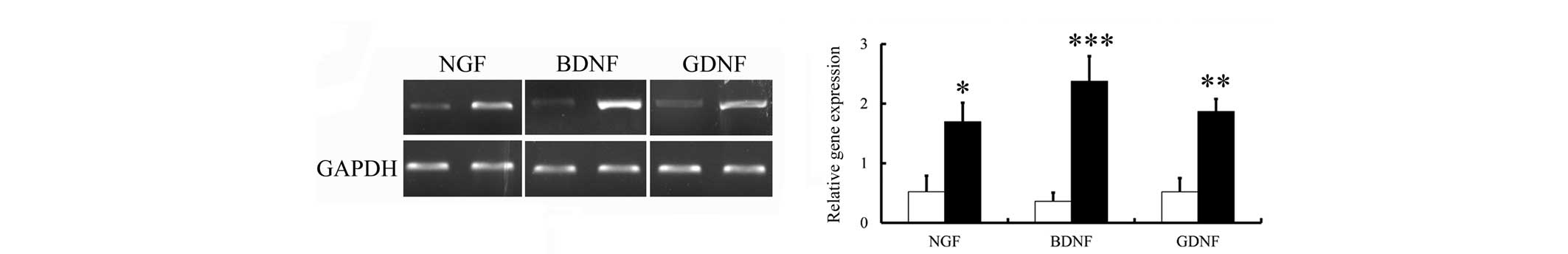
Expression levels of NGF, BDNF and GDNF
mRNA following differentiation
To examine the biological function of the
differentiated cells, the expression levels of the NGF, BDNF and
GDNF neurotrophic factors in the cells were measured via RT-qPCR
using specific primers, as described above. Neurotrophic factors
are polypeptides or proteins that are released by neurons or
non-neuronal cells that exert marked effects on the central and
peripheral nervous systems (23).
Exogenous administration of NGF has been reported to completely
prevent the death of axotomized sensory neurons (24,25).
Subsequent to co-culture of the hAECs with SCs, the expression
levels of NGF mRNA in the hAECs increased 1.55-fold as compared
with treatment with saline (Fig.
4). Subsequently, whether co-culture results in a sustained
increase in BDNF and GDNF mRNA expression levels was determined.
The BDNF and GDNF mRNA levels were also significantly increased
following co-culture (1.5-fold, P<0.05; and 1.8-fold, P<0.05,
respectively).
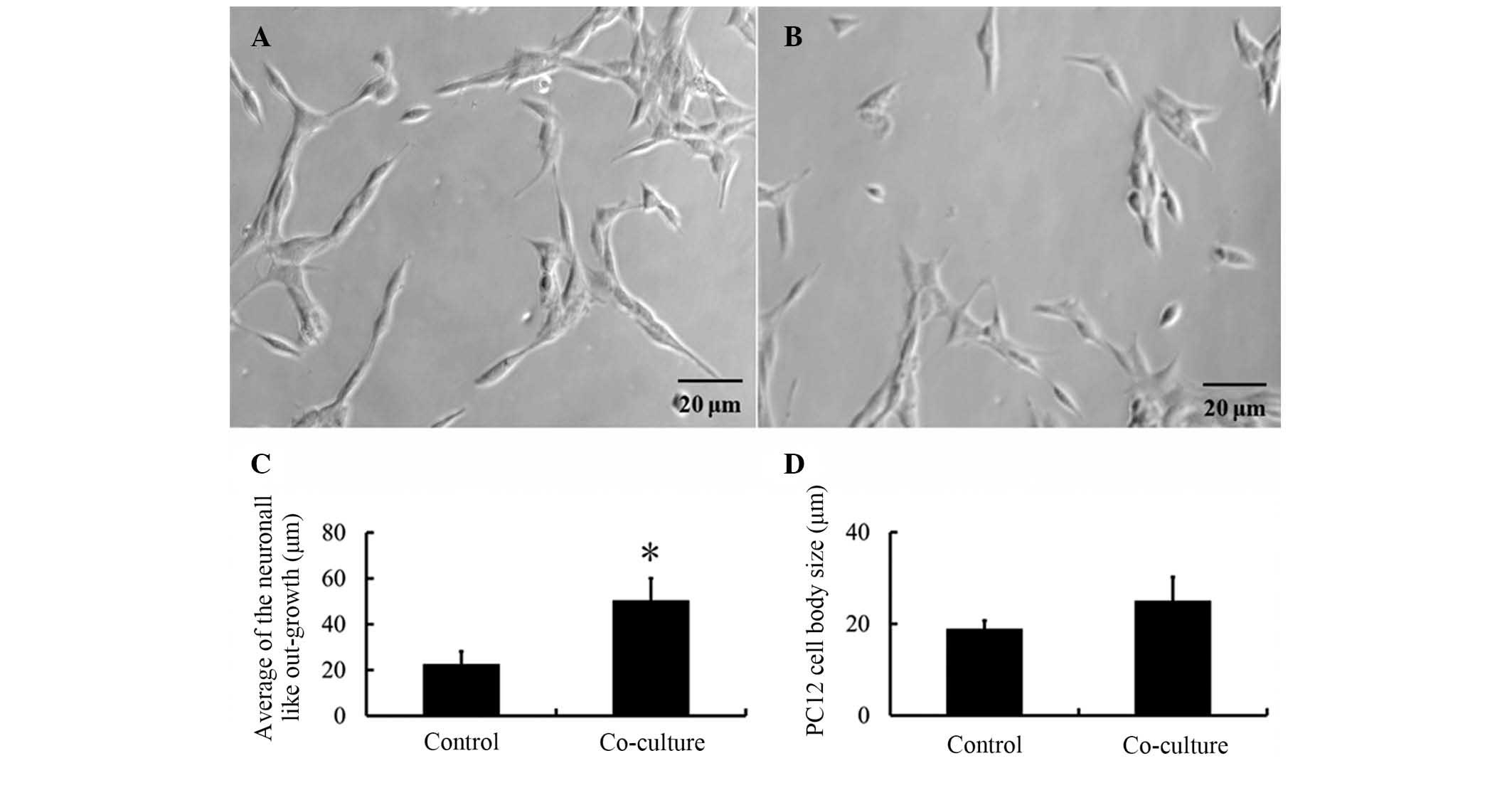
PC12 cell morphology
To determine the effect of differentiated hAECs on
PC12 cell axon regeneration subsequent to co-culture of PC12 cells
with differentiated hAECs, the number of PC12 cells that exhibited
axonogenesis, the cell body sizes and the overall length of the
outgrown fibers were quantified. Fig.
5 clearly presents the biological activity of the
differentiated cells. Statistical analysis of the neuron-like
outgrowth of the PC12 cells revealed significant induction of
axonogenesis following co-culture (P<0.05; 51.32±8.56 μm) as
compared with the control group (21.32±5.45 μm). In addition, the
average cell body size of the PC12 cells was determined during the
culture period since alterations in cell morphology and size
indicate higher metabolic activity induced by external stimuli. The
cell body sizes were measured for each group. The average PC12 cell
body size increased following incubation in the co-culture.
However, these marginal changes were not statistically
significant.
Discussion
In the present study, hAECs were differentiated into
SCs using a co-culture method. The results revealed that, following
differentiation, the hAECs exhibited a spindle-like morphology. The
hAECs were immunopositive for the S-100 SC markers. Subsequently,
the function of the differentiated hAECs was analyzed. The RT-qPCR
results revealed that NGF, BDNF and GDNF expression was upregulated
as compared with the control group. In addition, following the
co-culture of differentiated hAECs with PC12 cells, the
differentiated cells significantly increased the average axon
length of the PC12 cells.
In recent years, numerous strategies have been
developed to prepare nerve conduits involving biomaterials that
incorporate growth factors, cells and natural microarchitectural
structures (26,27). To more effectively repair nerve
defects, the construction of nerve grafts that integrate these
factors is a promising alternative to the employment of autologous
nerves. SCs are a good candidate for use as seed cells. However,
treatment with primary SCs as a cell therapeutic remains limited by
the associated donor site morbidity (28). In addition, for clinical
applications, the time period required to generate a sufficient
quantity of cells is not always feasible (29). Therefore, various studies have
attempted to identify an alternative method. hAECs, which exhibit
characteristics of embryonic and pluripotent stem cells, may be
utilized for cell therapy without legal or ethical problems
(30). The differentiation of
hAECs has been widely investigated. Using β-mercaptoethanol,
Banerjee et al (31) found
that human amniotic membranes were differentiated towards the SC
lineage in vitro. Following differentiation, the cells
expressed BDNF and GDNF, a finding concurrent with that of the
present study.
Contrary to BMSCs or ADSCs, hAECs are not stem
cells. However, hAECs do possess the multi-lineage differentiation
potential required to develop into tissues of three germ layers
(ectodermal, mesodermal and endodermal lineages), a finding which
has been demonstrated by multiple studies (32–38).
In addition, the demonstration of hAEC expression of pluripotent
and multipotent markers, such as octamer-binding protein (Oct)-3/4,
SRY-related HMG-box 2, Nanog, fibroblast growth factor 4, reduced
expression 1, frizzled class receptor 9, bone marrow stromal cell
antigen 1 and ATP-binding cassette sub-family G member 2, provides
further support for the multi-lineage differentiation potential of
hAECs (32,39,40).
In the present study, the overall proportion of
differentiated hAECs remained low. Two factors may account for this
phenomenon. On one hand, the quantity of co-cultured hAECs was
large. Incubation of an excessive number of cells together may not
only restrict cell growth and differentiation but also limit the
relative effect of the proteins secreted by the SCs. Conversely,
Miki et al (32) found that
hAECs grow as three layers when cultured in dishes: A layer of
cells in the supernatant, an intermediate layer and a layer of
cells attached to the culture dish. Furthermore, the expression
levels of the pluripotent markers Oct-4 and Nanog were elevated in
the cells that were present above the basal layer of hAECs, which
indicates that the cells above the basal layer exhibited greater
differentiation ability. This phenomenon may partially explain why
particular hAECs differentiate more quickly and easily. In the
present study, due to the only partial differentiation of hAECs
into SCs observed, the function of the cells following co-culture
was subsequently examined.
PC12 cells are a well-defined and widely used model
for the analysis of axon regeneration (41). In the present study, following
co-culturing, the average axon length was observed to increase from
21.32±5.45 to 51.32±8.56 μm. The PC12 cell body size may also have
increased, as alterations in cell morphology and size indicated a
higher metabolic activity induced by external stimuli. A marginal,
but not significant, increase in the average cell body size was
demonstrated. This result implied that unlike average axon length,
cell body size may not provide an appropriate marker of
regeneration.
In conclusion, in general, using a co-culture
technique, hAECs were demonstrated in the present study to
differentiate towards SCs in vitro. The differentiated hAECs
exhibited certain characteristics of SCs and greatly increased the
axon length of PC12 cells. These promising results indicate that
this method may provide a good strategy for the treatment of nerve
injuries in humans.
Acknowledgements
This study was supported by grants from Guangdong
Medical Science and Technology Research Fund (grant no. A2011163),
Doctoral Startup Project of Guangdong Natural Science Foundation
(grant no. S201204006336), the National High Technology Research
and Development Program of China (863 Program, grant no.
012AA020507), the Nature Science Grant of Guangdong China (grant
no. 9251008901000017, and 2012AA020507), the National Basic
Research Program of China (973 Program, grant no. 2014CB542201) and
the 985 program of Sun Yat-sen University (grant no.
90035-3283312).
References
|
1
|
He B, Zhu Q, Chai Y, et al: Safety and
efficacy evaluation of a human acellular nerve graft as a digital
nerve scaffold: a prospective, multicentre controlled clinical
trial. J Tissue Eng Regen Med. Feb 25–2013.(Epub ahead of print).
View Article : Google Scholar
|
|
2
|
Okui N, Yamamoto M, Fukuhira Y, Kaneko H
and Hirata H: A new nerve coaptation technique using a
biodegradable honeycomb-patterned film. Microsurg. 32:466–474.
2012. View Article : Google Scholar
|
|
3
|
Meng H1, Li M, You F, Du J and Luo Z:
Assessment of processed human amniotic membrane as a protective
barrier in rat model of sciatic nerve injury. Neurosci Lett.
496:48–53. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wangensteen KJ and Kalliainen LK: Collagen
tube conduits in peripheral nerve repair: a retrospective analysis.
Hand (N Y). 5:273–277. 2010. View Article : Google Scholar
|
|
5
|
Kehoe S, Zhang XF and Boyd D: FDA approved
guidance conduits and wraps for peripheral nerve injury: a review
of materials and efficacy. Injury. 43:553–572. 2012. View Article : Google Scholar
|
|
6
|
Elliot D, Lloyd M, Hazari A, Sauerland S
and Anand P: Relief of the pain of neuromas-in-continuity and
scarred median and ulnar nerves in the distal forearm and wrist by
neurolysis, wrapping in vascularized forearm fascial flaps and
adjunctive procedures. J Hand Surg Eur Vol. 35:575–582. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kline DG and Hudson AR: Vertebral artery
compression. J Neurosurg. 83:7591995.PubMed/NCBI
|
|
8
|
Schmidt CE and Leach JB: Neural tissue
engineering: Strategies for repair and regeneration. Annu Rev
Biomed Eng. 5:293–347. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He B, Liu SQ, Chen Q, et al:
Carboxymethylated chitosan stimulates proliferation of Schwann
cells in vitro via the activation of the ERK and Akt signaling
pathways. Eur J Pharmacol. 667:195–201. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yazdani SO, Golestaneh AF, Shafiee A, et
al: Effects of low level laser therapy on proliferation and
neurotrophic factor gene expression of human schwann cells in
vitro. J Photochem Photobiol B. 107:9–13. 2012. View Article : Google Scholar
|
|
11
|
Gambarotta G, Fregnan F, Gnavi S and
Perroteau I: Neuregulin 1 role in Schwann cell regulation and
potential applications to promote peripheral nerve regeneration.
Int Rev Neurobiol. 108:223–256. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hu J, Zhu QT, Liu XL, Xu YB and Zhu JK:
Repair of extended peripheral nerve lesions in rhesus monkeys using
acellular allogenic nerve grafts implanted with autologous
mesenchymal stem cells. Exp Neurol. 204:658–666. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sowa Y, Imura T, Numajiri T, Nishino K and
Fushiki S: Adipose-derived stem cells produce factors enhancing
peripheral nerve regeneration: Influence of age and anatomic site
of origin. Stem Cells Dev. 21:1852–1862. 2012. View Article : Google Scholar
|
|
14
|
Manuelpillai U, Tchongue J, Lourensz D, et
al: Transplantation of human amnion epithelial cells reduces
hepatic fibrosis in immunocompetent CCl4-treated mice.
Cell Transplant. 19:1157–1168. 2010. View Article : Google Scholar
|
|
15
|
Miki T, Marongiu F, Dorko K, Ellis EC and
Strom SC: Isolation of amniotic epithelial stem cells. Curr Protoc
Stem Cell Biol. 1:1E.32010.
|
|
16
|
Murphy S, Rosli S, Acharya R, et al:
Amnion epithelial cell isolation and characterization for clinical
use. Curr Protoc Stem Cell Biol. 1:1E.62010.
|
|
17
|
Parolini O, Alviano F, Bagnara GP, et al:
Concise review: Isolation and characterization of cells from human
term placenta: Outcome of the first international Workshop on
Placenta Derived Stem Cells. Stem Cells. 26:300–311. 2008.
View Article : Google Scholar
|
|
18
|
Tsutsumi S, Shimazu A, Miyazaki K, et al:
Retention of multilineage differentiation potential of mesenchymal
cells during proliferation in response to FGF. Biochem Biophys Res
Commun. 288:413–419. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Toda A, Okabe M, Yoshida T and Nikaido T:
The potential of amniotic membrane/amnion-derived cells for
regeneration of various tissues. J Pharmacol Sci. 105:215–228.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Akle CA, Adinolfi M, Welsh KI, Leibowitz S
and McColl I: Immunogenicity of human amniotic epithelial cells
after transplantation into volunteers. Lancet. 2:1003–1005. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fatimah SS, Ng SL, Chua KH, et al: Value
of human amniotic epithelial cells in tissue engineering for
cornea. Hum Cell. 23:141–151. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Weinstein DE and Wu R: Isolation and
purification of primary Schwann cells. Curr Protoc Neurosci.
3:3.172001.
|
|
23
|
Baloh RH, Enomoto H, Johnson EJ and
Milbrandt J: The GDNF family ligands and receptors - implications
for neural development. Curr Opin Neurobiol. 10:103–110. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rich KM, Yip HK, Osborne PA, Schmidt RE
and Johnson EJ Jr: Role of nerve growth factor in the adult dorsal
root ganglia neuron and its response to injury. J Comp Neurol.
230:110–118. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Taniuchi M, Clark HB and Johnson EJ:
Induction of nerve growth factor receptor in Schwann cells after
axotomy. Proc Natl Acad Sci USA. 83:4094–4098. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang L, Lv X, Tong X, Jia H and Li Z:
Study on molecular mechanism for improving neural regeneration
after repair of sciatic nerve defect in rat by acellular nerve
allograft. Synapse. 66:52–60. 2012. View Article : Google Scholar
|
|
27
|
Zheng C, Zhu Q, Liu X, et al: Effect of
platelet-rich plasma (PRP) concentration on proliferation,
neurotrophic function and migration of Schwann cells in vitro. J
Tissue Eng Regen Med. May 31–2013.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu Y, Liu L, Li Y, et al: Myelin-forming
ability of Schwann cell-like cells induced from rat adipose-derived
stem cells in vitro. Brain Res. 1239:49–55. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kingham PJ, Kalbermatten DF, Mahay D, et
al: Adipose-derived stem cells differentiate into a Schwann cell
phenotype and promote neurite outgrowth in vitro. Exp Neurol.
207:267–274. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xue S, Chen C, Dong W, et al: Therapeutic
effects of human amniotic epithelial cell transplantation on
double-transgenic mice co-expressing APPswe and PS1ΔE9-deleted
genes. Sci China Life Sci. 55:132–140. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Banerjee A, Nürnberger S, Hennerbichler S,
et al: In toto differentiation of human amniotic membrane towards
the Schwann cell lineage. Cell Tissue Bank. 15:227–239. 2014.
View Article : Google Scholar
|
|
32
|
Miki T, Lehmann T, Cai H, Stolz DB and
Strom SC: Stem cell characteristics of amniotic epithelial cells.
Stem Cells. 23:1549–1559. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ilancheran S, Michalska A, Peh G, et al:
Stem cells derived from human fetal membranes display multilineage
differentiation potential. Biol Reprod. 77:577–588. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hou Y, Huang Q, Liu T and Guo L: Human
amnion epithelial cells can be induced to differentiate into
functional insulin-producing cells. Acta Biochim Biophys Sin
(Shanghai). 40:830–839. 2008. View Article : Google Scholar
|
|
35
|
Miki T and Strom SC: Amnion-derived
pluripotent/multipotent stem cells. Stem Cell Rev. 2:133–142. 2006.
View Article : Google Scholar
|
|
36
|
Elwan MA and Sakuragawa N: Evidence for
synthesis and release of catecholamines by human amniotic
epithelial cells. Neuroreport. 8:3435–3438. 1997. View Article : Google Scholar
|
|
37
|
Wei JP, Zhang TS, Kawa S, et al: Human
amnion-isolated cells normalize blood glucose in
streptozotocin-induced diabetic mice. Cell Transplant. 12:545–552.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sakuragawa N, Enosawa S, Ishii T, et al:
Human amniotic epithelial cells are promising transgene carriers
for allogeneic cell transplantation into liver. J Hum Genet.
45:171–176. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Simat SF, Chua KH, Abdul Rahman H, Tan AE
and Tan GC: The stemness gene expression of cultured human amniotic
epithelial cells in serial passages. Med J Malaysia. 63(Suppl A):
53–54. 2008.PubMed/NCBI
|
|
40
|
Tan GC, Simat SF, Abdul Rahman H, Tan AE
and Chua KH: Quantitative RT PCR approach to evaluate the
neurogenic and gliagenic gene expression of cultured human amniotic
epithelial cells. Med J Malaysia. 63(Suppl A): 51–52.
2008.PubMed/NCBI
|
|
41
|
Zhou X, He X, He B, et al: Etifoxine
promotes glial-derived neurotrophic factor-induced neurite
outgrowth in PC12 cells. Mol Med Rep. 8:75–80. 2013.PubMed/NCBI
|