Introduction
Ovarian cancer is one of the most serious malignant
tumors that threaten the health of women, and the 5-year survival
rate in advanced ovarian cancer patients is estimated at only at
30.6% (1). Therefore, improving
the survival rate of these patients is a major clinical issue.
Since the 1990s, research studies have shed light on immunotherapy,
which may be the most important tool of the 21st century (2). The regulatory functions of
CD4+ CD25+ regulatory T (Treg) cells in the
maintainance of immune homeostasis, tumor immunity, allergic
reactions and microbial infection are well established (e.g.,
3). Active CD4+
CD25+ Treg cells can effectively inhibit the functions
of natural killer (NK) cells, B cells and dendritic cells (DCs)
based on cell-cell contact mechanisms or via the production of
soluble factors, such as transforming growth factor-β (TGF-β) and
interleukin-10 (IL-10) (4).
The expression of indoleamine 2,3-dioxygenase (IDO)
has been shown to be significantly increased in a variety of tumor
cells (5). Moreover, a recent
study reported that IDO can suppress the immune function of T cells
by inducing the differentiation of naive T cells to Treg cells
(6). A number of studies have
focused on the role of IDO in cancer development and therapy. Mei
et al (7) reported that
IDO1 enhances survival and invasiveness of endometrial stromal
cells via the activation of the JNK signaling pathway. Chen et
al (8) demonstrated that
attenuation of immune suppression via inhibition of the IDO1
enzymatic activity may be an important mechanism underlying
polyphenol-mediated chemoprevention or combinatorial cancer
therapy. In addition, a previous study reported that certain
phytochemicals markedly reduce the IDO1 activity, and that this
inhibition may at least in part explain their anti-cancer
properties (9). Furthermore, Wang
et al (10) revealed that
downregulation of IDO controls ovarian cancer progression by
activating NK cells, and proposed that IDO may be potentially
useful in the therapy of ovarian cancer. de Jong et al
(11) found that IDO-induced
immune escape may play an important role in ovarian cancer.
1-Methyl-D-tryptophan may promote anti-tumor immune escape by
increasing the IDO1 level in cancer cells (12). It is generally believed that the
combination of IDO and DCs is the major cause of tumor cell immune
tolerance induced by Treg cell proliferation (13). Due to the important roles played by
IDO and Treg cells, an important body of research has focused on
the identification of factors that may affect their activity,
including hypoxia. Hypoxia is considered one of the basic features
of the tumor microenvironment in the body (14). In the hypoxic environment, the
ovarian cancer cell adhesion ability was shown to be decreased,
while invasive ability is increased, inducing peritoneal metastases
or recurrence (15). Although a
number of studies have been published on hypoxia, the relationship
and interaction between the tumor hypoxic microenvironment and
tumor immunity still remains unclear.
In this study, the expression of IDO in ovarian
cancer cells was inhibited by hypoxia and enhanced by Treg cells.
In addition, the expression of interleukin-2 (IL-2), interferon-γ
(IFN-γ), perforin, IL-10 and TGF-β was significantly changed in
cultures containing Treg cells under hypoxic conditions.
Furthermore, our study indicated that Treg cells may significantly
enhance ovarian cancer cell apoptosis and invasive ability,
especially in hypoxia. Overall, our study explored the different
effects of IDO and Treg cells on ovarian cancer cells under hypoxic
conditions, and suggests that targeting IDO and Treg cels may
constitute a suitable therapeutic route for ovarian cancer.
Materials and methods
Cell cultures and study groups
The epithelial ovarian cancer cell line SKOV3-IP was
provided the by Institute of Obstetrics and Gynecology Hospital at
Fudan University. Treg cells, NK cells and cytotoxic T lymphocytes
(CTLs) were derived from peripheral blood of healthy adult
females.
SKOV3-IP cells (106/ml) were inoculated
with Dulbecco’s modified Eagle’s medium with Nutrient Mixture F-12
(DMEM-F12) supplemented with 10% Gibco® fetal bovine
serum (FBS) and Gibco® 1% penicillin/streptomycin (all
from Thermo Fisher Scientific, Waltham, MA, USA), and cultured at
37°C, in a 5% CO2 incubator. The medium was replaced
every other day. After cells had reached 80–90% confluence, they
were digested by a 0.25% trypsin-ethylene diamine tetraacetic acid
solution (Gibco®, Thermo Fisher Scientific) and
transferred to a new flask. Aerobically cultured cells were placed
in a 37°C incubator (95% air, 5% CO2). Hypoxia-cultured
cells were sealed in an anaerobic culture tank (1% O2,
5% CO2 and 94% N2 ) at 37°C.
The cells were divided into 6 groups: SKOV3-IP;
SKOV3-IP and Treg cells; SKOV3-IP and CTLs; SKOV3-IP and NK cells;
SKOV3-IP co-cultured with CTL and Treg cells; and SKOV3-IP
co-cultured with NK and Treg cells.
Reverse transcription- polymerase chain
reaction (RT-PCR)
Total RNA was extracted using the Invitrogen™ TRIzol
reagent (Thermo Fisher Scientific) following the manufacturer’s
instructions, and the quantity of RNA was analyzed by UV
spectrophotometry. RNA (4 μg) was reverse transcribed to cDNA with
Moloney murine leukemia virus reverse transcriptase in a 30-μl
reaction volume using oligo (dT)18 primers, RNase inhibitor and
buffers from the All-in-oneTM First-strand cDNA synthesis kit
(GeneCopoeia, Rockville, MD, USA), following the manufacturer’s
instructions. The synthesized cDNA was used for PCR, conducted on a
DNA thermocycler (Takara Bio, Inc., Shiga, Japan) with the
following conditions: Initial denaturation at 95°C for 3 min, and
then 35 cycles amplication (95°C for 20 sec, 60°C for 30 sec and
72°C for 30 sec. The reaction was performed on a 25-μl volume,
containing Taq DNA polymerase and PCR buffer provided in the
Phusion Blood Direct PCR kit (Finnzymes, Espoo, Finland), a dNTP
mix and primers for each gene. The glyceraldehyde-3-phosphate
dehydrogenase gene (GAPDH) was used as the internal control.
Primer sequences are listed in Table
I. Quantitative PCR was conducted on an iQ5 Multicolor
Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA,
USA) using a SYBR Green Real-time PCR Master Mix (Takara Bio,
Inc.). Data were calculated using the 2−ΔΔCt method
normalized to the individual internal control level.
 | Table IPrimers used in the present
study. |
Table I
Primers used in the present
study.
| Primer | Forward | Reverse |
|---|
| IDO |
5′-TTTGCTAAAGGCGCTGTTGG-3′ |
5′-CCTTCATACACCAGACCGTCTGA-3′ |
| GAPDH |
5′-CGGAGTCAACGGATTTGGTCGATA-3 |
5′-AGCCTTCTCCATGGTTGGTGAACAC-3′ |
Western blot analysis
The treated cells were washed twice with cold
Gibco® phosphate-buffered saline (Thermo Fisher
Scientific) and were lysed in RIPA buffer (Biocolor BioScience
& Technology Co., Ltd., Shanghai, China) in the presence of a
proteinase inhibitor (Kangchen Bio-tech, Shanghai, China). Protein
quantification was performed with the BCA-100 protein assay kit
(Biocolor BioScience & Technology Co., Ltd.). Samples were
subjected to 8% sodium dodecyl sulphate polyacrylamide gel
electrophoresis, transferred to polyvinylidene fluoride (PVDF)
membranes (EMD Millipore, Billerica, MA, USA) and blocked with 5%
fat-free milk for 2 h at room temperature. Equal loading in each
blot was confirmed by Coomassie staining (Beyotime Biotechnology,
Shanghai, China) of the membrane. The membrane was incubated
overnight with a mouse anti-human IDO monoclonal antibody (1:500;
Abcam, Cambridge, MA, USA) and mouse anti-human GAPDH monoclonal
antibody (1:5,000; Cell Signaling Technology, Inc., Beverly, MA,
USA) at 4°C. Then, the membrane was incubated with the horseradish
peroxidase (HRP)-labeled secondary antibody (1:7,000) for 1 h at
room temperature. Signals were visualized with an enhanced
chemiluminescence kit (Amersham Pharmacia Biotech, Piscataway, NJ,
USA) and quantified using an Odyssey® imaging system
(LI-COR Inc., Lincoln, NE, USA). The protein level of IDO was
normalized to the level of GAPDH.
Enzyme-linked immunosorbent assay
(ELISA)
TGF-β, IFN-γ, IL-2, IL-10 and perforin levels were
detected in the supernatant of cell culture medium by ELISA using
corresponding kits (eBioscience, San Diego, CA, USA) according to
the manufacturer’s instructions. The absorbance of the samples was
measured on an ELISA plate reader (eBioscience) at a 450 nm
wavelength, with a 610–630 nm reference filter. The concentration
was determined by standard curve analysis, based on the absorbance
of respective standards.
Apoptotic assay of ovarian cancer
cells
To analyze the apoptosis of SKOV3-IP cells, we used
the Invitrogen™ Apoptosis Assay kit (Thermo Fisher Scientific)
according to the manufacturer’s instructions. Briefly,
~1×105 SKOV3-IP cells were cultured in a 12-well plate.
In parallel, co-cultures with 5×105 Treg cells,
1×106 NK cells and 1×106 CTLs per well were
established. There were three replicates for each group. The cells
in each group were then subjected to different conditions i.e.,
72-h aerobic growth, 48-h normoxia and 24-h hypoxia, 24-h aerobic
and 48-h anaerobic growth, and 72-h hypoxia. Next, the supernatant
and the suspended cells were discarded and the cells were
dissociated with trypsin to obtain a single-cell suspension. Flow
cytometry (BD Biosciences, Franklin Lakes, NJ, USA) was used to
detect the apoptotic rate of ovarian cancer cells under the
different culture conditions.
Invasive ability of ovarian cancer
cells
The migratory ability of ovarian cancer cells was
assessed using Transwell chambers and Costar® cell
culture plates (all from Corning, Tewksbury, MA, USA). The
transwell chambers were placed in a 24-well plate. The bottom of
the chambers was coated with Matrigel (BD Biosciences),
2×105 SKOV3-IP cells per well were plated in the upper
chambers of the 24-well Transwell chamber and then 600 μl DMEM-F12
supplemented with 10% FBS were added to the lower chambers.
Following incubation for 24 h at 37°C in 5% CO2, cells
located on the upper membranes were removed with cotton swabs. The
cells that had invaded the lower surface of the membrane were fixed
in ethanol and stained with crystal violet. Images of invading
cells were acquired under a Leica DC 300F microscope (Olympus,
Tokyo, Japan) in five random fields (magnification, ×100). The
invasive ability of the cells in each group was determined by the
average number of invading cells in the five fields.
Statistical analysis
All experiments were repeated at least three times
and each experiment was performed at least in duplicate. The
results were presented as mean ± standard deviation (SD).
Statistical analysis was performed using a one-way analysis of
variance (ANOVA) and χ2 tests, implemented in the SPSS
11.5 (SPSS Inc., Chicago, IL, USA) or Excel (Microsoft, Bellevue,
WA, USA) software. P<0.05 was considered to indicate
statistically significant differences.
Results
Expression of the IDO mRNA under
different conditions
The IDO mRNA expression level showed a
decreasing trend (P<0.05) with prolonged exposure to hypoxia in
SKOV3-IP (Fig. 1A), and in
SKOV3-IP + CTL cells (P<0.05) (Fig.
1B). When SKOV3-IP cells were co-cultured with NK cells, the
IDO mRNA expression level was slightly increased in the 24-h
hypoxia group and was again decreased at 48 and 72 h (Fig. 1C). In the co-cultured system of
SKOV3-IP and Treg cells, the level was decreased in the early
hypoxia and increased at 72 h of hypoxia (Fig. 1D); similar changes were observed in
the SKOV3-IP + CTL + Treg group (Fig.
1E). In addition, a slight fluctuation in the IDO level
was observed in the SKOV3-IP + NK + Treg group (Fig. 1F).
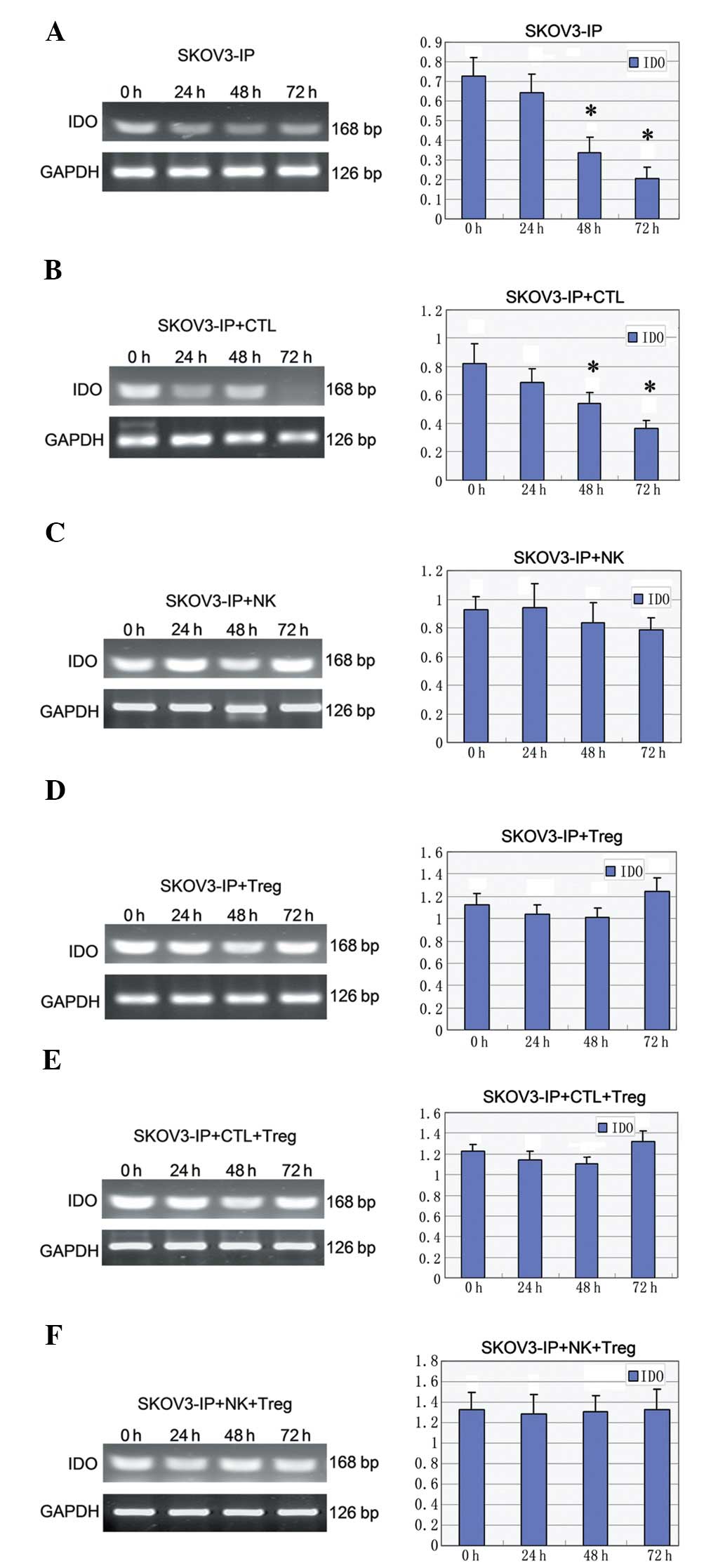 | Figure 1Expression of the IDO mRNA
under different conditions. IDO levels in the (A) SKOV3-IP,
(B) SKOV3-IP + CTL, (C) SKOV3-IP + NK, (D) SKOV3-IP + Treg, (E)
SKOV3-IP + CTL + Treg, and (F) SKOV3-IP + NK + Treg groups.
*P<0.05 vs. 0 h; IDO, indoleamine 2,3-dioxygenase;
CTL, cytotoxic T lymphocytes; NK, natural killer cells; Treg,
regulatory T cells; GAPDH, glyceraldehyde-3-phosphate
dehydrogenase. |
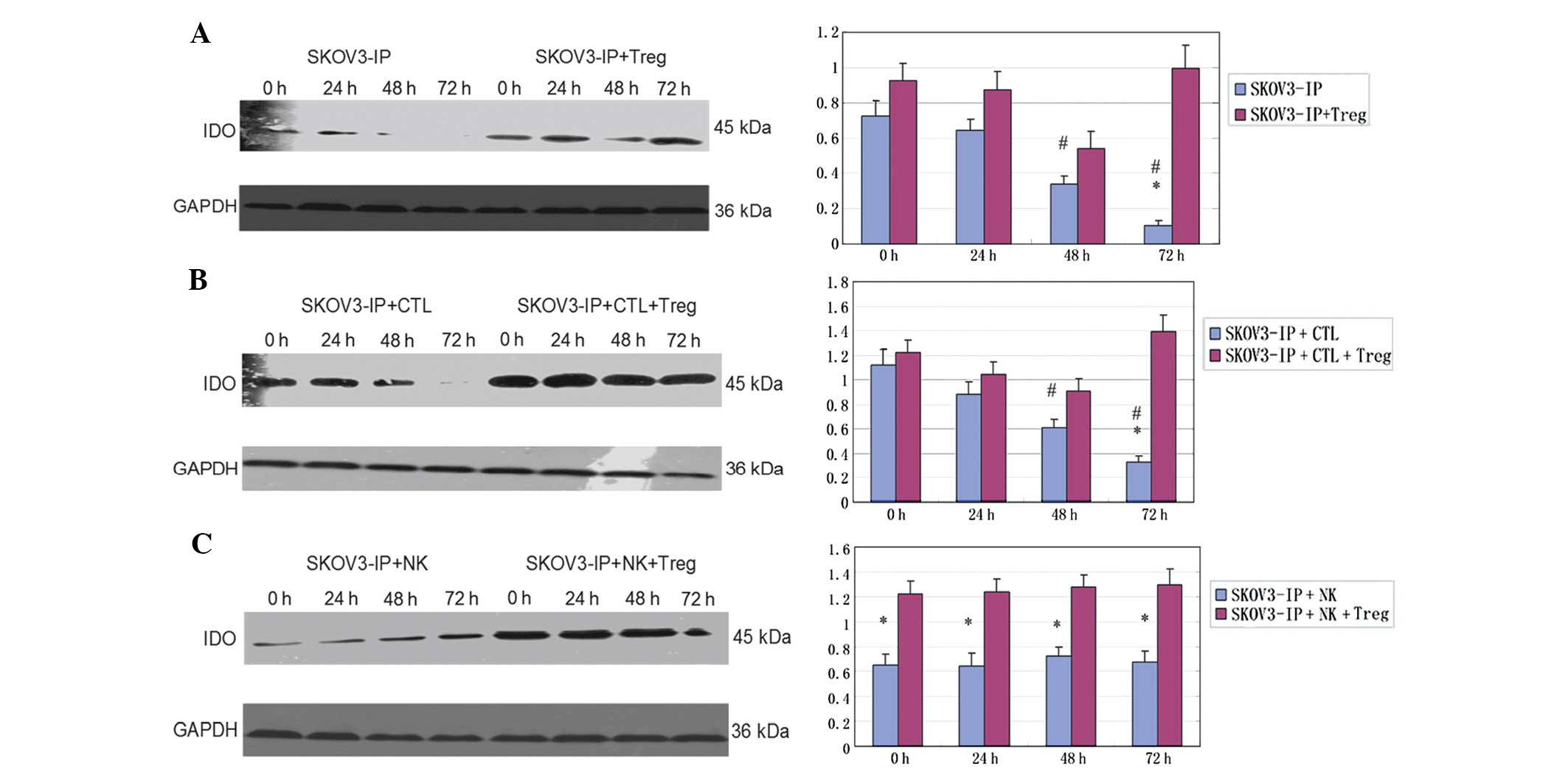
Expression of the IDO protein
The expression of the IDO protein was significantly
decreased in SKOV3-IP cells along the time of exposure to hypoxia
(P<0.05). The IDO level was higher in the SKOV3-IP + Treg group
compared to the group of SKOV3-IP cells, and decreased within 48 h,
then increased again at 72 h (P<0.05) (Fig. 2A). It is notable that a similar
profile was observed in the SKOV3-IP group co-cultured with CTLs
and the SKOV3-IP + CTL + Treg group (Fig. 2B). When SKOV3-IP cells were
cultured with NK cells, the expression level of IDO slightly
changed with the extension of exposure to hypoxia (P>0.05).
Addition of Treg cells markedly enhanced the expression of IDO
protein (P<0.05), but this effect only slightly changed with
time (P>0.05) (Fig. 2C).
The effect of Treg on ovarian cancer
cells
In order to investigate the effect of Treg cells on
the immunity of ovarian cancer cells, we measured the
concentrations of TGF-β, IFN-γ, IL-2, IL-10 and perforin in the
cell supernatants using ELISA.
The concentration of IL-2 in the SKOV3-IP + CTL
group with or without Treg cells was decreased with the extension
of exposure to hypoxia, and this decrease was significant when Treg
cells were present (P<0.05) (Fig.
3A). In the co-cultured systems of SKOV3-IP + NK cells, the
concentration of IL-2 fluctuated along the time of exposure to
hypoxia, with no statistically significant changes observed
(P>0.05) (Fig. 3A). In
addition, the secretion of IL-10 in SKOV3-IP cells alone, or
co-cultured with CTL or NK cells did not change during the exposure
to hypoxia (P>0.05) (Fig. 3B).
By contrast, the IL-10 level was found significantly and
time-dependently increased when Treg cells were present (P<0.05)
(Fig. 3B).
 | Figure 3The effect of Treg on the expression
of different proteins in ovarian cancer cells. (A, D and E)
Expression of IL-2, IFN-γ and perforin in the SKOV3-IP + CTL and
SKOV3-IP + NK groups, respectively, with or without Treg cells. (B
and C) Concentration of IL-10 and TGF-β in the SKOV3-IP, SKOV3-IP +
CTL and SKOV3-IP + NK groups, respectively with or without Treg
cells. *P<0.05 vs. group with Treg cells;
#P<0.05 vs. 0 h; Treg, regulatory T cells; IL-2,
interleukin-2; IFN-γ, interferon-γ; CTL, cytotoxic T lymphocytes;
NK, natural killer cells; IL-10, interleukin-10; TGF-β,
transforming growth factor-β. |
The concentration of TGF-β first increased and then
decreased at 24 and 72 h, respectively; this trend was observed in
SKOV3-IP, SKOV3-IP + CTL and SKOV3-IP + NK cells (Fig. 3C). When Treg cells were present,
the TGF-β level increased with the extension of hypoxia (P<0.05)
(Fig. 3C). In addition, the IFN-γ
level was decreased in the SKOV3-IP + CTL group and this decrease
was enhanced when Treg cells was present (P<0.05) (Fig. 3D). NK cells enhanced the secretion
of IFN-γ (P<0.05), and Treg cells inhibited the secretion of
IFN-γ; this inhibition become more apparent in the hypoxic state
(P<0.05) (Fig. 3D). The
expression profile of perforin was similar to that of IFN-γ
(Fig. 3E).
Apoptosis of ovarian cancer cells under
different conditions
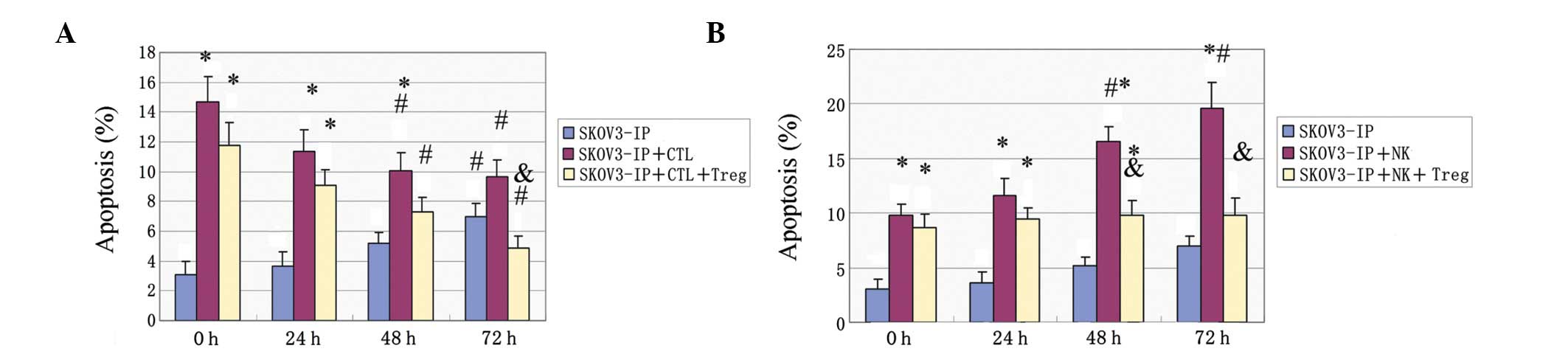
The apoptotic rate of SKOV3-IP cells was increased
with the extension of exposure to hypoxia (P<0.05). In addition,
CTLs were found to significantly enhance apoptosis under normoxic
conditions (P<0.05). This effect was attenuated with prolonged
hypoxia (P<0.05) and was also significant when Treg cells were
present (P<0.05) (Fig. 4A). NK
cells enhanced SKOV3-IP apoptosis, and this effect was further
enhanced with the hypoxic time extension (P<0.05) (Fig. 4B). When Treg cells were added in
the culture, the effect was reduced (P<0.05), but was only
slightly changed at different time-points (Fig. 4B).
Invasive ability of ovarian cancer cell
lines under normoxic and hypoxic conditions
The number of invading cells was increased when
SKOV3-IP cells were co-cultured with Treg cells under normoxic or
hypoxic conditions. In addition, hypoxia significantly enhanced the
invasive ability of SKOV3-IP cells co-cultured with Treg cells
(P=0.00109) or cultured alone (P=0.003171) (Table II).
| Table IIEffect of hypoxia and Treg cells on
the invasive ability of ovarian cancer cells. |
Table II
Effect of hypoxia and Treg cells on
the invasive ability of ovarian cancer cells.
| No. of migrating
cells | SKOV3-IP | SKOV3-IP +
Treg |
|---|
| Normoxia | 16.77±5.84 | 38.77±11.26 |
| Hypoxia | 33.66±9.73 | 89.47±22.45 |
| P-value | 0.003171 | 0.001090 |
Discussion
The expression of IDO in ovarian cancer cells showed
a significant trend to decrease at the mRNA and protein level along
the time of exposure to hypoxic conditions (Figs. 1 and 2), and these findings are consistent with
a previous study (16). The effect
of hypoxia on ovarian cancer cell apoptosis and invasive ability
was further investigated. The apoptotic rate of ovarian cancer
cells was significantly increased under hypoxic conditions
(Fig. 4). Previous studies have
shown that hypoxia inhibits DNA synthesis, induces cell cycle
arrest at the G0-G1 phase, as well as expression changes in
cell-cycle proteins. These changes included an increase in p27
expression, a decrease in Rb expression, reduction in the levels of
cyclin D1 and E leading to cell-cycle arrest, and inhibition of
ovarian cancer cell proliferation. It was further shown that these
changes were reverted under normoxia (17,18).
Hypoxia was shown to induce endothelial cell
apoptosis through nuclear factor-κB, and to mediate Bcl-2
suppression in vivo (19).
In our study, hypoxia was found to increase the invasive ability of
ovarian cancer cells (Table II).
Tumor invasion and metastasis are indicators of the degree of tumor
malignancy. The hypoxic microenvironment may induce the expression
of genes related to tumor cell invasion, while also reducing cell
adhesion and increasing cell motility and invasiveness to promote
tumor metastasis (20).
When Treg cells were added to the cultures, the
expression level of IDO was increased at both the mRNA and protein
level (Figs. 1 and 2). There is some debate on whether Treg
cells can promote the expression of IDO, and our findings support
the theory that Treg cells enhance IDO expression. In addition, our
results showed that IDO expression is reduced by addition of Treg
cells during early hypoxia (24–48 h) and is significantly increased
at 72 h of hypoxia. This result suggests that the direct effect of
hypoxia is to inhibit IDO expression, and that lowly expressed IDO
may stimulate immune cells to produce cytokines. Our results are
consistent with the findings of Munn and Mellor (21).
To study the effect of Treg cells on the immunity of
ovarian cancer cells, we quantified the secretion of the cytokines
TGF-β, IFN-γ, IL-2, IL-10 and perforin in the different co-culture
groups (Fig. 3). IL-2 plays a
crucial role in the maintenance of natural immunologic
self-tolerance (22). IL-10, a
cytokine with anti-inflammatory properties, has an important role
in infection by inhibiting the immune response to pathogens
(23). TGF-β has been found to
function in vascular development and vascular homeostasis
maintenance (24). IFN-γ was
reported to play a crucial role in autoimmunity (25). Unlike relatively redundant
individual granzymes, functional perforin is essential to cytotoxic
lymphocyte function and immune regulation in the host (26). In our study, the levels of IFN-γ,
IL-2, IL-10 and perforin were decreased when Treg cells were
present in the culture. This finding indicates that Treg cells
induce changes in the expression of cytokines; this is likely an
immune-escape mechanism. Importantly, Treg cells can suppress
immune responses and play an important role in the dominant immune
escape process in early tumor progression (27).
In addition, the apoptosis of SKOV3-IP cells was
studied under different conditions (Fig. 4). Apoptosis plays a crucial role in
the pathogenesis of a variety of cardiovascular diseases that are
caused by the loss of terminally differentiated cardiac myocytes
(28). In addition, in the absence
of adequate vasculature, tumor cells undergo hypoxia and
starvation, followed by apoptosis. A previous study reported that
hypoxia promotes tolerance and angiogenesis via recruiting Treg
cells (29). In our study, the
promoting effect of CTLs on apoptosis was inhibited by hypoxia,
while the effect of NK cells was enhanced under hypoxic conditions.
This may be due to changes in the activity of these cells in the
hypoxic environment. Moreover, Treg cells significantly inhibited
the cytotoxicity of CTLs, an effect that was more obvious under
hypoxic conditions. Treg cells also inhibited the effect of NK
cells, but this effect slightly changed between normoxia and
hypoxia. Our findings on the effects of Treg cells on cancer cell
apoptosis are consistent with a previous study, which showed that
Treg cells inhibit the function of NK cells, B cells and other
immunocytes (30). We propose that
these effects are caused by the increased activity of Treg cells in
the hypoxic state.
Intraperitoneal dissemination and distant metastasis
constitute important complications in ovarian cancer treatment, and
are closely related to the invasion of malignant cells (31). Despite the advances in chemotherapy
and considerable efforts made to improve early detection,
metastasis remains a major challenge in the clinical management of
ovarian cancer (31). Hypoxia was
demonstrated to reduce ovarian cancer cell adhesion, and promote
cancer cell invasion and metastasis (32). Our experiments confirmed that Treg
cells increase the number of invading cells by enhancing the
invasive ability of ovarian cancer cells under normoxia or hypoxia.
However, this enhancing effect was stronger in hypoxia compared to
normoxia. In addition, the invasive ability of SKOV3-IP cells was
significantly higher in hypoxic compared to normoxic conditions,
independently of the presence of Treg cells. Moreover, the SKOV3-IP
invasive ability was more enhanced when co-culturing with Treg
cells in hypoxia than in any other condition. Our results suggest
that Treg cells and hypoxia may induce the immune escape and
ovarian cancer cell metastasis, as previously proposed in other
studies (33,34).
In summary, in the ovarian cancer microenvironment,
IDO and Treg cells may mutually enhance their levels and
synergistically act to attenuate the cytotoxic effect of CTLs and
NK cells. These events are enhanced when cells are cultured under
hypoxic conditions, which indicates that oxygen depletion plays a
key role in the immune tolerance and escape. Our findings are
helpful for improving the effects of cancer immunotherapy via the
amelioration of the hypoxic microenvironment of malignant tumors.
However, further investigation is needed to study the effect of
hypoxia on immune escape in vivo.
Acknowledgements
This study was supported by grants from the National
Nature Science Foundation of China (no. 81001150) and the Shanghai
Science and Technology Department Funds (no. 10411960800).
Abbreviations:
|
NK
|
natural killer
|
|
DCs
|
dendritic cells
|
|
TGF-β
|
transforming growth factor-β
|
|
IL-10
|
interleukin-10
|
|
IDO
|
indoleamine 2,3-dioxygenase
|
|
CTL
|
cytotoxic T lymphocytes
|
|
RT-PCR
|
reverse transcription-polymerase chain
reaction
|
|
PVDF
|
polyvinylidene fluoride
|
|
HRP
|
horseradish peroxidase
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
SD
|
standard deviation
|
|
IFN-γ
|
interferon-γ
|
References
|
1
|
Fehrmann RS, Li XY, van der Zee AG, et al:
Profiling studies in ovarian cancer: a review. Oncologist.
12:960–966. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dougan M and Dranoff G: Immunotherapy of
cancer. Innate Immune Regulation and Cancer Immunotherapy. Wang RF:
Springer; New York: pp. 391–414. 2012, View Article : Google Scholar
|
|
3
|
Wållberg M, Wong FS and Green EA: An
islet-specific pulse of TGF-β abrogates CTL function and promotes β
cell survival independent of Foxp3+ T cells. J Immunol.
186:2543–2551. 2011. View Article : Google Scholar
|
|
4
|
Lu L, Zhou X, Wang J, Zheng SG and Horwitz
DA: Characterization of protective human
CD4+CD25+ FOXP3+ regulatory T
cells generated with IL-2, TGF-β and retinoic acid. PLoS One.
5:e151502010. View Article : Google Scholar
|
|
5
|
Teoh D and Secord AA: Antiangiogenic
agents in combination with chemotherapy for the treatment of
epithelial ovarian cancer. Int J Gynecol Cancer. 22:348–359. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stein P, Weber M, Prüfer S, et al:
Regulatory T cells and IL-10 independently counterregulate
cytotoxic T lymphocyte responses induced by transcutaneous
immunization. PLoS One. 6:e279112011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mei J, Li MQ, Ding D, et al: Indoleamine
2,3-dioxygenase-1 (IDO1) enhances survival and invasiveness of
endometrial stromal cells via the activation of JNK signaling
pathway. Int J Clin Exp Pathol. 6:431–444. 2013.PubMed/NCBI
|
|
8
|
Chen SS, Corteling R, Stevanato L and
Sinden J: Polyphenols inhibit indoleamine 3,5-dioxygenase-1
enzymatic activity - a role of immunomodulation in chemoprevention.
Discov Med. 14:327–333. 2012.PubMed/NCBI
|
|
9
|
Chen S, Corteling R, Stevanato L and
Sinden J: Natural inhibitors of indoleamine 3,5-dioxygenase induced
by interferon-γ in human neural stem cells. Biochem Biophys Res
Commun. 429:117–123. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang D, Saga Y, Mizukami H, et al:
Indoleamine-2,3-dioxygenase, an immunosuppressive enzyme that
inhibits natural killer cell function, as a useful target for
ovarian cancer therapy. Int J Oncol. 40:929–934. 2012.
|
|
11
|
de Jong RA, Nijman HW, Boezen HM, et al:
Serum tryptophan and kynurenine concentrations as parameters for
indoleamine 2,3-dioxygenase activity in patients with endometrial,
ovarian, and vulvar cancer. Int J Gynecol Cancer. 21:1320–1327.
2011.PubMed/NCBI
|
|
12
|
Opitz CA, Litzenburger UM, Opitz U, et al:
The indoleamine-2,3-dioxygenase (IDO) inhibitor
1-methyl-D-tryptophan upregulates IDO1 in human cancer cells. PLoS
One. 6:e198232011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fallarino F, Grohmann U, You S, et al: The
combined effects of tryptophan starvation and tryptophan
catabolites down-regulate T cell receptor ζ-chain and induce a
regulatory phenotype in naive T cells. J Immunol. 176:6752–6761.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Erkanli S, Bolat F, Kayaselcuk F, Demirhan
B and Kuscu E: COX-2 and survivin are overexpressed and positively
correlated in endometrial carcinoma. Gynecol Oncol. 104:320–325.
2007. View Article : Google Scholar
|
|
15
|
Schwandt A, Garcia JA, Elson P, et al:
Clinical and immunomodulatory effects of celecoxib plus
interferon-α in metastatic renal cell carcinoma patients with COX-2
tumor immunostaining. J Clin Immunol. 31:690–698. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hryniewicz A, Boasso A, Edghill-Smith Y,
et al: CTLA-4 blockade decreases TGF-β, IDO, and viral RNA
expression in tissues of SIVmac251-infected macaques. Blood.
108:3834–3842. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wolf AM, Wolf D, Steurer M, Gastl G,
Gunsilius E and Grubeck-Loebenstein B: Increase of regulatory T
cells in the peripheral blood of cancer patients. Clin Cancer Res.
9:606–612. 2003.PubMed/NCBI
|
|
18
|
Curiel TJ, Coukos G, Zou L, et al:
Specific recruitment of regulatory T cells in ovarian carcinoma
fosters immune privilege and predicts reduced survival. Nat Med.
10:942–949. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Matsushita H, Morishita R, Nata T, et al:
Hypoxia-induced endothelial apoptosis through nuclear factor-κB
(NF-κB)-mediated bcl-2 suppression: in vivo evidence of the
importance of NF-κB in endothelial cell regulation. Circ Res.
86:974–981. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
He X, Brenchley PE, Jayson GC, Hampson L,
Davies J and Hampson IN: Hypoxia increases heparanase-dependent
tumor cell invasion, which can be inhibited by antiheparanase
antibodies. Cancer Res. 64:3928–3933. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Munn DH and Mellor AL: Indoleamine
2,3-dioxygenase and tumor-induced tolerance. J Clin Invest.
117:1147–1154. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Setoguchi R, Hori S, Takahashi T and
Sakaguchi S: Homeostatic maintenance of natural Foxp3+
CD25+ CD4+ regulatory T cells by interleukin
(IL)-2 and induction of autoimmune disease by IL-2 neutralization.
J Exp Med. 201:723–735. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Saraiva M and O’Garra A: The regulation of
IL-10 production by immune cells. Nat Rev Immunol. 10:170–181.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Orlova VV, Liu Z, Goumans MJ and ten Dijke
P: Controlling angiogenesis by two unique TGF-β type I receptor
signaling pathways. Histol Histopathol. 26:1219–1230.
2011.PubMed/NCBI
|
|
25
|
Boniface K, Blumenschein WM, Brovont-Porth
K, et al: Human Th17 cells comprise heterogeneous subsets including
IFN-γ-producing cells with distinct properties from the Th1
lineage. J Immunol. 185:679–687. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Voskoboinik I, Dunstone MA, Baran K,
Whisstock JC and Trapani JA: Perforin: structure, function, and
role in human immunopathology. Immunol Rev. 235:35–54.
2010.PubMed/NCBI
|
|
27
|
Elpek KG, Lacelle C, Singh NP, Yolcu ES
and Shirwan H: CD4+CD25+ T regulatory cells
dominate multiple immune evasion mechanisms in early but not late
phases of tumor development in a B cell lymphoma model. J Immunol.
178:6840–6848. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee Y and Gustafsson AB: Role of apoptosis
in cardiovascular disease. Apoptosis. 14:536–548. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Facciabene A, Santoro S and Coukos G: Know
thy enemy: Why are tumor-infiltrating regulatory T cells so
deleterious? Oncoimmunology. 1:575–577. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Diao Z, Shi J, Zhu J, et al: TRAIL
suppresses tumor growth in mice by inducing tumor-infiltrating
CD4+CD25+ Treg apoptosis. Cancer Immunol
Immunother. 62:653–663. 2013. View Article : Google Scholar
|
|
31
|
Naora H and Montell DJ: Ovarian cancer
metastasis: integrating insights from disparate model organisms.
Nat Rev Cancer. 5:355–366. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tan DS, Agarwal R and Kaye SB: Mechanisms
of transcoelomic metastasis in ovarian cancer. Lancet Oncol.
7:925–934. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Facciabene A, Peng X, Hagemann IS, et al:
Tumour hypoxia promotes tolerance and angiogenesis via CCL28 and
T(reg) cells. Nature. 475:226–230. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Facciabene A, Motz GT and Coukos G:
T-regulatory cells: key players in tumor immune escape and
angiogenesis. Cancer Res. 72:2162–2171. 2012. View Article : Google Scholar : PubMed/NCBI
|