Introduction
Apoptosis is the major pathogenetic mechanism of
early tubular cell death in ischemia/reperfusion (I/R)-induced
acute kidney injury (AKI). Under I/R conditions, disruption of the
outer mitochondrial membrane potential and permeability, leading to
the release of multiple toxic proteins, has been implicated in
activating apoptotic signaling (1,2).
However, the cell death pathway exists in a state of dynamic
equilibrium between pro and anti-apoptotic effector molecules. The
inhibitor of apoptosis proteins (IAP), for example, negatively
regulate apoptotic signaling at a post-mitochondrial level. Among
the IAP family, X-linked inhibitor of apoptosis protein (XIAP)
appears to be the most potent caspase inhibitor by directly binding
and inhibiting caspase targets, including caspase-3, 7 and 9. In
addition, in response to apoptotic stimuli, the inhibitory function
of XIAP can be antagonized by second mitochondria-derived activator
of caspases (Smac)/direct IAP-binding protein with low PI (DIABLO)
(3–7), which is also released from
mitochondria as cytochrome c. In addition, interaction of XIAP with
Smac/DIABLO has been demonstrated to mediate apoptosis following
diverse insults, including ischemia (8–10),
oxidative stress (11) and
ultraviolet radiation (12,13).
Despite these findings, the role of Smac/DIABLO and XIAP in renal
I/R remains to be elucidated.
Heat shock protein 72 (HSP72), a major stress
inducible protein, functions as a molecular chaperone in protein
folding, transport and degradation. Previous studies from our
laboratory (Renal Section, Department of Medicine, Boston Medical
Center, Boston University, Boston, MA, USA) and elsewhere have
revealed that HSP72 protects renal epithelial cells from apoptosis
by reducing mitochondrial membrane injury and inhibiting
mitochondrial release of cytochrome c and apoptosis-inducing factor
(AIF) (7,14,15).
Furthermore, this evidence also suggests that HSP72 attenuates
renal fibrosis through inhibiting epithelial-to-mesenchymal
transition (16,17). Thus, induction of HSP72 may have
wide-ranging effects in the treatment of acute and chronic renal
injury. However, it remains to be elucidated whether HSP72 protects
against I/R-induced renal tubular cell injury through modulation of
Smac/DIABLO and XIAP signaling.
In the present study, it was hypothesized that HSP72
reduces mitochondrial Smac/DIABLO release, prevents XIAP
degradation and thereby promotes tubular cell survival in renal I/R
injury.
Materials and methods
Reagents and antibodies
Geranylgeranylacetone (GGA) was obtained from Eisai
China (Shanghai, China). Terminal deoxynucleotidyl
transferase-mediated dUTP nick end labeling (TUNEL) assay kits
(fluorescent), annexin V fluorescein isothiocyanate apoptosis
detection kits and protease inhibitors were obtained from
Calbiochem (San Diego, CA, USA). In addition, the following
antibodies were used: mouse anti-human HSP72 (1:1,000; Stressgen
Biotechnologies, Victoria, BC, Canada), rabbit anti-human XIAP
(1:1,000; BD Biosciences, San Jose, CA, USA), mouse anti-human
Smac/DIABLO (1:1,000; BD Biosciences), rabbit anti human pro
caspase 3 (1:500; Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA) and mouse anti-human β-actin (1:2,000; Boster, Wuhan, China).
Horseradish peroxidase-conjugated anti-mouse IgG and horseradish
peroxidase conjugated anti-rabbit IgG were obtained from Jackson
ImmunoResearch (West Grove, PA, USA). All remaining reagents were
purchased from Sigma-Aldrich (St. Louis, MO, USA).
Cell culture and treatment
An immortalized proximal tubule epithelial cell line
from normal adult human kidney (HK-2) was purchased from the
American Type Culture Collection (Rockville, MD, USA). Cells were
cultured at 37°C in a 5% carbon dioxide atmosphere in Dulbecco’s
modified Eagle’s medium mixed 1:1 (vol:vol) with F12 medium
(Invitrogen Life Technologies, Carlsbad, CA, USA) supplemented with
10% fetal bovine serum. Cells were grown to 70–80% confluence and
subjected to serum-deprivation for 24 h prior to experimental
manipulation.
Induction of HSP72
HSP72 protein content was enhanced by coinfecting
HK-2 cells with adenoviruses containing wild-type human HSP72 and
green fluorescent protein (AdvTR5/HSP72-GFP) located on separate
cistrons induced by a tetracyclin-regulated promoter (AdvCMV/tTA)
as described previously (16). To
induce optimal renal HSP72 expression, GGA was emulsified with 5%
gum arabic and 0.008% tocopherol and administered to rats as
previously described (16).
Briefly, rats received daily oral administration with 400 mg/kg
GGA, starting one day prior to surgery and continuing throughout
I/R or sham surgery. Control animals were administered the same
volume of components without GGA (vehicle).
Animals
The experiments were performed with adult male
Sprague-Dawley rats weighing 200–250 g maintained with free access
to water and standard food. Renal I/R studies were performed using
the protocol approved by the Animal Care and Use Committee of the
Sun Yat-sen University (Guangzhou, China). Rats were randomly
allocated into three groups: i) Sham-surgery controls
receiving vehicle (n=8); ii) I/R group receiving vehicle
(n=8); iii) I/R group receiving GGA (n=8).
Renal I/R injury was induced as previously described
(18). Briefly, rats were
anesthetized by an intraperitoneal injection of chloral hydrate
(350 mg/kg) and placed on a sterile disposable towel over a warming
pad. A midline incision was made and the renal pedicles were
bluntly dissected. The left renal pedicles were occluded with a
nontraumatic vascular clamp for 45 min and subsequently allowed to
reperfuse following removal of the clamp. The right kidney was
removed. The incision was closed with 3-0 silk and rats were
returned to cages to recover. The rats in the sham-surgery group
were treated in an identical manner, with the exception of clamping
of the renal pedicles. Animals were sacrificed following 24 h of
reperfusion. Subsequently, blood samples were collected by heart
puncture for measurement of creatinine and urea nitrogen and the
left kidney was harvested and subjected to further investigation as
described below.
Renal function examination
Blood urea nitrogen (BUN) and creatinine levels were
measured using a QuantiChrom BUN or Creatinine assay kit (BioAssay
Systems, Hayward, CA, USA).
Histological analysis and apoptosis
assay
Kidney tissues were fixed in 10% phosphate-buffered
formalin, embedded in paraffin, sectioned at 3 μm thickness and
then stained with hematoxylin and eosin or periodic acid-Schiff
(PAS). Histological examinations were performed in a blinded manner
and scores were calculated on the basis of the percentage of
damaged tubules in 20 randomly selected cortical tubules with
visible basement membranes at ×400 magnification (Axioplan 2
imaging; Carl Zeiss, Oberkochen, Germany). A five-point scale was
used: 0, Normal; 1, <10%; 2, 11–25%; 3, 25–50%; 4, 51–75% and 5,
>75%.
Apoptosis was quantified in histological sections
using a commercially available TACS TdT-Fluor In Situ
Apoptosis Detection kit (R&D Systems, Inc., Minneapolis, MN,
USA) according to the manufacturer’s instructions and our previous
study (16). Briefly,
paraffin-embedded kidney sections were deparaffinized,
permeabilized and rehydrated. Slides were incubated with a TUNEL
reaction mixture containing terminal deoxynucleotidyl transferase.
Positive staining was identified in the cell nucleus with DNA
fragmentation under confocal microscopy (Zeiss LSM 510 META; Carl
Zeiss) microscopy and expressed as apoptotic cells per high-power
field.
Western blot analysis
Kidney cortex and harvested cultured cells were
homogenized in lysis buffer supplemented with a protease inhibitor
cocktail (Cell Signaling Technology, Inc., Beverly, MA, USA).
Cytosolic protein fractions were obtained through incubation of
cells with digitonin buffer [10 mM
piperazine-N,N′-bis(2-ethanesulfonic acid), pH 6.8, 0.015% (wt/vol)
digitonin, 300 mM sucrose, 100 mM NaCl, 3 mM MgCl2, 5 mM
EDTA and 1 mM phenylmethylsulfonyl fluoride] for 10 min at 4°C
(7). The supernatants of tissue,
cell lysates and cytosolic protein extracts were extracted,
subjected to protein assay and mixed with sodium dodecyl sulfate
(SDS) loading buffer. Samples were loaded and separated by 12% SDS
polyacrylamide gels (SDS-PAGE), electrotransferred onto a
nitrocellulose membrane, blotted with the designated antibodies and
then detected by enhanced chemiluminescence (Amersham Pharmacia
Biotech, Amersham, UK). Densitometric quantification was performed
with the image analysis program (Fluorchem™ 8900; Alpha Innotech,
San Leandro, CA, USA).
Immunoprecipitation analysis
Cytosolic protein fractions were dissolved in
immunoprecipitation buffer (0.5–1 mg of protein/ml) as described
previously (17). The cell lysates
were incubated overnight at 4°C with a polyclonal rabbit antibody
directed against human XIAP (2 μg/mg protein/ml immunoprecipitation
buffer; BD Biosciences). The immunocomplexes were isolated by
incubation at 4°C with Protein A/G PLUS agarose beads (Pierce
Biotechnology Inc., Rockford, IL, USA) for 2 h, washed three times
with the immunoprecipitation buffer and analyzed using the
indicated antibody by SDS-PAGE and western blotting.
Statistical analysis
All results are expressed as the mean ± standard
error of the mean. Analysis was performed with standard statistical
software (SPSS 11.0; SPSS, Inc., Chicago, IL, USA). Comparison
among groups was performed using a one-way analysis of variance
followed by the Student-Newman-Keuls test. P<0.05 was considered
to indicate a statistically significant difference.
Results
GGA attenuates I/R-induced renal
injury
Previous studies from our laboratory and others have
demonstrated that orally administered GGA selectively enhances
expression of HSP72 in the kidney (16,19).
In order to assess the protective roles of GGA in acute kidney
injury, renal function was evaluated in a rat I/R injury model.
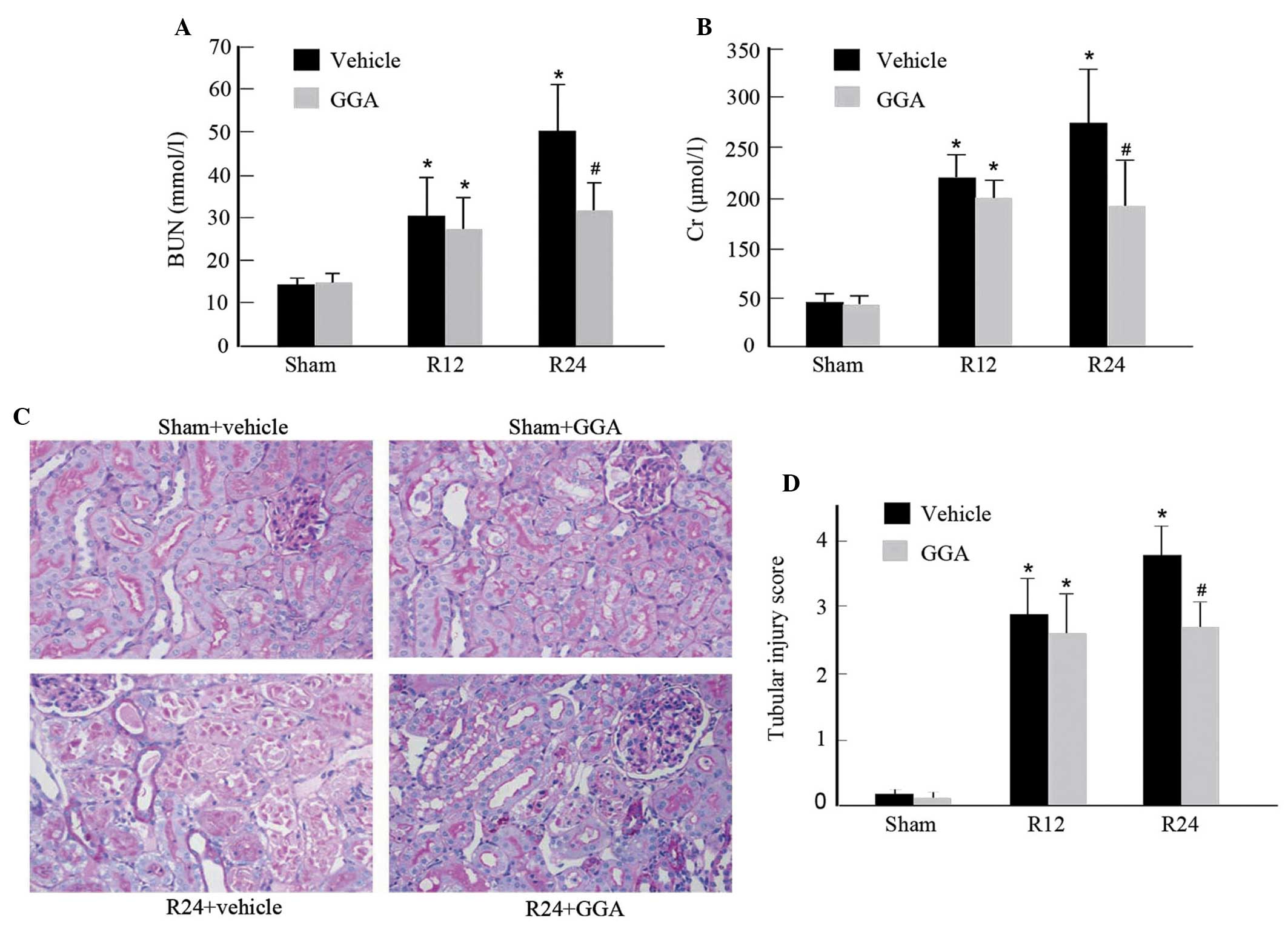
Compared with the sham-surgery group, the I/R rats with vehicle
alone exhibited marked and progressive elevation in the levels of
BUN and creatinine. By contrast, GGA administration significantly
improved renal dysfunction 24 h after reperfusion (Fig. 1A and B). Concordantly, histological
analysis of PAS staining revealed that I/R in the vehicle group
caused significant brush border loss, detached tubular epithelium,
cast formation and tubular dilation compared with the sham-surgery
group. However, GGA treatment exhibited significant improvements in
renal morphology (Fig. 1C) and
reduced the tubular injury score (Fig.
1D). Of note, GGA did not alter renal function and morphology
in rats subjected to sham I/R.
GGA inhibits apoptosis and XIAP
degradation
To examine the potential mechanism for the renal
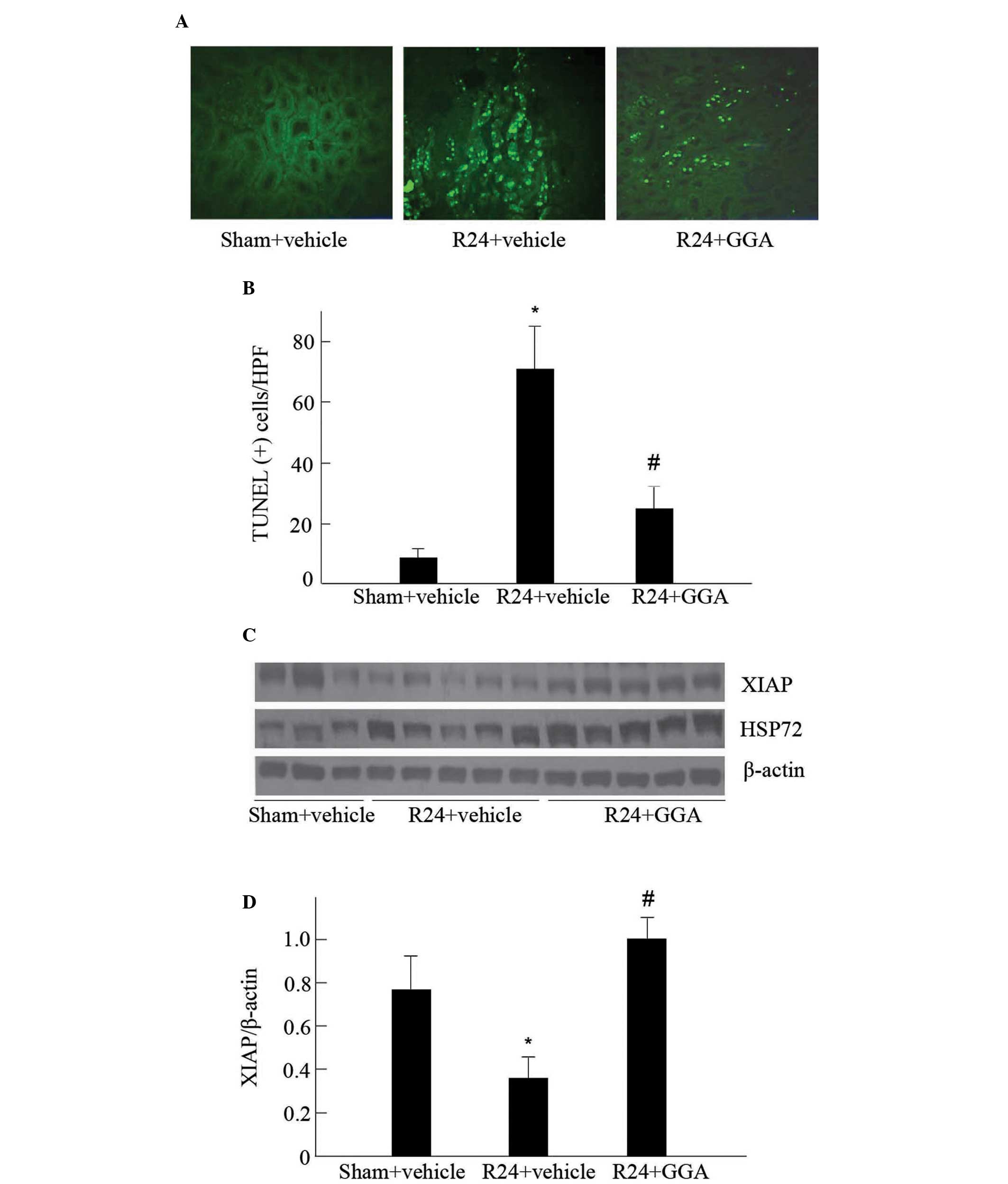
protective effect of GGA, the apoptosis of tubular cells was
detected using TUNEL staining. As shown in Fig. 2A, renal TUNEL-positive cells,
predominantly located at the proximal tubules, were increased in
vehicle-treated I/R kidneys, as compared with those of the
sham-surgery group. A significantly lower number of apoptotic cells
were observed in the GGA-treated kidneys subjected to I/R (Fig. 2A and B).
XIAP has been observed to suppress cell death by
directly inhibiting caspase activity (20). Therefore, the impact of GGA on the
steady-state levels of XIAP were examined. As shown in Fig. 2C and D, XIAP protein levels in the
kidney were significantly reduced following I/R injury in the
vehicle-treated rats, which was consistent with increased
apoptosis. The administration of GGA markedly enhanced HSP72
expression, corresponding with preservation of XIAP protein
contents. These findings suggest that HSP72 may protect the kidney
against I/R-induced injury, at least in part, through inhibiting
XIAP degradation.
HSP72 prevents Smac/DIABLO release and
caspase 3 activation
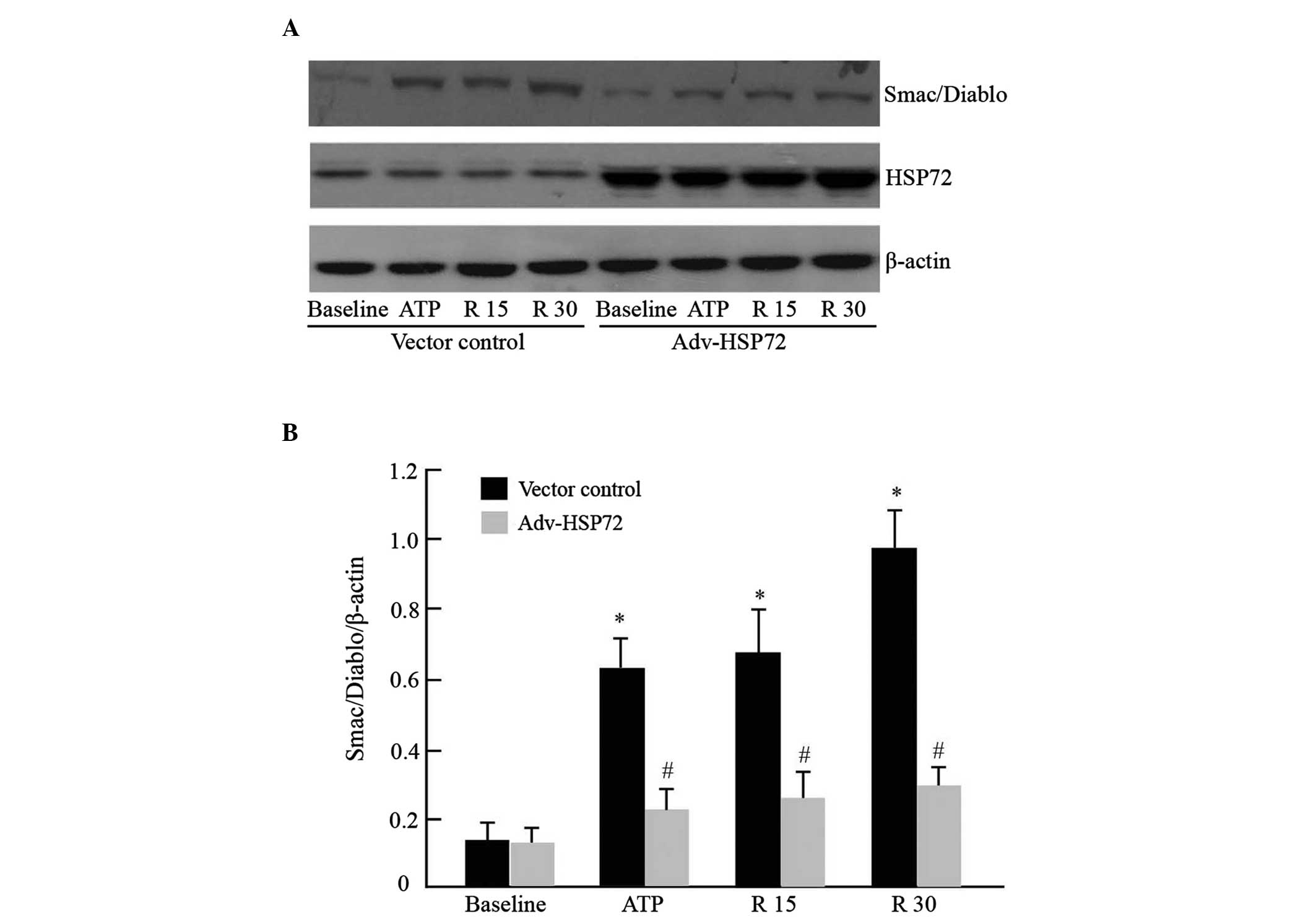
In response to apoptotic stimuli, Smac/DIABLO
protein is redistributed from the mitochondria to the cytosol,
binds to XIAP and activates the apoptosome complex. As GGA exposure
preserved XIAP protein levels, attenuated apoptosis and protected
against I/R renal injury in rats, the function of HSP72 in
regulating mitochondrial Smac/DIABLO translocation and caspase 3
activation were examined in cultured epithelial tubular cells
subjected to ATP depletion, as previously described (7). Under normal conditions, Smac/DIABLO
was not detected in the cytosol of empty vector cells. ATP
depletion caused a marked increase in cytosolic Smac/DIABLO. The
quantity of Smac/DIABLO progressively enhanced during recovery from
transient ATP depletion. However, HSP72 overexpression
significantly inhibited mitochondrial Smac/DIABLO release (Fig. 3A and B). Concordantly, levels of
pro-caspase 3 were reduced in metabolic inhibitor-treated control
cells, whereas caspase 3 activation was efficiently inhibited in
HSP72 overexpressing cells (Fig. 3C
and D). Without metabolic inhibitor exposure, HSP72
overexpression per se did not affect Smac/DIABLO leakage and
pro-caspase 3 content. This result suggested that HSP72 may confer
protective effects in part by inhibiting mitochondrial Smac/DIABLO
protein release.
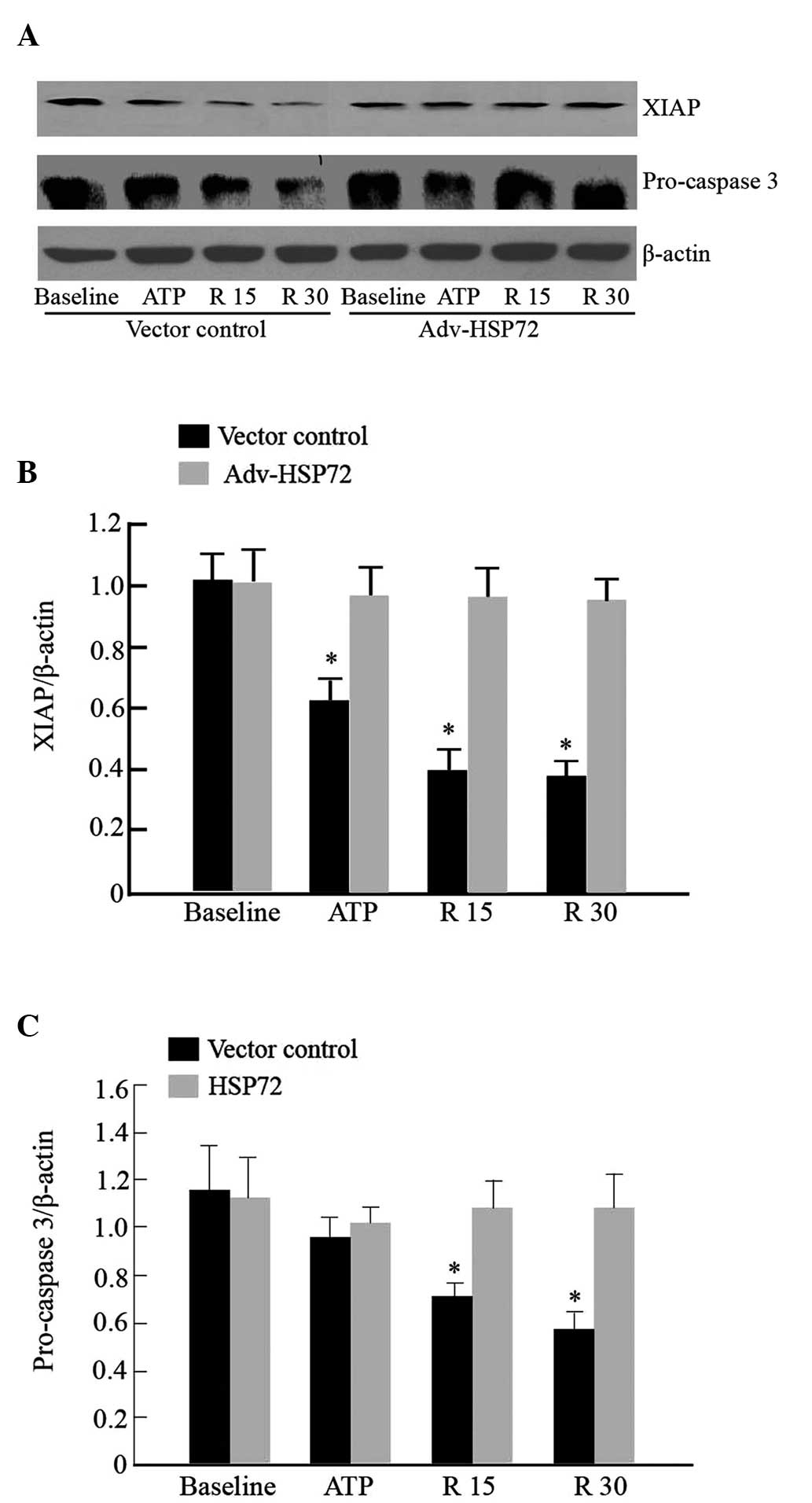
HSP72 stabilizes XIAP protein
XIAP is degraded during apoptosis induction
(21). Since HSP72 preserved XIAP
protein content in rats subjected to renal I/R, whether XIAP
degradation may be suppressed by HSP72 in HK-2 cells was further
examined. Consistent with our in vivo findings,
overexpression of HSP72 increased the stabilization of XIAP protein
following ATP depletion, compared with that in the empty vector
control cells (Fig. 4A and B). To
examine the possibility that HSP72 may prevent XIAP degradation by
binding to XIAP, immunoprecipitation of the two proteins was
performed. In the empty vector control cells, interaction between
HSP72 and XIAP was weak, whereas overexpression of HSP72
significantly elevated the interaction between these two proteins
under all experimental conditions. The immunoprecipitable XIAP
content was similar (Fig. 5). The
present results suggest that HSP72 may confer XIAP stability
against degradation through binding XIAP.
Discussion
The present study demonstrated that induction of
HSP72 by GGA preserved renal XIAP protein content, attenuated
tubular cell apoptosis and improved renal dysfunction following I/R
injury. In vitro experiments in HK-2 cells revealed that
overexpression of HSP72 inhibited mitochondrial Smac/DIABLO
release, interacted with XIAP and increased levels of XIAP and
pro-caspase 3 in ATP-depleted cells. These findings suggest that
elevated expression of HSP72 prevents the onset of tubular cell
apoptosis in renal I/R injury through suppression of Smac/DIABLO
release from mitochondria and restoring XIAP and pro-caspase 3
protein level.
Transient ischemia due to hypovolemia, hypotension
or heart failure commonly causes AKI, a disease associated with
high mortality, which is increasing in prevalence. Experimental and
human studies indicate that the mitochondrial-mediated apoptotic
pathway contributes to tubular cell detachment, loss and
dysfunction in the course of acute and chronic renal injury
(8,22–25).
The present study, as well as previous studies (22,24),
demonstrated that tubular apoptosis was markedly increased in the
I/R-subjected kidney compared with the sham-surgery-subjected
kidney. Notable evidence indicates that an alteration in
mitochondrial membrane integrity is crucial to the regulation of
apoptosis, as stress-mediated permeabilization permits
pro-apoptotic factors from the mitochondria into the cytosol and
activates caspase-dependent and independent pathways (1,2). The
present study demonstrated that mitochondrial Smac/DIABLO, like
cytochrome c and AIF (7,15) rapidly release into the cytosol of
renal tubular cells during exposure to metabolic inhibitors. During
recovery from ATP depletion and following release of mitochondrial
Smac/DIABLO, the protein levels of pro-caspase 3 and the caspase
inhibitory function of XIAP were reduced. These findings are in
agreement with those of previous studies and suggest that
Smac/DIABLO from mitochondria ensures continued caspase activation,
which is essential ultimately for cell death by inhibiting the
caspase inhibitory function of IAPs (3,5,26)
HSP72 is an abundant, inducible molecular chaperone.
Multiple studies have demonstrated that HSP72 may protect a variety
of cells, including renal tubule cells, against thermal, toxic and
ATP depletion-induced damage in vitro (7,15,27).
Overexpression of HSP72 also suppresses I/R-mediated myocardial,
liver, brain and renal injury in animals (28–30).
Through release of mitochondrial Smac/DIABLO, it is hypothesized
that HSP72 regulates Smac/DIABLO translocation and promotes cell
survival during ischemic AKI. In our previous study, administered
GGA specifically enhanced the expression of HSP72 in the kidney
(16). The present data revealed
that pre-conditioning rats with GGA attenuated tubular epithelial
cell injury, apoptosis and renal dysfunction. The in vitro
data substantiated the cytoprotective function of HSP72. As
observed following ischemia in vivo, overexpressed HSP72 in
cultured HK-2 cells attenuated mitochondrial Smac/DIABLO release
into the cytosol, degradation of XIAP and the activation of caspase
3. Since HSP72 has been observed to bind to various signaling
molecules, it may impact on cell survival. In the present study,
the interaction between HSP72 and XIAP was increased under
pathophysiological conditions and even elevated following
overexpression of HSP72 in the cells. Smac/DIABLO was found to
promote caspase activation by binding and neutralizing the IAPs,
including XIAP (3). The present
observations indicate that HSP72 functions as a regulator that
prevents XIAP degradation upon apoptotic signaling, competitively
inhibits the binding of XIAP and Smac/DIABLO and maintains the
association of XIAP with caspase, thereby preventing apoptosis.
In conclusion, the present study has revealed that
HSP72 prevents tubular apoptosis and dysfunction in renal I/R
injury by suppressing the release of mitochondrial Smac/DIABLO and
protecting the functions of pro-apoptotic proteins.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant nos. 81070553, 81270783 and
81130012) and the National Basic Research Program of China (grant
no. 2012CB517700-2012CB517706).
References
|
1
|
Suen DF, Norris KL and Youle RJ:
Mitochondrial dynamics and apoptosis. Genes Dev. 22:1577–1590.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brooks C, Wei Q, Cho SG and Dong Z:
Regulation of mitochondrial dynamics in acute kidney injury in cell
culture and rodent models. J Clin Invest. 119:1275–1285. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Du C, Fang M, Li Y, Li L and Wang X: Smac,
a mitochondrial protein that promotes cytochrome c-dependent
caspase activation by eliminating IAP inhibition. Cell. 102:33–42.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Deveraux QL, Takahashi R, Salvesen GS and
Reed JC: X-linked IAP is a direct inhibitor of cell-death
proteases. Nature. 388:300–304. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Verhagen AM, Ekert PG, Pakusch M, et al:
Identification of DIABLO, a mammalian protein that promotes
apoptosis by binding to and antagonizing IAP proteins. Cell.
102:43–53. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fulda S and Vucic D: Targeting IAP
proteins for therapeutic intervention in cancer. Nat Rev Drug
Discov. 11:109–124. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li F, Mao HP, Ruchalski KL, et al: Heat
stress prevents mitochondrial injury in ATP-depleted renal
epithelial cells. Am J Physiol Cell Physiol. 283:C917–C926. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bonventre JV and Weinberg JM: Recent
advances in the pathophysiology of ischemic acute renal failure. J
Am Soc Nephrol. 14:2199–2210. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Russell JC, Whiting H, Szuflita N and
Hossain MA: Nuclear translocation of X-linked inhibitor of
apoptosis (XIAP) determines cell fate after hypoxia ischemia in
neonatal brain. J Neurochem. 106:1357–1370. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Scarabelli TM, Stephanou A, Pasini E, et
al: Minocycline inhibits caspase activation and reactivation,
increases the ratio of XIAP to smac/DIABLO and reduces the
mitochondrial leakage of cytochrome C and smac/DIABLO. J Am Coll
Cardiol. 43:865–874. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saito A, Hayashi T, Okuno S, Nishi T and
Chan PH: Oxidative stress is associated with XIAP and Smac/DIABLO
signaling pathways in mouse brains after transient focal cerebral
ischemia. Stroke. 35:1443–1448. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
de Almagro MC and Vucic D: The inhibitor
of apoptosis (IAP) proteins are critical regulators of signaling
pathways and targets for anti-cancer therapy. Exp Oncol.
34:200–211. 2012.PubMed/NCBI
|
|
13
|
Hu W, Wang F, Tang J, et al: Proapoptotic
protein Smac mediates apoptosis in cisplatin-resistant ovarian
cancer cells when treated with the anti-tumor agent AT101. J Biol
Chem. 287:68–80. 2012. View Article : Google Scholar :
|
|
14
|
Mosser DD, Caron AW, Bourget L,
Denis-Larose C and Massie B: Role of the human heat shock protein
hsp70 in protection against stress-induced apoptosis. Mol Cell
Biol. 17:5317–5327. 1997.PubMed/NCBI
|
|
15
|
Ruchalski K, Mao H, Li Z, et al: Distinct
hsp70 domains mediate apoptosis-inducing factor release and nuclear
accumulation. J Biol Chem. 281:7873–7880. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mao H, Li Z, Zhou Y, et al: HSP72
attenuates renal tubular cell apoptosis and interstitial fibrosis
in obstructive nephropathy. Am J Physiol Renal Physiol.
295:F202–F214. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou Y, Mao H, Li S, et al: HSP72 inhibits
Smad3 activation and nuclear translocation in renal
epithelial-to-mesenchymal transition. J Am Soc Nephrol. 21:598–609.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Z, Gall JM, Bonegio RG, et al:
Induction of heat shock protein 70 inhibits ischemic renal injury.
Kidney Int. 79:861–870. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Suzuki S, Maruyama S, Sato W, Morita Y,
Sato F, Miki Y, Kato S, Katsuno M, Sobue G, Yuzawa Y and Matsuo S:
Geranylgeranylacetone ameliorates ischemic acute renal failure via
induction of Hsp70. Kidney Int. 67:2210–2220. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Eckelman BP, Salvesen GS and Scott FL:
Human inhibitor of apoptosis proteins: why XIAP is the black sheep
of the family. EMBO Rep. 7:988–994. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang Y, Fang S, Jensen JP, Weissman AM and
Ashwell JD: Ubiquitin protein ligase activity of IAPs and their
degradation in proteasomes in response to apoptotic stimuli.
Science. 288:874–877. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Havasi A and Borkan SC: Apoptosis and
acute kidney injury. Kidney Int. 80:29–40. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Oberbauer R, Rohrmoser M, Regele H,
Muhlbacher F and Mayer G: Apoptosis of tubular epithelial cells in
donor kidney biopsies predicts early renal allograft function. J Am
Soc Nephrol. 10:2006–2013. 1999.PubMed/NCBI
|
|
24
|
Kaushal GP, Basnakian AG and Shah SV:
Apoptotic pathways in ischemic acute renal failure. Kidney Int.
66:500–506. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sanz AB, Santamaria B, Ruiz-Ortega M,
Egido J and Ortiz A: Mechanisms of renal apoptosis in health and
disease. J Am Soc Nephrol. 19:1634–1642. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song Z, Yao X and Wu M: Direct interaction
between survivin and Smac/DIABLO is essential for the
anti-apoptotic activity of survivin during taxol-induced apoptosis.
J Biol Chem. 278:23130–23140. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao S, Lin Q, Li H, et al: Carbon
monoxide releasing molecule-2 attenuated ischemia/reperfusion
induced apoptosis in cardiomyocytes via a mitochondrial pathway.
Mol Med Rep. 9:754–762. 2014.
|
|
28
|
Pabla N, Dong G, Jiang M, et al:
Inhibition of PKCδ reduces cisplatin-induced nephrotoxicity without
blocking chemotherapeutic efficacy in mouse models of cancer. J
Clin Invest. 121:2709–2722. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tan Z, Turner RC, Leon RL, et al:
Bryostatin improves survival and reduces ischemic brain injury in
aged rats after acute ischemic stroke. Stroke. 44:3490–3497. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fang H, Liu A, Dahmen U and Dirsch O: Dual
role of chloroquine in liver ischemia reperfusion injury: reduction
of liver damage in early phase, but aggravation in late phase. Cell
Death Dis. 4:e6942013. View Article : Google Scholar : PubMed/NCBI
|