Introduction
Liver cirrhosis is a long-term consequence of
hepatitis virus infection or hepatic injury caused by alcohol
abuse, which is characterized by dysfunction with extensive
accumulation of fibrous tissue in the liver (1). Long-term and advanced cirrhosis is
usually irreversible and is often associated with variceal
hemorrhage or the development of hepatocellular carcinoma;
therefore, liver cirrhosis is a major worldwide cause of morbidity
and mortality (2). In the liver
tissue, quiescent hepatic stellate cells (HSCs) and liver
myofibroblasts are the main sources of hepatic extracellular matrix
(ECM) and play a key role in the pathogenesis (3,4).
Although the cell proliferation and migration of HSCs and portal
myofibroblasts are a pathogenic cause of hepatic fibrosis, previous
studies (5,6) have indicated that the apoptotic cell
death of liver cells may also be involved in the formation of
hepatic fibrosis and cirrhosis.
Previous studies have commonly reported that
endoplasmic reticulum (ER) stress plays a crucial role in the
induction and regulation of apoptosis (7,8).
There are numerous pathways involved in ER stress, such as
ER-associated protein degradation and the unfolded protein response
(UPR) (9) The UPR pathway also
activates several proteins, including inositol-requiring enzyme
(IRE) 1, protein kinase R-like ER kinase (Perk) and activating
transcription factor 6 (ATF6) (10). The above proteins can further
activate the apoptosis of the cells.
In the present study, the specific mechanisms for
the development of cirrhosis and hepatic fibrosis were investigated
in the liver tissue of a rat model of cirrhosis. It was proposed
that the elucidation of these mechanisms could lead the way to
therapeutic methods for patients with liver cirrhosis.
Materials and methods
Animal treatment and cirrhosis model
establishment
Wistar rats (Animal Center of Guiyang Medical
College, Guiyang, China) aged between four and six weeks (n=60)
were used in this study. All of the animal experiments were
performed according to the US National Institutes of Health 1991
guidelines, and approved by the Ethics Committee of Guiyang Medical
College (Guiyang, China). For the establishment of experimental
hepatic cirrhosis, rats (n=20) were intraperitoneally administered
with dimethylnitrosamine (Sigma-Aldrich, St. Louis, MO, USA)
dissolved in saline at 10 mg/kg rat body weight, on three
consecutive days a week for five continuous weeks. An equivalent
number of rats (n=20) were treated with saline only as a sham
group, and 20 untreated rats formed a control group.
Immunohistochemical examination
After the five week treatment period, the rats were
anaesthetized with intravenous pentobarbital sodium (40 mg/kg) and
sacrificed by cervical dislocation, and the liver tissues were
isolated and fixed with 10% neutralized formaldehyde or 70% ethanol
and embedded in paraffin. For the immunohistochemical examination
of desmin and α-smooth muscle actin (α-SMA), the tissue sections
were incubated with monoclonal anti-mouse desmin (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) and monoclonal anti-mouse
α-SMA (Santa Cruz Biotechnology, Inc.) antibodies for 1 h,
respectively. The immunohistochemical procedures were performed
according to the method described by Xu et al (11).
Hematoxylin and eosin (HE) staining
10 weeks after the start of the experiment the
animals were anaesthetized with 40 mg/kg intravenous pentobarbital
sodium and sacrificed and the livers were recovered for
histological analysis. The analysis was performed by an experienced
pathologist who was unaware of the experimental groups. Liver
specimens were fixed in buffered 10% formalin, embedded in paraffin
and stained with HE and picrosirius red to evaluate the extent of
liver fibrosis. A microscope (2124002; Sigma-Aldrich) was used to
evaluate the slides. The classification of the tissue slides was in
accordance with the methods of Goldani et al (12).
RNA extraction and semi-quantitative
reverse transcription-polymerase chain reaction (RT-PCR)
To measure the transcriptional levels of the
ER-stress associated agents, semi-quantitative RT-PCR assays were
designed, which included IRE1, cleaved ATF6 and p-Perk. β-actin
served as the internal control. Total RNA (10 μg) was obtained
using the RNAsimple Total RNA Kit [Tiagen Biotech (Beijing) Co.,
Ltd., China]. Reverse transcription was performed using the
SuperScript™ III First-Strand Synthesis System (Invitrogen Life
Technologies, Carlsbad, CA USA). Briefly, 2 μg total RNA, 200 U
MMLV reverse transcriptase and 50 pM oligo (dT20) were mixed in a
volume of 20 μl. The mixture was incubated at 50°C for 50 min and
inactivated by heating at 85°C for 5 min. To remove the RNA, 1 μl
RNase H was added to the mixture, followed by incubation at 37°C
for 20 min. Aliquots (2 μl) of the RT reaction products were
amplified by PCR in a volume of 50 μl under the following
conditions: 94°C for 30 sec, 54°C for 30 sec and 72°C for 40 sec.
Next, the products were electrophoresed on 1.4% agarose gels.
Images of each PCR product were then captured using a CCD camera
(Nikon Coporation, Tokyo, Japan) and analyzed using NIH Imager beta
version 2 (Bio-Rad Laborotories, Inc., Hercules, CA, USA). Relative
transcriptional values are presented as a ratio of the signal value
of the specific PCR product compared with individual β-actin.
Western blot analysis
In order to examine the protein expression using
western blot analysis, the cells were lysed in the sample buffer
for SDS-PAGE. The cell lysate was subjected to SDS-PAGE and
proteins were electroblotted onto polyvinylidene difluoride
membranes. Subsequent to blocking with phosphate-buffered saline
containing 5% skimmed milk, the membrane was incubated with abbit
anti-rat IRE1 monoclonal antibody (Santa Cruz Biotechnology, Inc.),
rabbit anti-rat cleaved ATF6 polyclonal antibody (Santa Cruz
Biotechnology, Inc.), rabbit anti-rat cleaved caspase-12 monoclonal
antibody (Santa Cruz Biotechnology, Inc.) and rabbit anti-rat CHOP
monoclonal antibody (Santa Cruz Biotechnology, Inc.).
Immunoreactive proteins were visualized using an enhanced
chemiluminescence reagent (Invitrogen Life Technologies).
Flow cytometry assay
Cell viability was assessed by flow cytometry, which
analyzed Annexin V-fluorescein isothiocyanate (FITC) binding and
propidium iodide (PI; Trevigen, Inc., Gaithersburg, MD, USA) uptake
simultaneously. Briefly, cells were harvested and resuspended in
Annexin V-FITC binding buffer (Invitrogen Life Technologies) and
incubated with Annexin V-FITC (Invitrogen Life Technologies, CA,
USA) and PI (5lg ml-1) (Invitrogen Life Technologies) in the dark
at room temperature for 15 min. Samples were then analyzed using a
FACScan flow cytometer (Becton-Dickinson, Oxford, UK). Fluorescence
was measured using a 530/30 band filter (FL-1) to monitor Annexin
V-FITC binding and a 585/42 band filter (FL-2) to monitor PI
uptake.
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical analyses were performed using the unpaired Student’s
t-test or analysis of variance. Differences were considered to be
statistically significant at P<0.05.
Results
Increased fibrosis in the cirrhosis rat
model
The analysis was performed in the 40 cirrhosis rats
(20 rats in the treated group and sham group, respectively) and 20
normal rats that survived through five weeks of the treatment
period by the time of sacrifice. Cirrhosis was significantly more
prevalent in the cirrhosis group than in the sham and control
groups (P<0.01); however, no significant difference was observed
between the sham and control groups (P>0.05). The histology of
the normal liver and liver tissues exhibiting fibrosis and
cirrhosis is shown in Fig. 1A. The
liver tissues were isolated and subjected to western blot analysis
in a consistent manner. The results indicated that hepatic
expression of α-SMA and desmin in the cirrhosis group was increased
compared with that in the sham and control groups (Fig. 1B). These findings strongly
suggested that the cirrhosis model was established
successfully.
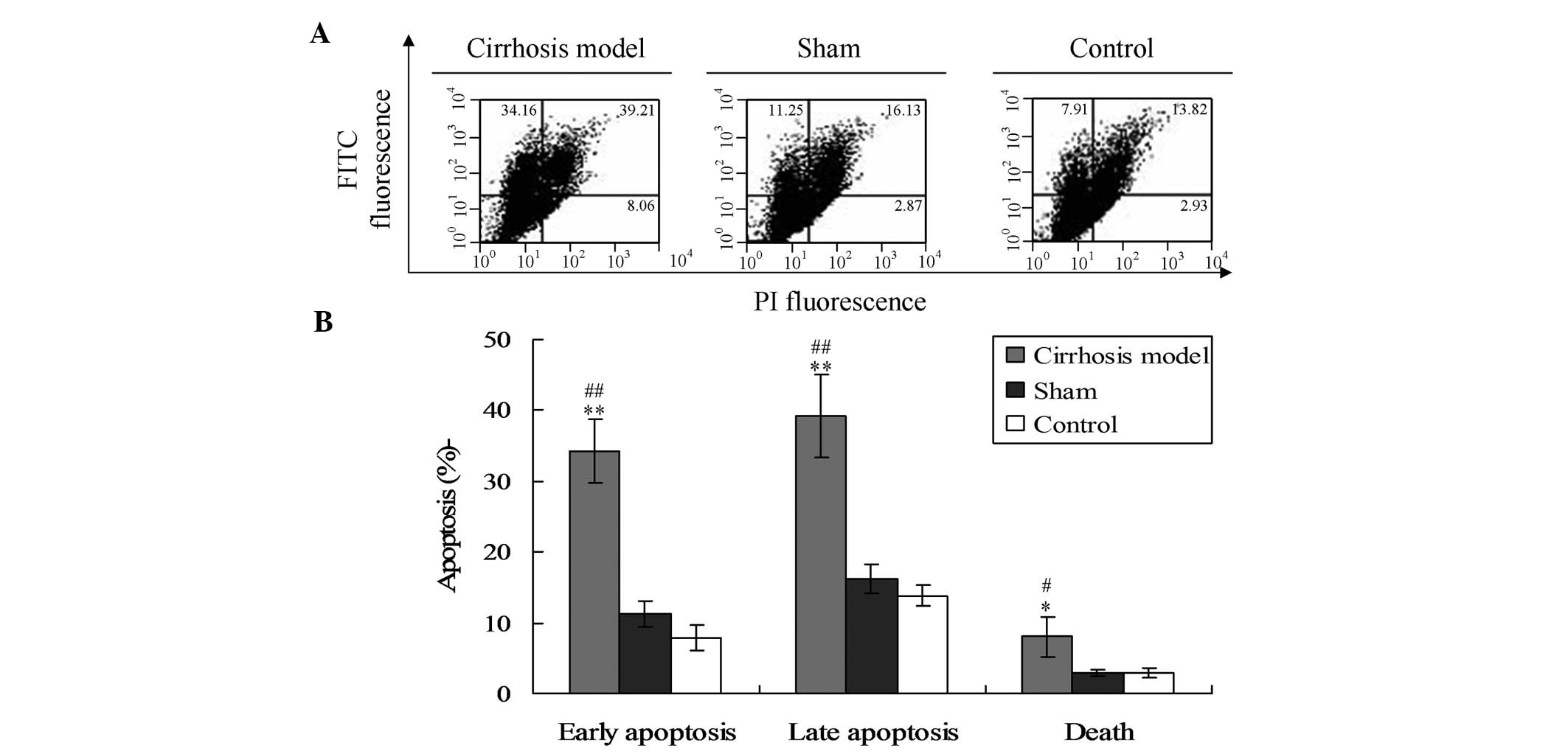
Induction of hepatic cell apoptosis in
cirrhotic rats
In the present study, it was hypothesized that
cirrhosis may be caused by the apoptosis of hepatic cells. Thus,
the apoptosis of the hepatic tissues was observed in each group.
Flow cytometric analysis showed that the levels of early apoptosis,
late apoptosis and cell death in the cirrhosis model were
significantly higher compared with those in the sham and control
groups (Fig. 2) (P<0.01 for
early and late apoptosis, P<0.05 for cell death).
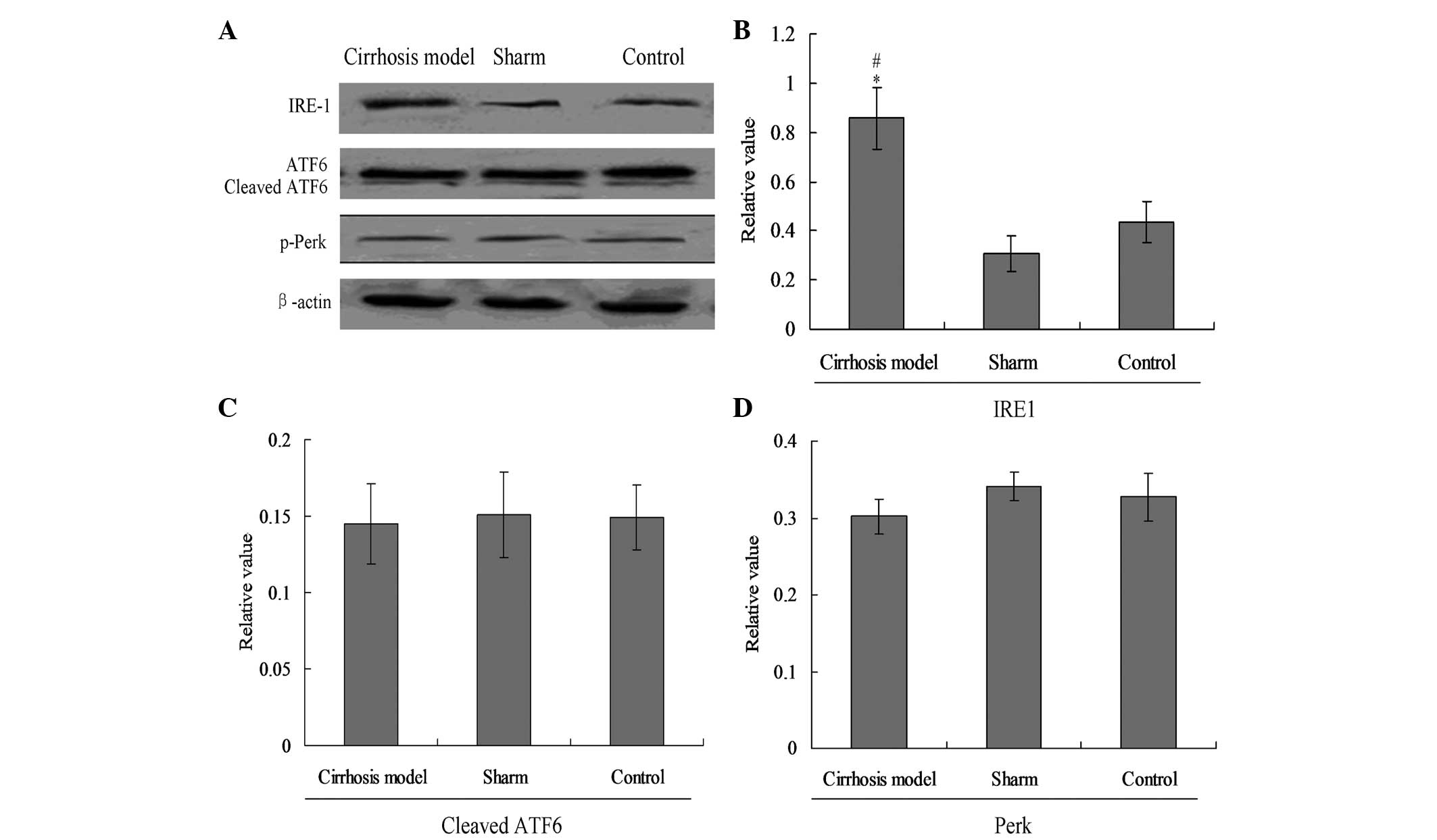
ER stress UPR pathways are involved in
cirrhosis-mediated apoptosis
In the present study, three main UPR factors of ER
stress, IRE1, Perk and ATF6, were analyzed by western blot assay
(Fig. 3) and the RT-PCR (Fig. 4). The results showed that the
expression of IRE1 in the cirrhosis group was increased
significantly compared with that in the sham and control groups
(Fig. 3A and B, P<0.05);
however, no evident changes were observed in cleaved ATF6 and
phosphorylated (p)-Perk protein among the three groups (Fig. 3A, C and D).
The expression of mRNA for the above UPR proteins
was analyzed with semi-quantitative PCR. As indicated in Fig. 4, when compared with the sham and
control groups, the mRNA levels of IRE1 were increased
significantly in the cirrhosis group (Fig. 4A, P<0.01). Notably, no
significant differences were observed in cleaved ATF6 and Perk
levels among the three groups (Fig. 4B
and C).
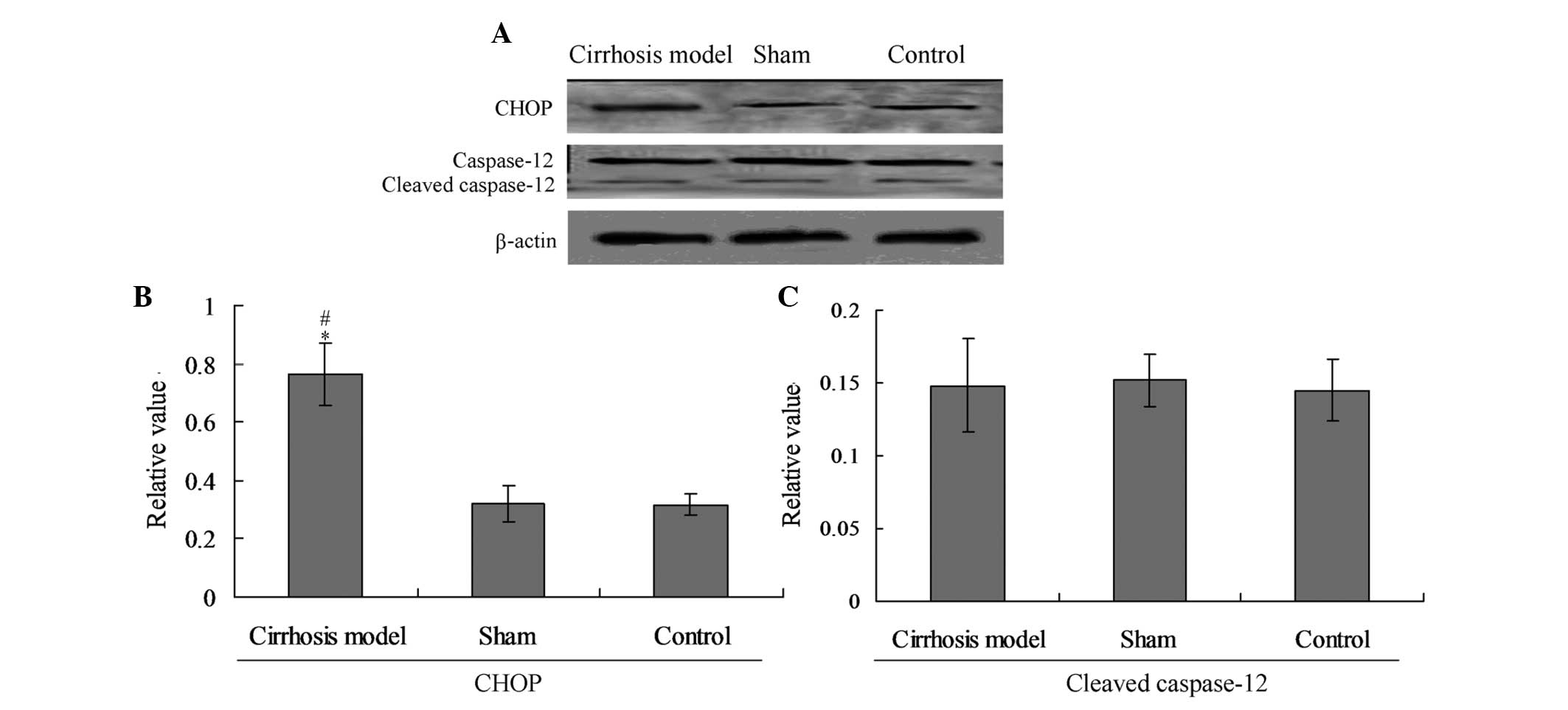
C/EBP homologous protein (CHOP) changes
mediate the ER stress-associated apoptosis in cirrhotic rats
To explore the specific pathway causing the
apoptosis in hepatic tissues, the cellular protein levels of
cleaved caspase-12 and CHOP were evaluated by individual western
blot analysis (Fig. 5A). The
results indicated that the expression of CHOP in the cirrhosis
model was significantly increased compared with that in the control
group (P<0.05). No significant differences were observed in
cleaved caspase-12 levels among the three groups (Fig. 5B).
Discussion
Activated HSCs and portal myofibroblasts act as
major cellular sources of ECM deposition in liver
fibrosis/cirrhosis following hepatic injury and/or inflammation.
Conversely, during spontaneous recovery from liver fibrosis,
activated HSCs and portal myofibroblasts undergo apoptotic cell
death (13). Thus, the apoptosis
of liver cells and portal myofibroblasts is closely involved in
recovery from liver fibrosis/cirrhosis. Although a few studies have
explored the association between cirrhosis and apoptosis (14,15),
the present study is the first, to the best of our knowledge, that
investigates the specific mechanism of cirrhosis-mediated apoptosis
in liver tissues.
In the present study, fibrosis formation was
observed in the cirrhosis rat model. The results indicated that the
fibrosis was significantly increased in the cirrhosis model group
compared with that in the normal control group. It has been shown
that, for fibrosis or cirrhosis formation, cells must express
fibrosis-related proteins (16);
thus, α-SMA and desmin proteins were detected in the hepatic
tissues in the present study. The marked increase in fibrous tissue
expansion and hepatic expression of α-SMA and desmin suggested that
the formation of the fibrosis or cirrhosis was caused by the α-SMA
and desmin expression. We speculated that the potential mechanism
for the enhanced level of α-SMA and desmin may have been due to the
apoptosis of hepatic cells.
In order to investigate the specific mechanism
underlying the cirrhosis-related apoptosis, the ER stress (UPR
pathway)-associated protein levels, including p-Perk, IRE1 and
cleaved ATF6, were detected (17).
It was found that IRE1 protein and mRNA were activated in the
hepatic tissue of the cirrhosis model; therefore, the IRE1 UPR
pathway may be involved in cirrhosis-related apoptosis. The results
of cirrhosis-related apoptosis may assist in examining the role of
fibrosis in the progression of liver cirrhosis. A previous study
showed that the activation of IRE1 could activate the ER-associated
apoptosis and cell death (18).
The data strongly indicated the emergence of ER stress for the
cirrhosis rat model; therefore, we propose that ER stress may
participate in the pathogenic process of fibrosis formation in
hepatic cirrhosis.
In a study by Zhao et al (19), ER stress-associated factors
(cleaved caspase-12 and CHOP) were detected to help identify the
specific apoptotic factors involved in the pathogenesis of
cirrhosis. The study reavealed that simvastatin protects MG63 cells
from oxidative stress-induced apoptosis through downregulation of
caspase-3, caspase-9 and caspase-12 activation. A number of studies
have found that cleaved caspase-12 can activate caspase-3 and
trigger apoptosis and that CHOP can directly induce ER
stress-associated apoptosis (20,21).
In the present study, no significant changes were observed in the
cleavage of caspase-12 protein (activated) among the groups
(P>0.05); however, the level of CHOP in the cirrhotic rats was
significantly increased compared with that in the sham and control
groups. From these results, it was deduced that the ER
stress-associated protein CHOP was activated, which subsequently
induced the apoptosis. This indicated that the fibrosis was caused
by CHOP activation induced by ER stress.
In conclusion, the protein expression of the UPR
pathway protein, inositol-requiring enzyme (IRE) 1, and the
expression of CHOP was increased significantly in the cirrhotic rat
tissues. The evident apoptosis observed in the hepatic tissue of
cirrhotic rats, which was caused by the activation of the ER
stress-mediated IRE1 and CHOP.
References
|
1
|
Zhang C, Wang Y, Chen H, Yang G, Wang S,
Jiang M, Cong L, Yuan L, Li H and Jia Y: Protective effect of the
herbal medicine Gan-fu-kang against carbon tetrachloride-induced
liver fibrosis in rats. Mol Med Rep. 8:954–962. 2013.PubMed/NCBI
|
|
2
|
Kim WH, Matsumoto K, Bessho K and Nakamura
T: Growth inhibition and apoptosis in liver myofibroblasts promoted
by hepatocyte growth factor leads to resolution from liver
cirrhosis. Am J Pathol. 166:1017–1028. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alcolado R, Arther MJP and Iredale JP:
Pathogenesis of liver fibrosis. Clin Sci (Lond). 92:103–112.
1997.
|
|
4
|
Yoshiji H, Noguchi R, Ikenaka Y, Kaji K,
Aihara Y, Douhara A, Yamao J, Toyohara M, Mitoro A, Sawai M,
Yoshida M, Morioka C, Fujimoto M, Uemura M and Fukui H: Combination
of branched-chain amino acid and angiotensin-converting enzyme
inhibitor improves liver fibrosis progression in patients with
cirrhosis. Mol Med Rep. 5:539–544. 2012.
|
|
5
|
Ramadori G and Saile B: Mesenchymal cells
in the liver - one cell type or two? Liver. 22:283–294. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cha JH, Bae SH, Kim HL, Park NR, Choi ES,
Jung ES, Choi JY and Yoon SK: Branched-chain amino acids ameliorate
fibrosis and suppress tumor growth in a rat model of hepatocellular
carcinoma with liver cirrhosis. PLoS One. 8:e778992013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yung HW, Korolchuk S, Tolkovsdy AM,
Charnock-Jones DS and Burton GJ: Endoplasmic reticulum stress
exacerbates ischemia-reperfusion-induced apoptosis through
attenuation of Akt protein synthesis in human choriocarcinoma
cells. FASEB J. 21:872–884. 2007. View Article : Google Scholar :
|
|
8
|
Moenner M, Pluquet O, Bouchecareilh M and
Chevet E: Integrated endoplasmic reticulum stress responses in
cancer. Cancer Res. 67:10631–10634. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Banjerdpongchai R, Punyati P, Nakrob A,
Pompimon W and Kongtawelert P: 4′-Hydroxycinnamaldehyde from
Alpinia galangal (Linn.) induces human leukemic cell apoptosis via
mitochondrial and endoplasmic reticulum stress pathways. Asian Pac
J Cancer Prev. 12:593–598. 2011.
|
|
10
|
Hung JY, Hsu YL, Ni WC, Tsai YM, Yang CJ,
Kuo PL and Huang MS: Oxidative and endoplasmic reticulum stress
signaling are involved in dehydrocostuslactone-mediated apoptosis
in human non-small cell lung cancer cells. Lung Cancer. 68:355–365.
2010. View Article : Google Scholar
|
|
11
|
Xu K, Liu XN, Zhang HB, An N, Wang Y,
Zhang ZC and Wang YN: Replication-defective HSV-1 effectively
targets trigeminal ganglion and inhibits viral pathopoiesis by
mediating interferon gamma expression in SH-SY5Y cells. J Mol
Neurosci. 53:78–86. 2014. View Article : Google Scholar
|
|
12
|
Goldani HA, Matte US, Ramos AR, Costa TG,
Winkelmann LV, Meureer L, Vieira SM, Kieling CO and Silveira TR:
The role of food restriction on CCl4-induced cirrhosis
model in rats. Exp Toxicol Pathol. 58:331–337. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Saile B, Matthes N, Neubauer K, Eisenbach
C, El-Armouche H, Dudas J and Ramadori G: Rat liver myofibroblasts
and hepatic stellate cells differ in CD95-mediated apoptosis and
response to TNF-alpha. Am J Physiol Gastrointest Liver Physiol.
283:G435–G444. 2002.PubMed/NCBI
|
|
14
|
Foster CR, Daniel LL, Daniels CR, Dalal S,
Singh M and Singh K: Deficiency of ataxia telangiectasia mutated
kinase modulates cardiac remodeling following myocardial
infarction: involvement in fibrosis and apoptosis. PLoS One.
8:e835132013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Puche JE, Saiman Y and Friedman SL:
Hepatic stellate cells and liver fibrosis. Compr Physiol.
3:1473–1492. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sheng L, Jiao B, Shao L, Bi S, Cheng C,
Zhang J and Jiang Y: Probucol inhibits hydrogen peroxide to induce
apoptosis of vascular smooth muscle cells. Mol Med Rep.
7:1185–1190. 2013.PubMed/NCBI
|
|
17
|
Xiao B, Han F, Wang HT and Shi YX:
Single-prolonged stress induces increased phosphorylation of
extracellular signal-regulated kinase in a rat model of
post-traumatic stress disorder. Mol Med Rep. 4:445–449.
2011.PubMed/NCBI
|
|
18
|
Lee JH, Kwon EJ and Kim DH: Calumenin has
a role in the alleviation of ER stress in neonatal rat
cardiomyocytes. Biochem Biophy Res Commun. 439:327–332. 2013.
View Article : Google Scholar
|
|
19
|
Zhao XH, Xu ZR, Zhang Q and Yang YM:
Simvastatin protects human osteosarcoma cells from oxidative
stress-induced apoptosis through mitochondrial-mediated signaling.
Mol Med Rep. 5:483–488. 2012.
|
|
20
|
Wang X, Shi Q, Xu K, Gao C, Chen C, Li XL,
Wang GR, Tian C, Han J and Dong XP: Familial CJD associated PrP
mutants within transmembrane region induced Ctm-PrP retention in ER
and triggered apoptosis by ER stress in SH-SY5Y cells. PLoS One.
6:e146022011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu K, Wang X, Shi Q, Chen C, Tian C, Li
XL, Zhou RM, Chu YL and Dong XP: Human prion protein mutants with
deleted and inserted octarepeats undergo different pathways to
trigger cell apoptosis. J Mol Neurosci. 43:225–234. 2011.
View Article : Google Scholar
|