Introduction
Epilepsy is a chronic disease characterized by
recurrent episodes of abnormal high-frequency neuronal discharge
resulting in the transient disturbance of cerebral function. ~80%
of all epilepsies are well-managed with drugs and numerous patients
have prolonged remissions. The remaining 20% are medically
intractable, often resistant to treatment with various
antiepileptic drugs (AEDs) at maximal dosages as monotherapies or
in combination, and are thus referred to as intractable epilepsy
(IE) cases (1). The underlying
mechanisms of IE remain elusive and represent a highly challenging
topic of epilepsy research that has attracted notable attention in
recent years.
D site of albumin promoter binding protein (DBP),
hepatic leukemia factor (HLF) and thyrotroph embryonic factor (TEF)
are the three members of the proline and acidic amino acid-rich
basic leucine zipper (PAR bZip) transcription factor family. All
three of these transcriptional regulatory proteins accumulate with
robust circadian rhythms in tissues with high amplitudes of clock
gene expression, including the suprachiasmatic nucleus (SCN) and
the liver. In mammals, the circadian timing system controls
numerous aspects of behavior and physiology, including
rest-activity cycles, heartbeat frequency, body temperature,
arterial blood pressure, endocrine functions, renal plasma flow,
intestinal peristalsis and metabolism (2). PAR bZip proteins control the
expression of numerous enzymes and regulators involved in
detoxification and drug metabolism, including cytochrome P450
enzymes, carboxylesterases and constitutive androstane receptor
(CAR) (3). Only DBP is capable of
efficiently activating transcription from the cholesterol
7α-hydroxylase (C7αH) promoter (4). It has been suggested that DBP may
have an important role in drug resistance.
However, DBP expresses at a nearly invariable level
in the majority of brain regions, in which clock gene expression
only cycles with low amplitude (4,5).
Klugmann et al (6) reported
that DBP expression gradually increased between postnatal day 1 and
day 60. Several studies have demonstrated that mice with a
deficiency of only one or two PAR bZip genes exhibit relatively
mild phenotypes (5,7,8),
whereas strong phenotypes are observed when all three PAR bZip
genes are inactivated. Approximately half of these animals tend to
have spontaneous and sound-induced epileptic seizures during the
first three months following birth (5). Of note, locomotor activity appears to
correlate with DBP levels (9). By
contrast, DBP-deficient mice demonstrated reduced baseline
locomotor activity and blunted behavioral responses to acute
methamphetamine stimulation in an open field test.
The mitogen-activated protein kinase (MAPK) pathway
is an extracellular signal pathway capable of causing a cell
nuclear reaction and even activating components of the programmed
cell death pathway, including extracellular signal-regulated
protein kinase 1/2 (ERK1/2), ERK3/4, ERK5, p38 and c-Jun N-terminal
kinase (JNK).
The MAPK pathway is widely expressed in the central
nervous system (CNS). Each type of extracellular stimulus signal,
including neurotransmitters, nerve trophic factors and growth
factors, may use this pathway to affect synaptic transmission,
neuronal remodeling, morphological differentiation and survival.
Therefore, this signaling pathway is involved in the pathological
processes associated with numerous types of nervous system
disorders, including epilepsy. Based on the important physiological
roles of DBP in the adult brain, it is important to examine whether
DBP is abnormally expressed in IE and whether it may serve as a
marker for the condition. MAPK and the signal pathways mentioned
above have both been reported in epilepsy and correlated to the
underlying mechanism of the disease. It may therefore be possible
that DBP, through the interaction with MAPK, participates in the
drug-resistant mechanisms of epilepsy. In order to examine the
possible roles of DBP in the pathogenesis of IE, the present study
detected DBP expression by immunohistochemically staining tissue
from patients with IE and examined the correlation between DBP and
MAPK by double-label immunofluorescence staining. It was
hypothesized that DBP and MAPK have a key role in the mechanisms
that underlie the pathogenesis of IE.
Materials and methods
IE group
In the present study, all of the patients with
epilepsy had typical clinical manifestations and characteristic
electroencephalograms. A total of 35 patients with IE were randomly
selected from the epilepsy brain tissue collections of The First
Affiliated Hospital of Chongqing Medical University (Chongqing,
China). The diagnosis of seizure type was confirmed according to
the 1981 International League Against Epilepsy. Prior to the
surgery, the epileptic lesion was localized in all patients by
brain magnetic resonance imaging (MRI) or computerized tomography
(CT), and 24 h electroencephalography (EEG) or video EEG;
sphenoidal electrode monitoring and intraoperative
electrocorticography (ECOG) were performed to localize the
epileptic lesion prior to resection in all patients. Two
neuropathologists reviewed each case independently. The 35 patients
were refractory to maximal doses of at least three AEDs, including
phenytoin, valproic acid, carbamazepine, phenobarbital and
topiramate. Brain MRI or CT, and the associated laboratory
inspection, did not discover the presence of other nervous system
diseases in any of the patients. Table
I summarizes the patients’ clinical features. In these epilepsy
patients, surgical removal of the epileptogenic zone in the
temporal neocortex was an alternative treatment option. The
pathological findings in the resected tissue included gliosis and
neuronal loss, as well as signs of neuronal degeneration. Following
lesion resection, the electrodes for intraoperative
electrocorticography were placed on the remaining edge of tissue to
ensure that the lesion was resected completely.
 | Table IClinical characteristics of temporal
lobe epilepsy patients. |
Table I
Clinical characteristics of temporal
lobe epilepsy patients.
| Patient no. | Gender | Age (years) | Duration (years) | Seizure type | AEDs | Resection tissue | Pathology |
|---|
| 1 | M | 28 | 2 | SGS | CBZ, PHT, VPA | TNr | nd |
| 2 | F | 36 | 16 | SGS | CBZ, PHT, VPA,
PB | TNl | nd, nl |
| 3 | F | 22 | 3 | SGS | CBZ, VPA, PHT | TNr | nd, nl, g |
| 4 | M | 15 | 9 | SGS | CBZ, PHT, VPA | TNl | g |
| 5 | M | 14 | 7 | CPS | CBZ, PHT, PB | TNl | nd, g |
| 6 | M | 20 | 10 | SGS | CBZ, PHT, VPA′ | TNr | nd, nl, g |
| 7 | F | 17 | 5 | CPS | CBZ, PB, CLB | TNr | nd, g |
| 8 | F | 58 | 40 | CPS | CBZ, PHT, CLB | TNl | nd, g |
| 9 | M | 8 | 5 | CPS | CBZ, PB, VPA | TNl | nd, g |
| 10 | M | 18 | 13 | SGS | CBZ, PRM, PHT | TNr | nd, g |
| 11 | M | 15 | 4 | SGS | CBZ, PHT, TPM | TNr | nd, g |
| 12 | M | 32 | 20 | SGS | CBZ, VPA, TPM | TNl | nl, g |
| 13 | F | 47 | 18 | CPS | CBZ, PB, CLB,
PRM | TNr | nd, nl, g |
| 14 | M | 41 | 20 | SGS | CBZ, PHT, VPA,
PB | TNr | nd, nl, g |
| 15 | M | 17 | 5 | SGS | TPM, VPA, PHT | TNr | nd, nl, g |
| 16 | M | 22 | 16 | SGS | CBZ, TPM, VPA,
PB | TNl | nl, g |
| 17 | M | 16 | 14 | SGS | CBZ, PHT, PB | TNl | nl, nd, g |
| 18 | M | 15 | 15 | SGS | CBZ, CLB, TPM | TNl | nd, g |
| 19 | M | 25 | 1 | SGS | CBZ, PRM, TPM,
PB | TNr | nd |
| 20 | M | 26 | 9 | SGS | CBZ, PHT, VPA,
CLB | TNr | nl, nd, g |
| 21 | F | 45 | 3 | CPS | CBZ, VPA, TPM | TNr | g |
| 22 | M | 26 | 14 | SGS | CBZ, TPM, VPA,
CLB | TNr | nl, nd, g |
| 23 | M | 22 | 21 | SGS | CBZ, VPA, TPM,
PB | TNl | nd, g |
| 24 | M | 22 | 12 | SGS | CBZ, VPA, PRM, NIM,
PB | TNr | nl, nd, g |
| 25 | M | 12 | 3 | SGS | CBZ, VPA, PB, CLB,
PHT | TNr | nl, nd, g |
| 26 | F | 15 | 2 | CPS | CBZ, TPM, VPA | TNl | nd, g |
| 27 | M | 22 | 1 | CPS | CBZ, PB, TPM | TNr | nl |
| 28 | M | 15 | 2 | SGS | CBZ, VPA, PHT | TNr | nd, g |
| 29 | M | 24 | 22 | SGS | CBZ, VPA, PB, TPM,
CLB | TNl | nl, nd, g |
| 30 | F | 22 | 5 | SGS | CBZ, VPA, CLB | TNr | nl |
| 31 | F | 18 | 16 | SGS | CBZ, PB, VPA | TNl | nd, g |
| 32 | F | 23 | 8 | SGS | CBZ, PB, TPM | TNl | nl |
| 33 | M | 16 | 8 | SGS | CBZ, VPA, TPM | TNl | g |
| 34 | F | 44 | 12 | CPS | CBZ, TMP, VPA | TNl | nd, g |
| 35 | F | 24 | 21 | SGS | CBZ, PB, PRM,
VPA | TNr | nl, g |
Control group
For comparison, 15 histologically normal temporal
neocortex samples were obtained from individuals treated for
increased intracranial pressure due to head trauma. The time
between mortality and surgery was <5 h. All of the patients were
diagnosed, by the same two pathologists, with brain trauma and had
no prior history of seizures or other neurological disorders. The
majority of the neurological structural features appeared normal.
Table II summarizes the clinical
features of the controls.
| Table IIClinical characteristics of control
patients. |
Table II
Clinical characteristics of control
patients.
| Gender | Age (years) | Etiological
diagnosis | Resection
tissue | Adjacent tissue
pathology |
|---|
| Male | 47 | Trauma | TNl | Normal |
| Female | 22 | Trauma | TNl | Normal |
| Male | 31 | Trauma | TNl | Normal |
| Male | 23 | Trauma | TNr | Normal |
| Female | 20 | Trauma | TNr | Normal |
| Male | 46 | Trauma | TNl | Normal |
| Male | 21 | Trauma | TNr | Normal |
| Male | 29 | Trauma | TNl | Normal |
| Female | 42 | Trauma | TNr | Normal |
| Male | 38 | Trauma | TNl | Normal |
| Male | 38 | Trauma | TNl | Normal |
| Female | 17 | Trauma | TNr | Normal |
| Male | 48 | Trauma | TNl | Normal |
| Female | 18 | Trauma | TNr | Normal |
| Female | 10 | Trauma | TNl | Normal |
The National Institutes of Health and the Ethics
Committee on Human Research at Chongqing Medical University
approved the present study (Chongqing, China). Informed consent was
obtained from the patients or their relatives for the use of any
data and tissues for the study.
Tissue processing
The resected brain tissues were fixed in 10%
buffered formalin. Following fixation in formalin for 24–48 h, the
specimens were routinely paraffin-embedded, sectioned and preserved
for future use at room temperature.
Immunohistochemistry
Each paraffin-embedded sample was cut into 5-μm
thick sections, which were spread on polylysine-coated slides. Each
specimen produced 10 slices, of which one was selected randomly for
immunohistochemical staining. Immunohistochemistry was performed
according to the manufacturer’s instructions of the DAB Detection
kit (Polymer, Zhongshan Golden Bridge, Inc., Beijing, China).
First, the paraffin sections were deparaffinized, hydrated through
graded alcohols and incubated in H2O2 (0.3%
for 10 min). The sections were heated in a microwave oven for 20
min at 98°C in a citrate buffer (pH 6.0) for antigen retrieval,
then cooled naturally to 37°C at room temperature and blocked in
normal goat serum (1:10) for 10 min (Zhongshan Golden Bridge Inc.,
Beijing, China). The sections were incubated in primary rabbit
anti-human DBP polyclonal antibody (1:200; Aviva Systems Biology
Corporation, San Diego, CA, USA) overnight at 4°C, followed by a
biotinylated anti-rabbit secondary antibody for 30 min at 37°C.
Immunoreactivity was observed with 3,30-diaminobenzidine
(DAB+; Zhongshan, Golden Bridge Inc.). Counterstaining
was conducted with Harris hematoxylin. The blank controls were
obtained by substituting the primary antibody with
phosphate-buffered saline (PBS).
Immunofluorescence
Each paraffin-embedded sample was cut into 10-μm
thick sections, which were spread onto polylysine-coated slides.
Each specimen produced 10 slices, of which one was selected
randomly for analysis. The paraffin sections were deparaffinized,
hydrated through graded alcohols and incubated in Triton (0.4% of
15 min). The sections were heated in a microwave oven for 20 min at
98°C in citrate buffer (pH 6.0) for antigen retrieval and then
cooled naturally to 37°C at room temperature. Subsequently, the
sections were blocked in calf serum mix followed by normal goat
serum for 5 h (Zhongshan Golden Bridge Inc., Beijing). The sections
were incubated in primary rabbit anti-human DBP polyclonal antibody
(1:100; Aviva Systems Biology Corporation) and mouse anti-human
MAPK polyclonal antibody (1:75; Zhongshan Golden Bridge Inc.)
overnight at 4°C, followed by tetramethylrhodamine-conjugated
AffiniPure goat anti-mouse immunoglobulin (Ig)G (1:50; Zhongshan
Golden Bridge Inc.) and fluorescein isothiocyanate-conjugated
AffiniPure goat anti-rabbit IgG (1:50; Zhongshan Golden Bridge
Inc.) for 3 h (under protection from light) at room temperature.
The sections were mounted with 50% glycerol and 50% PBS. The
fluorescent-stained sections were observed by confocal microscopy
(TCS SP2; Leica Microsystems GmbH, Wetzlar, Germany).
Statistical analysis
Data are expressed as the mean ± standard deviation.
Student’s t-test (SPSS version 17.0; SPSS, Inc., Chicago, IL, USA)
was used for statistical analysis of the differences between the IE
group and control group in humans.
Results
Demographic and clinical characteristics
of the patients
The mean age of the patents with drug-refractory IE
was 34.97±17.26 years and consisted of 12 male and 23 female
patients. A total of 48.5% of patients had at least a 10-year
history of seizure recurrence and 31.4% had a clinical history of
epilepsy of >15 years. The control group had a mean age of
30±12.39 years and consisted of nine male and six female subjects.
There were no significant differences in age or gender between the
IE and control groups (P>0.05).
Significant DBP staining in the temporal
neocortex of patients with IE
In the IE group, DBP was consistently observed in
all cases, whereas reduced DBP levels were apparent in the control
group (Fig. 1). The expression of
DBP was more apparent in the neurons of IE patients than in those
of the control group and it was almost absent in glial cells in
both groups. No immunoreactivity was observed in the negative
control sections, in which the primary antibody was omitted. DBP
expression was significantly higher in the brain tissues of
patients with IE as compared with that in the control group. The
mean absorbance of DBP expression in the IE group was 0.31±0.03,
whereas in the control group, it was 0.18±0.02 (P<0.05). The
expression of MAPK was upregulated in IE patients as compared with
that in the controls (Fig. 2). The
mean absorbance of MAPK expression in the IE group was 0.19±0.03,
whereas in the control group, it was 0.12±0.02 (P<0.05).
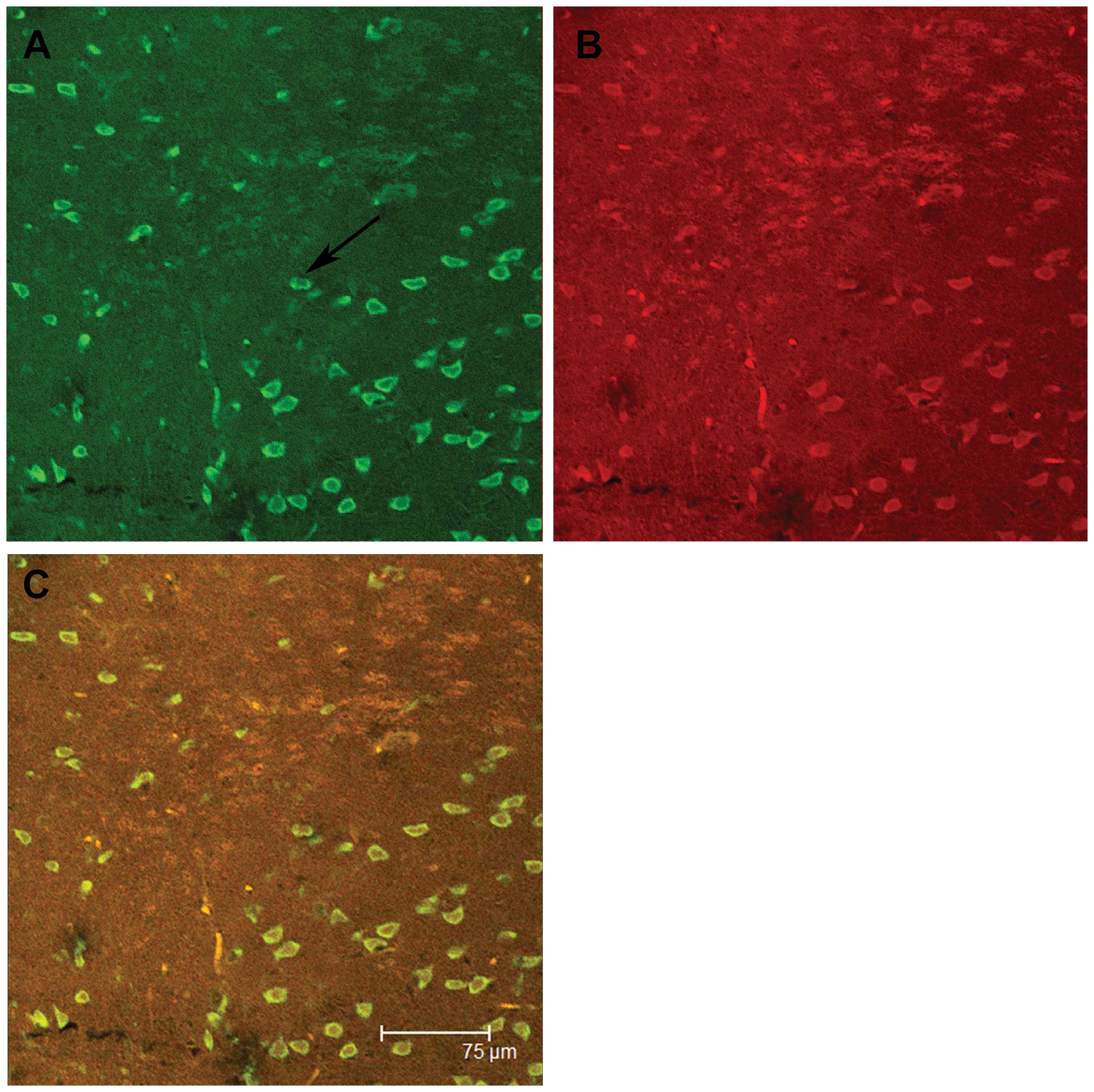
Immunofluorescence staining showed that in the IE group, the
expression of both DBP and MAPK was present in the same neurons,
whereas almost no immunoreactivity was apparent in the glia from
either group (Fig. 3).
Discussion
Epileptic seizures are characterized by
high-frequency perturbations of the activity of cells of the CNS,
which may subsequently initiate a series of complex nervous
biological effects. Each type of intranuclear adverse factor causes
a reaction through a series of signal transduction pathways. From
that moment, activated signaling molecules may develop continuous
cascade reactions, changing the expression of numerous genes,
exhibiting durability functions and finally, causing delayed nerve
cell death. The pathological mechanisms involved in IE are
intricate and, to date, not fully understood. Numerous studies have
reported that DBP and MAPK are associated with the pathogenesis of
epilepsy. However, it is not clear whether DBP, and its interaction
with MAPK, participate in mechanisms of drug resistance in
epilepsy. The aim of the present study was to clarify the role of
DBP and MAPK in the mechanisms that underlie the pathogenesis of
IE.
Immunohistochemical analysis of protein levels
demonstrated that DBP expression was significantly higher in the
brain tissues of patients with IE and that it was localized to
neurons specifically, while nearly absent in glia. It is therefore
concluded that DBP does not participate in gliosis but exerts its
biological effect through neurons, contributing to the mechanisms
that trigger epilepsy.
Several studies have suggested that PAR bZip
triple-knockout mice have high sensitivity to lethal sound-induced
and spontaneous seizures that are immediately followed by a
disturbance of neurotransmitter homeostasis (5). DBP-deficient mice do not exhibit such
major abnormalities and this is possibly due to functional
redundancy (10). The phenotype is
considered to be caused by a decrease in pyridoxal kinase (Pdxk),
which is encoded by a PAR bZIP target gene in the liver and the
brain. Pyridoxal kinase converts vitamin B6 derivatives into
pyridoxal phosphate (PLP), the coenzyme of numerous enzymes
involved in amino acid and neurotransmitter metabolism. PLP also
serves as the coenzyme of aromatic amino acid decarboxylase, an
enzyme involved in the synthesis of the monoamines serotonin and
dopamine (11).
These neurotransmitters have been associated with
the occurrence of epileptic events. Even moderate reductions in the
concentration of PLP have been demonstrated to provoke epileptic
seizures (12,13). During et al (13) suggested that DBP downregulation
paralleled anticonvulsive effects following glucagon-like peptide-1
receptor GLP-1R activation, which follows abnormally reduced
susceptibility to kainate-induced seizures. The present study
suggested that DBP levels in the brain may impact the modulation of
seizures. Matthias’ microarray analysis demonstrated that
hippocampal activation of GLP-1R, which is associated with improved
learning and neuroprotection, resulted in the suppression of DBP
(6). By contrast, the other two
PAR bZip transcription factor family members, TEF and HLF, did not
produce evident changes. The present study also demonstrated a
compensatory shut-down of DBP expression to counteract neuronal
overexcitation through glutamate receptors. However, it is unclear
whether persistent upregulation of DBP increases the sensitivity to
epilepsy. Studies have demonstrated that the persistent
upregulation of DBP may cause the upregulation of pdxk and induce
the activation of ERK1/2 and MAPK, as compared with the controls
(6). The ERK1/2 signaling pathway
belongs to the MAPK family and its major roles involve coordinating
neuronal responses to external signaling, affecting the remodeling
of synapses, axonal growth, long term potentiation and neuronal
excitability, by regulatory gene expression and protein synthesis
(14). The continual activation of
this pathway prompts persistent delivery of neurotransmitters. The
latter possibly maintains the activation of the ERK circuit through
phosphorylation, and ERK activation subsequently enhances the
delivery of synaptic transmitters through positive feedback.
MAPK expression has been primarily observed in
neurons following seizures and the present results confirmed that
while the upregulation of MAPK was expressed in glial cells and
neurons of both groups, it was mainly observed in neurons from
patients with IE. This suggests a significant role of MAPK in
neurons associated with chronic epilepsy. In the liver, DBP has an
important role in detoxification and drug metabolism. Epilepsy
causes the upregulation of DBP and MAPK, and while MAPK
participates in the progression and promotes the development of
epilepsy, DBP activates MAPK through positive feedback (15,16).
In conclusion, the present study demonstrated that
the expression of DBP and MAPK in patients with IE was upregulated.
The simultaneous co-expression of DBP and MAPK indicated that the
interaction of DBP and MAPK may participate in the remodeling of
synapses and the drug resistance mechanism of epilepsy. However, it
remains elusive how DBP and MAPK participate in this mechanism.
Further studies are required to determine the role of DBP and MAPK
in the mechanism underlying the pathogenesis of IE, and the present
study offers a new direction of inquiry into this concept.
Acknowledgements
The authors are grateful to the patients and their
families for their participation in the study, The First Affiliated
Hospital of Chongqing Medical University for their support in brain
tissue procurement, and the National Institutes of Health of China
and the Ethics Committee on Human Research of the Chongqing Medical
University. The authors are also grateful to the National Natural
Science Foundation of China (no. 81171225) for supporting the
study.
References
|
1
|
Shorvon SD: The epidemiology and treatment
of chronic and refractory epilepsy. Epilepsia. 37(Suppl 2): S1–S3.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gachon F, Nagoshi E, Brown SA, Ripperger J
and Schibler U: The mammalian circadian timing system: from gene
expression to physiology. Chromosoma. 113:103–112. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gachon F, Olela FF, Schaad O, Descombes P
and Schibler U: The circadian PAR-domain basic leucine zipper
transcription factors DBP, TEF, and HLF modulate basal and
inducible xenobiotic detoxification. Cell Metab. 4:25–36. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fonjallaz P, Ossipow V, Wanner G and
Schibler U: The two PAR leucine zipper proteins, TEF and DBP,
display similar circadian and tissue-specific expression, but have
different target promoter preferences. EMBO J. 15:351–362.
1996.PubMed/NCBI
|
|
5
|
Gachon F, Fonjallaz P, Damiola F, et al:
The loss of circadian PAR bZip transcription factors results in
epilepsy. Genes Dev. 18:1397–1412. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Klugmann M, Leichtlein CB, Symes CW, et
al: A novel role of circadian transcription factor DBP in
hippocampal plasticity. Mol Cell Neurosci. 31:303–314. 2006.
View Article : Google Scholar
|
|
7
|
Lopez-Molina L, Conquet F, Dubois-Dauphin
M and Schibler U: The DBP gene is expressed according to a
circadian rhythm in the suprachiasmatic nucleus and influences
circadian behavior. EMBO J. 16:6762–6771. 1997. View Article : Google Scholar
|
|
8
|
Franken P, Lopez-Molina L, Marcacci L,
Schibler U and Tafti M: The transcription factor DBP affects
circadian sleep consolidation and rhythmic EEG activity. J
Neurosci. 20:617–625. 2000.PubMed/NCBI
|
|
9
|
Niculescu AB III, Segal DS, Kuczenski R,
Barrett T, Hauger RL and Kelsoe JR: Identifying a series of
candidate genes for mania and psychosis: a convergent functional
genomics approach. Physiol Genomics. 4:83–91. 2000.PubMed/NCBI
|
|
10
|
Siow YL and Dakshinamurti K: Effect of
pyridoxine deficiency on aromatic L-amino acid decarboxylase in
adult rat brain. Exp Brain Res. 59:575–581. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sharma SK and Dakshinamurti K: Seizure
activity in pyridoxine-deficient adult rats. Epilepsia. 33:235–247.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schlesinger K and Lieff B: Levels of
pyridoxine and susceptibility to electroconvulsive and audiogenic
seizures. Psychopharmacologia. 42:27–32. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
During MJ, Cao L, Zuzga DS, et al:
Glucagon-like peptide-1 receptor is involved in learning and
neuroprotection. Nat Med. 9:1173–1179. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yin Y, Huang P, Han Z, et al: Collagen
nanofibers facilitated presynaptic maturation in differentiated
neurons from spinal-cord-derived neural stem cells through
MAPK/ERK1/2-Synapsin I signaling pathway. Biomacromolecules.
15:2449–2460. 2014.PubMed/NCBI
|
|
15
|
McClelland AC, Hruska M, Coenen AJ, et al:
Trans-synaptic EphB2-ephrin-B3 interaction regulates excitatory
synapse density by inhibition of postsynaptic MAPK signaling. Proc
Natl Acad Sci USA. 107:8830–8835. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mao LM, Reusch JM, Fibuch EE, Liu Z and
Wang JQ: Amphetamine increases phosphorylation of MAPK/ERK at
synaptic sites in the rat striatum and medial prefrontal cortex.
Brain Res. 1494:101–108. 2013. View Article : Google Scholar :
|