Introduction
Postoperative cognitive dysfunction (POCD) is
characterized by a decline of cognitive performance following
surgery, particularly following surgery of aged patients (1). It has been identified to occur in
30–80% of aged patients 1 week after surgery and in 10–15% of aged
patients 3 months after surgery (2–4). A
number of risk factors for POCD have been reported in previous
studies, including advanced age, educational level, mental illness,
preoperative level of recognition, operative type, and
postoperative infection (4,5).
However, the pathological mechanisms underlying POCD remain to be
elucidated. The toxicity of anesthetics was initially assessed,
with simple anesthesia impairing the learning and memory of aged
rats (1,6–8) and
mice (9). Notably, the duration
and type of anesthesia had no significant effects on the incidence
of POCD of aged patients at the late stage (>1 week post
surgery) (10). This suggested
that anesthetics were not important in POCD associated with aged
patients. In addition to surgery and anesthesia, another potential
mechanism underlying POCD is inflammation induced by surgery. He
et al (11) and Wang et
al (12) reported that
surgical trauma under general anesthesia causes distinct changes in
systemic and central pro-inflammatory cytokines in aged rats at an
early stage (<1 week post surgery), corresponding closely to the
dysfunction of cognition. Open tibial fractures impair the
cognitive function and induce the increase of tumor necrosis
factor-α (TNF-α) and interleukin-1 (IL-1) in adult mice (4,13).
Inhibiting the signals of TNF-α and IL-1 prior to surgery partly
rescued the cognitive dysfunction induced by surgery (4,13).
However, a similar incidence of POCD has been detected among aged
patients 3 months after surgery, although the extent of the damage
induced by surgery was independent of the anaesthetic and the type
of surgery (14). This suggested
that, in addition to the toxicity of anesthetics and inflammation
induced by surgery, other unknown factors also contribute to POCD
in aged patients.
The combination of mass spectrometry and
two-dimensional gel electrophoresis is a useful method to examine
the differential protein expression levels between different
treatments. It is usually used to examine novel mechanisms
underlying certain diseases (15).
The hippocampus is an important structure, closely involved with
learning and memory (16,17). During normal aging, the volume of
the hippocampus decreases gradually, particularly the dentate gyrus
and the CA3 area (18,19). During pathological aging, as
observed in Alzheimer’s disease, the hippocampal volume decreases
more rapidly (18,19). Accordingly, Chen et al
(20) found that the hippocampal
volume in aged patients with POCD is significantly smaller compared
with that of aged patients without POCD (20). In addition, the hippocampal volume
was negatively correlated with the score of cognitive function of
aged patients. These suggested that changes in the volume of the
hippocampus were closely involved in the occurrence of POCD in aged
patients. In the present study, differences in the protein
expression of the hippocampus were compared between aged rats with
POCD and normal aged rats.
Materials and methods
Animals and grouping
A total of 24 aged male Sprague-Dawley rats (22
months old) were purchased from the Experimental Animal Nursery of
Central South University (Changsha, China). The rats were
maintained under temperature-controlled environmental conditions
with a 12/12-h light/dark cycle and had ad libitum access to
food and water. The food consisted of standardized rodent pellets.
Ambient conditions remained constant at 23°C and a relative
humidity of 50–60%. All procedures were approved by the Medical
Ethics Committee of the Third Xiangya Hospital of Central South
University and conformed to the guidelines for animal experiments
of Central South University.
A total of 24 rats were handled, according to the
flow chart of the study design shown in Fig. 1. The rats were divided randomly
into a surgery group (n=12) and a normal control group (n=12). The
rats in the surgery group were treated with isoflurane anesthesia
for 2 h and splenectomy, while rats in the normal control group
received 40% oxygen for 2 h without surgery (1). The cognitive functions of the two
groups were examined using a Y-maze test. The upper limit of the
95% confidence interval of the mean training duration of rats in
the normal control group in the Y-maze test was used to decide
whether the rats in the surgery group had cognitive dysfunction
(21). The rats in the surgery
group were subdivided into POCD and no-POCD groups, according to
previously described methods (21). The protein profile from the
hippocampi of six aged rats (three rats with POCD and three rats
from the normal control group) were detected using two-dimensional
gel electrophoresis (2-DE) and matrix-assisted laser
desorption/ionization time of flight mass spectrometry
(MALDI-TOF-MS).
Y-maze test
The details of the Y-maze assessment (Fig. 2) was described in our previous
study (1). Briefly, the voltage
for the electric shock in the Y-maze was 50±5 V. The rats were
acclimatized to the Y-maze with the light on for 3 min 24 h after
treatment. Subsequently, the rats were placed in the stem arm and
were administered an electric shock. If the rats arrived at the lit
arms within 30 sec, the trial was deemed successful. The same trial
was repeated 20 times/day consecutively for 3 days. During the
assessment, the following parameters were recorded: The number of
correct reactions, which was the total number entering the lit arm
of the 20 trials each day; the total reaction duration, which rats
spent completing the 20 trials each day and the number of
initiative avoidance responses, which was the number of escape
responses of the rats within the 5 sec prior to the arms being
electrified.
Splenectomy surgery
Splenectomy surgery was performed using the method
reported in our previous study (11). Briefly, the rats were initially
placed in a closed box with an airflow of 3% isoflurane for 5 min.
Anesthesia was then administered through a mask using 2%
isoflurane. A 14-gauge catheter was inserted through the glottis.
The rats were artificially ventilated, through endotracheal
intubation, with room air supplemented with 2 l/min oxygen and 2%
isoflurane for 2 h (22). During
the anesthesia, the gas concentrations and respiratory rates (RR;
breaths per min)were continuously monitored using a multi-function
monitor (Datex-Ohmeda, Helsinki, Finland) (Table I). Rectal temperature was
maintained at 37±0.5°C. In addition, heart rate (HR), mean arterial
blood pressure (MAP) and pulse oximeter oxygen saturation (SpO2)
were measured continuously through a femoral artery catheter
(Table I). Under anesthesia, a
small incision was made in the upper left quadrant through the skin
and muscle wall. The spleen was mobilized, isolated and removed.
The wound was infiltrated with 0.25% bupivacaine and then closed
with sterile sutures. The rats in the normal control group received
40% oxygen for 2 h in a gas chamber without surgery.
 | Table IVital signs of the rats during and
after splenectomy surgery. |
Table I
Vital signs of the rats during and
after splenectomy surgery.
| Group | Index | 10 min
preoperative | 10 min
operative | 20 min
operative | 10 min
postoperative |
|---|
| Control | RR (bpm) | 82.20±3.35 | 72.06±4.68 | 71.33±3.29 | 78.50±2.67 |
| HR (bpm) | 374.83±11.70 | 359.83±16.49 | 362.35±20.11 | 365.00±16.07 |
| MAP (mmHg) | 95.33±10.17 | 83.83±10.32 | 85.29±13.25 | 88.67±7.23 |
| SpO2 (%) | 97.17±1.48 | 97.50±1.64 | 97.27±1.06 | 97.83±1.16 |
| Surgery | RR (bpm) | 80.00±4.70 | 71.50±6.78 | 73.75±7.69 | 75.60±8.41 |
| HR (bpm) | 372.57±12.11 | 348.28±11.33 | 355.88±13.68 | 361.78±12.52 |
| MAP (mmHg) | 92.21±8.32 | 83.21±9.07 | 85.64±8.33 | 87.14±8.37 |
| SpO2 (%) | 97.93±1.07 | 97.78±1.31 | 97.65±1.08 | 97.42±1.15 |
2-DE and MALDI-TOF-MS
Hippocampal tissue (250 mg) was suspended in 1 ml of
8 M urea, 2 M sulfocarbamide, 20 mM Tris-HCl, 1 mM CHAPS, 40 mM
EDTA, 65 mM dithiothreitol, 1 mM phenylmethylsulfonyl fluoride,
0.025 mM RNase, 0.05 mM DNase and 1 mM NaF. The suspension was
sonicated for ~30 sec and centrifuged at 150,000 × g for 45 min.
The protein concentration in the supernatant was examined using the
Coomassie blue method (23). In
first dimensional isoelectric focusing, 1,000 μg of each
sample was loaded into immobilized pH 3–10 nonlinear gradient
strips (18 cm). After 12 h of reswelling at 30 V, voltages of 100,
500, 1,000 and 3,000 V were administered for 1 h each and 8,000 V
for 6 h. The second dimensional separation was performed using 12%
sodium dodecyl sulfate-polyacrylamide gels (Sangon Biotech Shanghai
Co., Ltd, Shanghai, China) (24).
Following staining of the gels with colloidal Coomassie blue
(25), the gels were scanned using
a Umax PowerLook II scanner and Adobe Photoshop 8.0 image software
(Adobe Systems Inc., San Jose,. CA, USA), and the images were
analyzed using ETTAN ImageMaster 2D Elite 4.01 software (GE
Healthcare, Amersham, UK). This software was used for matching and
quantitative analysis of the protein spots on the gels. The average
gel was constructed as a representative gel for the three
hippocampal samples acquired from each group of rats. The average
mode of background subtraction was used for normalization of the
intensity volume that indicates the protein concentration or the
quantity on each spot. The average gel was then used for
determination of the existence of differences in the protein
expression levels between each group. A value of 2.0 indicated a
2-fold increase and 0.5 indicated a 2-fold reduction (26–29).
Biologically relevant differences were defined as changes
>2-fold.
MALDI-TOF-MS analysis was performed, as described
previously (25–29). The valuable spots in the coomassie
blue-stained gels were assessed. The extracts were redissolved in 1
μl extraction buffer and 1 μl matrix solution
(α-acyano-4-hydroxycinnamic acid; Sangon Biotech Shanghai Co., Ltd)
and targeted onto a MALDI-TOF plate. Following drying the samples
completely onto the targeting plate, MALDI-TOF/MS was performed
using a Voyager-DE STR mass spectrometer (Applied Biosystems,
Foster City, CA, USA) equipped with delay ion extraction. Mass
spectra were obtained over a mass range of 800–3,000 Da. For the
identification of proteins, peptide mass fingerprinting data were
used to search against the Mascot database (http://www.matrixscience.com) at the School of Life
(Central South University). Peptide matching and protein
identification were performed automatically, as described
previously (26–29). A Mascot score >58 was considered
to indicate statistical significance (P<0.05).
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR) for astrocyte-associated
proteins
To confirm the differential protein expression based
on MALDI-TOF-MS, the mRNA expression levels of the proteins of
interest were detected by RT-qPCR. Total RNA from the hippocampus
was isolated using the RNeasy plus mini kit (Qiagen, Valencia, CA,
USA), according to the manufacturer’s instructions. Prior to RT,
the total RNA was treated to remove genomic DNA using a DNA-free
kit (AM1906; Applied Biosystems). cDNA was synthesized using the
Retroscript kit (AM1710; Applied Biosystems). RT-qPCR was performed
using Power SYBR Green PCR master mix (Applied Biosystems). The rat
β-actin gene was used as a homogenous standard. The running
protocol extended to 40 cycles consisting of 95°C for 15 sec and
60°C for 1 min using an Applied Biosystems 7500 Fast Real-time PCR
system. The primers were designed using Primer 5.0 software
(Premier Biosoft International, Palo Alto, CA, USA; Table II).
| Table IIPrimers used for reverse
transcription quantitative polymerase chain reaction. |
Table II
Primers used for reverse
transcription quantitative polymerase chain reaction.
| Gene | Primer | Amplicon
length(bp) |
|---|
| Glutamate
dehydrogenase | Forward:
GCTCTGGACTCTTCCCAACA | 115 |
| Reverse:
AAATGCCACACGCCTACTTC |
| Glutamine
synthetase | Forward:
CGCTCTTCGTCTCGTTCTC | 119 |
| Reverse:
CTGCTTGATGCCTTTGTTCA |
| Fatty acid binding
protein 7, brain | Forward:
GAAGGGCAAGGATGGTAGATG | 133 |
| Reverse:
ACCGTTGGTTTGGTCACATT |
| β-actin | Forward:
GGAAATCGTGCGTGACATTA | 181 |
| Reverse:
GAAGGAAGGCTGGAAGAGAG |
The relative quantity of the target gene present was
calculated based on the expression of β-actin in the endogenous
control. The mean cycle threshold (Ct) values and standard
deviations were calculated. To calibrate the analysis, the value
obtained from the control rats was used. The factor difference was
also calculated using the standard relative quantitative method
(2−ΔΔCT method) (30,31).
Statistical analysis
All summary data are reported as the mean ± standard
error of the mean. The results of the present study were processed
using SPSS 17.0 statistical software (SPSS, Inc., Chicago, IL,
USA). The total reaction time in the Y maze was analyzed using a
repeated measures analysis of variance (ANOVA) with Bonferroni’s
post-hoc test. The number of correct reactions and initiative
avoidance occurrences were examined using a non-parametric
Mann-Whitney U test. The RR, HR and MAP values during surgery were
compared using a repeated measures ANOVA with Bonferroni’s post-hoc
test. The gel electrophoresis and RT-qPCR results were analyzed
using Student’s t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Surgery with inhaled anesthesia impairs
the cognitive function of aged rats
In the present study, a Y-maze test was used to
evaluate cognitive function (1). A
decrease of correct reactions and initiative avoidance number, and
an increase in total reaction time implied deteriorating cognitive
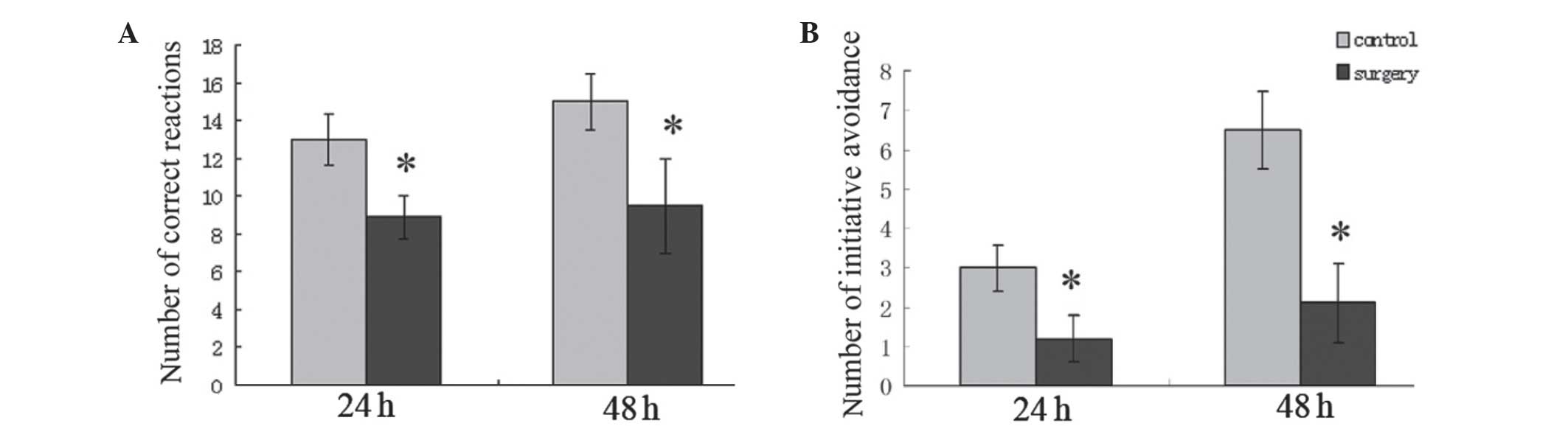
function. The findings demonstrated that the number of correct
reactions in the surgery group were 8.90±1.09 and 9.47±2.46 (24 and
48 h, respectively) following surgery, which was significantly
lower compared with the control group (P<0.05; Fig. 3A). The number of initiative
avoidance occurrences in the surgery group were 1.20±0.59 and
2.07±1.01 (24 h and 48 h, respectively) following surgery, which
was also significantly lower compared with the control group
(P<0.05; Fig. 3B). These
findings demonstrated that surgery with inhaled anesthesia impaired
the cognitive functions of aged rats at an early stage.
Surgery with inhaled anesthesia alters
the protein profile of the hippocampus in aged rats with POCD
The protein expression profile of the hippocampus
was assessed using MALDI-TOF-MS. The levels of 21 proteins in the
hippocampus of aged rats with POCD were markedly altered, compared
with the normal control. These proteins included dynamin-1,
superoxide dismutase, glutamine synthetase and coronin-1A (Table III, Fig. 4). Further functional clustering
revealed that three proteins were closely involved in synaptic
plasticity, four in oxidative stress, six in energy production
three in neuroinflammation and two in glutamate metabolism. The
protein expression levels were downregulated, with regard to
synaptic plasticity, oxidative stress, energy production and
glutamate metabolism; however, the protein expression levels
associated with neuroinflammation were upregulated in the POCD
rats.
| Table IIIDifferential protein expression
between the control group and postoperative cognitive dysfunction
group. |
Table III
Differential protein expression
between the control group and postoperative cognitive dysfunction
group.
| Protein symbol | Protein name | Number of spots in
gel | Changes in
expression (POCD, vs. control) | Protein
function |
|---|
| DNM1 | Dynamin-1 | 2 | ↓ | Synaptic
plasticity |
| DHPRP2 |
Dihydropyrimidinase-related protein 2 | 3 | ↓ | Synaptic
plasticity |
| GABA-T | GABA
transaminase | 19 | ↓ | Modulating neuronal
plasticity |
| SOD1 | Superoxide
dismutase (Cu-Zn) | 8 | ↓ | Antioxidant
protective role |
| GSTP1 | Glutathione
S-transferase P | 12 | ↓ | Antioxidant
protective role |
| PRDX2 |
Peroxiredoxin-2 | 5 | ↑ | Antioxidant
protective role |
| ADR | Aldose
reductase | 14 | ↑ | Oxidative
stress |
| DLST |
Dihydrolipoyllysine-residue
succinyltransferase | 4 | ↓ | Energy production
in mitochondria |
| ATP5D | ATP synthase δ
chain | 7 | ↓ | Energy production
in mitochondria |
| NDUFA10 | NADH dehydrogenase1
α subcomplex subunit 10 | 13 | ↓ | Energy production
in mitochondria |
| PGAM1 | Phosphoglycerate
mutase 1 | 11 | ↓ | Energy production
in mitochondria |
| TKT | Transketolase | 20 | ↓ | Energy production
and neurogenesis |
| PK-M1/M2 | Pyruvate kinase
isozymes M1/M2 | 21 | ↓ | Energy production
in mitochondria |
| CORO1A | Coronin-1A | 1 | ↑ | Neuroinflammation
marker |
| GMFB | Glia maturation
factor b | 6 | ↓ | Modulating
neuroinflammation |
| CRYAB | α crystallin B
chain | 11 | ↑ | Modulating
neuroinflammation |
| FABP7 | Fatty acid-binding
protein, | 9 | ↑ | Regulation of
astrocyte function |
| GS | Glutamine
synthetase | 17 | ↓ | Modulating
extracellular glutamate level |
| GLUD1 | Glutamate
dehydrogenase | 18 | ↑ | Modulating
extracellular glutamate level and energy production of
mitochondria |
| CCT2 | T-complex protein 1
subunit β | 23 | ↑ | Protein
folding |
| EF-Tu | Elongation factor
Tu | 16 | ↓ | Modulating
translation of DNA |
Astrocytes are the predominant type of cell in the
hippocampus, with important roles in supporting neurons. Thus, from
the identified differentially expressed proteins, three proteins
associated with astrocytes were selected [fatty acid binding
protein 7, brain (FABP7) (32),
glutamate dehydrogenase 1 (GLUD1) (33) and glutamine synthetase (GS)
(34)] to confirm the reliability
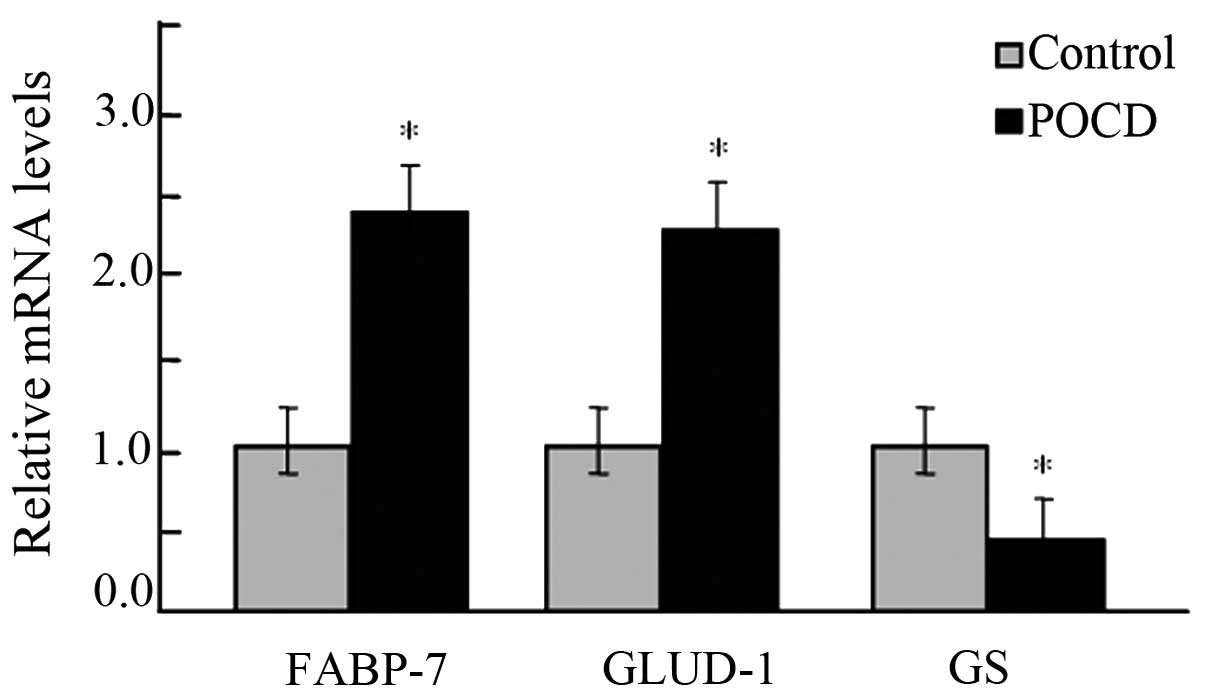
of the data from the MALDI-TOF-MS assessment. It was identified
that the mRNAs of FABP-7 and GLUD increased in aged rats with POCD
and the mRNAs of GS decreased, closely corresponding to the changes
observed in the MALDI-TOF-MS data (P<0.05; Fig. 5).
Discussion
In the present study, the protein profile of the
hippocampi in aged rats with POCD and normal aged rats were
screened. A total of 21 proteins were significantly altered in aged
rats with POCD compared with normal aged rats. These proteins were
functionally clustered in association with synaptic plasticity,
oxidative stress, energy production, glutamate metabolism and
neuroinflammation. These data provided important indications that
the hippocampus is functionally important in the POCD of aged
rats.
POCD is usually detected among aged patients
following surgery. It was initially reported by Bedford in 1955
(35). In the present study, the
cognitive function of aged rats was initially assessed with or
without surgery using a Y-maze. The number of correct reactions and
the number of initiative avoidance occurrences in the surgery group
were significantly lower compared with the control group
(P<0.05). These results suggested that surgery with inhaled
anesthesia impaired the cognitive function of aged rats at an early
stage. Based on these findings, changes in the protein expression
profile of the hippocampi of aged rats with POCD was assessed.
Compared with the normal control, 14 proteins increased in the aged
rats with POCD and seven proteins decreased (Table II). PubMed (http://www.ncbi.nlm.nih.gov/pubmed/) was used to
search for useful information using key words. Among the 21
proteins, three proteins, DNM1 (36–38),
DHPRP2 (39) and GABA-T (40,41)
were involved in synaptic plasticity and their expression were
decreased in aged rats with POCD. A total of four proteins, SOD1
(42,43), glutathione S-transferase P
(44,45), peroxiredoxin 2 (46) and ADR (47) were associated with oxidative
stress. In addition, six proteins, dihydrolipoyllysine-residue
succinyltransferase (48), ATP
synthase δ chain (49), NADH
dehydrogenase 1α (50),
phosphoglycerate mutase 1 (51),
transketolase (52) and pyruvate
kinase isozymes M1/M2 (53), were
involved in energy production and their expression levels were
decreased in the aged rats with POCD. A total of three proteins,
coronin-1A (54), glia maturation
factor b (55), α crystallin B
chain (56) were involved in
neuroinflammation. These findings suggested that surgery induced
neuronal damage in aged rats with POCD. Neuroinflammation and
oxidative stress were important in neuronal damage and the data
were consistent with previously reported results (4,11–13,20,57).
Chen et al (20) found that
aged patients with POCD had a smaller volume hippocampus and the
size of the hippocampus was negatively correlated with the
cognitive function of aged patients following surgery. It has been
found that surgery induced increased neuroinflammation in mice and
aged rats (4,11,12),
corresponding to cognitive dysfunction. In addition, An et
al (57) found that surgical
trauma induced oxidative stress in rats with POCD (57). Notably, the expression of FABP7
(32), a modulator of astrocyte
function, was higher in aged rats with POCD. At the same time, GS
(58) and GLUD1 (33,59–61),
which are two astrocytic enzymes modulating the level of
extracellular glutamate, were also changed in aged rats with POCD.
These findings suggested that astrocytes are important in the
mechanism underlying POCD, and astrocytes may offer a novel
direction in the investigation of POCD.
The present study did not distinguish the effect of
surgery in POCD from the anaesthetic agent, as our previous study
demonstrated that anesthesia and surgery can result in POCD. It has
been confirmed that surgery, rather than anesthetic agents lead to
long-term changes in exploratory behavior (62). It is disputed whether the mechanism
underlying POCD is associated with surgery and/or anesthesia,
however, it is considered to be a multi-factorial process. Further
investigation is required to discriminate each component of the
entire process.
Acknowledgments
The authors would like to thank all those who
assisted with this study, particularly Professor Jianbin Tong, who
provided technical assistance and critically edited the
manuscript.
References
|
1
|
Yan XB, Ouyang W, Li G and Duan KM:
Involvement of neuronal nitric oxide synthase in cognitive
impairment in isoflurane-treated rats. Neurosci Lett. 506:240–244.
2012. View Article : Google Scholar
|
|
2
|
Ramaiah R and Lam AM: Postoperative
cognitive dysfunction in the elderly. Anesthesiol Clin. 27:485–496.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Monk TG and Price CC: Postoperative
cognitive disorders. Curr Opin Crit Care. 17:376–381. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Terrando N, Monaco C, Ma D, Foxwell BM,
Feldmann M and Maze M: Tumor necrosis factor-alpha triggers a
cytokine cascade yielding postoperative cognitive decline. Proc
Natl Acad Sci USA. 107:20518–20522. 2012. View Article : Google Scholar
|
|
5
|
Rosczyk HA, Sparkman NL and Johnson RW:
Neuroinflammation and cognitive function in aged mice following
minor surgery. Exp Gerontol. 43:840–846. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Culley DJ, Baxter MG, Yukhananov R and
Crosby G: Long-term impairment of acquisition of a spatial memory
task following isoflurane-nitrous oxide anesthesia in rats.
Anesthesiology. 100:309–314. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Culley DJ, Baxter M, Yukhananov R and
Crosby G, Culley DJ, Baxter M, Yukhananov R and Crosby G: The
memory effects of general anesthesia persist for weeks in young and
aged rats. Anesth Analg. 96:1004–1009. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Culley DJ, Baxter MG, Crosby CA,
Yukhananov R and Crosby G: Impaired acquisition of spatial memory 2
weeks after isoflurane and isoflurane-nitrous oxide anesthesia in
aged rats. Anesth Analg. 99:1393–1397. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hudson AE and Hemmings HC Jr: Are
anaesthetics toxic to the brain? Br J Anaesth. 107:30–37. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rasmussen LS, Johnson T, Kuipers HM,
Kristensen D, Siersma VD, Vila P, Jolles J, Papaioannou A,
Abildstrom H, Silverstein JH, Bonal JA, Raeder J, Nielsen IK,
Korttila K, Munoz L, Dodds C, Hanning CD and Moller JT: ISPOCD2
(international study of postoperative cognitive dysfunction)
investigators: does anaesthesia cause postoperative cognitive
dysfunction? A randomised study of regional versus general
anaesthesia in 438 elderly patients. Acta Anaesthesiol Scand.
47:260–266. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He HJ, Wang Y, Le Y, Duan KM, Yan XB, Liao
Q, Liao Y, Tong JB, Terrando N and Ouyang W: Surgery upregulates
high mobility group box-1 and disrupts the blood-brain barrier
causing cognitive dysfunction in aged rats. CNS Neurosci Ther.
18:994–1002. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang Y, He HJ, Li D, Zhu WJ, Duan KM, Le
Y, Liao Y and Ouyang W: The role of the TLR4 signaling pathway in
cognitive deficits following surgery in aged rats. Mol Med Rep.
7:1137–1142. 2013.PubMed/NCBI
|
|
13
|
Terrando N, Eriksson LI, Ryu JK, Yang T,
Monaco C, Feldmann M, Jonsson Fagerlund M, Charo IF, Akassoglou K
and Maze M: Resolving postoperative neuroinflammation and cognitive
decline. Ann Neurol. 70:986–995. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Evered L, Scott DA, Silbert B and Maruff
P: Postoperative cognitive dysfunction is independent of type of
surgery and anesthetic. Anesth Analg. 112:1179–1185. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bi FF, Huang C, Tong JB, Qiu G, Huang B,
Wu Qx, Li F, Xu ZS, Xia XG and Zhou HX: Reactive astrocytes secrete
Lcn2 to assist neurons in degeneration. Proc Natl Acad Sci USA.
110:4069–4074. 2013. View Article : Google Scholar
|
|
16
|
Dede AJ, Wixted JT, Hopkins RO and Squire
LR: Hippocampal damage impairs recognition memory broadly,
affecting both parameters in two prominent models of memory. Proc
Natl Acad Sci USA. 1:6577–6582. 2013. View Article : Google Scholar
|
|
17
|
Da Silva WC, Cardoso G, Bonini JS, Benetti
F and Izquierdo I: Memory reconsolidation and its maintenance
depend on L-voltage-dependent calcium channels and CaMKII functions
regulating protein turnover in the hippocampus. Proc Natl Acad Sci
USA. 110:6566–6570. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Buckner RL: Memory and executive function
in aging and AD: multiple factors that cause decline and reserve
factors that compensate. Neuron. 44:195–208. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jagust W: Vulnerable neural systems and
the borderland of brain aging and neurodegeneration. Neuron.
77:219–234. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen MH, Liao Y, Rong PF, Hu R, Lin GX and
Ouyang W: Hippocampal volume reduction in elderly patients at risk
for postoperative cognitive dysfunction. J Anesth. 27:487–492.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen SQ, Wang PJ, Ten GJ, Zhan W, Li MH
and Zang FC: Role of myo-inositol by magnetic resonance
spectroscopy in early diagnosis of Alzheimer’s disease in APP/PS1
transgenic mice. Dement Geriatr Cogn Disord. 28:558–566. 2009.
View Article : Google Scholar
|
|
22
|
Covasala O, Stirn SL, Albrecht S, De Col R
and Messlinger K: Calcitonin gene-related peptide receptors in rat
trigeminal ganglion do not control spinal trigeminal activity. J
Neurophysiol. 108:431–440. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Krauspe R and Scheer A: Coomassie
brilliant blue G-250 dye-binding technique for determination of
autolytic protein breakdown in Euglena gracilis and comparison to
other methods of autolysis measurement. Anal Biochem. 153:242–250.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Oguri T, Takahata I, Katsuta K, Nomura E,
Hidaka M and Inagaki N: Proteome analysis of rat hippocampal
neurons by multiple large gel two-dimensional electrophoresis.
Proteomics. 2:666–672. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fountoulakis M and Gasser R: Proteomic
analysis of the cell envelope fraction of Escherichia coli. Amino
Acids. 24:19–41. 2003.PubMed/NCBI
|
|
26
|
Opii WO, Joshi G, Head E, Milgram NW,
Muggenburg BA, Klein JB, Pierce WM, Cotman CW and Butterfield DA:
Proteomic identification of brain proteins in the canine model of
human aging following a long-term treatment with antioxidants and a
program of behavioral enrichment: relevance to Alzheimer’s disease.
Neurobiol Aging. 29:51–70. 2008. View Article : Google Scholar
|
|
27
|
Berndt P, Hobohm U and Langen H: Reliable
automatic protein identification from matrix-assisted laser
desorption/ionization mass spectrometric peptide fingerprints.
Electrophoresis. 20:3521–3526. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nikolov M, Schmidt C and Urlaub H:
Quantitative mass spectrometry-based proteomics. Methods Mol Biol.
893:85–100. 2012.
|
|
29
|
Nagappan A, Park HS, Park KI, Kim JA, Hong
GE, Kang SR, Zhang J, Kim EH, Lee WS, Won CK and Kim GS: Proteomic
analysis of differentially expressed proteins in vitamin C-treated
AGS cells. BMC Biochem. 14:242013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nolan T, Hands RE and Bustin SA:
Quantification of mRNA using real-time RT-PCR. Nat Protoc.
1:1559–1582. 2006. View Article : Google Scholar
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C(T)). Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
32
|
Kipp M, Clarner T, Gingele S, Pott F, Amor
S, van der Valk P and Beyer C: Brain lipid binding protein (FABP7)
as modulator of astrocyte function. Physiol Res. 60(Suppl 1):
49–60. 2011.
|
|
33
|
Aubrecht J and Schomaker S: Serum
glutamate dehydrogenase as a potential biomarker of mitochondrial
dysfunction. Toxicol Sci. 134:2232013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Butterfield DA, Hensley K, Cole P, et al:
Oxidatively induced structural alteration of glutamine synthetase
assessed by analysis of spin label incorporation kinetics:
relevance to Alzheimer’s disease. J Neurochem. 68:2451–2457. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bedford PD: Adverse cerebral effects of
anaesthesia on old people. Lancet. 269:259–263. 1955. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mallozzi C, D’Amore C, Camerini S, Macchia
G, Crescenzi M, Petrucci TC and Di Stasi AM: Phosphorylation and
nitration of tyrosine residues affect functional properties of
Synaptophysin and Dynamin I, two proteins involved in
exo-endocytosis of synaptic vesicles. Biochim Biophys Acta.
1833:110–121. 2013. View Article : Google Scholar
|
|
37
|
Merriam LA, Baran CN, Girard BM, Hardwick
JC, May V and Parsons RL: Pituitary adenylate cyclase 1 receptor
internalization and endosomal signaling mediate the pituitary
adenylate cyclase activating polypeptide-induced increase in guinea
pig cardiac neuron excitability. J Neurosci. 33:4614–4422. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yamada H, Abe T, Satoh A, Okazaki N, Tago
S, Kobayashi K, Yoshida Y, Oda Y, Watanabe M, Tomizawa K, Matsui H
and Takei K: Stabilization of actin bundles by a dynamin
1/cortactin ring complex is necessary for growth cone filopodia. J
Neurosci. 33:4514–4526. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Koide T, Aleksic B, Ito Y, Usui H, Yoshimi
A, Inada T, Suzuki M, Hashimoto R, Takeda M, Iwata N and Ozaki N: A
two-stage case-control association study of the
dihydropyrimidinase-like 2 gene (DPYSL2) with schizophrenia in
Japanese subjects. J Hum Genet. 55:469–472. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang QP, Jammoul F, Duboc A, Gong J,
Simonutti M, Dubus E, Craft CM, Ye W, Sahel JA and Picaud S:
Treatment of epilepsy: the GABA-transaminase inhibitor, vigabatrin,
induces neuronal plasticity in the mouse retina. Eur J Neurosci.
27:2177–2187. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hwang IK, Kim DW, Yoo KY, Kim DS, Kim KS,
Kang JH, Choi SY, Kim YS, Kang TC and Won MH: Age-related changes
of gamma-aminobutyric acid transaminase immunoreactivity in the
hippocampus and dentate gyrus of the Mongolian gerbil. Brain Res.
1017:77–84. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wootz H, Fitzsimons-Kantamneni E,
Larhammar M, Rotterman TM, Enjin A, Patra K, André E, Van Zundert
B, Kullander K and Alvarez FJ: Alterations in the motor
neuron-renshaw cell circuit in the Sod1 (G93A) mouse model. J Comp
Neurol. 521:1449–1469. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kang SH, Li Y, Fukaya M, Lorenzini I,
Cleveland DW, Ostrow LW, Rothstein JD and Bergles DE: Degeneration
and impaired regeneration of gray matter oligodendrocytes in
amyotrophic lateral sclerosis. Nat Neurosci. 16:571–579. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Castro-Caldas M, Carvalho AN, Rodrigues E,
Henderson C, Wolf CR and Gama MJ: Glutathione S-transferase pi
mediates MPTP-induced c-Jun N-terminal kinase activation in the
nigrostriatal pathway. Mol Neurobiol. 45:466–477. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Korff A, Pfeiffer B, Smeyne M, Kocak M,
Pfeiffer RF and Smeyne RJ: Alterations in glutathione S-transferase
pi expression following exposure to MPP+ -induced
oxidative stress in the blood of Parkinson’s disease patients.
Parkinsonism Relat Disord. 17:765–768. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sánchez-Font MF, Sebastià J, Sanfeliu C,
Cristòfol R, Marfany G and Gonzàlez-Duarte R: Peroxiredoxin 2
(PRDX2), an antioxidant enzyme, is under-expressed in Down syndrome
fetal brains. Cell Mol Life Sci. 60:1513–1523. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tang WH, Martin KA and Hwa J: Aldose
reductase, oxidative stress and diabetic mellitus. Front Pharmacol.
3:872012. View Article : Google Scholar
|
|
48
|
Takase C, Nakano K, Ohta S, Nakagawa S and
Matuda SY: Different distribution of dihydrolipoamide
succinyltransferase, dihydrolipoamide acetyltransferase and ATP
synthase beta-subunit in monkey brain. In Vivo. 10:495–501.
1996.PubMed/NCBI
|
|
49
|
Ko YH, Pan W, Inoue C and Pedersen PL:
Signal transduction to mitochondrial ATP synthase: evidence that
PDGF-dependent phosphorylation of the delta-subunit occurs in
several cell lines, involves tyrosine and is modulated by
lysophosphatidic acid. Mitochondrion. 1:339–348. 2002. View Article : Google Scholar
|
|
50
|
Adam-Vizi V and Tretter L: The role of
mitochondrial dehydrogenases in the generation of oxidative stress.
Neurochem Int. 62:757–763. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Batke J, Nazaryan KB and Karapetian NH:
Complex of brain D-phosphoglycerate mutase and gamma enolase and
its reactivation by D-glycerate 2,3-bisphosphate. Arch Biochem
Biophys. 264:510–518. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhao Y, Pan X, Zhao J, Wang Y, Peng Y and
Zhong C: Decreased transketolase activity contributes to impaired
hippocampal neurogenesis induced by thiamine deficiency. J
Neurochem. 111:537–546. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tolle SW, Dyson RD, Newburgh RW and
Cardenas JM: Pyruvate kinase isozymes in neurons, glia,
neuroblastoma and glioblastoma. J Neurochem. 27:1355–1360. 1976.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ahmed Z, Shaw G, Sharma VP, Yang C,
McGowan E and Dickson DW: Actin-binding proteins coronin-1a and
IBA-1 are effective microglial markers for immunohistochemistry. J
Histochem Cytochem. 55:687–700. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zaheer A, Zaheer S, Sahu SK, Knight S,
Khosravi H, Mathur SN and Lim R: A novel role of glia maturation
factor: induction of granulocyte-macrophage colony-stimulating
factor and pro-inflammatory cytokines. J Neurochem. 101:364–376.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Shao W, Zhang SZ, Tang M, Zhang XH, Zhou
Z, Yin YQ, Zhou QB, Huang YY, Liu YJ, Wawrousek E, Chen T, Li SB,
Xu M, Zhou JN, Hu G and Zhou JW: Suppression of neuroinflammation
by astrocytic dopamine D2 receptors via αB-crystallin. Nature.
494:90–94. 2013. View Article : Google Scholar
|
|
57
|
An LN, Yue Y, Guo WZ, Miao YL, Mi WD,
Zhang H, Lei ZL, Han SJ and Dong L: Surgical trauma induces iron
accumulation and oxidative stress in a rodent model of
postoperative cognitive dysfunction. Biol Trace Elem Res.
151:277–283. 2013. View Article : Google Scholar
|
|
58
|
Cooper AJ: Possible treatment of end-stage
hyperammonemic encephalopathy by inhibition of glutamine
synthetase. Metab Brain Dis. 28:119–125. 2013. View Article : Google Scholar
|
|
59
|
Plaitakis A, Zaganas I and Spanaki C:
Deregulation of glutamate dehydrogenase in human neurologic
disorders. J Neurosci Res. 91:1007–1017. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Frigerio F, Karaca M, De Roo M, Mlynárik
V, Skytt DM, Carobbio S, Pajęcka K, Waagepetersen HS, Gruetter R,
Muller D and Maechler P: Deletion of glutamate dehydrogenase 1
(Glud1) in the central nervous system affects glutamate handling
without altering synaptic transmission. J Neurochem. 123:342–348.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Skytt DM, Klawonn AM, Stridh MH, Pajęcka
K, Patruss Y, Quintana-Cabrera R, Bolaños JP, Schousboe A and
Waagepetersen HS: siRNA knock down of glutamate dehydrogenase in
astrocytes affects glutamate metabolism leading to extensive
accumulation of the neuroactive amino acids glutamate and
aspartate. Neurochem Int. 61:490–497. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hovens IB, Schoemaker RG, van der Zee EA,
Heineman E, Nyakas C and van Leeuwen BL: Surgery-induced behavioral
changes in aged rats. Exp Gerontol. 48:1204–1211. 2012. View Article : Google Scholar
|