Introduction
Lung cancer is one of the most common causes of
cancer-associated mortality worldwide (1). Non-small cell lung cancer (NSCLC) is
predominantly divided into squamous cell carcinoma and
adenocarcinoma (1). The five-year
survival rate for NSCLC is ~10–15% (2). The postoperative resistance of
patients with lung cancer to chemotherapy drugs, including
cisplatin and tyrosine kinase inhibitors (TKI), is a significant
factor influencing disease prognosis. Therefore, exploration into
chemotherapy resistance may help improve the treatment of patients
with lung cancer.
Long non-coding RNAs (lncRNAs) belong to a novel
heterogeneous class of ncRNAs, and are involved in various
biological processes, including imprinting (3), histone-code regulation and
proliferation (4) of cancer cells,
through regulation of gene expression (5). lncRNA H19 is able to regulate the
induction of multidrug resistance protein 1-associated drug
resistance in hepatocellular carcinoma cells (6). A previous study demonstrated that
lncRNA UCA1 may enhance the chemoresistance of bladder cancer cells
via regulation of WNT signaling (7). A novel lncRNA ARA has been shown to
contribute to adriamycin resistance in cancer, through the
modulation of numerous signaling pathways (8). Maternally expressed gene 3 (Meg3) is
a tumor suppressor gene that encodes an lncRNA and is expressed in
numerous types of normal tissue; however, its expression is lost in
multiple cancer cell lines that originate from various types of
tissue (9). Previous studies have
demonstrated that Meg3 may inhibit proliferation (10–12)
and induce apoptosis (10,13) of tumor cells in vitro.
However, little is currently known regarding the role of Meg3 in
the development of chemoresistant phenotypes of lung cancer.
Cisplatin is a commonly used anti-cancer drug, which induces
apoptosis by suppressing the DNA replication process of cancer
cells (14).
Alterations in the WNT/β-catenin signaling pathway
are frequent amongst human malignancies. In NSCLC, mutations in
β-catenin are uncommon; however, WNT signaling is significant in
NSCLC cell lines and WNT inhibition results in increased levels of
apoptosis (15). WNT signaling has
a substantial impact on NSCLC tumorigenesis, prognosis and
resistance to therapy, and a loss of WNT signaling inhibitors by
promoter hypermethylation or other mechanisms appears to be
particularly important (15).
It was hypothesized that Meg3 may regulate
apoptosis, cell cycle distribution and drug resistance of cancer
cells via the inhibition of β-catenin/survivin by activated p53.
The present study therefore aimed to elucidate the mechanisms
underlying the effects of Meg3 on apoptosis, cell cycle and drug
resistance in NSCLC. The expression levels of Meg3 in A549/DDP
cisplatin-resistant lung cancer cells and A549 parental lung cancer
cells were evaluated and; furthermore, Meg3-mediated
chemosensitivity enhancement was investigated in these cell
lines.
Materials and methods
Cell culture and blood samples
A549 and A549/DDP human lung cancer cell lines were
used in the present study. The cells were cultured in RPMI-1640
medium (Invitrogen Life Technologies, Carlsbad, CA, USA)
supplemented with 10% fetal bovine serum (FBS; Invitrogen Life
Technologies) and penicillin (100 U/ml; Invitrogen Life
Technologies), at 37°C in an atmosphere containing 5%
CO2. Blood samples (n=27) were collected from patients
with advanced lung cancer that had received cisplatin-based
chemotherapy between May 2010 and December 2012, at the Affiliated
Hospital of Jiangnan University (Wuxi, China). The median patient
age was 64 (range, 49–85 years). The patients included 16 females
and 11 males. All specimens were histologically classified by a
professional pathologist, according to the national NCCN guidelines
for NSCLC version 3.2011 (16) in
a blinded manner. Written informed consent was obtained from all of
the patients recruited. The methodologies of the present study
conformed to the standards set by the Declaration of Helsinki; and
the study was approved by the Ethical Committee of the Affiliated
Hospital of Jiangnan University (Wuxi, China)
Plasmid construction and cell
transfection
The sequence of Meg3 was synthesized by Genewiz
(Suzhou, China). All recombinant lentiviruses were produced by
calcium phosphate-mediated transient transfection of HEK293T cells,
according to standard protocols. Briefly, HEK293T cells from
Clontech were cultured in Dulbecco's modified Eagle's medium
(DMEM), supplemented with 10% FBS and penicillin/streptomycin (100
U/ml). The subconfluent cells in a 10 cm culture dish were
co-transfected with lentiviral vector (10 µg), the
lentiviral packaging vectors pLv-GFP or pLv-Meg3 (2 µg),
pMDLg/pRRE (5 µg), and the vesicular stomatitis virus G
glycoprotein (VSVG) expression vector pMD2G (3 µg). The
viruses were collected from the culture supernatants on days 2 and
3 post-transfection. The cultured cells were incubated with
lentivirus at various concentrations for 24 h and cultured with
DMEM, containing 10% FBS. A pure population of infected cells were
sorted based on the GFP-expression by flow cytometry and >98% of
the cells were GFP-positive following sorting. The plasmid was
subcloned into a lentiviral vector, and then co-transfected into
HEK-293T human embryonic kidney cells using the Lentiviral
Packaging Mix. A549/DDP cells were transfected with the lentivirus,
and cells which highly expressed Meg3 (A549/DDP-Meg3 cells) were
identified by selection with G418. The Meg3 sequence (NR_002766.2)
was subcloned into an HIV type-1 lentiviral vector, pLV-GFP
(Addgene, Cambridge, MA, USA) by restriction digestion using
BamHI and MluI (New England Biolabs, Ltd.,
Hertfordshire, United Kingdom). The resulting constructs was termed
pLV-Meg3 and pLV-GFP was used as a control. Following
trypsinization, the cells were centrifuged at 1000 × g for 5 min
and the supernatant was removed. A total of 1×106 cells
were subsequently seeded into six-well plates at high density
(80–90% confluent). Lipofectamine 2000 (Invitrogen Life
Technologies) complexes were prepared, according to the standard
Lipofectamine protocol. Lipofectamine 2000 reagent (10 µl)
and 100 pmol siRNA were each diluted in 250 µl Opti-MEM
medium. Each mix was pooled and incubated for 5 min prior to the
DNA-reagent complex being added to the cells (500 µl per
well). The cells were incubated for 6 h at 37°C and subsequently
incubated with DMEM, containing 10% FBS for 24 h.
Transfection of small interfering
(si)RNAs
The A549 cells were seeded at 50–80% cell density
into six-well plates and transfected with 50 nM siRNAs specifically
targeting Meg3 (siRNA/Meg3-243: 5′-GAUCCCACCAACAUACAAATT-3′;
siRNA/Meg3-405: 5′-GCUCAUACUUUGACUCUAUTT-3′; siRNA/Meg3-852:
5′-CCCUCUUGCUUGUCUUACUTT-3′) (Shanghai Gene Pharma Co., Ltd.,
Shanghai, China) using Lipofectamine®2000 (Invitrogen
Life Technologies), according to the manufacturer's instructions,
and were subsequently incubated for 24 h at 37°C in a 5%
CO2 incubator. The control siRNA sequence was as
follows: 5′-UUAAGACGGUUGAAACUAG-3′.
Isolation of total RNA and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the cells and blood
samples using TRIzol® (Invitrogen Life Technologies) and
was subsequently reverse transcribed into cDNA using a reverse
transcriptase kit (Takara Bio, Inc., Otsu, Japan). The relative
mRNA expression levels of Meg3 were examined by RT-qPCR using SYBR
Premix Ex Taq (Takara Biotechnology Co., Ltd., Dalian, China), and
normalized to GAPDH. The RT-qPCR was performed using the ABI 7500
Fast Real-Time PCR system (Applied Biosystems, Foster City, CA,
USA). The PCR conditions were as follows: 94°C for 2 min, 94°C for
30 sec, 60°C for 30 sec and 72°C for 1 min for 30 cycles, and 72°C
for 10 min. The PCR primer sequences used were as follows: MEG3,
forward: 5′-ATCATCCGTCCACCTCCTTGTCTTC-3′ and reverse:
5′-GTATGAGCATAGCAAAGGTCAGGGC-3′; GAPDH, forward:
5′-AATGCCTCCTGCACCACCAAC-3′ and reverse:
5′-AAGGCCATGCCAGTGAGCTTC-3′. Calculations of expression were made
using the 2−ΔΔCT method.
Flow cytometric analysis of apoptosis and
cell cycle distribution
Apoptosis of the cultured A549 and A549/DDP cells
was evaluated using Annexin V labeling. An Annexin
V-Allophycocyanin-Labeled Apoptosis Detection kit (Abcam,
Cambridge, UK) was used according to the manufacturer's
instructions. Staining with annexin V/PI was performed using an
Annexin V-APC Apoptosis detection kit (Invitrogen Life
Technologies). Briefly, an aliquot containing 2×106
cells was resuspended in 0.5 ml binding buffer, labelled with 1
µl annexin V-APC plus 5 µl PI and were incubated for
10 min in the dark. The samples were immediately analyzed. Annexin
negative and PI negative cells were designated as live cells and
annexin positive and PI negative as early apoptotic cells. Staining
with PI was performed using cell cycle detection kit (invitrogene,
USA). Briefly, the cells were centrifuged at 500 × g for 10 min at
room temperature and the supernatant was removed. An aliquot of
~1×106 was incubated in ethyl alcohol at −20°C for 24 h
and was subsequently incubated in 50 µg/ml PI in the dark,
at room temperature for 30 min. The cells undergoing cell-cycle
analysis were stained with propidium iodide, using the BD Cycletest
Plus DNA Reagent kit (BD Biosciences, Shanghai, China), according
to the manufacturer's instructions. The results were analyzed using
a FACScan flow cytometer (BD Biosciences). The percentage of cells
in each of the subG0/G1, G1, S or
G2/M phases were counted and compared. Each experiment
was performed at least three times.
MTT assay
The chemosensitivity of the cisplatin-resistant
A549/DPP and parental A549 cells to cisplatin was determined by MTT
assay. Briefly, the cells were seeded into 96-well plates
(3.5×103 cells/well) and treated with various
concentrations of cisplatin (0, 1, 5, 8, 10, 15, 18, 20, 22 and 24
µg/ml). At 0, 24, 48, 72 and 96 h post-cisplatin
application, cell survival was assessed using 0.5 mg/ml MTT
(Sigma-Aldrich, St. Louis, MO, USA) solution. Following a 4 h
incubation at 37°C, the medium was replaced with 150 µl
dimethyl sulfoxide (Sigma-Aldrich) and vortexed for 10 min. The
absorbance of each well at 490 nm was measured using a microplate
reader 550 (Bio-Rad, Richmond, CA, USA). Each experiment was
performed at least three times.
Western blot analysis
The A549/DDP cells were washed in phosphate-buffered
saline (PBS) and lysed in radioimmuno-precipitation assay lysis
buffer supplemented with a protease inhibitor cocktail (Roche
Diagnostics, Mannheim, Germany). The total protein was quantified
using a Bicinchoninic Acid Protein Assay kit (Beyotime Institute of
Biotechnology, Haimen, China). Equal amounts (30 µg) of the
whole cell lysates were separated by 5% SDS-PAGE (Beyotime
Institute of Biotechnology, Jiangsu, China) and transferred onto
polyvinylidene difluoride membranes (Merck Millipore GmbH,
Eschborn, Germany). The blots were blocked in bovine serum albumin
(5% w/v in PBS+0.1% Tween 20) for 1 h at room temperature and
immunostained with the following antibodies at 4°C overnight: Mouse
anti-human p53 (1:2,000; monoclonal, cat. no. ab26), mouse
anti-human β-catenin (1:1,000; monoclonal, cat. no. ab6301), mouse
anti-human survivin (1:500; monoclonal, cat. no. ab118046) and
mouse anti-human GAPDH (1:5,000; monoclonal, cat. no. ab9484).
These antibodies were purchased from Abcam (Cambridge, MA, USA).
The membranes were washed four times with PBS, containing 0.1%
Tween 20. The secondary antibody, goat anti-mouse (1:50,000;
polyclonal, cat. no. ab97040; Abcam), was added in PBS, containing
0.1% Tween 20 for 1 h at 37°C. The membranes were then washed three
times for 15 min with PBS, containing 0.1% Tween 20
(Sigma-Aldrich). The results were visualized using an enhanced
chemiluminescent detection system (Pierce ECL Substrate Western
Blot Detection system; Thermo Fisher Scientific, Waltham, MA, USA)
and using a Molecular Imager ChemiDoc XRS system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
Statistical analysis was performed using STATA 11
(StataCorp LP, College Station, TX, USA), and presented using
GraphPad Prism 4.0 software (GraphPad Software, Inc., La Jolla, CA,
USA). The results obtained from the in vitro assays are
presented as the mean ± standard error of the mean from five
separate experiments performed in triplicate. P<0.05 was
considered to indicate a statistically significant difference.
Results
Meg3 expression levels are markedly lower
in the A549/DDP cisplatin-resistant cell line, compared with those
in the A549 parental cell line
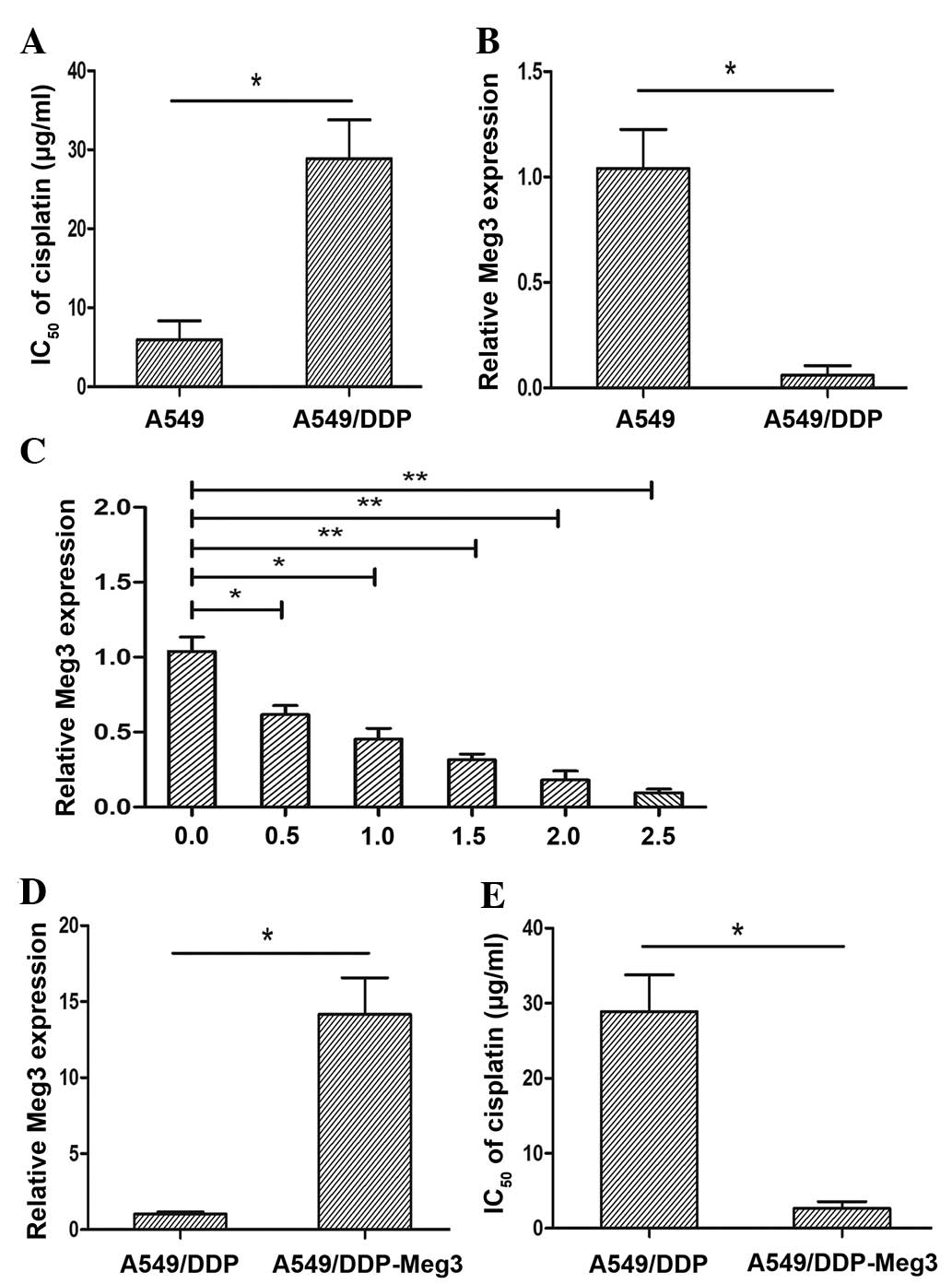
A549/DDP is a cisplatin-resistant lung cancer cell
line. The present study conducted an MTT assay to determine the
half maximal inhibitory concentration (IC50) of
cisplatin in the A549/DDP and parental A549 cell lines. The
IC50 of cisplatin in the A549 and A549/CDDP cells was
5.68±2.54 µg/ml and 29.24±1.62 µg/ml, respectively
(P<0.05; Fig. 1A). The A549/DDP
cells exhibited a 5.15-fold greater resistance to cisplatin
compared with that of the parental A549 cell line. To further
investigate whether Meg3 had a significant role in the acquired
cisplatin resistance of lung cancer cells, the expression levels of
Meg3 were detected in the A549/DDP and parental A549 cells by
RT-qPCR. The expression levels of Meg3 were significantly lower in
the A549/DDP cisplatin-resistant cell line, as compared with in the
A549 parental cell line (P<0.05; Fig. 1B).
Treatment with cisplatin reduces
expression of Meg3
The results above demonstrated that the expression
levels of Meg3 were upregulated in A549 cells. The present study
aimed to determine the expression levels of Meg3 in response to
various concentrations of cisplatin in A549 cells. The cells were
treated with various concentrations of cisplatin (0.0, 0.5, 1.0,
1.5, 2.0 and 2.5 µg/ml) for 24 h. The relative expression
levels of Meg3 were decreased in response to treatment with
increasing concentrations of cisplatin (Fig. 1C). These results suggested that a
reduction in Meg3 expression levels in the A549 lung cancer cell
line was induced in response to cisplatin treatment.
Meg3 upregulation reverses the cisplatin
resistance of A549/DPP cells
To further evaluate the effects of Meg3 on the
development of cisplatin resistance in lung cancer cells, a
lentivirus overexpressing Meg3 was stably transfected into the
A549/DDP cells (A549/DDP-Meg3). The transfection efficiency was
validated by RT-qPCR. The expression levels of Meg3 in the
A549/DDP-Meg3 cells were significantly upregulated, compared with
those of the A549/DDP control cells (P<0.05; Fig. 1D). Furthermore, overexpression of
Meg3 led to a reduction in the IC50 of cisplatin in
A549/DDP cells by ~9.79-fold (P<0.05; Fig. 1E).
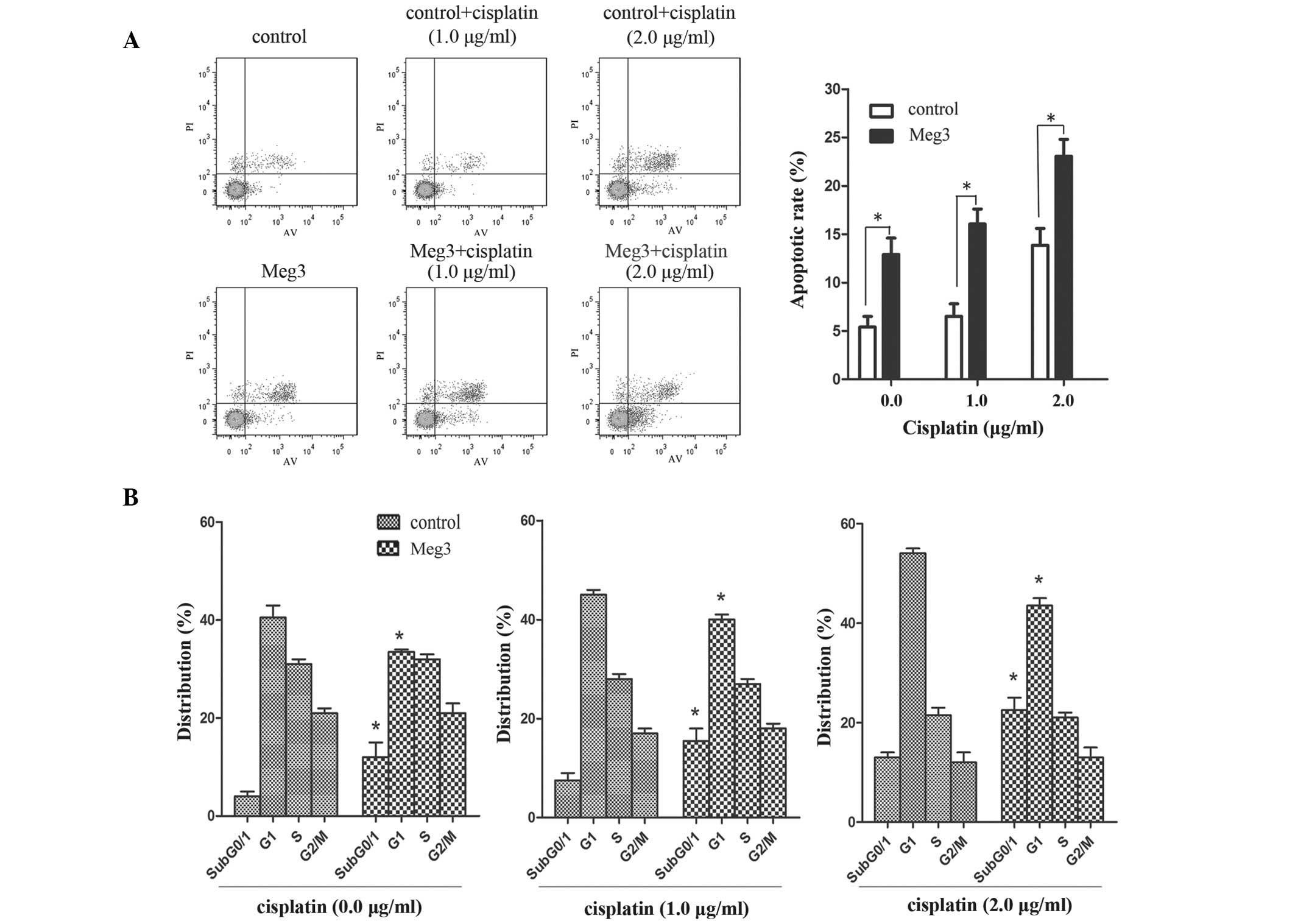
In the A549/DDP-Meg3 cells treated with cisplatin
(0.0, 1.0 and 1.5 µg/ml), upregulation of Meg3 resulted in
an increased rate of cisplatin-induced apoptosis (P<0.05;
Fig. 2A). Furthermore, the
percentage of A549/DDP-Meg3 cells in the
subG0/G1 and G1 phases,
particularly at checkpoint subG0/1, of the
cell cycle gradually increased with the increasing doses of
cisplatin (P<0.05; Fig. 2B).
These results suggested that upregulation of Meg3 may reverse the
cisplatin resistance of A549/DDP cells by inducing apoptosis and
cell cycle arrest.
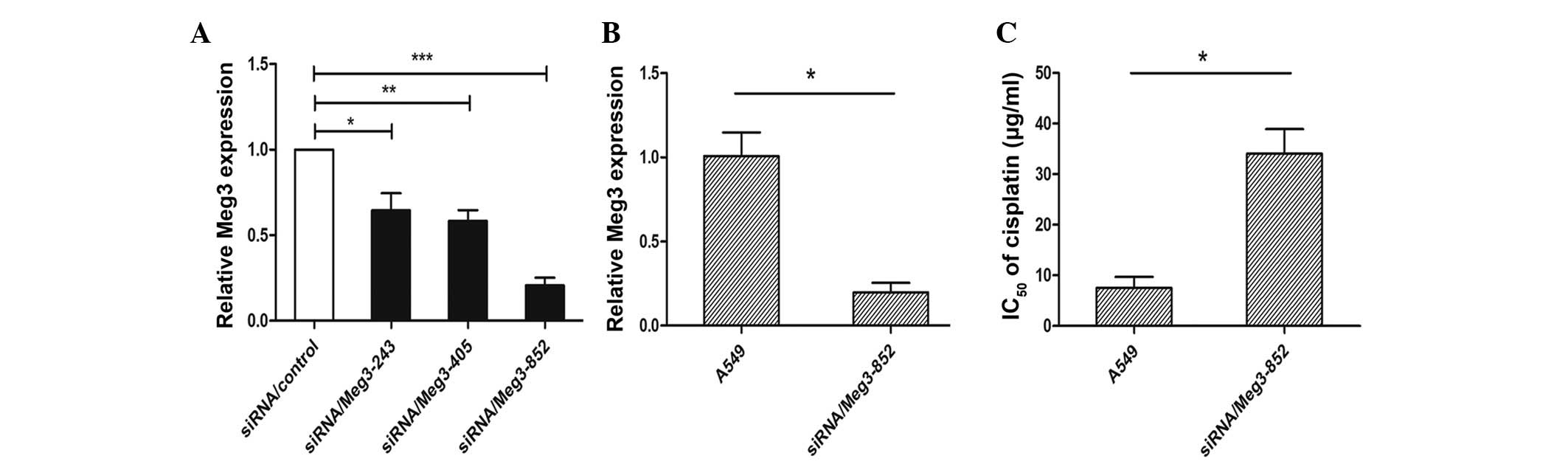
siRNA knockdown of Meg3 reduces the rate
of cisplatin-induced apoptosis and alters cell cycle distribution
in A549 cells
A549 cells were transfected with siRNA/control,
siRNA/Meg3-243, siRNA/Meg3-405 or siRNA/Meg3-852, in order to
analyze the role of Meg3 in determining the sensitivity of lung
cancer cells to cisplatin. At 48 h post-transfection, the
expression levels of Meg3 were significantly decreased by ~75.5% in
the A549-siRNA/Meg3-852 cells compared with the
siRNA/control-transfected cells (P<0.01; Fig. 3A). The Meg3 inhibition efficiency
was higher with this siRNA than that of siRNA/Meg3-243 (30.1%) and
siRNA/Meg3-405 (35.3%). Based on these results, siRNA/Meg3-852 was
used to suppress the expression of Meg3 in A549 cells (Fig. 3B). The effects of Meg3 expression
on the IC50 of cisplatin in A549 cells were subsequently
investigated. An MTT assay demonstrated that siRNA/Meg3-852
increased the IC50 of cisplatin in A549 cells
(P<0.05; Fig. 3C). Furthermore,
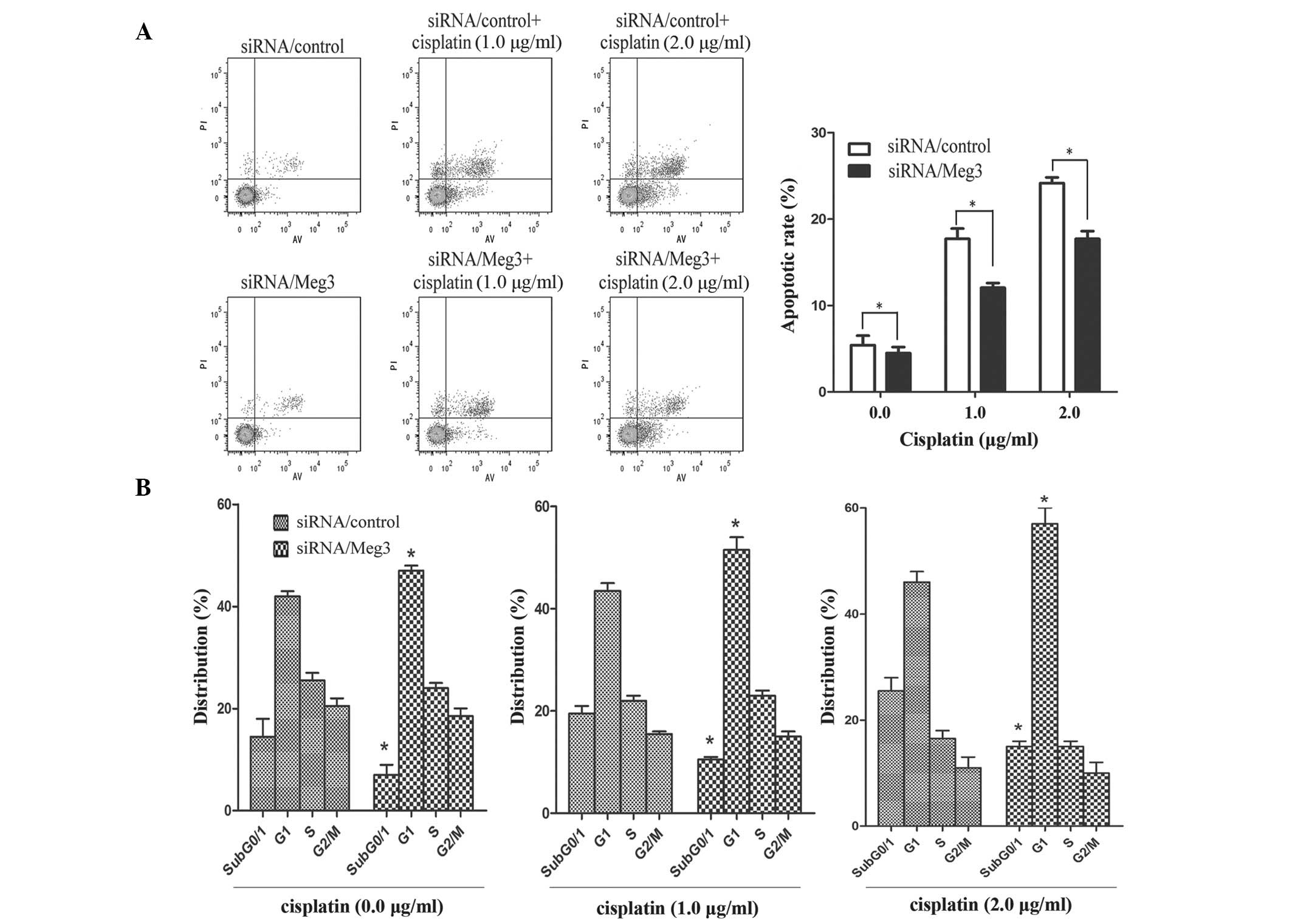
A549-siRNA/Meg3-852 cells were treated with various concentrations
of cisplatin (0.0, 1.0 and 2.0 µg/ml). An apoptosis assay
demonstrated that knockdown of Meg3 by RNA interference reduced the
rate of cisplatin-induced apoptosis in lung cancer cells
(P<0.05; Fig. 4A). In addition,
the percentage of siRNA/Meg3-transfected A549 cells in
subG0/G1 and G1 phases,
particularly in checkpoint G1, of the cell cycle
increased with increasing doses of cisplatin (P<0.05; Fig. 4B).
Upregulation of Meg3 in A549/DDP cells
decreases cisplatin resistance through inhibition of WNT/ß-catenin
signaling
A previous study reported that Meg3 was able to
regulate the specificity of p53 transcriptional activation
(17). The tumor suppressor gene
p53 has a central role in tumor suppression, and mediates the
functions of numerous tumor suppressors, in particular those
associated with apoptosis and drug resistance of cancer cells
(18). Furthermore, it has been
demonstrated that the overexpression of wild-type p53, by
transfection or DNA damage, downregulates the expression of
β-catenin in human cells (19).
The inhibitory effects of p53 on β-catenin have been revealed to be
mediated by the ubiquitin-proteasome system and require activation
of glycogen synthase kinase-3β (GSK-3β) (19). WNTs regulate embryonic development
and determine cell fate, and dysregulation of WNT/β-catenin
signaling may induce the development and/or progression of various
human diseases, including cancer (20). Therefore, analyzing the effects of
potential therapies on the WNT/β-catenin signaling pathway may
represent a novel method for the screening of putative anti-cancer
drugs. Gao et al (21)
identified potential drugs that were able to inhibit WNT/β-catenin
signaling, including cisplatin, using a drug screening platform
based on β-catenin/transcription factor-mediated transcriptional
activity. In addition, cisplatin was shown to activate
WNT/β-catenin signaling in A549/DDP cells, and inactivate it in
A549 cells (21). Based on these
prior studies, the present study hypothesized that Meg3 may
regulate the cisplatin resistance of cancer cells via the
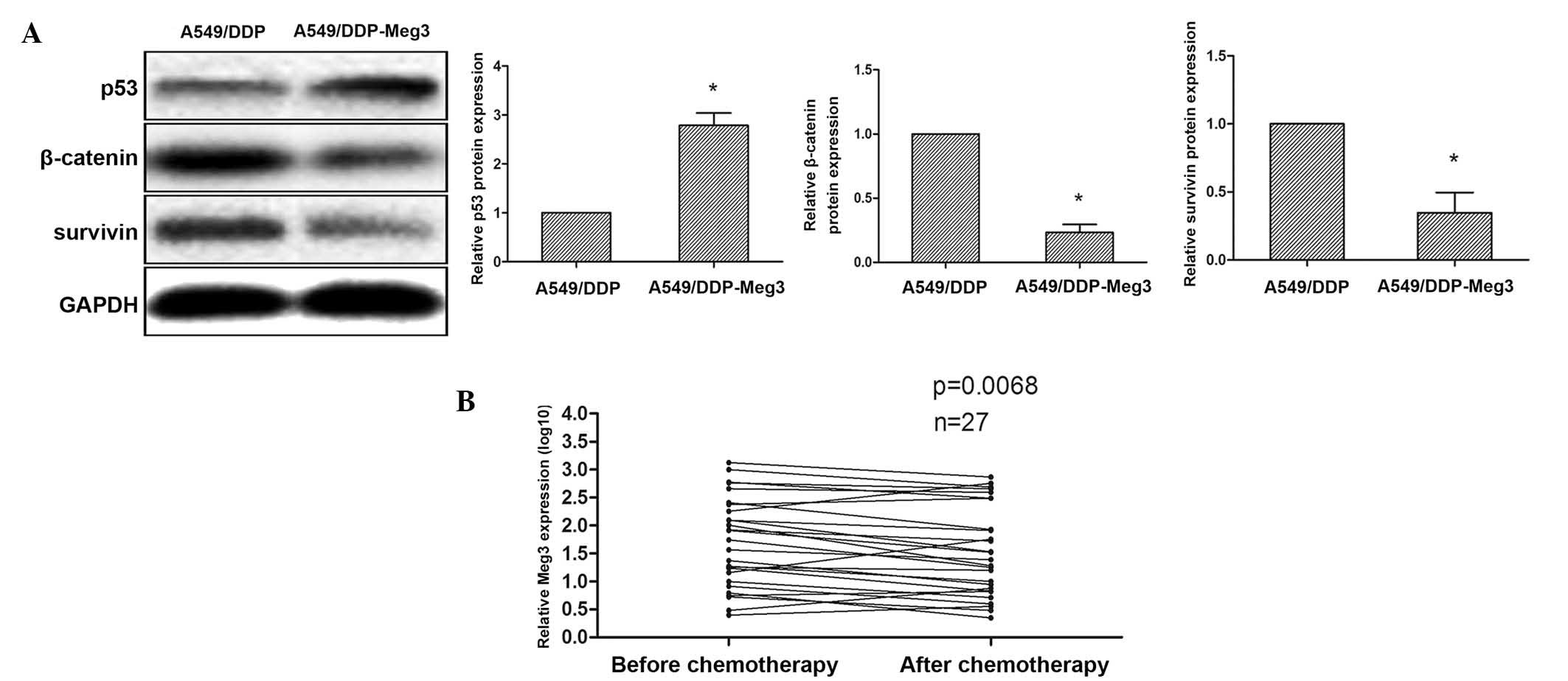
inhibition of β-catenin/survivin by activated p53. To examine this
hypothesis, the protein expression levels of p53, β-catenin and
survivin were determined in A549/DDP-Meg3 and A549/DDP cells by
western blot analysis. Upregulation of p53 and downregulation of
β-catenin/survivin was observed in the A549/DDP-Meg3 cells, and
conversely the expression levels of these proteins were reversed in
the A549/DDP cells (Fig. 5A).
These findings suggested that upregulation of Meg3 in A549/DDP
cells may decrease cisplatin resistance via inhibition of
WNT/β-catenin signaling.
Meg3 expression levels are downregulated
in blood samples from patients with advanced lung cancer following
cisplatin-based chemotherapy
In a clinical setting, cisplatin is frequently used
to treat patients with advanced lung cancer. In the present study,
blood samples (n=27) were collected from patients with advanced
lung cancer in order to determine the effects of cisplatin
treatment on Meg3 expression. RT-qPCR analysis demonstrated that
the expression levels of Meg3 were decreased in patients with
advanced lung cancer following treatment with cisplatin-based
chemotherapy, compared with those prior to cisplatin-based
chemotherapy (P=0.0068; Fig. 5B).
These results suggested that Meg3 may have a significant role as a
therapeutic target in lung cancer.
Discussion
Lung cancer is one of the most common causes of
cancer-associated mortality worldwide. The postoperative resistance
of patients to chemotherapy drugs, including cisplatin and TKI, is
a significant factor in determining the prognosis of patients with
lung cancer. Therefore, exploration into the mechanisms underlying
cisplatin resistance are crucial for improving treatment outcomes.
lncRNAs are a class of non-coding RNAs, which have been implicated
in the development of various cancers. Emerging literature has
demonstrated that numerous lncRNAs are associated with the
biological processes underlying various types of cancer (22–24).
The lncRNA SChLAP1 has been shown to contribute to the development
of lethal types of prostate cancer, at least in part by
antagonizing the tumor-suppressive functions of the switch/sucrose
nonfermentable complex (25). In
hepatocellular cancer, the lncRNA low expression in tumor is
suppressed by histone deacetylase 3, and may contribute to
hypoxia-mediated metastasis (26).
The present study investigated the association of the lncRNA Meg3
with the cell cycle, apoptosis and cisplatin resistance of human
lung cancer cell lines.
It remains a major therapeutic challenge amongst
numerous patients with cancer who are treated with anti-cancer
drugs, that they exhibit significant chemoresistance, including
intrinsic and acquired resistance (27). The study of tumor chemotherapy drug
resistance has continued to grow; however, the potential mechanisms
underlying the resistance to chemotherapeutic agents remain
unclear. Recent studies have demonstrated that substantial
epigenetic alterations may contribute to facilitating successful
chemotherapeutic treatment in drug-resistant cancer cells (28,29).
The results of the present study demonstrated that the expression
levels of Meg3 were markedly lower in the cisplatin-resistant cell
line A549/DDP, as compared with those in the A549 parental cell
line. In addition, reduced expression levels of Meg3 were detected
in the A549 lung cancer cell line in response to treatment with
cisplatin. Furthermore, upregulation of Meg3 resulted in an
increased rate of cisplatin-induced apoptosis of A549/DDP-Meg3
cells; whereas, knockdown of Meg3 expression by RNA interference
reduced the cisplatin-induced apoptosis of lung cancer cells.
The percentage of A549/DDP-Meg3 cells in
subG0/G1 and G1 phase,
particularly at checkpoint subG0/1, of the
cell cycle gradually increased with increasing doses of cisplatin,
whereas the percentage of siRNA/Meg3 transfected A549 cells in
subG0/G1 and G1 phase,
particularly at checkpoint G1, of the cell cycle
gradually increased with increasing doses of cisplatin. Braconi
et al (30) previously
demonstrated that Meg3 inhibited the percentage of hepatocellular
cancer cells in subG0/1 phase of the cell
cycle, without significantly modulating the percentage of cells at
checkpoint G0/1 or G2/M. These
results were concordant with the findings of the present study. The
tumor suppressor gene p53 contributes to the apoptosis, cell cycle
and drug resistance of cancer cells. Meg3 has previously been
suggested to regulate the specificity of p53 transcriptional
activation (17). Butz and Patoc
(31) reported that Meg3 activates
p21CIP1 through p53, and its checkpoint is G1/S in the
pituitary gland. However, Zhou et al (17) reported that Meg3 does not stimulate
p21CIP1 expression. These contradictory results suggest that
changes in apoptosis, cell cycle distribution and drug resistance
of cancer cells should not be attributed to alterations of just
one, or a small quantity of genes.
Cisplatin resistance is associated with altered
cellular signaling, including the phosphoinositide-3-kinase-Akt,
p53, and WNT/β-catenin signaling pathways (32–34).
β-catenin is a key component of the WNT signaling pathway and is
subsequently phosphorylated by GSK-3β, which targets β-catenin for
ubiquitination and proteolytic degradation. Activation of
WNT/β-catenin signaling inhibits formation of the multi-protein
complex, and phosphorylation of β-catenin by GSK-3β. A previous
study demonstrated that high levels of survivin inhibited the
induction of apoptosis by cisplatin and resulted in drug resistance
in A549/DDP cells; however, a lack of survivin enhanced
cisplatin-mediated apoptosis in A549 cells (35). Based on the results of previous
studies, the present study hypothesized that Meg3 may regulate
apoptosis, cell cycle distribution and drug resistance of cancer
cells via the inhibition of β-catenin/survivin by activated p53.
The present study aimed to elucidate the mechanism underlying the
effects of Meg3 on apoptosis, cell cycle and drug resistance within
the limits of NSCLC. However, the biological and pathological
functions of Meg3 in general remain to be determined.
In conclusion, to the best of our knowledge, the
present study was the first to demonstrate that the expression of
Meg3 was decreased in the A549/DDP cisplatin-resistant cell line,
as compared with A549 cells. The expression levels of Meg3 were
also shown to be downregulated in blood samples taken from patients
with advanced lung cancer following treatment with cisplatin-based
chemotherapy. The present study also demonstrated that
downregulation of Meg3 enhanced the cisplatin resistance of lung
cancer cells through activation of the WNT/β-catenin signaling
pathway, thus indicating that Meg3 may have a significant role as a
therapeutic target in lung cancer.
Acknowledgments
The authors of the present study would like to thank
Dr Junwei Tang at The First Affiliated Hospital of Nanjing Medical
University (Nanjing, China) for language revision.
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sánchez de Cos J, Sojo González MA,
Montero MV, et al: Non-small cell lung cancer and silent brain
metastasis. Survival and prognostic factors. Lung Cancer.
63:140–145. 2009. View Article : Google Scholar
|
|
3
|
Chen CL, Ip SM, Cheng D, Wong LC and Ngan
HY: Loss of imprinting of the IGF-II and H19 genes in epithelial
ovarian cancer. Clin Cancer Res. 6:474–479. 2000.PubMed/NCBI
|
|
4
|
Yang F, Zhang L, Huo XS, et al: Long
noncoding RNA high expression in hepatocellular carcinoma
facilitates tumor growth through enhancer of zeste homolog 2 in
humans. Hepatology. 54:1679–1689. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lai MC, Yang Z, Zhou L, et al: Long
non-coding RNA MALAT-1 overexpression predicts tumor recurrence of
hepatocellular carcinoma after liver transplantation. Med Oncol.
29:1810–1816. 2012. View Article : Google Scholar
|
|
6
|
Tsang WP and Kwok TT: Riboregulator H19
induction of MDR1-associated drug resistance in human
hepatocellular carcinoma cells. Oncogene. 26:4877–4881. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fan Y, Shen B, Tan M, et al: Long
non-coding RNA UCA1 increases chemoresistance of bladder cancer
cells by regulating Wnt signaling. FEBS J. 281:1750–1758. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jiang M, Huang O, Xie Z, et al: A novel
long non-coding RNA-ARA: Adriamycin resistance-associated. Biochem
Pharmacol. 87:254–283. 2014. View Article : Google Scholar
|
|
9
|
Gejman R, Batista DL, Zhong Y, et al:
Selective loss of MEG3 expression and intergenic differentially
methylated region hypermethylation in the MEG3/DLK1 locus in human
clinically nonfunctioning pituitary adenomas. J Clin Endocrinol
Metab. 93:4119–4125. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun M, Xia R, Jin F, et al: Downregulated
long noncoding RNA MEG3 is associated with poor prognosis and
promotes cell proliferation in gastric cancer. Tumour Biol.
35:1065–1073. 2014. View Article : Google Scholar
|
|
11
|
Ying L, Huang Y, Chen H, et al:
Downregulated MEG3 activates autophagy and increases cell
proliferation in bladder cancer. Mol Biosyst. 9:407–411. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang P, Ren Z and Sun P: Overexpression of
the long non-coding RNA MEG3 impairs in vitro glioma cell
proliferation. J Cell Biochem. 113:1868–1874. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qin R, Chen Z, Ding Y, Hao J, Hu J and Guo
F: Long non-coding RNA MEG3 inhibits the proliferation of cervical
carcinoma cells through the induction of cell cycle arrest and
apoptosis. Neoplasma. 60:486–492. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao X, Xu Z, Wang Z, et al: RNA silencing
of integrin-linked kinase increases the sensitivity of the A549
lung cancer cell line to cisplatin and promotes its apoptosis. Mol
Med Rep. 12:960–966. 2015.PubMed/NCBI
|
|
15
|
Stewart DJ: Wnt signaling pathway in
non-small cell lung cancer. J Natl Cancer Inst. 106:djt3562014.
View Article : Google Scholar
|
|
16
|
Ward JH: NCCN Guidelines and the
International Community. J Natl Compr Canc Netw. 9:133–134.
2011.PubMed/NCBI
|
|
17
|
Zhou Y, Zhong Y, Wang Y, et al: Activation
of p53 by MEG3 non-coding RNA. J Biol Chem. 282:24731–24742. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Miao W, Liu X, Wang H, et al: p53
upregulated modulator of apoptosis sensitizes drug-resistant U251
glioblastoma stem cells to temozolomide through enhanced apoptosis.
Mol Med Rep. 11:4165–4173. 2015.PubMed/NCBI
|
|
19
|
Sadot E, Geiger B, Oren M and Ben-Ze'ev A:
Down-regulation of beta-catenin by activated p53. Mol Cell Biol.
21:6768–6781. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu MY, Xie X, Xu ZK, et al: PP2A
inhibitors suppress migration and growth of PANC-1 pancreatic
cancer cells through inhibition on the Wnt/beta-catenin pathway by
phosphorylation and degradation of β-catenin. Oncol Rep.
32:513–522. 2014.PubMed/NCBI
|
|
21
|
Gao Y, Liu Z, Zhang X, et al: Inhibition
of cytoplasmic GSK-3β increases cisplatin resistance through
activation of Wnt/β-catenin signaling in A549/DDP cells. Cancer
Lett. 336:231–239. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ren S, Liu Y, Xu W, et al: Long noncoding
RNA MALAT-1 is a new potential therapeutic target for castration
resistant prostate cancer. J Urol. 190:2278–2287. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schmidt LH, Spieker T, Koschmieder S, et
al: The long noncoding MALAT-1 RNA indicates a poor prognosis in
non-small cell lung cancer and induces migration and tumor growth.
J Thorac Oncol. 6:1984–1992. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xi YN, Xin XY and Ye HM: Effects of HMGA2
on malignant degree, invasion, metastasis, proliferation and
cellular morphology of ovarian cancer cells. Asian Pac J Trop Med.
7:289–292. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Prensner JR, Iyer MK, Sahu A, et al: The
long noncoding RNA SChLAP1 promotes aggressive prostate cancer and
antagonizes the SWI/SNF complex. Nat Genet. 45:1392–1398. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang F, Huo XS, Yuan SX, et al: Repression
of the long noncoding RNA-LET by histone deacetylase 3 contributes
to hypoxia-mediated metastasis. Mol Cell. 49:1083–1096. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chang A: Chemotherapy, chemoresistance and
the changing treatment landscape for NSCLC. Lung Cancer. 71:3–10.
2011. View Article : Google Scholar
|
|
28
|
Strathdee G: Epigenetic markers and
response to chemotherapy in cancer. Dis Markers. 23:43–49. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu Z, Sun M, Lu K, et al: The long
noncoding RNA HOTAIR contributes to cisplatin resistance of human
lung adenocarcinoma cells via downregualtion of p21 (WAF1/CIP1)
expression. PloS One. 8:e772932013. View Article : Google Scholar
|
|
30
|
Braconi C, Kogure T, Valeri N, et al:
microRNA-29 can regulate expression of the long non-coding RNA gene
MEG3 in hepatocellular cancer. Oncogene. 30:4750–4756. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Butz H and Patocs A: Pituitary
tumorigenesis: role of regulation of Wee1 kinase by microRNAs.
Tumors of the Central Nervous System. Hayat M: 10. Springer;
Dordrecht: pp. 141–150. 2013
|
|
32
|
McCubrey JA, Steelman LS, Abrams SL, et
al: Roles of the RAF/MEK/ERK and PI3K/PTEN/AKT pathways in
malignant transformation and drug resistance. Adv Enzyme Regul.
46:249–279. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bouwman P and Jonkers J: The effects of
deregulated DNA damage signalling on cancer chemotherapy response
and resistance. Nat Rev Cancer. 12:587–598. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cui J, Jiang W, Wang S, Wang L and Xie K:
Role of Wnt/β-catenin signaling in drug resistance of pancreatic
cancer. Curr Pharm Des. 18:2464–2471. 2012. View Article : Google Scholar
|
|
35
|
Yang D, Welm A and Bishop JM: Cell
division and cell survival in the absence of survivin. Proc Natl
Acad Sci USA. 101:15100–15105. 2004. View Article : Google Scholar : PubMed/NCBI
|