Introduction
Atherosclerosis is the major cause of mortality in
contemporary society (1). It is a
chronic inflammatory disease with a complicated pathogenesis.
Numerous risk factors are involved, which ultimately enhance the
adherence of circulating monocytes to vascular endothelial cells,
which are inflamed, damaged or exposed to turbulent shear-stress
(2). This monocyte-endothelial
adherence is considered to be the primary initiator of
atherosclerosis (3). Adherent
monocytes transmigrate into the arterial intima where they
propagate, mature and accumulate lipids. These intimal monocytes
transform into macrophage foam cells, a hallmark of atherosclerosis
(4). In addition, adherent
monocytes damage the vascular endothelium, causing the release of
chemoattractants and inflammatory factors (5). The inflammatory reactions are
therefore, self-reinforcing, eventually resulting in the formation
of the atherosclerotic plaque (6).
Therefore, monocyte-endothelial adhesion not only initiates the
pathological process of atherosclerosis, but enhances its
progression. Therefore, it is important to understand the mechanism
of this adhesion and the factors, which control it.
A number of factors facilitate this initial step,
including hemodynamic turbulence, secretion of inflammatory factors
and dysfunction of the endothelial cells (7). Previous studies have indicated roles
for connexin (Cx) hemichannels and gap junctions in the pathology
of atherosclerosis (8,9).
Connexins are a large family of proteins, which form
hemichannels or gap junctions to promote transmembrane and
intercellular coordination of tissue activity. Previous studies
have indicated that connexins are involved in atherogenesis
(8–12). There are three predominant
connexins expressed in endothelial cells, Cx37, Cx40 and Cx43.
Changes in their expression pattern have been assessed in cell
lines and different animal models of atherosclerosis (9). Cx37 is considered to be
atheroprotective. It was reported that Cx37 hemichannels regulate
the initiation of atherosclerosis by inhibiting autocrine
ATP-dependent modulation of monocyte-endothelial adhesion (8). Deletion of Cx40 promotes
monocyte-endothelial adhesion and accelerates atherosclerosis,
which is associated with gap junctions composed of Cx40 propagating
adenosine-evoked anti-inflammatory signals (9). In our previous study, it was
determined that the protective effects of Cx43 in cell adhesion of
monocytes may be regulated by Cx43-stimulated ATP release (11). However, the function of the
expression of Cx43 in endothelial cells remains to be
elucidated.
Cx43 is the most widespread Cx in the cardiovascular
system and is involved in normal physiology and several
cardiovascular pathologies, including atherosclerosis (11). Cx43 is upregulated in human
atheromatous plaque and in certain animal models of
atherosclerosis. It enhances monocyte intimal migration,
proliferation and apoptosis (12,13).
In the present study, the effects of Cx43 expression in human
umbilical vein endothelial cells (HUVEC) on monocyte-endothelial
adhesion were investigated.
Materials and methods
Cell lines and cell culture
HUVEC were enzymatically isolated from fresh
umbilical cord, as previously described (11). The HUVEC were cultured in human
endothelial-SFM medium (Invitrogen Life Technologies, Carlsbad, CA,
USA), containing 15% fetal bovine serum (Invitrogen Life
Technologies), 100 U/ml penicillin-streptomycin (Invitrogen Life
Technologies), 100 µg/ml heparin (Sigma-Aldrich, St. Louis,
MO, USA) and 150 µg/ml endothelial cell growth supplement
(Becton-Dickinson, Franklin Lakes, NJ, USA). For experiments, HUVEC
between passages 3 and 5 were selected. The present study conformed
to the ethical guidelines of the 1975 Declaration of Helsinki, with
the approval of the Institutional Medical Ethics Committee of the
Third Affiliated Hospital of Sun Yat-Sen University (Guangdong,
China). All donors provided written informed consent. Human U937
monocytic cells were purchased from American Type Culture
Collection (Manassas, VA, USA) and were cultured in RPMI-1640
medium (Invitrogen Life Technologies), supplemented with 15% fetal
bovine serum and 100 U/ml penicillin-streptomycin. HUVEC and U937
cells were cultured at 37°C in a 5% CO2 incubator with
90% humidity (Thermo Fisher Scientific, Waltham, MA, USA).
Adhesion assays
The U937 cells were labeled with 5 µM
calcein-acetoxymethyl ester (Invitrogen Life Technologies) for 30
min at 37°C in a 5% CO2 incubator at 90% humidity
(Thermo Fisher Scientific). The cells were subsequently washed
twice with phosphate buffered saline (PBS; Invitrogen Life
Technologies) and were resuspended in medium without serum.
Following this, the cells were seeded onto confluent monolayers of
HUVEC, which had been treated overnight with recombinant mouse
tumor necrosis factor-α (TNF-α; 20 ng/ml; PeproTech, Rocky Hill,
NJ, USA). Following incubation for 1 h at 37°C, the plates were
rinsed twice with medium without serum. The adherent U937 cells
remaining on the confluent monolayers of the HUVEC were counted
using a fluorescence microscope (IX71; Olympus, Tokyo, Japan). For
each condition, eight different visual fields (magnification, ×200)
in the middle of the dish were selected for analysis. HUVEC were
pretreated with the Cx43 channel inhibitor, oleamide (50
µM), or enhancer, retinoic acid (25 µM;
Sigma-Aldrich), for different durations prior to adhesion assays
and western blotting.
Overexpression of Cx43
Cx43 was expressed in HUVEC using a pcDNA3.1-Cx43
vector, kindly gifted by Professor Ryan Jensen and Professor Peter
M. Glazer (Departments of Therapeutic Radiology and Genetics, Yale
University School of Medicine, CT, USA). Transfection into the
HUVEC was performed using Lipofectamine 2000 (Invitrogen Life
Technologies), according to the manufacturer's instructions. The
expression was assessed 72 h after transfection by western
blotting.
Inhibition of Cx43 expression by small
interfering (si)RNA transfection
An siRNA targeting the human Cx43 gene (5′-CAG UCU
GCC UUU CGU UGUA-3′) and a non-specific control siRNA were used
(Invitrogen Life Technologies). For HUVEC, transfection with siRNA
was performed at 50% confluence in 6-well plates. A mixture of 50
nM siRNA, Lipofectamine 2000 (5 µl) and 500 µl
Opti-MEM (Invitrogen Life Technologies), were pre-incubated for 20
min at room temperature and the mixture was subsequently added to
the 6-well plates, containing 1,500 µl complete medium
without antibiotics for 6 h in a 5% CO2 incubator with
90% humidity. Following 72 h incubation, the knockdown efficiency
was assessed by western blotting.
Western blotting
Western blotting was performed, as described
previously (10). Whole-cell
lysates for western blotting were prepared by washing the cells
thrice with cell wash buffer [0.01 mol/l PBS, 0.138 mol/l NaCl,
0.02% NaN3 (pH 7.4)] followed by a 2-h incubation in lysis buffer
(Nanjing Keygen Biotech Co., Ltd., Nanjing, China) at 4°C using
0.05 ml/cm2. Protein concentrations were measured via
the bicinchoninic acid method (Nanjing Keygen Biotech Co., Ltd.).
Cell lysates (10 µg) or purified connexin preparations (10
µl) were separated by SDS-PAGE on 13% Tris-glycine mini-gels
(Invitrogen Life Technologies) and transferred onto polyvinylidene
difluoride membrane (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The membranes were blocked for 30 min at room temperature
with 5% non fat dry milk (Sigma-Aldrich) and immunoblotted using
anti-Cx43 antibody (1:3,000; mouse monoclonal Cx43 antibody raised
against human; cat. no. C8093; Sigma-Aldrich) overnight at 4°C.
Following several washes, the membranes were incubated for 1 h at
room temperature with anti-mouse horseradish peroxidase
(HRP)-conjugated secondary antibodies (1:3,000, goat polyclonal
antibody raised against mouse IgG; cat. no. M6898; Sigma-Aldrich).
The membranes were also incubated with anti-vascular cell adhesion
molecule-1 (VCAM-1; mouse monoclonal VCAM-1 antibody raised against
human; cat. no. sc-13160; Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) and anti-intercellular cell adhesion molecule-1
(ICAM-1; mouse monoclonal ICAM-1 antibody raised against human;
cat. no. sc-8439; Santa Cruz Biotechnology, Inc.) at a dilution of
1:200 overnight at 4°C and anti-mouse HRP-conjugated secondary
antibodies (1:2,000; goat polyclonal antibody raised against mouse
IgG; cat. no. M6898; Sigma-Aldrich) for 1 h at room temperature.
Anti-Cx37 (goat polyclonal Cx37 antibody raised against human; cat.
no. sc-27712; Santa Cruz Biotechnology, Inc.) and anti-Cx40
antibodies (mouse monoclonal Cx40 antibody raised against human;
cat. no. sc-365107; Santa Cruz Biotechnology, Inc.) were both used
at a dilution of 1:200. Secondary antibodies of Cx37 (1:2,000;
donkey anti-goat IgG-HRP; cat. no. sc-2020; Santa Cruz
Biotechnology, Inc.) was used at 1:1,000. Anti-mouse HRP-conjugated
secondary antibodies of Cx40 (1:2,000; goat polyclonal antibody
raised against mouse IgG; cat. no. M6898; Sigma-Aldrich) was used
at 1:1,000. Mouse monoclonal anti-β-actin antibody raised against
human (cat. no. A1978; Sigma-Aldrich) was used at a dilution of
1:10,000 and anti-mouse HRP-conjugated secondary antibodies (goat
polyclonal antibody raised against mouse IgG; cat. no. M6898;
Sigma-Aldrich) was used at 1:10,000. All primary antibodies were
incubated overnight at 4°C and secondary antibodies were incubated
for 1 h at room temperature. The protein bands were detected with
an enhanced chemiluminescence system (KGP1125; Nanjing KeyGen
Biotech. Co., Ltd.) Protein band sizes were estimated using Alpha
View software (version 2.2.14407; ProteinSimple, Santa Clara, CA,
USA).
Extracellular ATP measurements
Cellular ATP release was determined using an ATP
bioluminescent assay kit (Sigma-Aldrich), according to the
manufacturer's instructions (11).
The supernatants of the HUVEC cultures were collected on ice. A
total of 100 µl supernatant was added to 100 µl ATP
assay mix solution. The luminescence was measured using a Cary
Eclipse Fluorescence Spectrophotometer (FL0811M005; Bio/Chemi
luminescence mode; Agilent Technologies, Palo Alto, CA, USA) in a
96-well culture plate.
Statistical analysis
Statistical analysis was performed using SPSS 15.0
software (SPSS, Inc., Chicago, IL, USA). Multiple comparisons among
groups were analyzed using one-way analysis of variance, followed
by Tukey post hoc comparisons. P<0.05 was considered to indicate
a statistically significant difference.
Results
Effects of oleamide and retinoic acid
treatment on the expression of Cx43, ATP release and U937-HUVEC
adhesion in HUVEC
To investigate the role of HUVEC connexins on
monocyte-endothelial adhesion, HUVEC were treated with oleamide,
which rapidly inhibited Cx channels, including Cx43 (14), or with retinoic acid, which our
previous study demonstrated increases the expression of Cx43 in
these cells (15). The effects of
oleamide and retinoic acid treatment on U937-HUVEC adhesion, Cx43
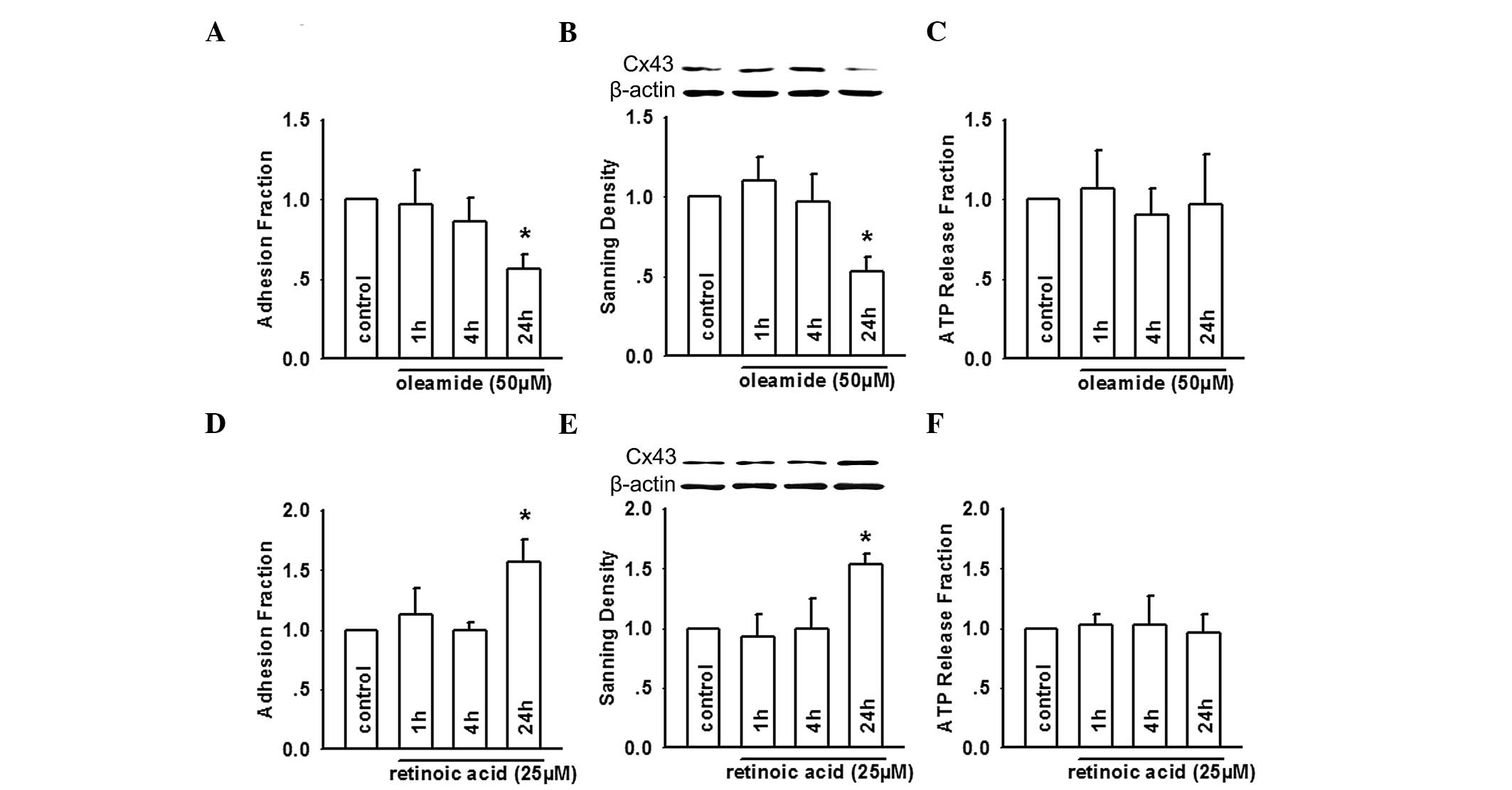
expression and ATP release were determined. As shown in Fig. 1A, oleamide (50 µM) revealed
no rapid effect on adhesion, however, did modestly attenuate
adhesion at 24 h, which correlated with a decrease in the
expression of Cx43 in HUVEC (Fig.
1B). Previous studies suggest that a rapid effect on adhesion
is always associated with ATP release, particularly from monocytes,
which inhibits monocyte-endothelial adhesion, and the depression of
ATP release may thereby enhance atherosclerotic progression
(11,16). These studies suggested that ATP
release can markedly affect cell adhesion. Therefore, the present
study detected ATP release from HUVEC exposed to oleamide. No
effects were observed on ATP release (Fig. 1C). This suggested that in HUVEC,
Cx43 channel activity was not involved in adhesion, however, a
decrease in Cx43 expression may attenuate it. To further
characterize the association between the expression of Cx43 and
U937-HUVEC cell adhesion, the effects of retinoic acid, a commonly
used potentiator of Cx expression and function was assessed.
Treatment of HUVEC with retinoic acid (25 µM) induced
increased expression of Cx43 at 24 h, however, not at shorter
durations, which correlated with increased adhesion (Fig. 1D and E). ATP release from HUVEC
exposed to retinoic acid remained unchanged (Fig. 1F). These findings suggested that
U937-HUVEC cell adhesion was positively modulated by the expression
of Cx43 in HUVEC, and was essentially unaffected by altering of the
Cx43 function.
Direct modulation of the expression of
Cx43 in HUVEC modulates U937-HUVEC adhesion
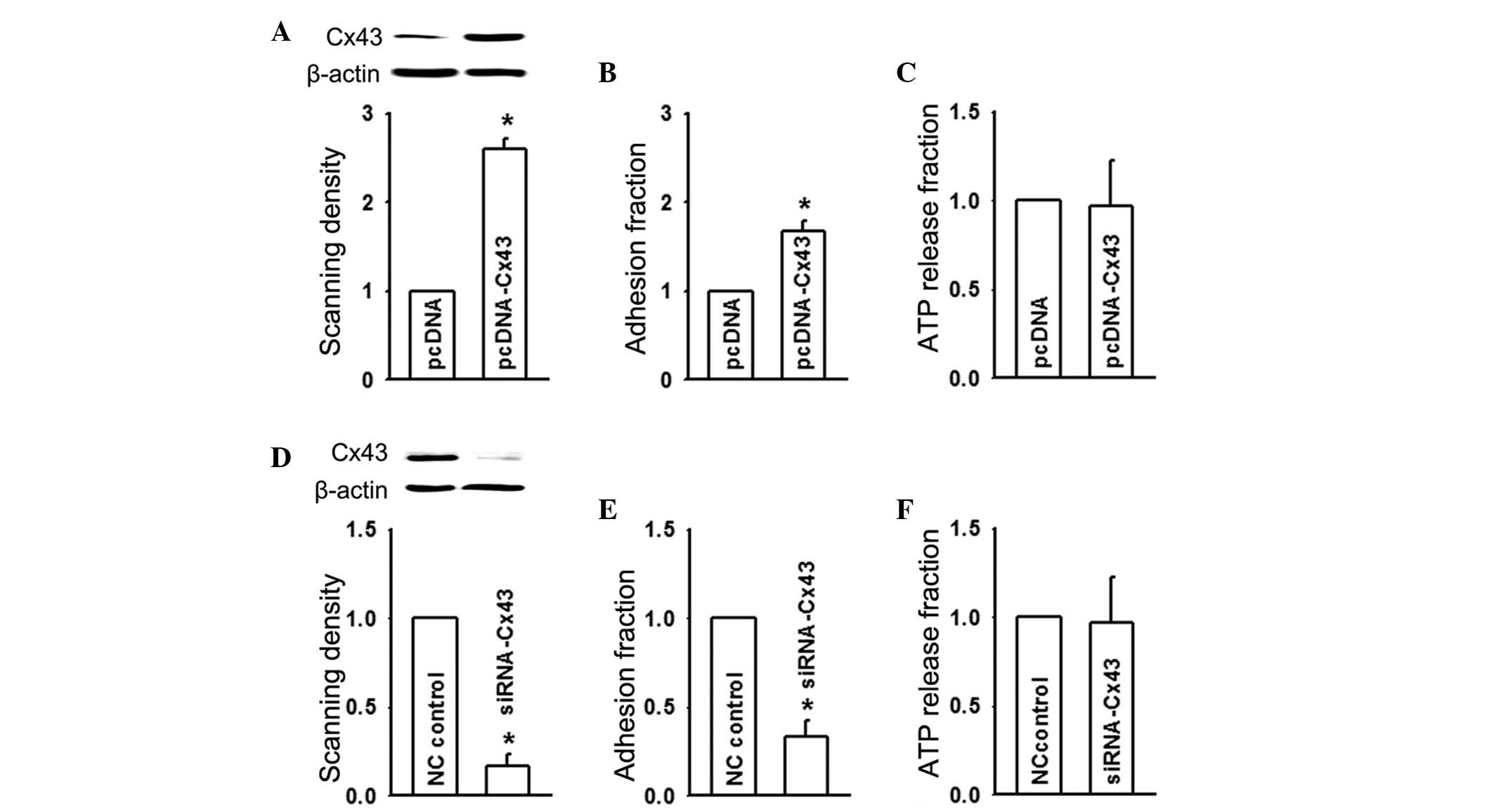
To more directly and specifically establish the role
of Cx43 in U937-HUVEC adhesion, the expression of Cx43 was
specifically altered by two methods: Over-expression by
transfection with pcDNA-Cx43 and knock-down using siRNA-Cx43. As
shown in Fig. 2A and B, Cx43
expression in HUVEC was enhanced nearly two-fold following
pcDNA-Cx43-transfection, and this markedly increased adhesion to
monocytes. However, ATP release from HUVEC remained unaffected by
pcDNA-Cx43-transfection, which correlated with HUVEC exposed to the
Cx43 potentiator, retinoic acid (Figs.
1C and 2C). Fig. 2D and E demonstrated that the
expression of Cx43 was downregulated by siRNA-Cx43 and this led to
a decrease in adhesion, however, ATP release from HUVEC remained
unaffected (Fig. 2F). These
results, in combination with those in Fig. 1, demonstrated that even though Cx43
channel function in HUVEC revealed no effect on adhesion, the
expression of Cx43 in HUVEC positively regulated it.
Direct modulation of the expression of
Cx43 regulates cell adhesion proteins, VCAM-1 and ICAM-1, in
HUVEC
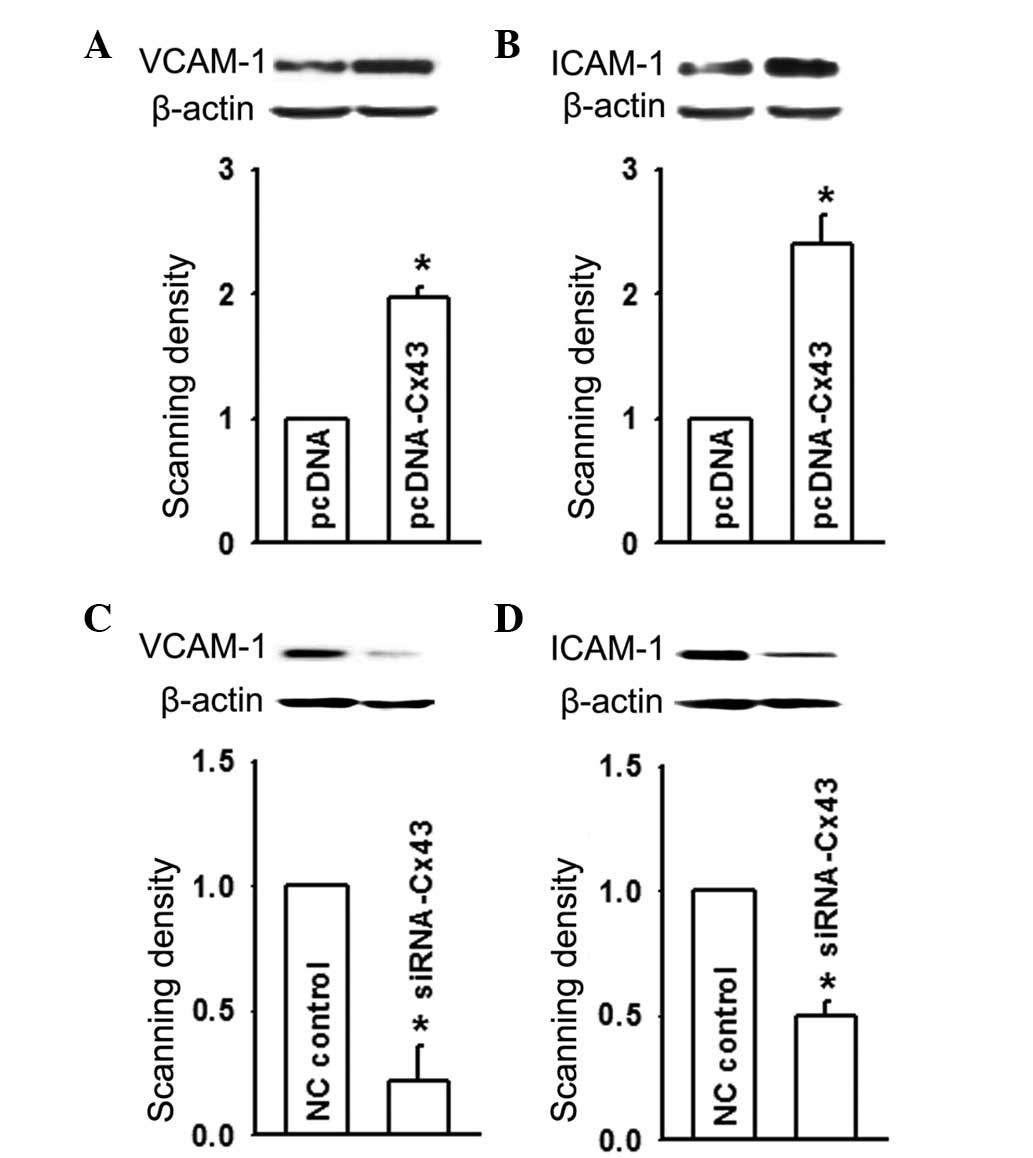
The cell adhesion proteins, VCAM-1 and ICAM-1,
belong to the immunoglobulin superfamily, whose endothelial
expression is closely associated with the development of
atherosclerosis (17). Since the
results shown in Figs. 1 and
2 indicated that the expression of
Cx43 in HUVEC positively regulated U937-HUVEC cell adhesion, the
present study next determined whether alteration of the expression
of Cx43 HUVEC affected the expression levels of VCAM-1 and ICAM-1.
The results demonstrated that when Cx43 expression was increased by
pcDNA-Cx43-transfection, VCAM-1 and ICAM-1 expression were markedly
upregulated (Fig. 3A and B). By
contrast, when the expression of Cx43 was reduced by siRNA-Cx43 in
HUVEC, the expression levels of VCAM-1 ICAM-1 declined as the
expression of Cx43 decreased (Fig. 3C
and D). This suggested that the expression of Cx43 in HUVEC
promoted cell adhesion by increasing the expression of VCAM-1 and
ICAM-1.
Altering the expression of Cx43 in HUVEC
reveals no effect on the expression levels of Cx37 or Cx40
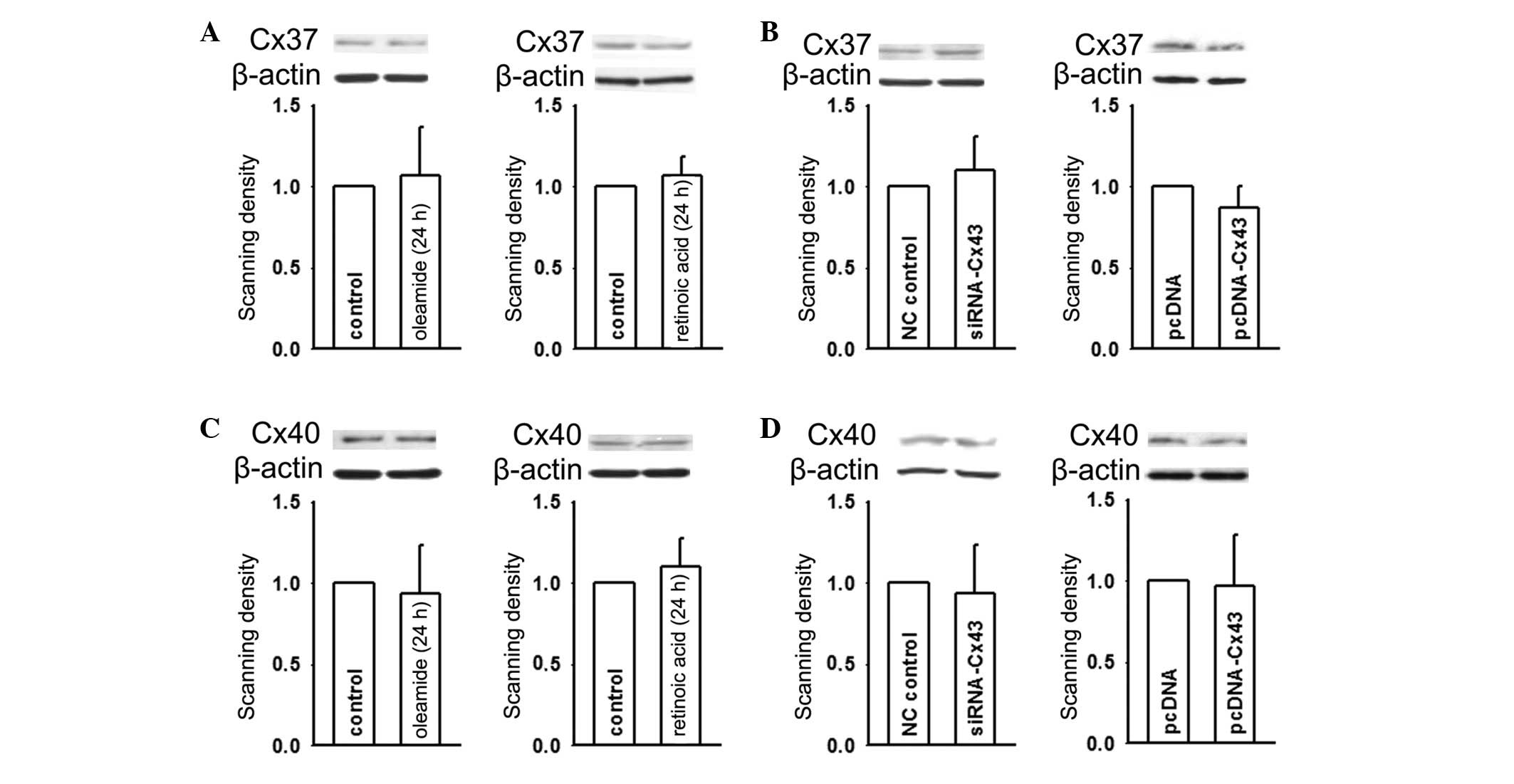
Previous studies demonstrated three different types
of connexins expressed in HUVEC, Cx37, Cx40 and Cx43 (18–20).
It was previously demonstrated that Cx37 promotes ATP release,
which reduced monocyte adhesion to endothelial cells, thereby
interfering with atherosclerotic development (8,9). It
was also demonstrated that Cx40-mediated gap junction communication
contributed to the attenuation of leukocyte adhesion to the
endothelium (8,9). In order to exclude the effects of
Cx37 and Cx40 on cell adhesion, the present study assessed the
expression levels of Cx37 and Cx40 when Cx43 expression was
altered. Fig. 1 demonstrated that
at 24 h, the expression of Cx43 was markedly changed along with
cell adhesion when HUVEC were exposed to oleamide or retinoic acid.
However, the expression of Cx37 revealed no change under identical
conditions (Fig. 4A). Notably,
when Cx43 in HUVEC was overexpressed by transfection with
pcDNA-Cx43 or knocked down with siRNA-Cx43, Cx37 expression
remained unchanged (Fig. 4B). An
identical result was obtained for Cx40 (Fig. 4C and D). These results indicated
that Cx43 expression, which regulated cell adhesion, was
independent of the expression levels of Cx37 or Cx40 in HUVEC.
Discussion
The present study investigated the effect of Cx43
function and expression in vascular endothelial cells on
monocyte-endothelial adhesion, the initial step in atherosclerotic
development. It was confirmed that in HUVEC, upregulation of the
expression of Cx43 enhanced cell adhesion and downregulation
markedly decreased cell adhesion. Notably, two different types of
crucial cell adhesion protein, VCAM-1 and ICAM-1, were affected by
changes in the expression of Cx43, which mediated the recruitment
of circulating monocytes onto endothelial cells. A previous study
demonstrated that ATP release is important in monocyte-endothelial
adhesion, particularly from Cx37 channels, which contributed to a
rapid decrease in adhesion (8).
However, the possible contribution of Cx43 in HUVEC remains to be
elucidated. It has been well documented that Cx43 is impermeable to
ATP in monocytes, which may modulate monocyte-endothelial adhesion
(11), however, in the present
study, it was revealed that ATP release from HUVEC revealed no
effect on cell adhesion and that monocyte-endothelial adhesion may
be regulated by changes in the expression of Cx43 modulating the
cell adhesion proteins, VCAM-1 and ICAM-1.
Downregulation of Cx43 had been demonstrated to
down-regulate the expression of VCAM-1 in other systems (21). The specific mechanism remains to be
elucidated, however, there were several likely pathways. One
pathway involves interactions between the carboxyl-terminal domain
of Cx43 and elements of various cellular signaling pathways,
including cytoskeletal components, Src, PKA and PKC, which have
downstream effects on cell adhesion molecules (22). In particular, it is known that the
expression levels of VCAM-1 and ICAM-1 are closely associated with
the activation of NF-κB and PKC, and that Cx43 amplifies the
intercellular signaling, which activates NF-κB (23).
Several previous studies revealed that Cx
hemichannels release cytosolic compounds, including ATP (8). ATP release through hemichannels may
be dynamically regulated by several factors, including changes in
intracellular Ca2+, ischaemic stress, hypoxia and,
important in the context of atherosclerosis, cytokines (24,25).
Atherosclerosis is an inflammatory disease in which the release of
cytokines is prominent. Cytokines, including TNF-α, interleukin,
IFN-γ and TGF-β, inhibit the activity of Cx43 channels (26,27).
Therefore, cytokine release in atherosclerotic progression may
attenuate ATP release from monocytes, enhancing adhesion in a
positive-feedback inflammatory cycle. Although Cx43 was expressed
by U937 and HUVEC, it was notable that Cx43 hemichannel activity
appeared to be differentially regulated in the two cell types. In
our previous study, monocyte-endothelial adhesion was revealed to
be modulated by Cx43 through ATP release in U937 cells (11). However, in the present study, Cx43
in HUVEC did not exert this function. This difference may arise
from interactions of the carboxyl-terminal domain of Cx43 with
different regulatory binding partners and signaling pathways in the
two cell types (28).
To date, three different types of connexins, Cx37,
Cx40 and Cx43, have been revealed to be expressed in HUVEC, which
may influence monocyte-endothelial adhesion and protect against
atherosclerosis through various mechanisms (8,9). In
the present study, four distinct methods were used to assess the
effects of Cx43 on monocyte-endothelial adhesion: Inhibition using
oleamide; enhancement with retinoic acid; overexpression of Cx43 by
transfection with pcDNA-Cx43 and knock-down of Cx43 expression with
siRNA-Cx43. The results revealed that Cx43 expressed in HUVEC
influenced monocyte-endothelial adhesion by modulating the
expression levels of cell adhesion proteins, VCAM-1 and ICAM-1. The
upregulation of Cx43 expression enhanced monocyte-endothelial
adhesion and downregulation of Cx43 markedly decreased it. In order
to confirm the specific function of Cx43 on monocyte-endothelial
adhesion, whether modulation of Cx43 was dependent on Cx37 or Cx40
was investigated. Fig. 4
illustrated that neither Cx37 nor Cx40 were affected by the altered
expression of Cx43, which indicated that the effects of Cx43
expression in HUVEC on monocyte-endothelial adhesion were specific
and distinctive, independent of Cx37 or Cx40.
The role of the expression and/or function of Cx43
in the pathogenesis of atherosclerosis were complex, suggesting
various roles in different phases of atherosclerosis. For example,
Cx43 modulated monocyte migration to the arterial intima through
the interaction between the carboxyl-terminal tails of Cx43 and
microtubules (29,30). These subsequently became foam cells
in arterial intima and this process was also regulated by the
expression of Cx43 (12). The
expression of Cx43 was always markedly increased at the curved
areas or the branch points of large arteries and the oscillatory
shear stress in those locations, which contributed to
atherosclerotic plaques formation (31). Reduced endothelial expression of
Cx43 was associated with plaque stability (13).
The present study focused on the initiating step of
atherosclerosis, monocyte-endothelial adhesion. However, the
present study differed compared with our previous investigation,
which focused on the expression of Cx43 in monocytes and the
effects of endothelial cells expressing Cx43 on adhesion. The
present findings, along with our previous conclusion, suggested
that altered expression of Cx43 in monocytes and endothelial cells
may regulate cell adhesion. Notably, it was determined that Cx43
channel-stimulated ATP release in monocytes can affect rapid
adhesion, however, the expression of Cx43 in HUVEC did not exert
this function. The differences described here offer distinct basis
for targeted intervention.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (nos. 81170449 and 81401628), the key
project of Natural Science Foundation of Guangdong Province, China
(no. S2011020002780) and the Medical Research Foundation of
Guangdong Province (no. B2014141).
References
|
1
|
Burnier L, Fontana P, Angelillo-Scherrer A
and Kwak BR: Intercellular communication in atherosclerosis.
Physiology (Bethesda). 24:36–44. 2009. View Article : Google Scholar
|
|
2
|
Boon RA and Horrevoets AJ: Key
transcriptional regulators of the vasoprotective effects of shear
stress. Hamostaseologie. 29:39–43. 2009.PubMed/NCBI
|
|
3
|
Binder CJ, Chang MK, Shaw PX, Miller YI,
Hartvigsen K, Dewan A and Witztum JL: Innate and acquired immunity
in atherogenesis. Nat Med. 8:1218–1226. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sumiyoshi A: Pathological processes of
coronary atherosclerosis. Nihon Rinsho. (Suppl 52): S85–S89.
1994.In Japanese.
|
|
5
|
Rebrov AP and Voskoboi IV: Effect of
inflammatory and infectious factors in the development of
atherosclerosis. Ter Arkh. 76:78–82. 2004.In Russian.
|
|
6
|
Insull W Jr: The pathology of
atherosclerosis: Plaque development and plaque responses to medical
treatment. Am J Med. 122(Suppl 1): S3–S14. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tzoulaki I, Murray GD, Price JF, Smith FB,
Lee AJ, Rumley A, Lowe GD and Fowkes FG: Hemostatic factors,
inflammatory markers and progressive peripheral atherosclerosis:
The Edinburgh Artery Study. Am J Epidemiol. 163:334–341. 2006.
View Article : Google Scholar
|
|
8
|
Wong CW, Christen T, Roth I,
Chadjichristos CE, Derouette JP, Foglia BF, Chanson M, Goodenough
DA and Kwak BR: Connexin37 protects against atherosclerosis by
regulating monocyte adhesion. Nat Med. 12:950–954. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chadjichristos CE, Scheckenbach KE, van
Veen TA, Richani Sarieddine MZ, de Wit C, Yang Z, Roth I, Bacchetta
M, Viswambharan H, Foglia B, et al: Endothelial-specific deletion
of connexin40 promotes atherosclerosis by increasing CD73-dependent
leukocyte adhesion. Circulation. 121:123–131. 2010. View Article : Google Scholar
|
|
10
|
Luo C, Yuan D, Li X, Yao W, Luo G, Chi X,
Li H, Irwin MG, Xia Z and Hei Z: Propofol attenuated acute kidney
injury after orthotopic liver transplantation via inhibiting gap
junction composed of connexin 32. Anesthesiology. 122:72–86. 2015.
View Article : Google Scholar
|
|
11
|
Lee ES, Van Spyk EN, Chun KC, Pitts RL, Wu
MH and Yuan SY: Monocytic adhesion molecule expression and
monocyte-endothelial cell dysfunction are increased in patients
with peripheral vascular disease versus patients with abdominal
aortic aneurysms. J Surg Res. 177:373–381. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Derouette JP, Wong C, Burnier L, Morel S,
Sutter E, Galan K, Brisset AC, Roth I, Chadjichristos CE and Kwak
BR: Molecular role of Cx37 in advanced atherosclerosis: A
micro-array study. Atherosclerosis. 206:69–76. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wong CW, Burger F, Pelli G, Mach F and
Kwak BR: Dual benefit of reduced Cx43 on atherosclerosis in LDL
receptor-deficient mice. Cell Commun Adhes. 10:395–400. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ransjö M, Sahli J and Lie A: Expression of
connexin43 mRNA in microisolated murine osteoclasts and regulation
of bone resorption in vitro by gap junction inhibitors. Biochem
Biophys Res Commun. 303:1179–1185. 2003. View Article : Google Scholar
|
|
15
|
Tanmahasamut P and Sidell N: Up-regulation
of gap junctional intercellular communication and connexin43
expression by retinoic acid in human endometrial stromal cells. J
Clin Endocrinol Metab. 90:4151–4156. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Eltzschig HK, Eckle T, Mager A, Küper N,
Karcher C, Weissmüller T, Boengler K, Schulz R, Robson SC and
Colgan SP: ATP release from activated neutrophils occurs via
connexin43 and modulates adenosine-dependent endothelial cell
function. Circ Res. 99:1100–1108. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fotis L, Agrogiannis G, Vlachos IS,
Pantopoulou A, Margoni A, Kostaki M, Verikokos C, Tzivras D,
Mikhailidis DP and Perrea D: Intercellular adhesion molecule
(ICAM)-1 and vascular cell adhesion molecule (VCAM)-1 at the early
stages of atherosclerosis in a rat model. In Vivo. 26:243–250.
2012.PubMed/NCBI
|
|
18
|
Dhein S, Gaertner C, Georgieff C, Salameh
A, Schlegel F and Mohr FW: Effects of isoprenaline on endothelial
connexins and angiogenesis in a human endothelial cell culture
system. Naunyn Schmiedebergs Arch Pharmacol. 388:101–108. 2015.
View Article : Google Scholar
|
|
19
|
Godecke S, Roderigo C, Rose CR, Rauch BH,
Godecke A and Schrader J: Thrombin-induced ATP release from human
umbilical vein endothelial cells. Am J Physiol Cell Physiol.
302:C915–C923. 2012. View Article : Google Scholar
|
|
20
|
Haussig S, Schubert A, Mohr FW and Dhein
S: Sub-chronic nicotine exposure induces intercellular
communication failure and differential down-regulation of connexins
in cultured human endothelial cells. Atherosclerosis. 196:210–218.
2008. View Article : Google Scholar
|
|
21
|
Toubas J, Beck S, Pageaud AL, Huby AC,
Mael-Ainin M, Dussaule JC, Chatziantoniou C and Chadjichristos CE:
Alteration of connexin expression is an early signal for chronic
kidney disease. Am J Physiol Renal Physiol. 301:F24–F32. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Giepmans BN: Gap junctions and
connexin-interacting proteins. Cardiovasc Res. 62:233–245. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Martin FJ and Prince AS: TLR2 regulates
gap junction intercellular communication in airway cells. J
Immunol. 180:4986–4993. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Faigle M, Seessle J, Zug S, El Kasmi KC
and Eltzschig HK: ATP release from vascular endothelia occurs
across Cx43 hemi-channels and is attenuated during hypoxia. PLoS
One. 3:e28012008. View Article : Google Scholar
|
|
25
|
Anselmi F, Hernandez VH, Crispino G,
Seydel A, Ortolano S, Roper SD, Kessaris N, Richardson W, Rickheit
G, Filippov MA, et al: ATP release through connexin hemichannels
and gap junction transfer of second messengers propagate
Ca2+ signals across the inner ear. Proc Natl Acad Sci
USA. 105:18770–18775. 2008. View Article : Google Scholar
|
|
26
|
Lim MC, Maubach G and Zhuo L: TGF-beta1
down-regulates connexin43 expression and gap junction intercellular
communication in rat hepatic stellate cells. Eur J Cell Biol.
88:719–730. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kimura K and Nishida T: Role of the
ubiquitin-proteasome pathway in downregulation of the gap-junction
protein Connexin43 by TNF-{alpha} in human corneal fibroblasts.
Invest Ophthalmol Vis Sci. 51:1943–1947. 2010. View Article : Google Scholar
|
|
28
|
Laird DW: The gap junction proteome and
its relationship to disease. Trends Cell Biol. 20:92–101. 2010.
View Article : Google Scholar
|
|
29
|
Rhee DY, Zhao XQ, Francis RJ, Huang GY,
Mably JD and Lo CW: Connexin43 regulates epicardial cell polarity
and migration in coronary vascular development. Development.
136:3185–3193. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Behrens J, Kameritsch P, Wallner S, Pohl U
and Pogoda K: The carboxyl tail of Cx43 augments p38 mediated cell
migration in a gap junction-independent manner. Eur J Cell Biol.
89:828–838. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Inai T, Mancuso MR, McDonald DM, Kobayashi
J, Nakamura K and Shibata Y: Shear stress-induced upregulation of
connexin43 expression in endothelial cells on upstream surfaces of
rat cardiac valves. Histochem Cell Biol. 122:477–483. 2004.
View Article : Google Scholar : PubMed/NCBI
|