Introduction
Lung cancer is the leading cause of
cancer-associated mortality worldwide. It was estimated that
226,160 novel cases and 160,340 mortalities from cancer occurred in
the United States in 2012 (1).
Non-small cell lung cancer (NSCLC) accounts for nearly 80–85% of
all lung cancer types. Approximately 40–50% of patients with NSCLC
present at an advanced stage, with metastatic or locally advanced
disease (2). Despite all recent
improvements in chemotherapy strategies and surgeries, the
five-year survival rate has remained <15% due to the emergence
of serious adverse effects, cancer recurrence and resistance to
conventional chemotherapy. Furthermore, among these patients with
tumor recurrence, >70% present with distant metastasis (3). Cancer cell invasion and migration are
critical processes determining the lethality of a malignancy, and
>90% of cancer mortalities have been attributed to the
metastatic spread of the disease rather than to the primary tumor.
Tumor metastasis is the main obstacle on the treatment of lung
cancer. Therefore, novel targeted therapeutic agents with high
efficacy and low toxicity for inhibiting metastasis are urgently
required.
Curcuminoids are the major active components of
turmeric with significant derivatives including curcumin (Cur),
demethoxycurcumin (DMC) and bisdemethoxycurcumin (BDMC). Curcumin
possesses a wide variety of biological activities, including
anti-proliferative, anti-oxidant, anti-inflammatory,
anti-angiogenic and anti-tumor effects (4,5). It
has been shown that curcumin exerts marked anti-cancer effects
through modulating the expression of a variety of molecules
involved in cancer progression (6). Curcumin has been demonstrated to
suppress lung cancer cell invasion and metastasis through multiple
signaling pathways, including DnaJ-like heat shock protein 40
(HLJ1) (7), matrix
metalloproteinase-2, metal-loproteinase-9, vascular endothelial
growth factor (8) and Janus
kinase-signal transducer and activator of transcription 3 signaling
pathway (9). However, the
stability of natural curcumin is poor and it can be easily degraded
in vivo. Therefore, it is necessary to use other stable
curcuminoids. BDMC is thought to be more stable than the other two
curcuminoids (10). Previous
studies have shown that BDMC has the highest anti-metastatic
potency in HT1080 human fibrosacroma among the three curcuminoids
(11). However, the underlying
molecular mechanisms of BDMC on tumor invasion and migration have
largely remained to be identified.
Autophagy is known to be a double-edged sword in
tumorigenesis and cancer development. Manipulation of autophagy has
been reported to have the potential to enhance the anti-cancer
effects of chemotherapeutics in the clinic (12). It has been reported that CLEFMA, a
synthetic analog of curcumin, induced autophagy in lung
adenocarcinoma H441 cells (13).
Furthermore, curcumin induced autophagy in A549 cells (14). However, little is known about the
association between autophagy induced by curcumin and the migratory
and invasive potential of NSCLC cells.
The present study investigated the effects of BDMC
on the proliferation, migration and invasion of 95D cells and
explored the underlying molecular mechanisms by detecting vimentin
and E-cadherin expression. In addition, the role of autophagy in
the invasion and migration of 95D cells was investigated.
Materials and methods
Reagents and antibodies
BDMC was purchased from Sigma-Aldrich (St. Louis,
MO, USA). BDMC (>98% pure) was dissolved in dimethyl sulfoxide
(DMSO; Sigma-Aldrich) to prepare a 100-mM stock solution, which was
diluted with medium to the required concentrations. The final
concentration of DMSO was not more than 0.01%. Polyclonal
anti-GAPDH antibody (cat. no. G9545) was purchased from
Sigma-Aldrich and rabbit polyclonal antibody against vimentin (cat.
no. sc-5565) was procured from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). Rabbit monolconal antibody against E-cadherin
(cat. no. 3195) was from Cell Signaling Technology (Beverly, MA,
USA). Goat anti-rabbit immunoglobulin G horseradish
peroxidase-conjugated secondary antibody (cat. no. ZDR-5306) was
from Zhongshan Golden Bridge Biotechnology (Beijing, China).
Cell lines, cell culture and
transfection
The highly metastatic large-cell lung cancer 95D
cell line was obtained from the Chinese Academy of Science Cell
Bank (Shanghai, China). Cells were cultured in RPMI 1640 medium (GE
Healthcare Life Sciences, Logan, UT, USA) supplemented with 10%
(v/v) fetal bovine serum (FBS; Gibco-BRL, Invitrogen Life
Technologies, Carlsbad, CA, USA) in a humidified atmosphere at 37°C
with 5% CO2. Transient transfection was performed with
green fluorescence protein-Beclin 1 plasmid using Lipofectamine
2000 (Invitrogen Life Technologies) according to the manufacturer's
instructions.
Cytotoxicity and cell proliferation
assay
The cytotoxicity and proliferation of 95D cells
after exposure to BDMC was measured using the cell counting kit 8
(CCK-8) assay kit (Beyotime Institute of Biotechnology, Haimen,
China). Briefly, 4×103 cells in the logarithmic growth
phase suspended in 100 µl complete medium were seeded into
each well of a 96-well flat-bottomed plate (Nest Biotech Co. Ltd.,
Wuxi, China) and incubated at 37°C in a humidified
5%-CO2 atmosphere overnight. The cells were then treated
with various concentrations of BDMC (0, 5, 10, 20, 40 or 80
µM) for 24 h. Subsequently, 10 µl CCK8 reagent was
added to each well containing 100 µl of culture medium,
followed by incubation for 1 h at 37°C. The optical density (OD) at
450 nm was measured using a microplate spectrophotometer (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). All of the experiments were
performed three times.
In vitro invasion assay
A total of 1×105 cells 95D cells in the
logarithmic growth phase in 0.5 ml serum-free RPMI 1640 medium were
seeded into 8 µm-pore polycarbonate membrane chamber inserts
in a Transwell plate (Corning-Costar, Corning, NY, USA) coated with
Matrigel (BD Biosciences, Franklin Lakes, NJ, USA). 600 µl
RPMI 1640 containing 20% FBS was added to the lower chamber. After
the cells were incubated for 12–24 h, cells on the upper surface of
the insert were removed by wiping with a cotton swab. Cells that
transgressed through the membrane to the bottom surface of the
insert were fixed in 100% methanol for 10 min, stained in 0.5%
crystal violet (Sigma-Aldrich) for 10 min, rinsed with
phosphate-buffered saline and then subjected to microscopic
inspection (magnification, x100; Olympus CX22, Olympus Corporation,
Tokyo, Japan). The invasion was quantified by counting five fields
per membrane. Values are and expressed as the average of three
independent experiments.
Cell motility assay
Cell migration assays were performed using Boyden
chambers containing polycarbonate membrane inserts (8 µm
pore size; Corning Costar) Briefly, 1×104 cells in 0.1
ml serum-free RPMI-1640 medium were seeded into the upper chamber.
600 µl RPMI-1640 containing 20% FBS was added to the lower
chamber as a chemoattractant. After the cells were incubated for 6,
12 or 24 h at 37°C in a 5%-CO2 incubator, the number of
cells that had migrated to the lower surface was counted. The
experiments were repeated a minimum of three times.
RNA interference of Beclin 1
95D cells were seeded in 24-well plates
(1.5×105 cells/well) and incubated overnight. A negative
control random small interfering RNA (NC-siRNA) and
Beclin1-targeted siRNA (1 µg/well) were purchased from
Genepharma (Shanghai, China). NC-siRNA and Beclin1-siRNA were
transfected using Lipofectamine 2000 according to the
manufacturer's instructions.
Western blot analysis
To determine the levels of protein expression,
whole-cell lysate extracts of BDMC-treated cells (0, 1, 5 and 10
µM for 24 h and 10 µM for 0, 6, 12 and 24 h) were
prepared and separated by 10% SDS-PAGE. After electrophoresis,
proteins were electroblotted onto a polyvinylidene difluoride
membrane (Millipore, Billerica, MA, USA), which was then incubated
with the primary antibodies (anti-GAPDH, 1:500; anti-E-cadherin,
1:1,000, anti-vimentin, 1:500). Subsequently, the blots were
incubated with the secondary antibody (1:2,000 dilution) for 1 h at
room temperature. Blots were visualized using an enhanced
chemiluminescence western blotting system (BeyoECL Plus kit;
Beyotime Institute of Biotechnology). The relative amount of
protein on the blots was determined by densitometry using Image J
software 1.49 (National Institutes of Health, Bethesda, MD,
USA).
Statistical analysis
All experiments were performed in triplicate. Values
are expressed as the mean ± standard deviation. The Student's
t test was used for paired comparisons, and the one-way
analysis of variance test was used for multiple comparisons. SPSS
17.0 (SPSS, Inc., Chicago, IL, USA) was used to analyze the data.
P<0.05 was considered to indicate a statistically significant
difference.
Results
BDMC inhibits the proliferation of 95D
cells
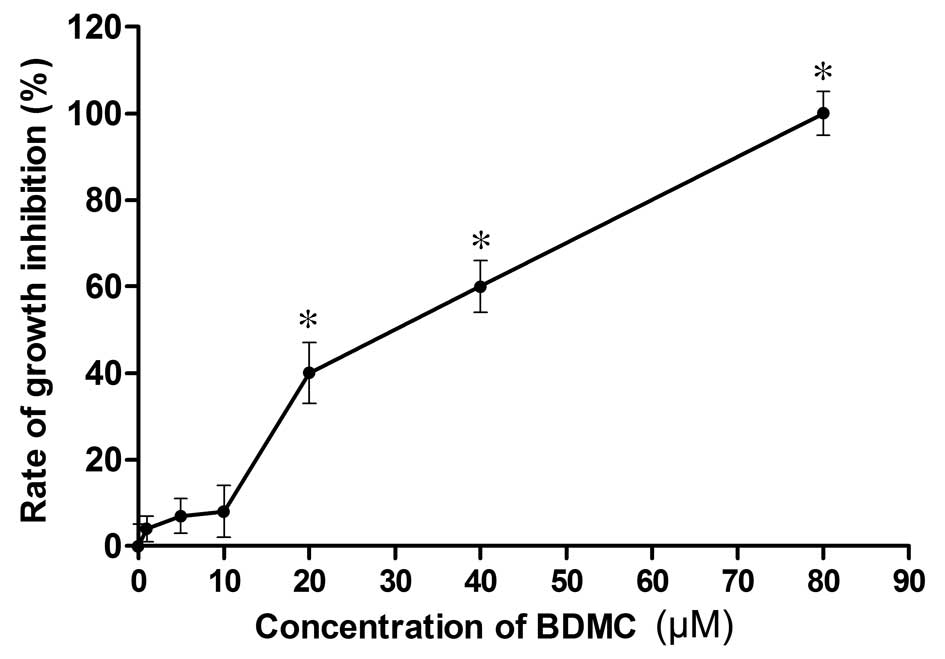
In order to investigate whether BDMC inhibits
95D-cell growth, the cells were incubated with increasing
concentrations of BDMC (0, 1, 5, 10, 20, 40 or 80 µM) for 24
h and subjected to the CCK8 assay. As shown in Fig. 1, the viability of 95D cells treated
with BDMC was not significantly reduced compared with that of the
untreated control cells at BDMC concentrations of up to 10
µM, while the proliferation was significantly inhibited by
BDMC at ≥20 µM. As treatment with BDMC at concentrations of
0–10 µM for 24 h had no cytotoxic effect on the highly
metastatic 95D cells, this concentration range was used in the
subsequent experiments.
BDMC suppresses 95D-cell migration and
invasion
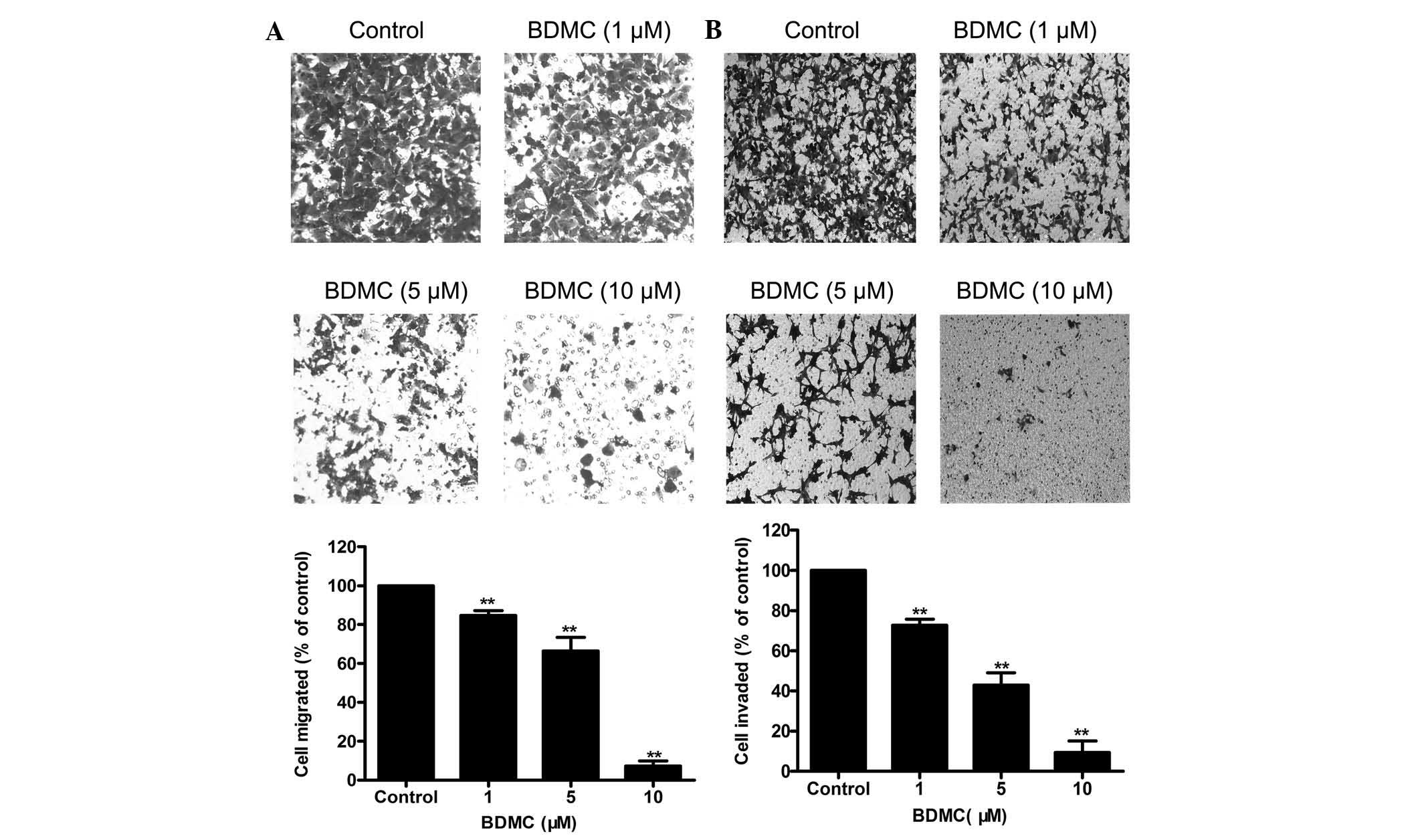
The present study examined the effects of
sub-cytotoxic concentrations of BDMC on the invasive and migratory
potential of highly invasive 95D cells. A Boyden chamber assay
showed that BDMC (1–10 µM) reduced the migratory ability of
95D cells in a dose-dependent manner (Fig. 2A). Similarly, the Transwell
invasion assay showed that BDMC (1–10 µM) decreased the
invasive ability of 95D cells in a dose-dependent manner (Fig. 2B). These results suggested that
BDMC at 1–10 µM significantly suppressed the invasion and
migration but not the viability of 95D cells.
Autophagy is involved in BDMC-mediated
suppression of 95D-cell invasion and migration
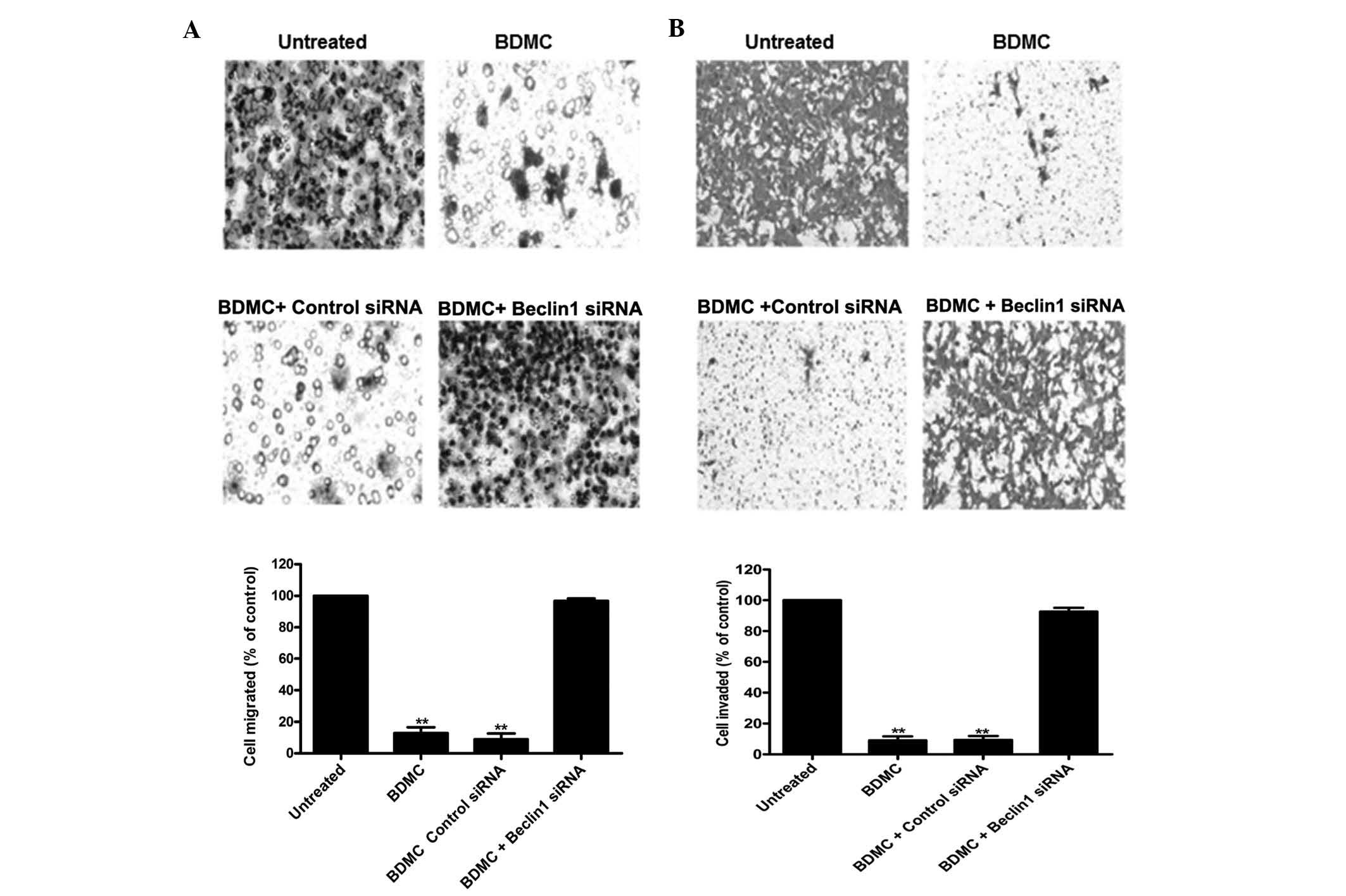
At present, autophagy is an important field of
cancer research. The effects of autophagy in the tumor are likely
to vary depending on the context. Based on the positive regulatory
effects of curcumin on autophagy, the present study hypothesized
that BDMC suppressed 95D-cell invasion and migration via
upregulating autophagy. Accordingly, a decrease in autophagy is
expected to reverse the effects of BDMC and facilitate invasion and
migration. To test this hypothesis, 95D cells were transfected with
Beclin 1-siRNA in order to interfere with the formation of
autophagosomes (15). Beclin 1 has
an important role in the formation of autophagosomes. As shown in
(Fig. 3), inhibition of autophagy
by Beclin 1 knockdown attenuated the inhibitory effects of BDMC on
95D-cell migration and invasion. This result suggested that
BDMC-mediated suppression of 95D-cell invasion and migration
proceeds via the autophagic pathway.
BDMC inhibits 95D-cell migration and
invasion by modulating the expression of E-cadherin and
vimentin
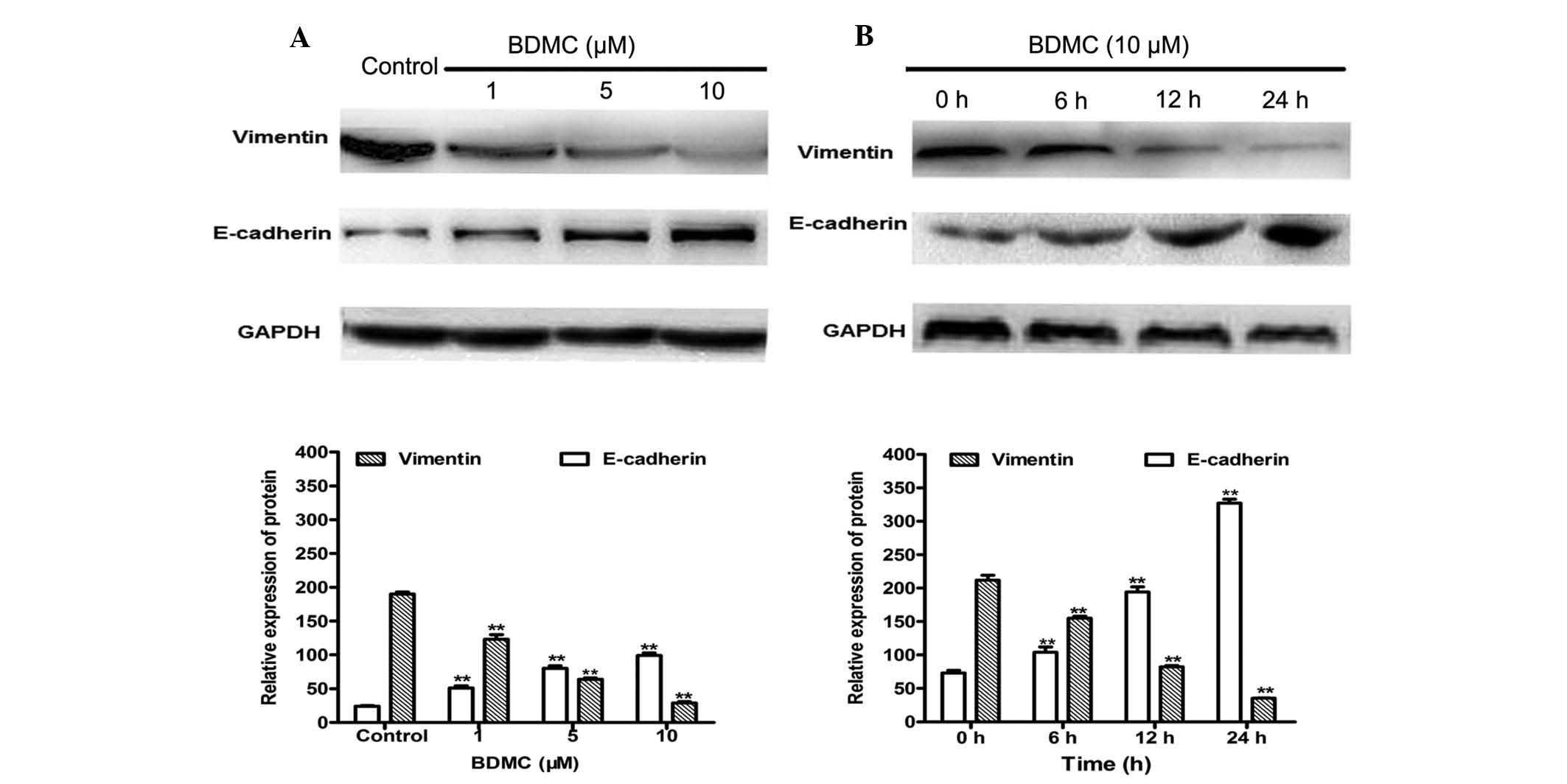
To elucidate the possible underlying mechanisms of
anti-metastatic activities of BDMC on 95D cells, its effect on
vimentin and E-cadherin expression were assessed by western blot
analysis. As shown in Fig. 4, BDMC
treatment significantly decreased the expression of vimentin in a
dose- and time-dependent manner. Treatment with 1, 5 or 10
µM BDMC for 24 h decreased the protein expression of
vimentin by 35.2, 66.3 and 84.7%, respectively, compared with that
in the control group. In addition, 10 µM BDMC treatment for
6, 12 and 24 h decreased vimentin protein expression by 26.7, 61.3
and 83.3%, respectively. Treatment with 1, 5 and 10 µM BDMC
for 24 h caused a 2.1-, 3.3-, 4.1-fold increase in E-cadherin
expression, respectively. BDMC treatment at 10 µM for 6, 12
or 24 h caused 1.4-, 2.6- and 4.5-fold increases in E-cadherin
protein expression, respectively. These findings suggested that
downregulation of vimentin and upregulation of E-cadherin may be
involved in the inhibition of invasion and migration of 95D cells
by BDMC.
Discussion
Accumulating evidence proved that curcumin has a
potent anti-cancer effects in vitro and in vivo on a
variety of cancer cell types, including breast cancer (16), prostate cancer (17), melanoma cells (18), ovarian carcinoma (19), pancreatic cancer (20), leukemia (21), head and neck cancer (22) and Burkitt's lymphoma (23). The anti-cancer activities of
curcumin are associated with multiple biological pathways involved
in tumor growth, oncogene expression, cell cycle regulation,
apoptosis, tumorigenesis and metastasis. Although multiple
anti-cancer mechanisms of curcumin have been elucidated, the
underlying mechanisms by which curcumin inhibits tumor metastasis
signaling have remained to be identified. Furthermore, the
inhibitory effects of BDMC on the invasion and migration of 95D
cells have not yet been investigated. Several studies have shown
that curcumin inhibits lung cancer cell invasion and migration
(7–9). In agreement with these previous
studies, the present study also observed marked inhibitory effects
of BDMC on 95D-cell invasion and migration.
A recent study indicated that the induction of the
epithelial-to-mesenchymal transition (EMT) has an important role in
cancer progression (24). During
EMT, epithelial cells lose cell-cell contacts, cell polarity and
epithelial markers, particularly E-cadherin, while acquiring
mesenchymal markers including vimentin and fibronectin;
furthermore, the epithelial cells acquire mesenchymal phenotypes
with increased motility and invasiveness (25). Therefore, loss of E-cadherin
expression and acquisition of vimentin are the major features of
EMT. EMT is considered to be regulated by transcription factors,
including Twist, Zeb2 (SIP1), Snail1 and Slug (Snail2) (26,27).
E-cadherin is a transmembrane protein mediating cell-cell adhesion
between epithelial cells, which has an important role in cell
migration and is thus involved in tumor metastasis. Low E-cadherin
expression in NSCLC tumors has been reported to be associated with
a more aggressive behavior of tumor epithelial cells and with an
unfavorable prognosis (28).
E-cadherin expression is associated with tumor differentiation,
lymph node metastasis and prognosis in patients with NSCLC
(29). Vimentin is one of the
markers of tumor differentiation and invasion in lung cancer, which
predicts the occurrence of metastases in NSCLCs. Vimentin
expression was found to be an independent prognostic marker for
poor survival in resected NSCLC patients (30). Vimentin overexpression in cancer is
correlated with accelerated tumor growth, invasion and poor
prognosis (31,32).
In addition, it has been reported that
death-effector domain-containing DNA-binding protein activated
autophagy and attenuated EMT by inducing the degradation of Snail
and Twist in human breast cancer (33). Furthermore, the EMT has been
associated with drug resistance and cancer cell metastasis
(33). During EMT, the expression
of E-cadherin, a transmembrane protein essential for stable
adherens junctions, is downregulated, while the expression of the
mesenchymal molecules vimentin, fibronectin, and/or N-cadherin is
upregulated. In order to investigate the underlying molecular
mechanisms of the inhibitory effects of BDMC on 95D-cell migration
and invasion, the present study assessed the effects of BDMC on
alterations in the protein levels of vimentin and E-cadherin in 95D
cells. It was observed that BDMC markedly decreased vimentin
expression, while upregulating E-cadherin expression in 95D cells
in a dose- and time-dependent manner. These results indicated that
BDMC suppressed NSCLC cell invasion and metastasis through the
inhibition of vimentin and the promotion of E-cadherin
expression.
Autophagy, a strategy of self-adaption, has been
regarded as a double-edged sword in tumor metastasis. The role of
autophagy in cell migration and invasion has been studied in depth
(34). Autophagy is known to exert
anti-metastatic effects; however, it also has pro-metastasis
activity in certain cases (35).
In the present study, autophagy was identified as a target of BDMC,
via which its anti-cancer effects, including the inhibition of
invasion and migration in NSCLC, were mediated in vitro.
Interfering with the autophagic pathway may affect
the migration and invasion of tumor cells. The present study was
the first, to the best of our knowledge, to demonstrate that the
inhibition of autophagy by knocking down beclin-1 significantly
attenuated the inhibitory effects of BDMC on the migration and
invasion of 95D cells, suggesting that autophagy was involved in
BDMC-mediated suppression of 95D-cell invasion and migration.
It was therefore hypothesized that BDMC inhibited
the invasion and migration of 95D cells by inducing autophagy,
which modulated the EMT process through promotion of the
degradation of transcriptional factors, including Twist and Snail.
The specific mechanisms require further study.
In conclusion, the present study was the first, to
the best of our knowledge, to show that BDMC significantly
suppressed the invasion and migration of highly metastatic human
lung cancer cells through downregulating vimentin and upregulating
E-cadherin expression. Furthermore, autophagy was shown to be
involved in the anti-tumor effects of BDMC in NSCLC. The mechanisms
of action of BDMC are required to be addressed more systematically
in further studies.
Acknowledgments
The authors would like to thank Dr Hu Luo and Miss
Qian Qiu, from the department of Respiratory Medicine of Southwest
Hospital (Chongqing, China) for their keen interest and help.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dempke WC, Suto T and Reck M: Targeted
therapies for non-small cell lung cancer. Lung Cancer. 67:257–274.
2010. View Article : Google Scholar
|
|
3
|
Kawachi R, Tsukada H, Nakazato Y, Takei H,
Furuyashiki G, Koshi-ishi Y and Goya T: Early recurrence after
surgical resection in patients with pathological stage I non-small
cell lung cancer. Thorac Cardiovasc Surg. 57:472–475. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ji C, Cao C, Lu S, Kivlin R, Amaral A,
Kouttab N, Yang H, Chu W, Bi Z, Di W and Wan Y: Curcumin attenuates
EGF-induced AQP3 up-regulation and cell migration in human ovarian
cancer cells. Cancer Chemother Pharmacol. 62:857–865. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zou P, Helson L, Maitra A, Stern ST and
McNeil SE: Polymeric curcumin nanoparticle pharmacokinetics and
metabolism in bile duct cannulated rats. Mol Pharm. 10:1977–1987.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sakaue-Sawano A, Kurokawa H, Morimura T,
Hanyu A, Hama H, Osawa H, Kashiwagi S, Fukami K, Miyata T, Miyoshi
H, et al: Visualizing spatiotemporal dynamics of multicellular
cell-cycle progression. Cell. 132:487–498. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen HW, Lee JY, Huang JY, Wang CC, Chen
WJ, Su SF, Huang CW, Ho CC, Chen JJ, Tsai MF, et al: Curcumin
inhibits lung cancer cell invasion and metastasis through the tumor
suppressor HLJ1. Cancer Res. 68:7428–7438. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin SS, Lai KC, Hsu SC, Yang JS, Kuo CL,
Lin JP, Ma YS, Wu CC and Chung JG: Curcumin inhibits the migration
and invasion of human A549 lung cancer cells through the inhibition
of matrix metalloproteinase-2 and -9 and Vascular Endothelial
Growth Factor (VEGF). Cancer Lett. 285:127–133. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang CL, Liu YY, Ma YG, Xue YX, Liu DG,
Ren Y, Liu XB, Li Y and Li Z: Curcumin blocks small cell lung
cancer cells migration, invasion, angiogenesis, cell cycle and
neoplasia through Janus kinase-STAT3 signalling pathway. PLoS One.
7:e379602012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sandur SK, Pandey MK, Sung B, Ahn KS,
Murakami A, Sethi G, Limtrakul P, Badmaev V and Aggarwal BB:
Curcumin, demethoxycurcumin, bisdemethoxycurcumin,
tetrahydrocurcumin and turmerones differentially regulate
anti-inflammatory and anti-proliferative responses through a
ROS-independent mechanism. Carcinogenesis. 28:1765–1773. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yodkeeree S, Chaiwangyen W, Garbisa S and
Limtrakul P: Curcumin, demethoxycurcumin and bisdemethoxycurcumin
differentially inhibit cancer cell invasion through the
down-regulation of MMPs and uPA. J Nutr Biochem. 20:87–95. 2009.
View Article : Google Scholar
|
|
12
|
Ikeda T, Ishii KA, Saito Y, Miura M,
Otagiri A, Kawakami Y, Shimano H, Hara H and Takekoshi K:
Inhibition of autophagy enhances sunitinib-induced cytotoxicity in
rat pheochromocytoma PC12 cells. J Pharmacol Sci. 121:67–73. 2013.
View Article : Google Scholar
|
|
13
|
Lagisetty P, Vilekar P, Sahoo K, Anant S
and Awasthi V: CLEF MA-an anti-proliferative curcuminoid from
structure-activity relationship studies on 3,5-bis (benzylidene)
-4-piperidones. Bioorg Med Chem. 18:6109–6120. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiao K, Jiang J, Guan C, Dong C, Wang G,
Bai L, Sun J, Hu C and Bai C: Curcumin induces autophagy via
activating the AMPK signaling pathway in lung adenocarcinoma cells.
J Pharmacol Sci. 123:102–109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Maejima Y, Kyoi S, Zhai P, Liu T, Li H,
Ivessa A, Sciarretta S, Del Re DP, Zablocki DK, Hsu CP, et al: Mst1
inhibits autophagy by promoting the interaction between Beclin1 and
Bcl-2. Nat Med. 19:1478–1488. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boonrao M, Yodkeeree S, Ampasavate C,
Anuchapreeda S and Limtrakul P: The inhibitory effect of turmeric
curcuminoids on matrix metalloproteinase-3 secretion in human
invasive breast carcinoma cells. Arch Pharm Res. 33:989–998. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Killian PH, Kronski E, Michalik KM,
Barbieri O, Astigiano S, Sommerhoff CP, Pfeffer U, Nerlich AG and
Bachmeier BE: Curcumin inhibits prostate cancer metastasis in vivo
by targeting the inflammatory cytokines CXCL1 and -2.
Carcinogenesis. 33:2507–2519. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Siwak DR, Shishodia S, Aggarwal BB and
Kurzrock R: Curcumin-induced antiproliferative and proapoptotic
effects in melanoma cells are associated with suppression of
IkappaB kinase and nuclear factor kappaB activity and are
independent of the B-Raf/mitogen-activated/extracellular
signal-regulated protein kinase pathway and the Akt pathway.
Cancer. 104:879–890. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin YG, Kunnumakkara AB, Nair A, Merritt
WM, Han LY, Armaiz-Pena GN, Kamat AA, Spannuth WA, Gershenson DM,
Lutgendorf SK, et al: Curcumin inhibits tumor growth and
angiogenesis in ovarian carcinoma by targeting the nuclear
factor-kappaB pathway. Clin Cancer Res. 13:3423–3430. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Glienke W, Maute L, Wicht J and Bergmann
L: Curcumin inhibits constitutive STAT3 phosphorylation in human
pancreatic cancer cell lines and downregulation of survivin/BIRC5
gene expression. Cancer Invest. 28:166–171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Anuchapreeda S, Tima S, Duangrat C and
Limtrakul P: Effect of pure curcumin, demethoxycurcumin and
bisdemethoxycurcumin on WT1 gene expression in leukemic cell lines.
Cancer Chemother Pharmacol. 62:585–594. 2008. View Article : Google Scholar
|
|
22
|
Gao W, Chan JY, Wei WI and Wong TS:
Anti-cancer effects of curcumin on head and neck cancers.
Anticancer Agents Med Chem. 12:1110–1116. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qiao Q, Jiang Y and Li G: Inhibition of
the PI3 K/AKT-NF-kappaB pathway with curcumin enhanced
radiation-induced apoptosis in human Burkitt's lymphoma. J
Pharmacol Sci. 121:247–256. 2013. View Article : Google Scholar
|
|
24
|
Termen S, Tan EJ, Heldin CH and Moustakas
A: p53 regulates epithelial-mesenchymal transition induced by
transforming growth factor β. J Cell Physiol. 228:801–813. 2013.
View Article : Google Scholar
|
|
25
|
Wang H, Zhang H, Tang L, Chen H, Wu C,
Zhao M, Yang Y, Chen X and Liu G: Resveratrol inhibits
TGF-β1-induced epithelial-to-mesenchymal transition and suppresses
lung cancer invasion and metastasis. Toxicology. 303:139–146. 2013.
View Article : Google Scholar
|
|
26
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: at the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang YL, Chen MW and Xian L: P rognostic
and clinicopathological significance of downregulated E-cadherin
expression in patients with non-small cell lung cancer (NSCLC): A
meta-analysis. PLoS One. 6:e997632014. View Article : Google Scholar
|
|
29
|
Liu D, Huang C, Kameyama K, Hayashi E,
Yamauchi A, Kobayashi S and Yokomise H: E-cadherin expression
associated with differentiation and prognosis in patients with
non-small cell lung cancer. Ann Thorac Surg. 71:949–954. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Al-Saad S, Al-Shibli K, Donnem T, Persson
M, Bremnes RM and Busund LT: The prognostic impact of NF-kappaB
p105, vimentin, E-cadherin and Par6 expression in epithelial and
stromal compartment in non-small-cell lung cancer. Br J Cancer.
99:1476–1483. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Satelli A and Li S: Vimentin in cancer and
its potential as a molecular target for cancer therapy. Cell Mol
Life Sci. 68:3033–3046. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kokkinos MI, Wafai R, Wong MK, Newgreen
DF, Thompson EW and Waltham M: Vimentin and epithelial-mesenchymal
transition in human breast cancer-observations in vitro and in
vivo. Cells Tissues Organs. 185:191–203. 2007. View Article : Google Scholar
|
|
33
|
Lv Q, Wang W, Xue J, Hua F, Mu R, Lin H,
Yan J, Lv X, Chen X and Hu ZW: DEDD interacts with PI3KC3 to
activate autophagy and attenuate epithelial-mesenchymal transition
in human breast cancer. Cancer Res. 72:3238–3250. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Macintosh RL, Timpson P, Thorburn J,
Anderson KI, Thorburn A and Ryan KM: Inhibition of autophagy
impairs tumor cell invasion in an organotypic model. Cell Cycle.
11:2022–2029. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Z, Yang L, Xie L, Xu L, Xu D and Liu
X: Statins, autophagy and cancer metastasis. Int J Biochem Cell
Biol. 45:745–752. 2013. View Article : Google Scholar
|