Introduction
Obesity is an increasingly prevalent metabolic
disorder and has become an epidemic condition in developed
countries in recent decades (1).
In addition, the prevalence of metabolic syndromes and various
chronic diseases, including diabetes, fatty liver, coronary artery
disease and hypertension, are increasing due to the rise of obesity
(2). Increases in body fat mass
result from an increase in the number and size of adipocytes
(3,4). Previous studies have demonstrated
that the induction of obesity, resulting in alterations in the
number of adipocytes (adipogenic differentiations) and adipocyte
size (lipid accumulations) can be initiated by dietary factors
(3,4). A previous study demonstrated that
increases in the number of adipocytes during the aging process may
influence the development of obesity observed in older individuals
(5). Thus, adipogenesis may be an
important factor in the development of obesity.
Adipocytes differentiate from mesenchymal stem
cells, which have the capacity for differentiation into myoblasts,
chondroblasts, osteoblasts or adipocytes (6). Tissue-specific differentiation is
regulated by a variety of differentiation factors in accordance
with the cells' conditions. Among the differentiation factors
influencing adipogenesis, peroxisome proliferator-activated
receptor γ (PPARγ) and CCAAT/enhancer binding proteins (C/EBP-α,
C/EBP-β and C/EBP-γ) are considered the key factors for the
induction of adipogenesis in mesenchymal stem cell-mediated
differentiation. These two factors are required for the expression
of adipocyte-specific genes such as adiponectin (7).
Sphingosine-1-phosphate (S1P) is part of a key group
of signaling sphingolipids recognized to serve diverse roles in a
variety of cellular processes, including apoptosis, migration,
differentiation and proliferation in a variety of cell types,
including endothelial cells, smooth muscle cells, mesenchymal stem
cells and macrophages (8–10). S1P has been demonstrated to act as
a ligand of G-protein-coupled receptors, namely S1P receptors
(11). Five members of the S1P
receptor group (S1P1–5) have been identified in mammals,
which possess distinct expression profiles and affinities toward
S1P (12,13). In particular, S1P2
receptors are widely expressed throughout the body, including in
the brain, heart, lung, thymus, kidney, spleen and adipose tissues
(14,15). S1P2 receptors are
associated with differentiation of various tissue types, including
those associated with the central nervous system (CNS), as well as
the differentiation of mesenchymal stem cells and osteoblasts
(10,16,17).
During early stages of CNS development, S1P2 receptors
have been identified to be expressed in young animals and
differentiate into neuronal cells (16). In addition, S1P2
receptors upregulate myogenic differentiation of myoblast cells and
adipose-derived mesenchymal stem cells (10,17,18).
However, to date, the effect of S1P-mediated S1P2
receptors on adipogenic differentiation has remained to be fully
elucidated.
The present study investigated the hypothesis that
S1P inhibits adipocyte differentiation via the regulation of
S1P2 receptors. Therefore the effects of up- or
downregulation of S1P2 receptors on differentiation of
adipocytes were examined. Furthermore, the levels of the adipogenic
differentiation markers PPARγ, C/EBP-α and adiponectin were
assessed.
Materials and methods
Reagents
S1P was purchased from Cayman Chemical Company (Ann
Arbor, MI, USA). S1P was prepared as a 2-mM solution in 0.3 M NaOH
and then further diluted in cell culture medium.
3-isobutyl-1-methylxanthine, dexamethasone and insulins were
purchased from Sigma-Aldrich (St. Louis, MO, USA). S1P2
antibody and normal goat immunoglobulin G (IgG) were purchased from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Cell culture and differentiation
Pre-adipocyte cell line 3T3-L1 cells and human
embryonic kidney cell line HEK 293 cells were obtained from the
American Type Culture collection (Rockville, MD, USA). The 3T3-L1
cells were maintained in Dulbecco's modified Eagle's medium (DMEM)
containing 10% fetal bovine serum and antibiotics: 100 µg/ml
gentamycin (Invitrogen Life Technologies, Carlsbad, CA, USA) and
100 µg/ml penicillin-streptomycin (HyClone, Logan, UT, USA).
To induce differentiation, 2 days post-confluent 3T3-L1 cells were
incubated in MDI induction media (DMEM containing 10% fetal bovine
serum, 0.5 mM 3-isobutyl-1-methylxanthine, 1 µm
dexamethasone and 1 µg/ml insulin) for 2 days. In certain
experiments, S1P (10 µM) was added at the time of the
induction of differentiation. Two days subsequent to the addition
of MDI (day 2), the media was replaced with insulin media. The
AdipoRed assay and detection of glycerol release contents were
performed on day 7.
Construction of recombinant
adenoviruses
Mouse S1P2-expressing adenoviruses and
empty vector adenoviruses were purchased from Genenmed, Inc.
(Seoul, Korea; Gen-E008-001). Recombinant adenoviruses were
amplified in human embryonic kidney HEK-293 cells and purified
using the Vivapure AdenoPACK kit (Sartorius AG, Göttingen, Germany)
according to the manufacturer's instructions (19).
S1P2 RNA interference
3T3-L1 cells were transfected with validated
Stealth™ small interfering (si)RNAs (Invitrogen Life Technologies)
directed against S1P2 using Lipofectamine 2000
(Invitrogen Life Technologies) RNA interference transfection
protocol. Sequences of the S1P2 siRNAs used were as
follows: Sense, 5′-AGAAGAUUCUCCACCACGAUGGCGC-3′ and anti-sense,
5′-GCGCCAUCGUGGUGGAGAAUCUUCU-3′. The Stealth™ RNA interference
negative control (medium G/C) were also obtained from Invitrogen
Life Technologies.
Quantification of lipid content
Lipid content was quantified using the commercially
available AdipoRed Assay Reagent (Lonza, Verviers, Belgium) in
accordance with the manufacturer's instructions. In brief,
pre-adipocytes were grown in 24-well plates and then incubated with
MDI media with or without S1P during the adipogenic phase. On day
7, the culture supernatant was removed and the cells were carefully
washed with 500 µl phosphate-buffered saline (PBS). The
wells were then filled with 300 µl PBS and 30 µl
AdipoRed reagent, followed by incubation for 10 min at 37°C.
Fluorescence was measured with excitation at 485 nm and emission at
572 nm.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from 3T3-L1 cells treated
with S1P using the Easy-spin™ total RNA extraction kit (Intron
Biotechnology, Inc., Seongnam, Korea). cDNA synthesis was performed
following the instructions of the Takara Prime Script™ 1st Strand
cDNA Synthesis kit (Takara Bio, Inc., Otsu, Japan). For RT-qPCR, 1
µl gene primers with iTaq SYBR Green supermix (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) in a 20-µl reaction
volume was used. The sequences of the primers used for RT-qPCR were
as follows: S1P2 forward, 5′AACAGCAAGTTCCACTCAGCAATG3′
and reverse, 5′GGCGGAGAGCGTGATGAAGG3′; PPARγ forward,
5′CGGAAGCCCTTTGGTGACTTTATG3′ and reverse,
5′GCAGCAGGTTGTCTTGGATGTC3′; C/EBP-α forward,
5′CGGGAACGCAACAACATCGC3′ and reverse, 5′TGTCCAGTTCACGGCTCAGC3′;
adiponectin forward, 5′TGACGGCAGCACTGGCAAG3′ and reverse,
5′TGATACTGGTCGTAGGTGAAGAGAAC3′; and β-actin forward,
5′TGAGAGGGAAATCGTGCGTGAC3′ and reverse,
5′GCTCGTTGCCAATAGTGATGACC3′. All primers were purchased from
Bioneer Inc. (Daejeon, Korea).
All reactions with iTaq SYBR Green Supermix were
performed on the CFX96 Real-Time PCR Detection system (Bio-Rad
Laboratories, Inc.). The PCR program was as follows: Denaturation
(95°C for 10 min), amplification and quantification for 40 cycles
(95°C for 10 sec, 55–60°C for 30 sec, and 72°C for 30 sec with a
single fluorescent measurement), melting curve analysis (65–95°C,
with a heating rate 0.2°C/sec and continuous fluorescence
measurement), and final cooling to 12°C.
The amplification of specific RT-qPCR products was
confirmed by performing a melting-curve step at the end of each
run. Across all the assays, none of the quantification cycle (Cq)
values were higher than 40. No-template and no-reverse
transcription controls were run to determine any contamination or
the generation of primer dimers. All amplifications were run in
triplicate.
Western blot analysis
The 3T3-L1 cells were lysed in lysis buffer [25 mM
4-(2-hydroxyethyl)-1-piperazineethane-sulfonic acid; pH 7.4, 100 mM
NaCl, 1 mM EDTA, 5 mM MgCl2, 0.1 mM dithiothreitol and
protease inhibitor mixture]. All reagents for cell lysis were
purchased from Sigma-Aldrich. Proteins were electrophoretically
resolved by 8–15% SDS-PAGE and immunoblotting was performed as
previously described (20). Images
were captured using the Fusion FX7 Acquisition system (Vilbert
Lourmat GmbH, Eberhardzell, Germany). The immunoreactive bands were
detected with an enhanced chemiluminescence detection system
(Thermo Fisher Scientific). The antibodies used for immunoblotting
were PPARγ (cat. no. sc-7273; Santa Cruz Biotechnology, Inc.),
S1P2 (cat. no. sc-31577; Santa Cruz Biotechnology),
phosphorylated-c-Jun N-terminal kinase (p-JNK) (cat. no. 9255; Cell
Signaling Technology, Inc., Danvers, MA, USA) and β-actin (cat. no.
A5441; Sigma-Aldrich). Goat, (cat. no. sc-3887) mouse (cat. no.
sc-2025) and rabbit (cat. no. sc-2027) secondary antibodies were
purchased from Santa Cruz Biotechnology, Inc.
Statistical evaluation
All values are expressed as the mean ± standard
error and were compared using Student's t-test and analysis of
variance with Duncan's test. The SAS statistical package version
8.1 (SAS Institute, Inc., Cary, NC, USA) was used for analysis.
P<0.05 was considered to indicate a statistically significant
difference.
Results
S1P2 receptor overexpression
suppresses adipogenesis and enhances the anti-adipogenic effects of
S1P
To verify the differentiative role of
S1P2 receptors in adipogenesis, 3T3-L1 cells were
infected with adenovirus for the expression of either the
S1P2 receptor or an adenovirus carrying an empty vector
at multiplicity of infection values of 500, 1,000 and 2,000,
followed by incubation in MDI. Overexpression of S1P2
receptor proteins reduced the triglyceride accumulation induced by
MDI-mediated adipocyte differentiation, whereas empty
vector-transfected cells showed similar amounts of lipid
accumulation to those in the control group incubated in MDI only
(Fig. 1A and B). To confirm the
anti-adipogenic function of the S1P2 receptor,
S1P2 receptor-overexpressing cells were co-treated with
S1P. The results showed that S1P treatment inhibited MDI-mediated
adipogenesis and that simultaneous S1P2 receptor
overexpression markedly enhanced the inhibition of MDI-mediated
adipogenesis of 3T3-L1 adipocytes (Fig. 1C and D). These results provided
further evidence for the anti-adipogenic effects of S1P being
mediated via the activation of S1P2 receptor
signaling.
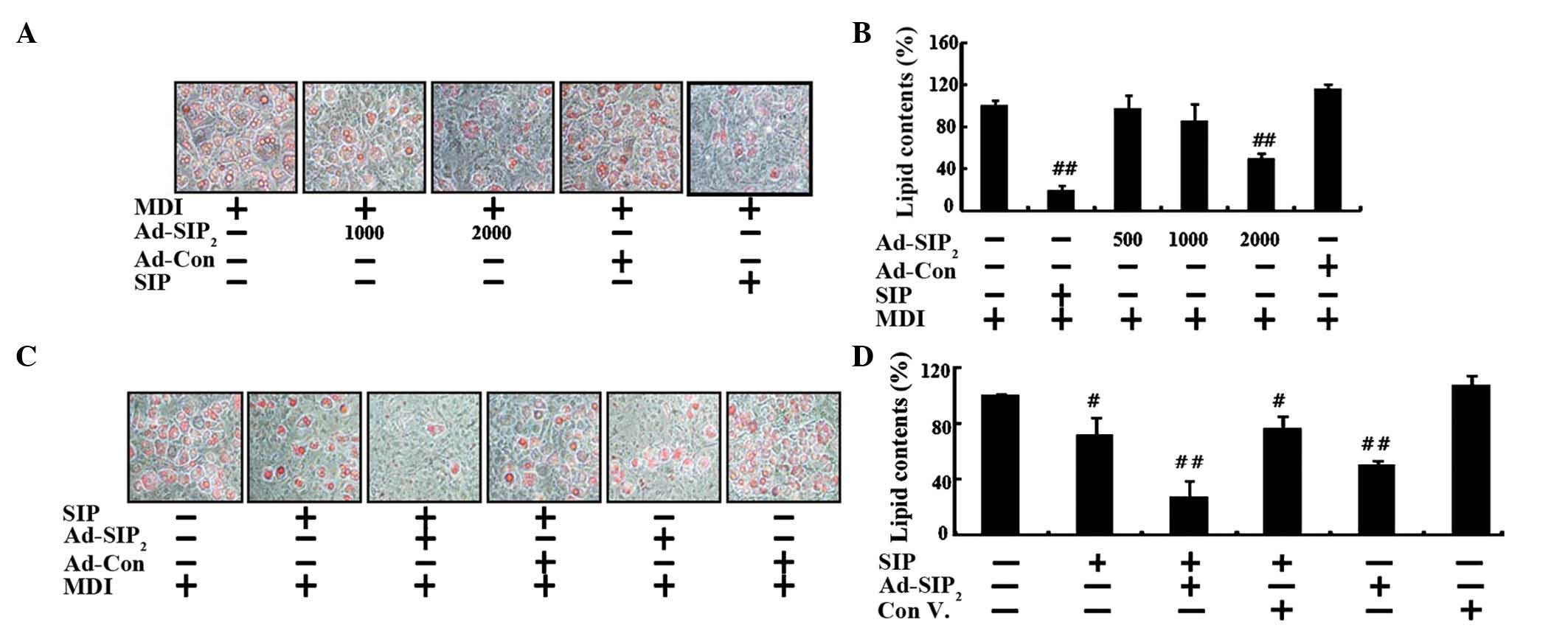 | Figure 1Overexpression of the S1P2
receptor suppresses adipogenesis and enhances anti-adipogenic
effects resulting from S1P treatment. (A) 3T3-L1 cells infected
with adenoviruses expressing empty vector (Ad-Con; MOI, 2,000) or
mouse S1P2 receptor (Ad-S1P2) for 48 h were
induced to differentiate for 6 days. AdipoRed assays were
performed. Adipocytes are indicated by red staining (magnification,
x200). (B) Quantified fluorescence intensities. (C) 3T3-L1 cells
infected with adenoviruses expressing empty vector (Ad-Con; MOI,
2,000) or mouse S1P2 (Ad-S1P2) for 48 h were
treated with S1P (0.1 µM) and induced to differentiate for 6
days. AdipoRed assays were performed as described (magnification,
x200). (D) Quantified fluorescence intensities. Values are
expressed as the mean ± standard error (n=3).
#P<0.05, ##P<0.01 vs. MDI control. The
experiments were performed in triplicate and data represent three
independent experiments. S1P2, sphingosine 1-phosphate
2; Ad, adenovirus; Con, control; MOI, multiplicity of infection;
MDI, media. |
S1P2 receptor overexpression
suppresses mRNA expression of adipogenic factors
The present study further investigated whether the
anti-adipogenic effects of S1P2 receptors are involved
in mediating the mRNA and protein expression levels of adipogenic
transcriptional factors (Fig.
2A–C). The expression levels of PPARγ, C/EBP-α and adiponectin
mRNA were observed to be increased in MDI-treated adipocytes, and
the elevated mRNA expression levels were suppressed by
S1P2 receptor overexpression. Western blot analysis
confirmed that the S1P2 receptor was overexpressed in
the adipocytes treated with the adenoviral S1P2
overexpression vector; furthermore, S1P2 receptor
overexpression downregulated the phosphorylation of JNK and
expression of PPARγ protein (Fig.
2D). These results indicated that the activation of the
S1P2 receptor caused by S1P2 receptor
overexpression suppressed mRNA and protein expression levels of
adipogenic factors, which exerted anti-adipogenic effects on 3T3-L1
adipocytes.
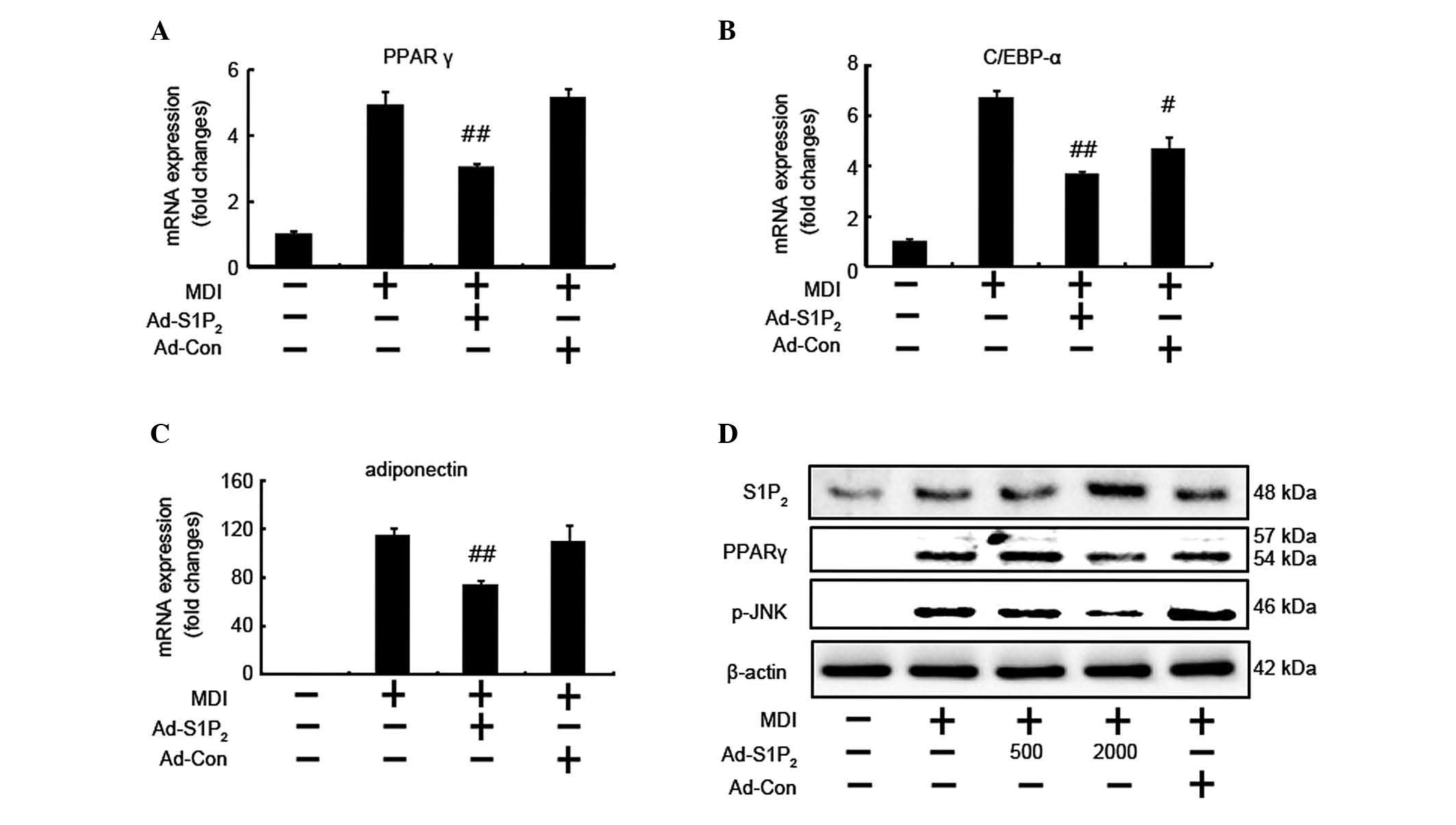 | Figure 2Overexpression of the S1P2
receptor suppresses adipogenic factors. (A–C) 3T3-L1 cells infected
with adenoviruses expressing empty vector (Ad-Con; multiplicity of
infection, 2,000), or mouse S1P2 (Ad-S1P2)
for 48 h were induced to differentiate for 2 days and harvested at
day 2 during the differentiation period. The mRNA expression of (A)
PPARγ, (B) C/EBP-α and (C) adiponectin was analyzed by reverse
transcription-quantitative polymerase chain reaction. Values are
expressed as the mean ± standard deviation of data from three
separate experiments; each experiment was performed in triplicate.
(D) Pre-adipocytes were treated as described. At day 2, the protein
levels of S1P2, PPARγ and p-JNK were analyzed by western
blot analysis. The experiments were repeatedly performed to confirm
the results. The data were analyzed using Student's t-test.
#P<0.05, ##P<0.01 vs. MDI control.
S1P2, sphingosine 1-phosphate 2; Ad, adenovirus; Con,
control; PPARγ, peroxisome proliferator-activated receptor γ;
p-JNK, phosphorylated c-Jun N-terminal kinase; MDI, media. |
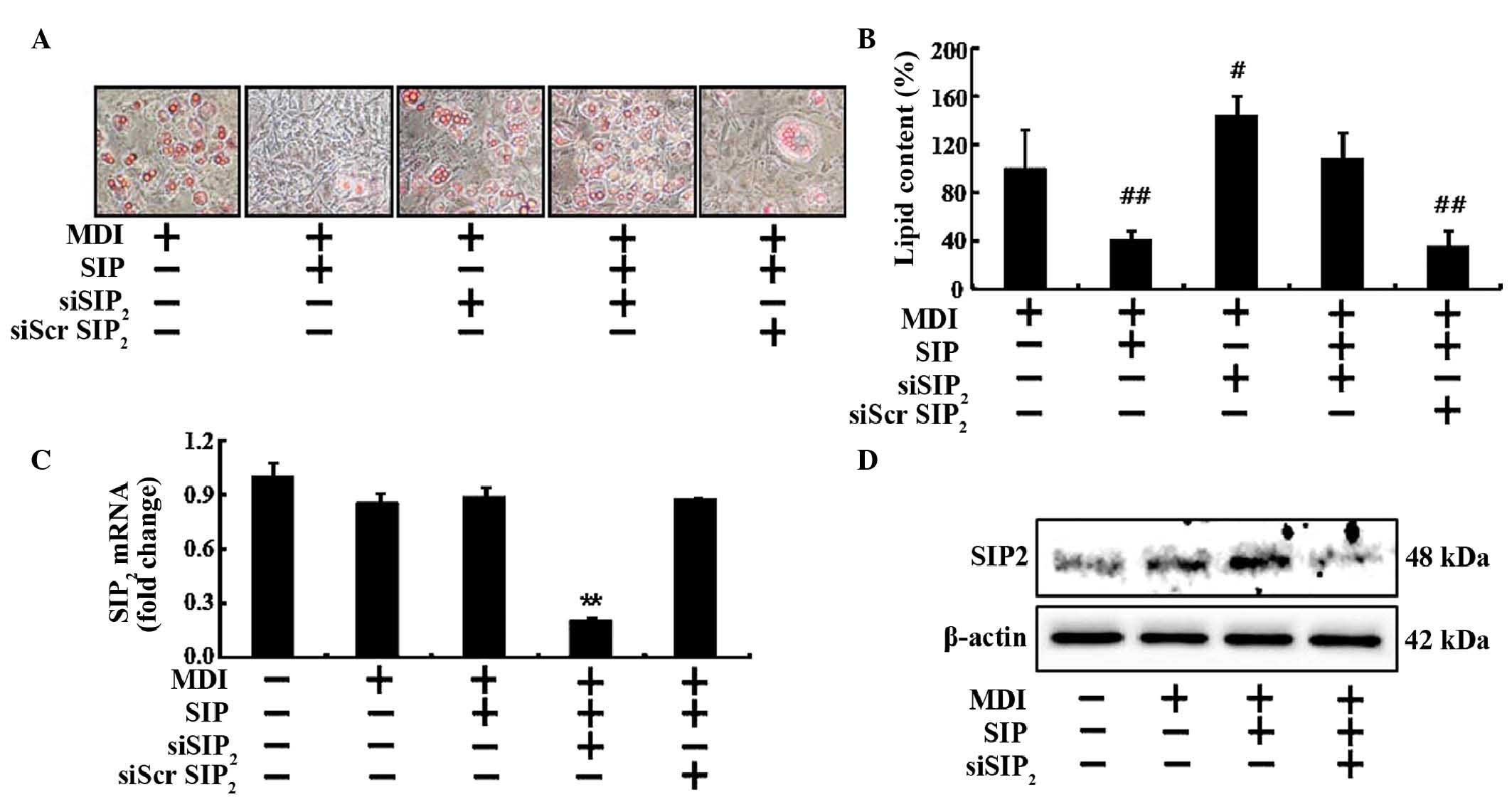
Silencing of the S1P2 receptor
abolishes the inhibition of lipid accumulation by S1P
siRNA was used to eliminate the S1P2 in
pre-adipocytes and determine the effects of S1P2
knockdown on the S1P-mediated adipocyte differentiation. 3T3-L1
cells were transfected with either S1P2 siRNA or
negative control siRNA, and 3T3-L1 cells at 2 days post-confluence
were then incubated in MDI induction media with or without S1P (10
µM) for 2 days. The media was then replaced with insulin
media to further induce adipogenesis. Following S1P2
knockdown, S1P completely lost its ability to impair adipogenic
differentiation (Fig. 3A and B).
S1P2 mRNA and protein expression levels were markedly
reduced in cells treated with S1P2 siRNA, relative to
those in cells treated with control siRNA (Fig. 3C and D). Of note, S1P2
protein expression was upregulated following treatment with S1P,
indicating that S1P may inhibit adipogenic differentiation via
upregulation of S1P2 (Fig.
3D).
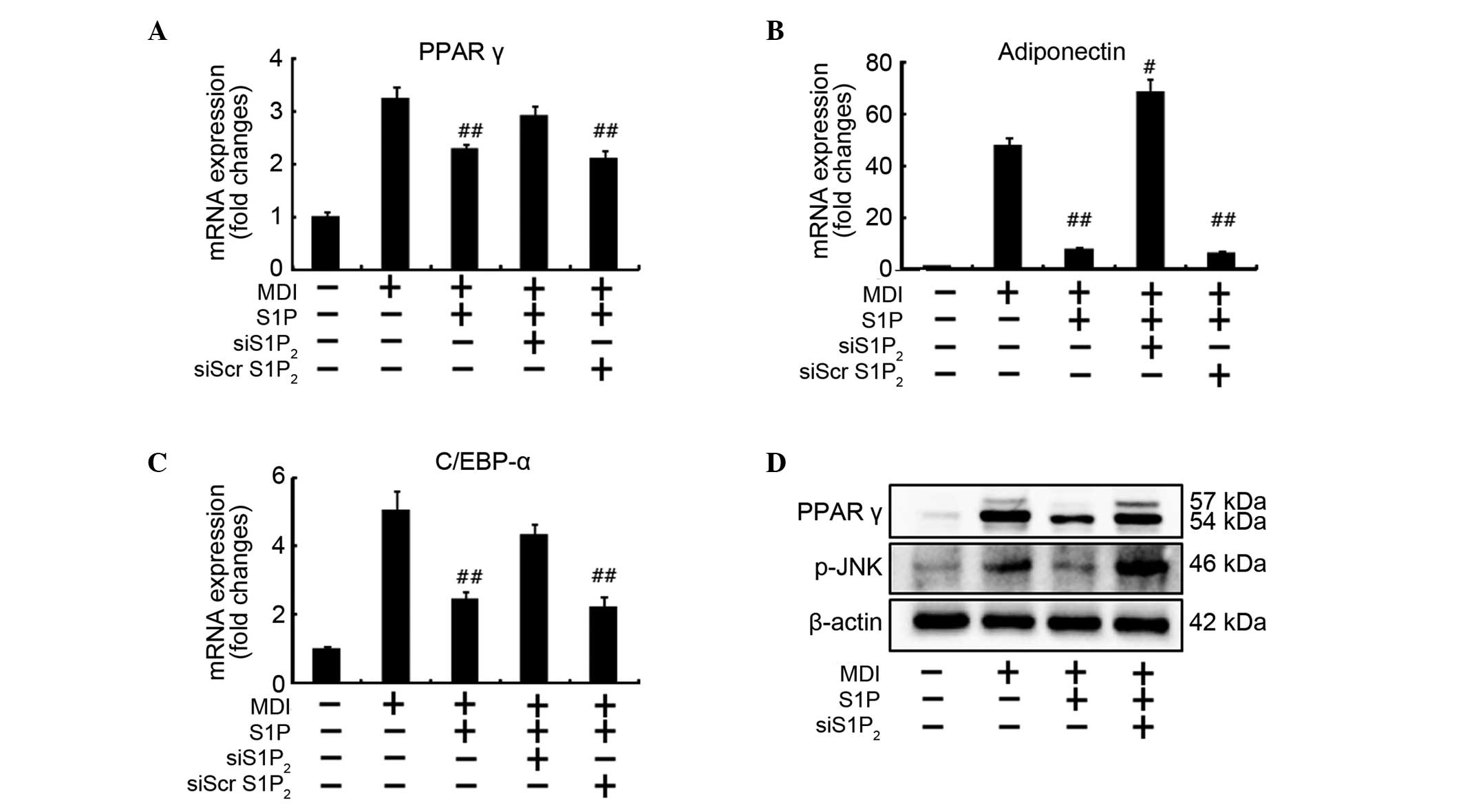
Silencing of the S1P2 receptor
abolishes the S1P-induced downregulation of PPARγ, C/EBP-α and
adiponectin expression
The present study further investigated whether the
downregulation of the expression of major adipogenic
transcriptional factors by S1P was a S1P2
receptor-mediated process. Silencing of the S1P2
receptor resulted in restoration of the mRNA expression levels of
PPARγ, C/EBP-α to levels similar to those in the control group
incubated with MDI only, and an increased adiponectin expression
compared to that in the MDI-non-treated group (Fig. 4A–C). The results indicated that S1P
inhibits adipogenic differentiation by downregulation of the major
adipogenic transcriptional factors, which were involved in
S1P2 receptor-mediated signaling. Silencing of the
S1P2 receptor additionally blocked the downregulation of
PPARγ protein and phospho-JNK protein induced by S1P treatment
(Fig. 4D). These results
demonstrated that silencing of the S1P2 receptor using
S1P2 siRNA abolishes the inhibitory effect of S1P on
adipogenesis, indicating that the S1P2 receptor may
serve a pivotal role in the regulation of adipogenic
differentiation.
Discussion
Adipocytes are generated by differentiation of
mesenchymal stem cells (6).
Mesenchymal stem cells possess the ability to differentiate into
numerous cell types, including adipocytes, osteoblasts,
chondrocytes and smooth muscle cells. Previous studies have
suggested that myogenesis, adipo-genesis and fibrogenesis are
competitive processes in the differentiation of mesenchymal stem
cells (6,10). In addition, Nincheri et al
(10) demonstrated that adipose
tissue-derived mesenchymal stem cells differentiated into smooth
muscle cells via the upregulation of S1P2 receptors.
Thus, it was hypothesized that upregulation of S1P2
receptors may inhibit the differentiation of progenitor adipocytes
into adipocytes. The results of previous studies are in agreement
with those of the present study, indicating that overexpression of
S1P2 receptors inhibits the differentiation of 3T3-L1
pre-adipocytes into adipocytes.
The recruitment of fat cells in adipose tissue
requires the differentiation of pre-adipocytes into adipocytes
(adipogenesis), a process tightly controlled by the transcription
factors PPARγ and C/EBP-α (21,22).
In particular, PPARγ is regarded as the key regulator of
adipogenesis. Forced expression of PPARγ is sufficient to induce
adipocyte differentiation in fibroblasts, and no factor is known
that promotes adipogenesis in the absence of PPARγ (21–23).
In the present study, overexpression of S1P2 had
inhibitory effects on PPARγ and C/EBP-α expression in 3T3-L1
adipocytes. In addition, S1P2 knockdown abrogated the
downregulation of PPARγ and C/EBP-α. These observations suggested
that S1P2 activation may have anti-adipogenic effects in
adipogenic differentiation.
Studies have been conducted on the effects of S1P on
cell differentiation. S1P has been reported to act as a regulator
of osteoclast differentiation (24) in addition to myogenic
differentiation (18,25). It is widely accepted that S1P and
the S1P2 receptor are associated with myogenic
differentiation of mesenchymal stem cells through G(i)-coupled S1P
receptor interactions. In addition, S1P interferes with the
differentiation of human monocytes into competent dendritic cells
(26). Numerous signaling pathways
that are activated in response to stimulation of cells by S1P are
initiated by activation of S1P-specific receptors (10,27,28).
However, it had yet to be clarified whether direct treatment with
S1P is able to influence adipogenic differentiation (10,17,18).
Therefore, the direct effect of S1P was examined in the present
study, and the results demonstrated that S1P exerted
anti-adipogenic effects via upregulation of S1P2 protein
levels.
The extracellular signal-regulated kinase, p38 and
JNK pathways are intracellular mitogen-activated protein kinase
(MAPK) signaling pathways that serve pivotal roles in numerous
essential cellular processes, including proliferation and
differentiation (3,18,29).
MAPKs are activated by a large variety of stimuli and one of their
major functions is to connect cell surface receptors to
transcription factors in the nucleus, which consequently triggers
long-term cellular responses (29). Overexpression of S1P2
was observed to inhibit MDI-induced phosphorylation of JNK in the
present study. When induced to differentiate, growth-arrested
3T3-L1 pre-adipocytes synchronously re-enter the cell cycle and
undergo mitotic clonal expansion (MCE). MCE is a pre-requisite for
differentiation of 3T3-L1 pre-adipocytes into adipocytes (30).
In conclusion, the results of the present study
suggested that the anti-adipogenic activity of S1P is mediated via
S1P2. The present study identified for the first time,
to the best of our knowledge, that the inhibitory effect of S1P on
adipogenic differentiation proceeded via the upregulation of
S1P2 and additionally suggested that S1P2
activation may be a therapeutic target for obesity. Therefore, the
development of S1P2 receptor sub-type-specific ligands
may be beneficial for potential medical interventions.
Acknowledgments
The present study was supported by a grant from the
National Research Foundation of Korea, funded by the Korean
government (grant no. 2013R1A1A2063931).
References
|
1
|
Lei F, Zhang XN, Wang W, Xing DM, Xie WD,
Su H and Du LJ: Evidence of anti-obesity effects of the pomegranate
leaf extract in high-fat diet induced obese mice. Int J Obes
(Lond). 31:1023–1029. 2007. View Article : Google Scholar
|
|
2
|
Boyle KB, Hadaschik D, Virtue S, Cawthorn
WP, Ridley SH, O'Rahilly S and Siddle K: The transcription factors
Egr1 and Egr2 have opposing influences on adipocyte
differentiation. Cell Death Differ. 16:782–789. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim KJ, Lee OH and Lee BY: Fucoidan, a
sulfated polysac-charide, inhibits adipogenesis through the
mitogen-activated protein kinase pathway in 3T3-L1 preadipocytes.
Life Sci. 86:791–797. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gregoire FM, Smas CM and Sul HS:
Understanding adipocyte differentiation. Physiol Rev. 78:783–809.
1998.PubMed/NCBI
|
|
5
|
Yanagiya T, Tanabe A and Hotta K:
Gap-junctional communication is required for mitotic clonal
expansion during adipogenesis. Obesity (Silver Spring). 15:572–582.
2007. View Article : Google Scholar
|
|
6
|
Rayalam S, Della-Fera MA and Baile CA:
Phytochemicals and regulation of the adipocyte life cycle. J Nutr
Biochem. 19:717–726. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xing Y, Yan F, Liu Y and Zhao Y: Matrine
inhibits 3T3-L1 preadipocyte differentiation associated with
suppression of ERK1/2 phosphorylation. Biochem Biophys Res Commun.
396:691–695. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Johnstone ED, Chan G, Sibley CP, Davidge
ST, Lowen B and Guilbert LJ: Sphingosine-1-phosphate inhibition of
placental trophoblast differentiation through a G(i)-coupled
receptor response. J Lipid Res. 46:1833–1839. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Goetzl EJ, Wang W, McGiffert C, Liao JJ
and Huang MC: Sphingosine 1-phosphate as an intracellular messenger
and extracellular mediator in immunity. Acta Paediatr Suppl.
96:49–52. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nincheri P, Luciani P, Squecco R, Donati
C, Bernacchioni C, Borgognoni L, Luciani G, Benvenuti S, Francini F
and Bruni P: Sphingosine 1-phosphate induces differentiation of
adipose tissue-derived mesenchymal stem cells towards smooth muscle
cells. Cell Mol Life Sci. 66:1741–1754. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schüppel M, Kurschner U, Kleuser U,
Schäfer-Korting M and Kleuser B: Sphingosine 1-phosphate restrains
insulin-mediated keratinocyte proliferation via inhibition of Akt
through the S1P2 receptor subtype. J Invest Dermatol.
128:1747–1756. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bieberich E: There is More to a lipid than
just being a fat: Sphingolipid-guided differentiation of
oligodendroglial lineage from embryonic stem cells. Neurochem.
36:1601–1611. 2011. View Article : Google Scholar
|
|
13
|
Pyne S and Pyne NJ: Sphingosine
1-phosphate signalling in mammalian cells. Biochem J. 349:385–402.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang G, Contos JJ, Weiner JA, Fukushima N
and Chun J: Comparative analysis of three murine G-protein coupled
receptors activated by sphingosine-1-phosphate. Gene. 227:89–99.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ishii I, Friedman B, Ye X, Kawamura S,
McGiffert C, Contos JJ, Kingsbury MA, Zhang G, Brown JH and Chun J:
Selective loss of sphingosine 1-phosphate signaling with no obvious
phenotypic abnormality in mice lacking its G protein-coupled
receptor, LP(B3)/EDG-3. J Biol Chem. 276:33697–33704. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Maclennan AJ, Marks L, Gaskin AA and Lee
N: Embryonic expression pattern of H218, a G-protein coupled
receptor homolog, suggests roles in early mammalian nervous system
development. Neuroscience. 79:217–224. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Roelofsen T, Akkers R, Beumer W, Apotheker
M, Steeghs I, van de Ven J, Gelderblom C, Garritsen A and Dechering
K: Sphingosine-1-phosphate acts as a developmental stage specific
inhibitor of platelet-derived growth factor-induced chemotaxis of
osteoblasts. J Cell Biochem. 105:1128–1138. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Donati C, Meacci E, Nuti F, Becciolini L,
Farnararo M and Bruni P: Sphingosine 1-phosphate regulates myogenic
differentiation: A major role for S1P2 receptor. FASEB J.
19:449–451. 2005.
|
|
19
|
Seo JS, Moon MH, Jeong JK, Seol JW, Lee
YJ, Park BH and Park SY: SIRT1, a histone deacetylase, regulates
prion protein-induced neuronal cell death. Neurobiol Aging.
33:1110–1120. 2012. View Article : Google Scholar
|
|
20
|
Moon MH, Jeong JK, Seo JS, Seol JW, Lee
YJ, Xue M, Jackson CJ and Park SY: Bisphosphonate enhances TRAIL
sensitivity to human osteosarcoma cells via death receptor 5
upregulation. Exp Mol Med. 43:138–145. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Simon MF, Daviaud D, Pradère JP, Grès S,
Guigné C, Wabitsch M, Chun J, Valet P and Saulnier-Blache JS:
Lysophosphatidic acid inhibits adipocyte differentiation via
lysophosphatidic acid 1 receptor-dependent downregulation of
peroxisome proliferator-activated receptor gamma2. J Biol Chem.
280:14656–14662. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fu L, Tang T, Miao Y, Zhang S, Qu Z and
Dai K: Stimulation of osteogenic differentiation and inhibition of
adipogenic differentiation in bone marrow stromal cells by
alendronate via ERK and JNK activation. Bone. 43:40–47. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rosen ED and MacDougald OA: Adipocyte
differentiation from the inside out. Nat Rev Mol Cell Biol.
7:885–896. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ryu J, Kim HJ, Chang EJ, Huang H, Banno Y
and Kim HH: Sphingosine 1-phosphate as a regulator of osteoclast
differentiation and osteoclastosteoblast coupling. EMBO J.
25:5840–5851. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Meacci E, Cencetti F, Donati C, Nuti F,
Farnararo M, Kohno T, Igarashi Y and Bruni P: Down-regulation of
EDG5/S1P2 during myogenic differentiation results in the specific
uncoupling of sphingosine 1-phosphate signalling to phospholipase
D. Biochim Biophys Acta. 1633:133–142. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Martino A, Volpe E, Auricchio G, Izzi V,
Poccia F, Mariani F, Colizzi V and Baldini PM: Sphingosine
1-phosphate interferes on the differentiation of human monocytes
into competent dendritic cells. Scand J Immunol. 65:84–91. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiang L, Liu T, Song K, Guan S, Li X and
Ge D: Stimulation of sphingosine-1-phosphate on cardiomyogenic
differentiation of mesenchymal stem cells. Sheng Wu Gong Cheng Xue
Bao. 29:1617–1628. 2013.In Chinese.
|
|
28
|
Zhao Z, Chen Z, Zhao X, et al:
Sphingosine-1-phosphate promotes the differentiation of human
umbilical cord mesenchymal stem cells into cardiomyocytes under the
designated culturing conditions. J Biomed Sci. 18:372011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bost F, Aouadi M, Caron L and Binétruy B:
The role of MAPKs in adipocyte differentiation and obesity.
Biochimie. 87:51–56. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tang QQ, Otto TC and Lane MD: Mitotic
clonal expansion: A synchronous process required for adipogenesis.
Proc Natl Acad Sci USA. 100:44–49. 2003. View Article : Google Scholar :
|