Introduction
Traumatic brain injury (TBI) causes primary neuronal
degeneration in the contusion site and also initiates secondary
injury in the surrounding tissue (pericontusion). The secondary
nonmechanical injury is progressive and can last for days, weeks or
months (1). Several pathological
processes are reported to be responsible for the neuronal cell
death, which occurs as a result of secondary damage following TBI,
including inflammation, apoptosis and oxidative stress (2–4).
Among these, inflammation is key in secondary injury (3,5).
Microglial activation has been widely-accepted as histological
evidence of brain injury (6). It
is also considered to be the major source of proinflammatory
cytokines following TBI (7–9). A
number of investigations have specifically targeted microglial
activation in order to retard the pathological processes associated
with inflammation following TBI (10,11).
Mammalian target of rapamycin (mTOR) is a rapamycin
sensitive serine/threonine protein kinase, which is involved in
modulating the initiation of protein synthesis (12). Growth factors, mitogens, hormones
and nutrients are able to activate the phosphoinositide 3-kinase
pathway, leading to the phosphorylation and activation of mTOR
(13,14). Activated mTOR exerts its functions
in stimulating translation through the phosphorylation and
activation of the p70S6K S6 kinase, which phosphorylates the S6RP
ribosomal protein (15). The
phosphorylation of S6RP is a critical effector of mTOR in the
regulation of protein synthesis. The mTOR pathway is important in
the synthesis of proinflammatory cytokines. Inactivation of the
mTOR pathway attenuates the expression of proinflammatory
cytokines, including interleukin (IL)-1β and tumor necrosis
factor-α (TNF-α), which promote adipose-derived stromal cell
viability and antiapoptotic/proangiogenic efficacy in vivo
(16). Thus, the mTOR pathway may
be a significant proinflammatory factor due to its ability to
induce proinflammatory cytokines, and targeting this pathway may
offer a reasonable therapeutic strategy against the inflammatory
response following TBI.
Rapamycin is a macrolide antibiotic product from
Streptomyces hygroscopicus, which specifically inhibits the
activity of mTOR. To exert its inhibitory effect on mTOR, rapamycin
binds to immunophilin FK-506-binding protein 12, which prevents the
phosphorylation of mTOR (17,18).
The function of rapamycin has been an intensive area of
investigation, and a focus of attention in cancer, development,
metabolism and central nervous system diseases (14). Rapamycin has also been investigated
for its neuroprotective effects in closed head injury TBI models.
The injection of rapamycin increases the survival of neurons and
significantly improves the functional recovery of the brain
(19). However, the effect of
rapamycin on the inflammatory reaction, which follows TBI remains
to be fully elucidated. Thus, in the present study, the possible
involvement of rapamycin on the decreased expression of
proinflammatory cytokines was examined in a mouse TBI model. In
addition, the functional significance of rapamycin on the
neurological outcome, associated with attenuating the inflammatory
reaction following TBI, was assessed.
Materials and methods
Animals
Male imprinting control region (ICR) mice
(Experiment Animal Centre of Nanjing Medical University, Jiangsu,
China) aged between 6–8 weeks, weighing 28–32 g were used in the
present study. The experimental procedures were approved by the
Animal Care and Use Committee of Nanjing Jinling Hospital and
conformed to the Guide for the Care and Use of Laboratory Animals
by the National Institutes of Health (Bethesda, MD, USA). The mice
were housed on a 12 h light/dark cycle with ad libitum
access to food and water.
Model of TBI
The model of TBI used in the present study was a
weight-drop model (Beyotime Institute of Biotechnology, Shanghai,
China), as described by Flierl et al (20). The mice were anesthetized with an
intraperitoneal (i.p.) injection of chloral hydrate (1%; 5 ml/kg;
Beyotime Institute of Biotechnology) and then placed onto a
platform directly below the weight of the weight-drop device. A 1.5
cm midline longitudinal scalp incision was made and the skull was
exposed. Subsequent to locating the left anterior frontal area (1.5
mm lateral to the midline on the mid-coronal plane) as the impact
area, a 200 g weight was released and dropped onto the skull from a
height of 2.5 cm. The mortality rate resulting from apnea was
reduced by early respiratory support. The scalp wound was closed
using standard suture material, and the mice were returned to
cages, where they had ad libitum access to water and food.
Sham-injured animals underwent the same procedures, but did not
undergo the weight-drop.
Experimental procedure
The male ICR mice were divided into four groups
(n=45 per group): Sham group, TBI group, TBI + dimethyl sulfoxide
(DMSO) group and TBI + rapamycin group. In the TBI + rapamycin
group, rapamycin (cat. no. S1039; Selleckchem, Munich, Germany) was
dissolved in DMSO (50 mg/ml; Sigma-Aldrich, St. Louis, MO, USA) and
injected into the mice 30 min after TBI (2 mg/kg; i.p.). The mice
in the TBI + DMSO group received equal volumes of vehicle (5% DMSO)
at 30 min subsequent to TBI.
Brain tissue processing
For isolation of the proteins, the animals were
anesthetized with a solution of chloral hydrate (1%, 5 ml/kg) 24 h
after TBI and were perfused intracardially with 30–40 ml cold (4°C)
heparinized 0.9% saline (Beyotime Institute of Biotechnology). The
left, ipsilateral, cerebral cortex (pericontusion) was collected,
immediately frozen in liquid nitrogen and then transferred to a
−80°C freezer until use. For immunohistochemical analysis, the
animals were sacrificed 24 h after TBI in the following way.
Following anesthesia, induced with chloral hydrate (1%, 5 ml/kg),
the animals were intracardially perfused with 30–40 ml cold
heparinized 0.9% saline followed by 20–30 ml cold 4%
paraformaldehyde (Beyotime Institute of Biotechnology). The whole
brain was removed and immersed in 4% paraformaldehyde overnight at
4°C. For immunofluorescence, the brain was subsequently immersed in
20% sucrose (Beyotime Institute of Biotechnology) followed by 30%
sucrose.
Western blot analysis
The protein concentrations were determined using the
Bradford assay (Beyotime Institute of Biotechnology) (21). Equal quantities of protein (50
µg/per lane) were separated by 10% sodium dodecyl
sulfate-polyacrylamide gel (Beyotime Institute of Biotechnology)
electrophoresis and transferred onto polyvinylidene-difluoride
membranes (EMD Millipore, Billerica, MA, USA). The membranes were
blocked for 2 h in blocking buffer (Tris-buffered saline/0.05%
Tween 20; TBST; Beyotime Institute of Biotechnology) containing 5%
skim milk) and were incubated overnight at 4°C with the following
primary antibodies in blocking buffer: Rabbit monoclonal
phospho-mTOR (Ser2448) (cat..no. #5536; 1:1,000; Cell Signaling
Technology, Danvers, MA, USA), rabbit monoclonal phospho-S6
ribosomal protein (Ser235/236) (cat..no. #4858; 1:2,000; Cell
Signaling Technology) and rabbit polyclonal GAPDH (cat. no. AP0063;
1:5,000; Bioworld Technology, Minneapolis, MN, USA). Following
washing of the membranes three times with TBST (10 min each), the
membranes were incubated with polyclonal goat anti-rabbit
horseradish peroxidase conjugated immunoglobulin G (cat. no.
BS13278; 1:5,000; Bioworld Technology) for 2 h at room temperature.
The protein bands were visualized using enhanced chemiluminescence
western blotting detection reagents (EMD Millipore) and exposure to
X-ray film (Carestream, Xiamen, China). The developed films were
digitized using an Epson Perfection 2480 scanner (Seiko Corp.,
Nagano, Japan). The band density was quantified using Un-Scan-It
6.1 software (Silk Scientific Inc., Orem, UT, USA) and the data
were normalized to GAPDH.
Immunofluorescence
For immunofluorescence, serial 8-µm coronal
sections were obtained using a cryostat (RM2235; Leica Microsystems
GmbH, Wetzlar, Germany). A total of four sets of five evenly spaced
(300 µm apart) sections, spanning the injured cortex, were
collected from each brain. Based on established immunostaining
procedures (22), slides (Beyotime
Institute of Biotechnology) were incubated in blocking buffer,
containing 10% normal goat serum in phosphate-buffered saline (PBS)
and 0.1% Triton X-100 (Beyotime Institute of Biotechnology) for 2
h, followed by incubation at 4°C overnight with the primary
antibody, rabbit anti-ionized calcium-binding adapter molecule 1
(IBA-1; cat. no. 019-19741; 1:5,000; Wako, Osaka, Japan). On the
following day, the slides were washed with PBS three times for 5
min and incubated with the appropriate secondary antibodies for 1 h
at room temperature. The slides were then washed three times in
PBS. Cover slips were applied using mounting medium. Images of the
immunofluorescence were captured using an Axio Observer A1
microscope system (Carl Zeiss, Oberkochen, Germany) and analyzed
using Image-Pro Plus 6.0 software (Media Cybernetics, Rockville,
MD, USA). The specificity of the immunofluorescence reaction was
evaluated by replacement of the primary antibody with PBS. A total
of six visual fields (magnification, ×200) surrounding the
contusion in each coronary section were randomly selected, and the
mean number of microglia in the six fields was calculated. A total
of four sections from each animal were used for quantification. The
data for each sample was the mean number of microglia in the four
sections. Data are presented as the mean number of microglia per
magnification field (magnification, ×200). All analyses were
performed by two investigators in a blinded-manner.
Enzyme-linked immunosorbent assay
(ELISA)
The expression levels of interluekin (IL)-1β and
TNF-α were analyzed using an ELISA (Biocalvin Company, Suzhou,
China), according to the manufacturer's instructions. The protein
concentrations were measured using a Bradford assay. Equal
quantities of lysate were used for the analyses of TNF-α and IL-1β,
with values expressed as pg/mg protein.
Cresyl violet (Nissl) staining
Tissue sections were stained with Cresyl Violet
(Nissl; Sigma-Aldrich), as described previously (23). Normal neurons have relatively large
cell bodies and are rich in cytoplasm, with one or two large round
nuclei. By contrast, damaged cells exhibit shrunken cell bodies,
condensed nuclei, a dark cytoplasm and numerous empty vesicles
(23). The counting of cells was
restricted to the lesion boundary zone (where the most damage was
observed). A total of six high-power fields (magnification, ×400)
in each coronary section were randomly selected, and the mean
number of surviving neurons in the six views were calculated for
each section using the Axio Observer A1 microscope system. A total
of four sections from each animal were used for quantification and
the average quantity of the four sections was calculated for each
sample. Data are presented as the quantity of neurons per
high-power field. All analyses were performed by two investigators
in a blinded-manner.
Neurobehavioral evaluation
The neurological statuses of the mice were evaluated
24 h and 72 h after TBI using a grip test and neurological severity
score (NSS). The grip test was developed to assess the gross
vestibulomotor function (24). The
mouse was placed on a thin, horizontal metal wire, measuring 45 cm
in length, which was suspended 45 cm above a foam pad between two
vertical poles. Each mouse was graded on its ability to grip,
attach and move, as described in Table
I. The grip test was performed in triplicate, with the total
score calculated for each mouse. In the NSS, the ability of each
mouse to perform 10 different tasks, which demonstrate motor
function, balance and alertness, was evaluated. A single point was
scored for failing to perform each of the tasks; thus, 0=minimum
deficit and 10=maximum deficit (Table
II) (3,20). The severity of the injury was
defined by the initial NSS, evaluated 1 h after TBI, providing a
reliable predictor of the later outcome. All neurobehavioral
assessments were performed by two investigators in a
blinded-manner.
 | Table IGrip test scoring system. |
Table I
Grip test scoring system.
| Task | Score |
|---|
| Unable to grasp
wire for 30 sec | 0 |
| Grip wire for 30
sec with one or two paws | 1 |
| Jump up and grasp
wire with four paws | 2 |
| Grasp wire with
four paws and wrap tail around | 3 |
| Crawl along the
wire for ≥5 cm | 4 |
| Crawl along the
wire to the end and dismount | 5 |
| Maximum score | 5 |
| Table IINeurological severity score scoring
system. |
Table II
Neurological severity score scoring
system.
| Task | Number of points
(success/failure) |
|---|
| Exit a circle of 30
cm diameter within 3 min | 0/1 |
| Paresis of upper
and/or lower limb of the contralateral side | 0/1 |
| Able to walk
straight | 0/1 |
| Presence of startle
reflex | 0/1 |
| Presence of seeking
behavior | 0/1 |
| Able to balance on
a beam of 7 mm width for 10 sec | 0/1 |
| Able to balance on
a round stick of 5 mm diameter for 10 sec | 0/1 |
| Able to cross a 30
cm long beam of 3 cm width | 0/1 |
| Able to cross a 30
cm long beam of 2 cm width | 0/1 |
| Able to cross a 30
cm long beam of 1 cm width | 0/1 |
| Maximum total | 10 |
Statistical analysis
Each experiment was repeated at least three times
and the data are expressed as the mean ± standard error of the
mean. For the behavioral assessment, two-way analysis of variance
was used followed by a Bonferroni post hoc test. For the other
assays, one-way analysis of variance was used followed by Tukey's
test. SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA) was used
for the statistical analyses. P<0.05 was considered to indicate
a statistically significant difference.
Results
Rapamycin improves the neurobehavioral
scores of mice following TBI
In the grip test, the scores of the
rapamycin-treated mice were significantly improved, compared with
those of the vehicle-treated mice 24 h and 72 h after TBI
(P<0.01 and P<0.05, respectively; Fig. 1A). As shown in Fig. 1B, the NSS scores of the TBI and the
TBI + DMSO groups were lower at 72 h than at 24 h. At 24 h
following TBI, the NSS scores of the rapamycin-treated mice were
significantly lower compared with those of the vehicle treated mice
(P<0.01), and at 72 h, the scores of the two groups had
improved, maintaining a significant difference (P<0.05).
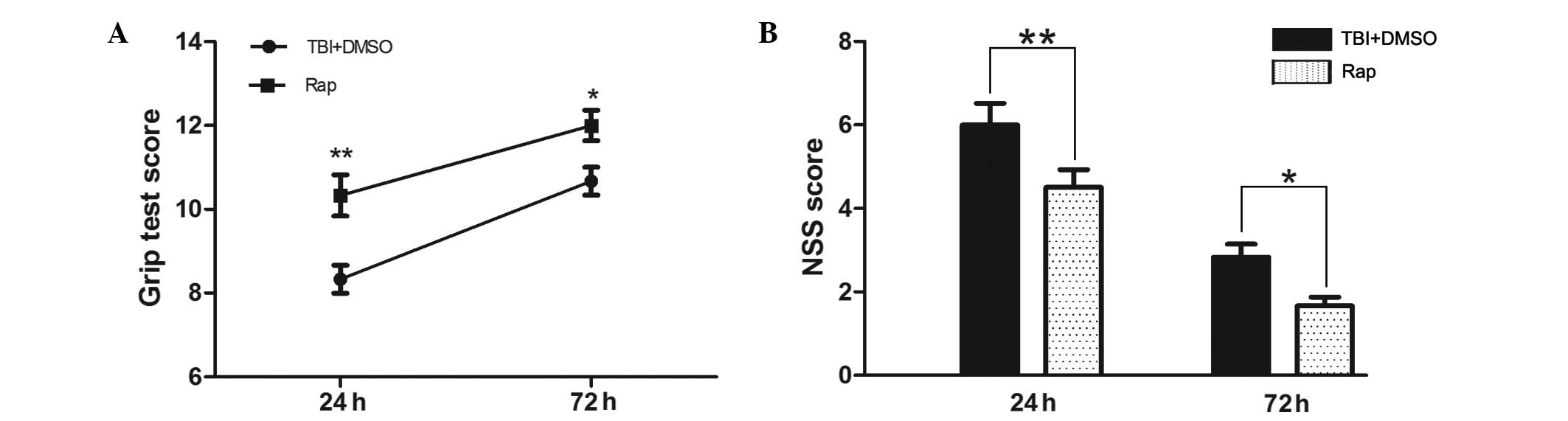 | Figure 1Rapamycin improves the neurobehavioral
performance of mice following TBI. At 24 h and 72 h following TBI,
the mice pretreated with rapamycin had significantly improved
neurological functions compared with the mice pretreated with DMSO,
as assessed by a (A) grip test and (B) NSS. Data are presented as
the mean ± standard error of the mean (n=8). **P<0.01
and *P<0.05, compared with the sham group,
nsP>0.05 compared with the TBI group, #P<0.05,
compared with the TBI + DMSO group. TBI, traumatic brain injury;
DMSO, dimethyl sulfoxide; NSS, neurological severity score; Rap,
rapamycin. |
Rapamycin protects neurons in the
pericontusional region from degeneration following TBI
Nissl staining was used to evaluate neuronal
survival in the pericontusional cortex 24 h after TBI (Fig. 2). Compared with the sham group, TBI
decreased the number of neurons in the pericontusional cortex
(P<0.01). A large proportion of neurons in the TBI group were
damaged, exhibiting extensive degenerative changes, which included
sparse cellular arrangements, loss of integrity, shrunken
cytoplasma and misshapen nuclei. The sham group was observed to
contain clear and intact neurons. Rapamycin significantly increased
the proportion of surviving neurons at 24 h after TBI (P<0.05)
and may, therefore, have alleviated the severity of neuronal
degeneration.
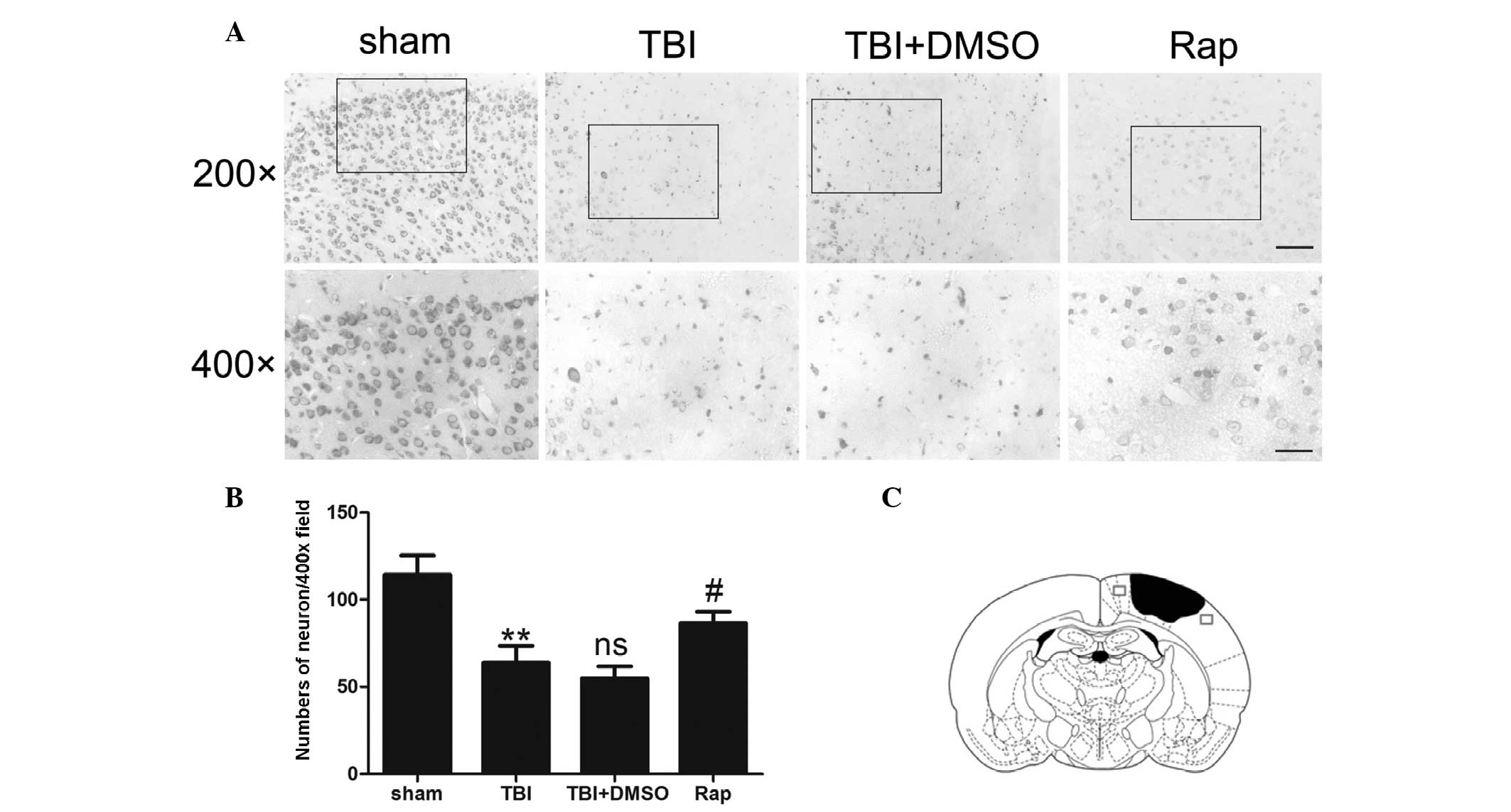 | Figure 2Rapamycin protects pericontusional
neurons from degeneration 24 h after TBI. (A) Representative
photomicrographs of Nissl staining of the pericontusional cortex in
the sham group, TBI, TBI + DMSO and TBI + rapamycin groups. Scale
bar=50 µm at magnification, ×200; scale bar=25 µm at
magnification, ×400. (B) Total neurons in each visual field
(magnification, ×400). Compared with the sham group, TBI decreased
the number of neurons in the pericontusional cortex. Rapamycin
treatment significantly increased the proportion of surviving
neurons 24 h after TBI. (C) Diagram of a coronal mouse brain
section, revealing the association between the lesion cavity (solid
black) and the regions where the images were captured (squares).
Data are expressed as the mean ± standard error of the mean (n=6).
**P<0.01, compared with sham group;
nsP>0.05, compared with TBI group;
#P<0.05, compared with TBI + DMSO group. TBI,
traumatic brain injury; Nissl, cresyl violet; DMSO, dimethyl
sulfoxide; Rap, rapamycin; ns, not significant. |
Rapamycin treatment decreases the
production of proinflammatory cytokines in the injured cortex
following TBI
To evaluate the association between rapamycin and
proinflammatory cytokines, an ELISA was used to detect the protein
levels of proinflammatory factors 24 h after TBI (Fig. 3). The expression levels of IL-1β
and TNF-α increased significantly following TBI compared with the
sham group (P<0.01 and P<0.001, respectively). No difference
was observed between the TBI + DMSO group and the TBI group
(P>0.05). The expression levels of IL-1β and TNF-α were reduced
in the rapamycin-treated groups compared with those in the
DMSO-treated group (P<0.05).
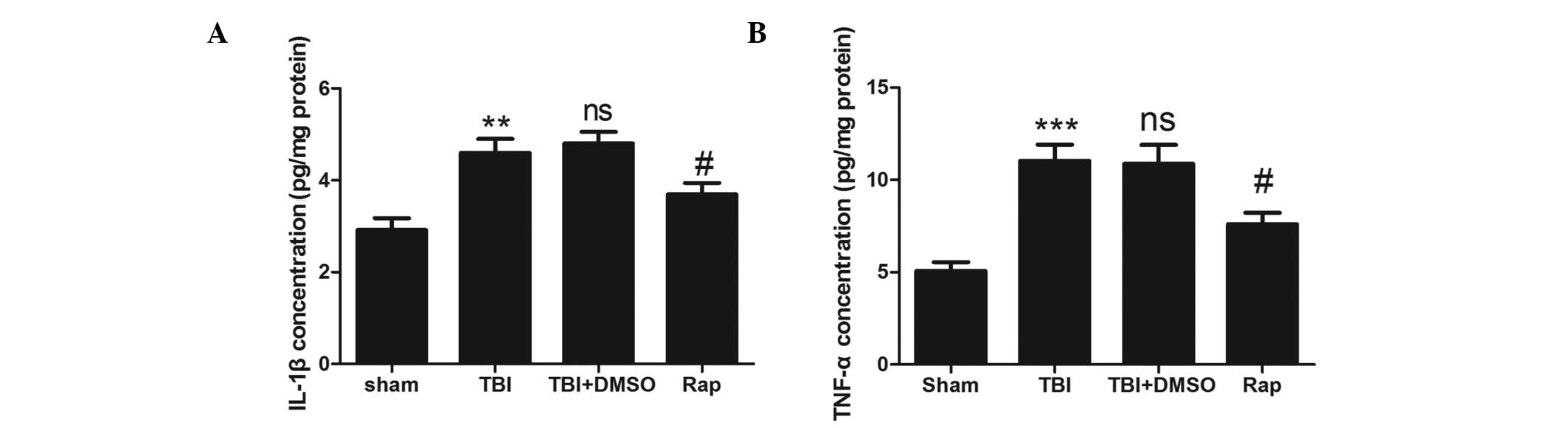 | Figure 3Rapamycin attenuates the upregulation
of proinflammatory cytokines in injured brain tissue following TBI.
(A) Concentration of IL-1β was increased significantly in the TBI
and TBI + DMSO groups compared with the sham group. Treatment with
rapamycin significantly attenuated the levels of IL-1β in the
ipsilateral cortex compared with DMSO treatment. (B) Concentration
if TNF-α was increased significantly in the TBI and TBI + DMSO
groups compared with the sham group. Treatment with rapamycin
significantly attenuated the levels of TNF-α in the ipsilateral
cortex compared with DMSO treatment. Data are expressed as the mean
± standard error of the mean (n=6). ***P<0.001 and
**P<0.01, compared with the sham group;
nsP>0.05, compared with the TBI group,
#P<0.05, compared with the TBI + DMSO group. TBI,
traumatic brain injury; DMSO, dimethyl sulfoxide; Rap, rapamycin;
IL, interleukin; TNF, tumor necrosis factor; ns, not
significant. |
Rapamycin suppresses microglial
activation in the pericontusional cortex following TBI
The levels of microglial activation were
investigated by detecting the immunofluorescence of IBA-1. As shown
in Fig. 4, the microglia in the
sham group exhibited long branching processes and small cellular
bodies. In response to brain injury, the number of microglia
increased significantly (P<0.01) compared with the sham group,
and the branches of the IBA-1-stained microglia became short,
retracted and thick, indicating activated microglia. Compared with
the TBI and TBI + DMSO groups, rapamycin injection decreased the
number of IBA-1-stained cells (P<0.05) and microglia exhibited
smaller cellular bodies.
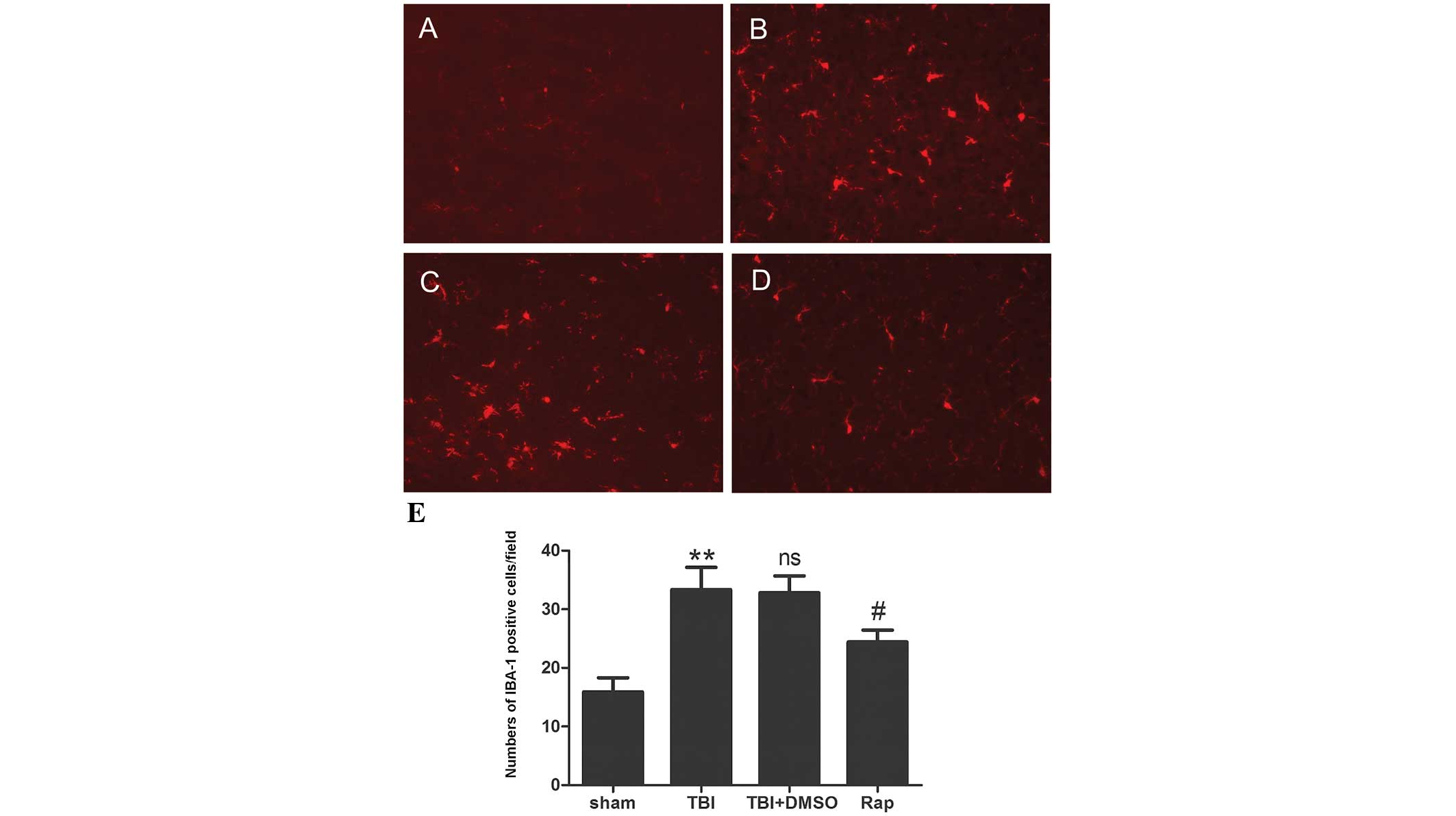 | Figure 4Rapamycin decreases microglial
activation 24 h following TBI. (A–D) Representative
photomicrographs of immunofluorescence staining for IBA-1 in the
pericontusive cortex. (A) Immunofluorescence indicated that
microglia of the sham group exhibited long branching processes and
a small cellular body. (B and C) In response to TBI, the number of
microglia increased and the branches of the microglia became short,
retracted and thick. (D) Rapamycin injection decreased the number
of IBA-1-stained cells, and the microglia exhibited a smaller
cellular body. Scale bar=50 µm. (E) Quantification of
immunofluorescence. Data are expressed as the mean ± standard error
of the mean (n=6). **P<0.01, compared with the sham
group; nsP>0.05, compared with the TBI group;
#P<0.05, compared with the TBI + DMSO group. TBI,
traumatic brain injury; IBA-1, ionized calcium-binding adapter
molecule 1DMSO, dimethyl sulfoxide; Rap, rapamycin; IBA-1, ionized
calcium-binding adapter molecule 1; ns, not significant. |
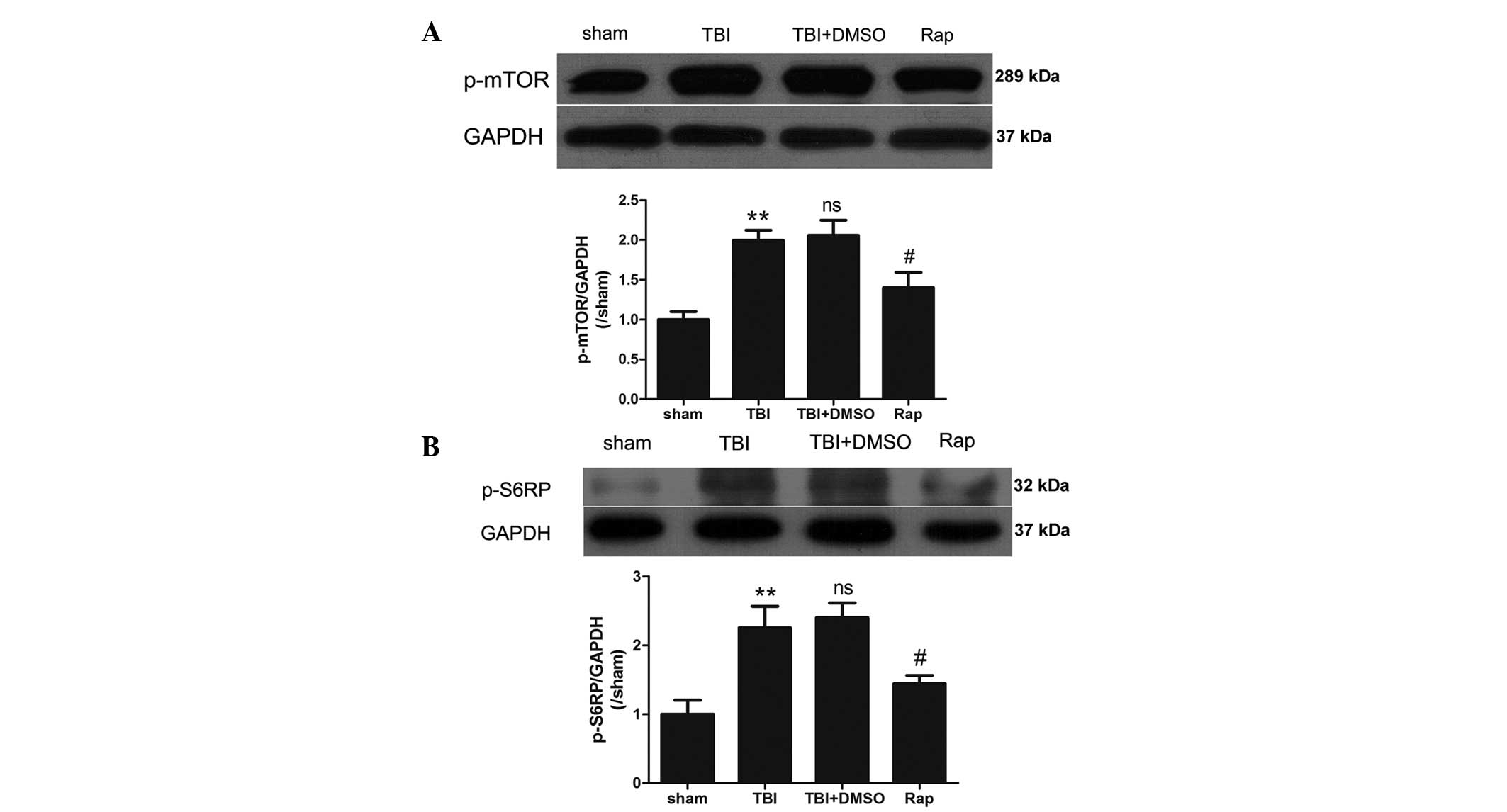
Rapamycin dephosphorylates the mTOR
pathway following TBI
To confirm the inactivation of the mTOR pathway
induced by rapamycin, the protein expression levels of
phosphorylated mTOR and S6RP were investigated using western
blotting. As shown in Fig. 5, the
expression levels of p-mTOR and p-S6RP were significantly increased
in the pericontusional cortex 24 h after TBI (P<0.01). No
statistically significant differences were identified between the
TBI group and the TBI + DMSO group (P>0.05). Compared with the
TBI + DMSO group, rapamycin administration markedly decreased the
expression levels of p-mTOR and p-S6RP (P<0.05).
Discussion
In the present study, the effects of rapamycin on
the upregulation of proinflammatory cytokines and the activation of
microglia were evaluated following TBI in a mouse model. The data
revealed that rapamycin administration significantly inhibited
microglial activation and decreased the expression levels of IL-1β
and TNF-α. Furthermore, rapamycin treatment increased the number of
neurons in the pericontusional cortex and improved neurological
function following TBI.
Neuroinflammation is important in the pathogenesis
of secondary brain injury following TBI (25,26).
Elevated levels of proinflammatory cytokines in the injured cortex,
including TNF-α and IL-1β, are considered to contribute to the
level of cerebral damage (27–30).
TNF-α has been observed as a major initiator of neuroinflammation
and is released early following an inflammatory stimulus (31). In inflammatory states, TNF-α, in
addition to a number of other proinflammatory mediators, is
produced predominantly by activated microglia (5). The contribution of TNF-α to tissue
damage has been assessed in animal models, whereby recombinant TNF
injected into the brain induced cerebral inflammation, breakdown of
the blood-brain barrier and the recruitment of intracranial
leukocytes (29). IL-1β is an
important initiator of the immune response, which is involved in
the onset and development of a complex inflammatory cascade.
Elevated levels of IL-1β have been detected in the cerebrospinal
fluid and brain tissue within the early hours following brain
injury (32), and neuronal damage
is attenuated when recombinant IL-1 receptor antagonist is injected
intracerebroventricularly following brain injury in rats (33). In the present study, the increased
number of neurons in the pericontusional cortex and the improved
neurological status following rapamycin administration may result
from the ability of rapamycin to decrease the expression levels of
IL-1β and TNF-α, thereby inhibiting the inflammatory reaction
following TBI.
As proinflammatory cytokines are predominantly
secreted by microglia, and activated microglia are important in the
progression of the inflammatory reaction following TBI, the effect
of rapamycin on microglial activation was further investigated. The
data revealed that rapamycin injection markedly suppressed the
levels of microglial activation following TBI. The activation of
microglia is widely accepted as histological evidence of brain
injury (34). Activated microglia
produce cytotoxic molecules, including nitric oxide, oxygen
radicals, arachidonic acid derivatives and cytokines (35), which induce microglial-mediated
neurotoxicity and aggravate the secondary injury following TBI.
Microglial activation leads to progressive and cumulative neuronal
cell loss (14), thereby inducing
irreversible neurological dysfunction. Therefore, in the present
study, the downregulated levels of proinflammatory cytokines may
have resulted from the inactivation of microglia, induced by
rapamycin.
The inner mechanisms of microglial activation
following treatment with various compounds have been reported in a
number of studies associated with microglial activation in
vitro (36–40). mTOR is key in microglial viability
(40) and the process of
microglial activation (39,40).
The mTOR pathway is an essential cellular signaling pathway
involved in a number of important physiological functions,
including cell growth, proliferation, metabolism, protein synthesis
and autophagy (41). Under basal
conditions, the inhibition of mTOR significantly reduces microglial
viability, and following activation of microglial cells using a
mixture of proinflammatory cytokines, mTOR inhibition reduces
microglial proliferation in a significant and dose-dependent manner
(40). In addition, the mTOR
pathway is important in the progression of protein synthesis. The
overexpression of p70S6K increases cell size (42), whereas p70S6K-knockout mice or the
inhibition of mTOR using rapamycin results in smaller cell sizes
(42,43). Thus, in the present study, the
activation of microglia following TBI may have resulted from cell
proliferation or enlargement, induced by phosphorylation and
activation of the mTOR pathway. Similar to previous studies, the
present data demonstrated that the mTOR pathway was important for
microglial activation, and inactivation of the mTOR pathway by
rapamycin resulted in a significant decrease in the activation of
microglia.
In conclusion, the present study demonstrated that
administration of rapamycin following TBI resulted in a
significantly decreased quantity of activated microglia in the
pericontusional cortex, suggesting that the mTOR pathway is
important in the process of microglial activation. However, there
were certain limitations of the present study, including the
limited investigation of the mechanism underlying the activation if
microglia by mTOR, short-term and single time-point observationa
following TBI, and the lack of investigation of the long-term
effects of rapamycin injection. Therefore, the long-term
consequences of rapamycin treatment following TBI requires further
investigation.
Acknowledgments
The present study was supported by the Project
Planning of Nanjing General Hospital (grant no. 2013067).
References
|
1
|
Cernak I: Animal models of head trauma.
NeuroRx. 2:410–422. 2005. View Article : Google Scholar
|
|
2
|
Yatsiv I, Grigoriadis N, Simeonidou C, et
al: Erythropoietin is neuroprotective, improves functional recovery
and reduces neuronal apoptosis and inflammation in a rodent model
of experimental closed head injury. FASEB J. 19:1701–1703.
2005.PubMed/NCBI
|
|
3
|
Xu J, Wang H, Ding K, et al: Inhibition of
cathepsin S produces neuroprotective effects after traumatic brain
injury in mice. Mediators Inflamm. 2013:1878732013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cornelius C, Crupi R, Calabrese V, et al:
Traumatic brain injury: oxidative stress and neuroprotection.
Antioxid Redox Signal. 19:836–853. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ziebell JM and Morganti-Kossmann MC:
Involvement of pro- and anti-inflammatory cytokines and chemokines
in the pathophysiology of traumatic brain injury.
Neurotherapeutics. 7:22–30. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Venkatesan C, Chrzaszcz M, Choi N and
Wainwright MS: Chronic upregulation of activated microglia
immunoreactive for galectin-3/Mac-2 and nerve growth factor
following diffuse axonal injury. J Neuroinflammation. 7:322010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cao T, Thomas TC, Ziebell JM, Pauly JR and
Lifshitz J: Morphological and genetic activation of microglia after
diffuse traumatic brain injury in the rat. Neuroscience. 225:65–75.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kelley BJ, Lifshitz J and Povlishock JT:
Neuroinflammatory responses after experimental diffuse traumatic
brain injury. J Neuropathol Exp Neurol. 66:989–1001. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang R, Liu Y, Yan K, et al:
Anti-inflammatory and immunomodulatory mechanisms of mesenchymal
stem cell transplantation in experimental traumatic brain injury. J
Neuroinflammation. 10:1062013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ng SY, Semple BD, Morganti-Kossmann MC and
Bye N: Attenuation of microglial activation with minocycline is not
associated with changes in neurogenesis after focal traumatic brain
injury in adult mice. J Neurotrauma. 29:1410–1425. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
D'Avila JC, Lam TI, Bingham D, et al:
Microglial activation induced by brain trauma is suppressed by
post-injury treatment with a PARP inhibitor. J Neuroinflammation.
9:312012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dazert E and Hall MN: mTOR signaling in
disease. Curr Opin Cell Biol. 23:744–755. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Reynolds TH IV, Bodine SC and Lawrence JC
Jr: Control of Ser2448 phosphorylation in the mammalian target of
rapamycin by insulin and skeletal muscle load. J Biol Chem.
277:17657–17662. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chong ZZ, Shang YC, Zhang L, Wang S and
Maiese K: Mammalian target of rapamycin: hitting the bull's-eye for
neurological disorders. Oxid Med Cell Longev. 3:374–391. 2010.
View Article : Google Scholar
|
|
15
|
Park J, Zhang J, Qiu J, et al: Combination
therapy targeting Akt and mammalian target of rapamycin improves
functional outcome after controlled cortical impact in mice. J
Cereb Blood Flow Metab. 32:330–340. 2012. View Article : Google Scholar :
|
|
16
|
Fan W, Cheng K, Qin X, et al: mTORC1 and
mTORC2 play different roles in the functional survival of
transplanted adipose-derived stromal cells in hind limb ischemic
mice via regulating inflammation in vivo. Stem Cells. 31:203–214.
2013. View Article : Google Scholar
|
|
17
|
Vignot S, Faivre S, Aguirre D and Raymond
E: mTOR-targeted therapy of cancer with rapamycin derivatives. Ann
Oncol. 16:525–537. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hosoi H, Dilling MB, Shikata T, et al:
Rapamycin causes poorly reversible inhibition of mTOR and induces
p53-independent apoptosis in human rhabdomyosarcoma cells. Cancer
Res. 59:886–894. 1999.PubMed/NCBI
|
|
19
|
Erlich S, Alexandrovich A, Shohami E and
Pinkas-Kramarski R: Rapamycin is a neuroprotective treatment for
traumatic brain injury. Neurobiol Dis. 26:86–93. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Flierl MA, Stahel PF, Beauchamp KM, Morgan
SJ, Smith WR and Shohami E: Mouse closed head injury model induced
by a weight-drop device. Nat Protoc. 4:1328–1337. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ding K, Wang H, Wu Y, Zhang L, Xu J, Li T,
Ding Y, Zhu L and He J: Rapamycin protects against apoptotic
neuronal death and improves neurologic function after traumatic
brain injury in mice via modulation of the mTOR-p53-Bax axis. J
Surg Res. 194:239–247. 2015. View Article : Google Scholar
|
|
22
|
Wang J and Doré S: Heme oxygenase-1
exacerbates early brain injury after intracerebral haemorrhage.
Brain. 130:1643–1652. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhuang Z, Zhou Ml, You WC, et al:
Hydrogen-rich saline alleviates early brain injury via reducing
oxidative stress and brain edema following experimental
subarachnoid hemorrhage in rabbits. BMC Neurosci. 13:472012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bermpohl D, You Z, Korsmeyer SJ, Moskowitz
MA and Whalen MJ: Traumatic brain injury in mice deficient in Bid:
effects on histopathology and functional outcome. J Cereb Blood
Flow Metab. 26:625–633. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kumar A and Loane DJ: Neuroinflammation
after traumatic brain injury: opportunities for therapeutic
intervention. Brain Behav Immun. 26:1191–1201. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lu J, Goh SJ, Tng PY, Deng YY, Ling EA and
Moochhala S: Systemic inflammatory response following acute
traumatic brain injury. Front Biosci (Landmark Ed). 14:3795–3813.
2009. View Article : Google Scholar
|
|
27
|
Rothwell NJ: Annual review prize lecture
cytokines-killers in the brain? J Physiol. 514:3–17. 1999.
View Article : Google Scholar
|
|
28
|
Ramilo O, Saez-Llorens X, Mertsola J, et
al: Tumor necrosis factor alpha/cachectin and interleukin 1 beta
initiate meningeal inflammation. J Exp Med. 172:497–507. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim KS, Wass CA, Cross AS and Opal SM:
Modulation of blood-brain barrier permeability by tumor necrosis
factor and antibody to tumor necrosis factor in the rat. Lymphokine
Cytokine Res. 11:293–298. 1992.PubMed/NCBI
|
|
30
|
Smith RM and Giannoudis PV: Trauma and the
immune response. J R Soc Med. 91:417–420. 1998.PubMed/NCBI
|
|
31
|
Hesse DG, Tracey KJ, Fong Y, et al:
Cytokine appearance in human endotoxemia and primate bacteremia.
Surg Gynecol Obstet. 166:147–153. 1988.PubMed/NCBI
|
|
32
|
Winter CD, Iannotti F, Pringle AK, Trikkas
C, Clough GF and Church MK: A microdialysis method for the recovery
of IL-1beta, IL-6 and nerve growth factor from human brain in vivo.
J Neurosci Methods. 119:45–50. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Loddick SA and Rothwell NJ:
Neuroprotective effects of human recombinant interleukin-1 receptor
antagonist in focal cerebral ischaemia in the rat. J Cereb Blood
Flow Metab. 16:932–940. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Morganti-Kossmann MC, Satgunaseelan L, Bye
N and Kossmann T: Modulation of immune response by head injury.
Injury. 38:1392–1400. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Min KJ, Jou I and Joe E:
Plasminogen-induced IL-1beta and TNF-alpha production in microglia
is regulated by reactive oxygen species. Biochem Biophys Res
Commun. 312:969–974. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dean JM, Wang X, Kaindl AM, et al:
Microglial MyD88 signaling regulates acute neuronal toxicity of
LPS-stimulated microglia in vitro. Brain Behav Immun. 24:776–783.
2010. View Article : Google Scholar
|
|
37
|
Pinteaux-Jones F, Sevastou IG, Fry VA,
Heales S, Baker D and Pocock JM: Myelin-induced microglial
neurotoxicity can be controlled by microglial metabotropic
glutamate receptors. J Neurochem. 106:442–454. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pais TF, Figueiredo C, Peixoto R, Braz MH
and Chatterjee S: Necrotic neurons enhance microglial neurotoxicity
through induction of glutaminase by a MyD88-dependent pathway. J
Neuroinflammation. 5:432008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dello RC, Lisi L, Feinstein DL and Navarra
P: mTOR kinase, a key player in the regulation of glial functions:
relevance for the therapy of multiple sclerosis. Glia. 61:301–311.
2013. View Article : Google Scholar
|
|
40
|
Dello Russo C, Lisi L, Tringali G and
Navarra P: Involvement of mTOR kinase in cytokine-dependent
microglial activation and cell proliferation. Biochem Pharmacol.
78:1242–1251. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wong M: Mammalian target of rapamycin
(mTOR) pathways in neurological diseases. Biomed J. 36:40–50. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fingar DC, Salama S, Tsou C, Harlow E and
Blenis J: Mammalian cell size is controlled by mTOR and its
downstream targets S6K1 and 4EBP1/eIF4E. Genes Dev. 16:1472–1487.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shima H, Pende M, Chen Y, Fumagalli S,
Thomas G and Kozma SC: Disruption of the p70(s6k)/p85(s6k) gene
reveals a small mouse phenotype and a new functional S6 kinase.
EMBO J. 17:6649–6659. 1998. View Article : Google Scholar : PubMed/NCBI
|