Introduction
Lung cancer is the life-threatening disease and is
the leading cause of mortality among malignant tumors worldwide
(1). Of all lung cancer cases,
~85% develop into non-small cell lung cancer (NSCLC), with a 5-year
survival rate that remains low at 18%, despite developments in
systemic therapy, including surgery, radiotherapy and chemotherapy,
in the previous decades (2). Based
on clinical observations, the lack of effective treatment for
relapse and invasion of lung cancer has become a major obstacle in
improving survival rates (3).
Accumulative data have demonstrated that the effort of controlling
the metastasis-associated pathways may delay the progression of
NSCLC (4–6). Therefore, it is necessary to identify
effective and safe drugs to treat NSCLC invasion and
metastasis.
It has been reported that Chinese medicine benefits
patients with NSCLC via increasing therapeutic effects and reducing
radiotherapy toxicity in clinical studies (7–9).
Calycosin (Cal) is a purified isoflavone with a defined chemical
structure (Fig. 1A) and is
isolated from the traditional Chinese herbs, Astragalus
membranaceus (Fisch.) Bge. or Astragalus membranaceus
(Fisch.) Bge. var. mongholicus (Bge.) Hsiao (10). Cal has been reported to have
various pharmacologic effects with antitumor, neuroprotective and
anti-inflammatory properties (11–14).
Previous studies have demonstrated that Cal inhibits cancer growth
via apoptosis in 143B osteosarcoma cells and MCF-7 breast cancer
cells (15,16). However, the antitumor activities of
Cal on NSCLC metastasis and invasion, and the underlying mechanism
remains to be elucidated. Therefore, the present study examined the
A549 human lung adenocarcinoma cell line to further understand the
effect of Cal on the migration and invasion of these cells.
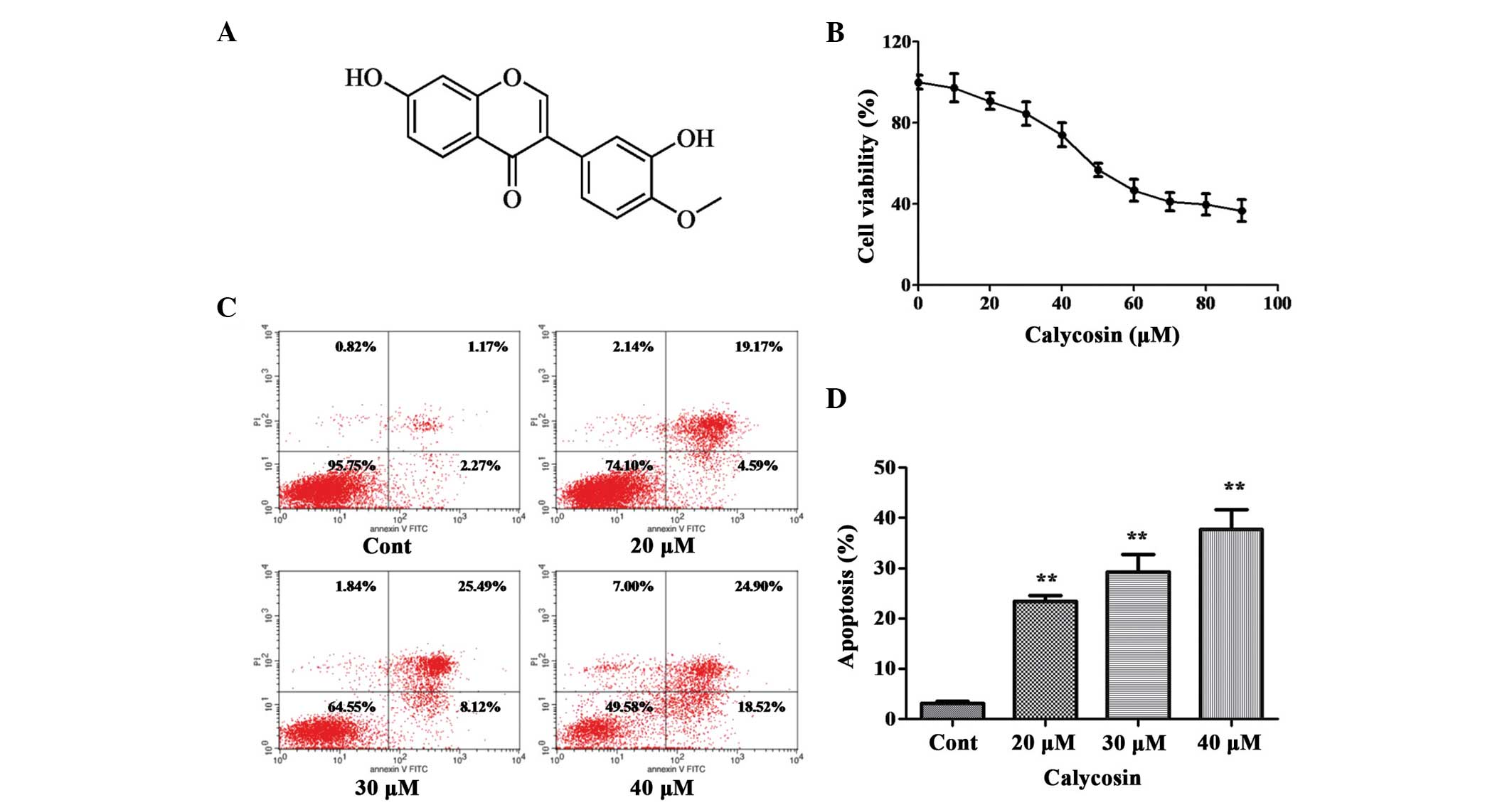 | Figure 1Effect of Cal on the proliferation
and apoptosis of A549 cells. (A) Chemical structure of Cal. (B)
A549 cells were treated with Cal at various concentrations (0, 10,
20, 30, 40, 50, 60, 70, 80 and 90 µM) for 24 h, and cell
viability was measured using an MTT assay. The results are
expressed as a percentage of the control and presented as the mean
± standard deviation (n=6). (C) A549 cells were treated with Cal
(20, 30 and 40 µM) and incubated for 24 h. The control group
received the same volume of dimethyl sulfoxide. Apoptotic cells
were detected using Annexin V and PI staining. (D) Apoptotic rate
obtained from three independent experiments and presented the mean
± standard deviation (n=3). **P≤0.01, vs. control. Cal,
calycosin; Cont, control; PI, propidium iodide; FITC, fluorescein
isothiocyanate. |
PKC-α is a member of the PKC family, which can be
activated by calcium and the second messenger, diacylglycerol
(17). PKC-α is involved in
diverse cellular signaling pathways and cellular functions,
including cell proliferation, differentiation, apoptosis and
survival (18). PKC-α serves as a
major receptor for phorbol-12-myristate-13-acetate (TPA), a class
of tumor promoter (19). The
overexpression of PKC-α, induced by TPA, promotes the migration and
invasion of GBM8401 glioma cells (20). Previous studies have demonstrated
that inhibition of the expression of PKC-α decreases hepatocellular
carcinoma cell invasion and breast cancer metastasis (21,22).
The results of these studies indicate that there is a correlation
between PKC-α and cancer cell metastasis. However, whether PKC-α is
involved in human lung cancer cells remains to be elucidated.
Therefore, the present study investigated whether PKC-α is involved
in the inhibition of A549 cell migration and invasion by Cal.
Extracellular signal-regulated protein kinase
(ERK)1/2, one of the important members of the mitogen-activated
protein kinase (MAPK) family, is an essential signaling pathway by
which cell survival, differentiation, apoptosis, proliferation,
migration and invasion are regulated (23). Previous studies have reported that
activated ERK1/2 signaling due to phosphorylation induces cancer
cell proliferation, migration and invasion (24). Studies have demonstrated that Cal
interacts with estrogen receptors on the cell membrane and
modulates the MAPK signaling pathway to inhibit growth and induce
apoptosis of ER-positive breast cancer cells (25,26).
It has also been demonstrated that the phosphorylation of ERK1/2
contributes to lung cancer migration and invasion (27). Therefore, the present study also
aimed to investigate whether Cal inhibits lung cancer migration and
invasion by regulating the ERK1/2 signal pathway, as well as its
downstream biomarkers.
The present study aimed to investigate the
antimetastic activity of Cal on A549 cells by assessing migration
and invasion abilities and the expression levels of potential
pathway proteins. The results may reveal the possible mechanisms
underlying the effects of Cal on A549 cells and may provide a basis
for the safety and anticancer efficacy of Cal in further clinical
applications.
Materials and methods
Chemicals and reagents
Cal (C16H12O5) was
obtained from Standard Biotech Co., Ltd (Shanghai, China). The A549
human lung adenocarcinoma cell line was purchased from American
Type Culture Collection (Manassas, VA, USA). TPA and bovine serum
albumin (BSA) were provided by Sigma-Aldrich (St. Louis, MO, USA).
Transwell chambers were purchased from Corning Incorporated
(Corning, NY, USA). MMP2, MMP-9, E-cadherin (E-Cad), integrin β1,
PKC-α antibodies were obtained from Boster Systems, Inc.
(Pleasanton, CA, USA). AEB071, a PKC-α inhibitor, was provided by
Sellek Chemicals (Houston, TX, USA). The ERK1/2 inhibitor
(PD98059), penicillin/streptomycin, trypsin, EDTA, RNase, 1% Triton
X-100, SDS-PAGE gel (10%) and polyvinylidene difluoride (PVDF)
membranes were purchased from Beyotime Institute of Biotechnology
(Jiangsu, China). AnnexinV/ propidium iodide (PI) was provided by
Immunotech (Marseille, France). Basal Dulbecco's modified Eagle's
medium (DMEM) and fetal bovine serum (FBS) were purchased from
Gibco Life Technologies (Carlsbad, CA, USA). Crystal violet, acetic
acid and pure methanol were purchased from Aladdin Shanghai
Biochemical Technology Co., Ltd.(Shanghai, China). Other reagents
used were of analytical grade and obtained from commercial
sources.
Cell culture
DMEM supplemented with 10% FBS, 1% nonessential
amino acids, 100 U/ml penicillin and 100 µg/ml streptomycin
were used as the medium for A549 cell cultivation. The medium was
replaced every 2 days. The incubation conditions were maintained at
37°C with a humidified atmosphere of 5% CO2.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT)
assay for cell viability
The cell viability inhibiting effect of Cal on the
A549 cells was determined using an MTT assay. Briefly, the
logarithmic growth phase A549 cells were digested in 0.1% trypsin,
DMEM supplemented with 10% FBS was added, and the cells were plated
in 96-well plates at a final density of 0.6×104
cells/well. The cells were treated with a series of diluted
concentrations of Cal (10, 20, 30, 40, 50, 60, 70, 80 or 90
µM) and incubated at 37°C for 24 h. Following incubation,
MTT (5 mg/ml, 10 µl) solution was added to each well and
incubated for 4 h at 37°C. Subsequently, the supernatant in each
well was discarded and dimethylsulfoxide (DMSO; 100 µl) was
added. The optical density (OD) value at 570 nm was measured using
a Spectra Max 190 microplate reader (Molecular Devices, Sunnyvale,
CA, USA). Cell viability was determined by the OD value, and was
calculated as the percentage of viable cells. The measurement was
performed in three independent experiments.
Flow cytometric analysis
The A549 cells (6×105/well) were seeded
into 6-well plates and treated with different concentration of Cal
(20, 30 and 40 µM), which were then cultured at 37°C for 24
h. The cells were collected and digested with 0.25% trypsin and
0.02% EDTA (1:1) at 37°C for 3-4 min. The cells were then pipetted
gently and collected and centrifuged at 112 g at room temperature
for 5 min. Subsequently the cells were washed with cold
phosphate-buffered saline (PBS; 0.01 M; pH 7.4) twice and
resuspended in the residual PBS. Following the addition of 1 ml
pre-chilled (−20°C) 80% ethanol, the cells were stored at −20°C
overnight. Following washing twice with PBS, 60–80 µl RNAase
(1 mg/ml) was added, and the cells incubated at 37°C for 30 min.
Following chilling on ice for 2 min, Annexin V-fluorescein
isothiocyanate (FITC)/PI solution (100 mg/l PI, 0.1% TritonX-100)
was added, and the sample was incubated in the dark at room
temperature for 30 min. Cell apoptosis was analyzed using an FC 500
flow cytometer (Beckman Coulter, Brea, CA, USA). Data were acquired
by the RXP software of the machine.
Adhesion analysis
A cell adhesion experiment was performed in 96-well
plates coated with Matrigel (BD Biosciences, Franklin Lakes, NJ,
USA) and air dried in a Logic+ laminar hood (Labconco,
Kansas City, MO, USA) overnight. The wells were blocked with 2% BSA
(50 µl/well) and incubated at 37°C for 2 h. The A549 cells
were stimulated with 80 nM TPA, as described previously (28). Subsequently, the cells were treated
with different concentrations of Cal (20, 30 and 40 µM) for
24 h. Then cells were inoculated into the 96-well plate at a
concentration of 1×104 cells/well and incubated at 37°C
for 30 min. The non-adherent cells were removed with 200 µl
PBS (pH 7.4) following incubation. Subsequently, the adherent cells
in each well were stained with 0.1% crystal violet and lysed with
30% acetic acid, and measured optical density at 550 nm on the
Spectra Max 190 microplate reader. The adhesion rate was calculated
from the OD values of triplicate experiments.
Wound healing analysis
A scratch assay was performed, as described
previously. Briefly, the A549 cells were detached using 0.1%
trypsin and resuspended in serum-free DMEM, seeded at a
concentration of 5×106 cells/cm2 into 6-well
plates in the medium containing 10% FBS. Following overnight
incubation at 37°C, a cell-monolayer was yielded for a wound
healing assay. To introduce the wound, three wound tracks were
scored in the monolayer, ~5 mm in distance, in each well using a
200 µl pipette tip. The suspended cells were washed twice
with DMEM, and the wounded cell monolayer was incubated in FBS-free
medium containing different concentrations of Cal (20, 30 and 40
µM), stimulated with 80 nM TPA, at 37°C for 24 h. Images of
the wound area were captured at 0 and 24 h using an IX73 microscope
(Olympus Corporation, Tokyo, Japan). The areas in the scratch
wound, which were not covered in cells were quantified using ImageJ
2.1.4.7 software (Media Cybernetics, Inc., Rockville, MD, USA). The
closure rate was determined as the percentage of the area at 0 h.
The experiments were performed in triplicate.
Cell migration analysis
To determine cell migration, experiments were
performed using Transwell chambers. The A549 cells were stimulated
with TPA and incubated in the presence or absence of various
concentrations of Cal (20, 30 and 40 µM) for 24 h. Following
incubation, the cells were detached using trypsin and resuspended
in serum-free medium. Medium containing 10% FBS was added to the
lower chamber as a chemotactic factor, and the cells were seeded in
the upper chamber at a density of 1×104 cells/well in 50
µl serum-free medium. Following incubation for 8 h at 37°C,
any A549 cells, which did not penetrate the polycarbonate membrane
were removed using cotton swabs. The cells, which had penetrated
through membrane were fixed with methanol and stained with 0.1%
crystal violet for 10 min. The chambers were visualized in six
randomly-selected visual fields under an IX73 microscope, in which
the number of cells were counted. Each experiment was performed in
triplicate.
Cell invasion analysis
To determine cell invasion, experiments were
performed in Transwell chambers coated with Matrigel, as described
previously (29). Briefly, 50
µl Matrigel was coated on the membrane at the base of the
Transwell chamber and air dried in a Logic+ laminar hood
(Labconco) overnight. Following blocking with 2% BSA (50
µl/well), the chambers were incubated at 37°C for 2 h and
were then rinsed with PBS. The cells, which had been exposed to
different concentration Cal (20, 30 and 40 µM) were placed
into the upper layer of the Transwell chamber at a concentration of
2×104 cells/well. The medium (600 µl/well)
containing chemotactic factor (10% FBS) was added to the lower
layer of the Transwell chamber. The cells were cultured at 37°C for
24 h. Cotton swabs were used to remove the cells, which did not
penetrate the polycarbonate membrane. The cells, which penetrated
through and adhered to the membrane were fixed with methanol and
stained with 0.1% crystal violet for 10 min. Following the crystal
violet staining, the cells were rinsed with distilled water to
removing excess dye. Subsequently six randomly-selected visual
fields in each well were selected and visualized under an IX73
microscope. The number of cells that penetrated the membrane were
counted, and the invasion rate was quantified by the number of
permeated cells associated with the OD value. Each experiment was
repeated three times.
Western blot analysis
To analyze the expression levels of
migration-associated proteins in the A549 cells affected by Cal,
western blot analysis was performed. Briefly, analyses were
performed following stimulation with TPA and treatment with Cal
(20, 30 or 40 µM), and with or without the PKC inhibitor
(AEB071) or ERK1/2 inhibitor (PD98059), following TPA stimulation
and Cal (30 µM) treatment. The A549 cells were suspended in
250 µl lysis buffer, containing 25 mM Tris-HCl (pH 7.6), 150
mM NaCl, 1% NP-40, 0.1% SDS, 1% sodium deoxycholate and protease
inhibitors (Thermo Fisher Scientific, Waltham, MA, USA). The cell
lysate was centrifuged at 10,000 g for 20 min at 4°C. Equal
quantities of proteins from each sample (50 µg) were
subjected to 10% sodiumdodecyl sulfate (SDS)-polyacrylamide gel
electrophoresis. The proteins were then transferred onto PVDF
membranes and the membranes were blocked with 5% (w/v) BSA for 2 h
and washed in Tris-buffered saline with Tween 20 (TBST) three
times. Subsequently, the membranes were incubated with the
following primary detection antibodies: E-Cad, integrin β1, MMP-2,
MMP-9, PKC-α, ERK1/2 and phosphorylated (p)-ERK1/2 (1:1,000)
overnight at 4°C. The membranes were then washed and incubated with
horseradish-peroxidase-conjugated IgG for 1 h at room temperature
and then washed in TBST three times. Chemiluminescence reagents of
western blotting were added for visualization of the protein bands,
and quantification of the proteins bands was performed using ImageJ
software.
Statistical analysis
All data in the present study were obtained from
three independent experiments and are expressed as the mean ±
standard deviation. One-way analysis of variance was used for
multiple comparisons and Student's t test was used to
evaluate the differences between two groups. All analyses were
performed using SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Cal inhibits the viability of A549
cells
The effect of Cal on cell viability was assessed
using an MTT assay. The A549 cells were treated with increasing
doses (0–90 µM) of Cal for 24 h. As shown in Fig. 1B, following exposure to Cal, the
viability of A549 cells decreased in a dose-dependent manner. No
significant change in cell viability were observed, compared with
the 0 µM (DMSO treatment only) group, following 24 h
treatment with Cal at concentration between 0 and 40 µM,
indicating that Cal was not toxic to the A549 cells at these
concentrations. Following treatment with Cal at concentrations
>40 µM, cell viability reduced significantly at 24 h.
These results indicated that treatment with Cal at doses >50
µM for 24 h resulted in the dose-dependent loss of cell
viability in the A549 cells, however, doses <40 µM for 24
h did not cause cytotoxicity. Therefore, concentrations of
Cal<40 µM was selected for the subsequent
experiments.
Effect of Cal on cell apoptosis
To understand whether the effect of Cal on A549 cell
proliferation had any association with apoptotic rates, the binding
of Annexin V to phosphatidylserine, exposed on the cell membrane,
was measured, which is generally recognized as an early indicator
of apoptosis. As shown in Fig. 1C and
D, the total percentages of Annexin V+/PI-cells (right lower
quadrant representing early apoptosis) and Annexin V+/PI+ cells
(right upper quadrant representing late apoptosis and necrosis)
increased between 23.39 and 43.77% following treatment of A549
cells with Cal at 20, 30 and 40 µM for 24 h, compared with
3.44% apoptosis in the control group. These data indicated that Cal
induced A549 cell apoptosis in a dose-dependent manner, which was
associated with the inhibition of proliferation.
Cal suppresses A549 cell adhesion induced
by TPA
To investigate the inhibition of Cal on TPA-treated
A549 cell adhesion, a cell matrix adhesion assay was performed. As
shown in (Fig. 2A), following
treatment with Cal at concentrations of 20, 30 and 40 µM,
the cell adhesion rates of the A549 cells were 86.58, 75.40 and
62.38% of that in the TPA-induced group, respectively (P<0.01).
These data suggested that Cal inhibited the adhesion ability of the
A549 cells to the cell matrix.
 | Figure 2Effect of Cal on the adhesion,
migration and invasion of TPA-induced A549 cells. The A549 cells
were treated with 0, 20, 30 or 40 µM Cal, in the presence or
absence of TPA (80 nM) for 24 h, and were analyzed for (A) adherent
ability and (B) wound healing. (C) Migration ability was determined
by the closure rate of migrating cells at 24 h, vs. 0 h. (D) A549
cells were inoculated in Transwell chambers treated with Cal for 10
h to assess migration with an IX73 microscope and crystal violet
staining (magnification, ×100). (E) Permeated cells, compared with
the TPA-induced group. (F) A549 cells were inoculated into
Matrigel-coated Transwell chambers and treated with Cal for 24 h to
assess cell invasiveness with an IX73 microscope following staining
with crystal violet (magnification, ×100). (G) Rate of cell
invasion through the membrane, compared with the TPA-induced group.
The results were obtained from triplicate experiments and are
presented the mean ± standard deviation (n=3). *P≤0.05,
vs. control; **P≤0.01 vs. control. Cal, calycosin; TPA,
phorbol-12-myristate-13-acetate. |
Cal inhibits A549 cell migration induced
by TPA
The effect of Cal on TPA-induced A549 cell migration
capability was estimated by a wound-healing assay and Transwell
chamber assay. In the wound-healing assay (Fig. 2B and C), the cells treated with TPA
covered 49.91% of the wound area, which was significantly higher
than the untreated cells following incubation (13.31%). The wound
closure rates were 41.81, 36.62 and 22.98% following treatment with
Cal at 20, 30 and 40 µM, respectively, which were
significantly lower than that of the TPA-treated group
(P<0.05).
Following 8 h Transwell chamber migration, the
percentage of A549 cells that penetrated the membrane in the
non-TPA-stimulated group was only 28%. The percentages of cells
that penetrated through the membrane following TPA treatment were
significantly decreased when exposed to Cal concentrations of at
20, 30 and 40 µM to 72.46, 52.46 and 48.23%, respectively,
compared with the TPA-treated only group (Fig. 2D and E; P<0.01).
Cal suppresses TPA-induced A549 cell
invasion
A Transwell coated with Matrigel was used to
determine the suppression of Cal on A549 cell invasion. As shown in
Fig. 2F and G, the relative
percentages of penetrated cells in the non-induced cell group
increased between 26.69 and 100% when exposed to TPA. This results
indicated that the invasion capability of the A549 cells induced by
TPA was increased significantly, compared with control group.
Compared with the TPA-treated only group, the increased number of
penetrated cells innduced by TPA was significantly suppressed by
Cal at concentrations of 20, 30 and 40 µM (62.72, 52.99 and
41.70%, respectively) in a dose-dependent manner. (P<0.05).
Cal reduces the mobility of A549 cells,
suppresses the expression of integrin β1 and increases the
expression of E-cad
The increase in tumor cell mobility is important in
the metastasis processes. The expression levels of E-Cad and
integrin β1, associated with cell migration, were detected in the
present study using western blot analysis As shown in Fig. 3A and B, the relative expression
level of E-Cad decreased between 221 and 100% when treated with
TPA. However, the expression level was increased to 106.00, 137.09
and 185.94% following exposure to different concentrations of Cal
for 24 h. The relative activity of integrin β1 activitiy was
increased between 57.34 and 100% following stimulation with TPA.
Cal inhibited this TPA-induced integrin β1 activity to 84.50, 71.04
and 61.28% following exposure to 20, 30 and 40 µM Cal for 24
h. These results indicated that Cal suppressed the migration
ability of the A549 cells by regulating the expression of E-Cad and
reducing the expression of integrin β1.
 | Figure 3Effect of Cal on the expression
levels of PKC-α, p-ERK1/2, E-Cad, integrin β1, MMP-2 and MMP-9. (A)
A549 cells were treated with various concentrations (0, 20, 30 and
40 µM) of Cal in the presence or absence of TPA (80 nM) for
24 h, and then subjected to western blotting to analyze the protein
levels of E-cad and integrin β1. (B) Quantification of the protein
level of E-cad and integrin β1. (C) A549 cells were treated with
various concentrations (0, 20, 30 and 40 µM) of Cal in the
presence or absence of TPA (80 nM) for 24 h, and then subjected to
western blotting to analyze the protein levels of MMP-2 and MMP-9.
(D) Quantification of the protein level of MMP-2 and MMP-9. (E)
A549 cells were treated with various concentrations (0, 20, 30 and
40 µM) of Cal in the presence or absence of TPA (80 nM) for
24 h, and then subjected to western blotting to analyze the protein
levels of PKC-α. (F) Quantification of the protein level of PKC-α.
(G) A549 cells were treated with various concentrations (0, 20, 30
and 40 µM) of Cal in the presence or absence of TPA (80 nM)
for 24 h, and then subjected to western blotting to analyze the
protein levels of p-ERK1/2 and ERK1/2 (H) Quantification of the
proteins level of p-ERK1/2 and ERK1/2. Values are presented as the
mean ± standard deviation of three independent experiments,
performed in triplicate. *P≤0.05 and
**P≤0.01, vs. TPA-induced group. Cal, calycosin; TPA,
phorbol-12-myristate-13-acetate; PCK, protein kinase C; p-ERK,
phosphorylated extracellular signal-regulated kinase; E-Cad,
E-cadherin; MMP, matrix metalloproteinase. |
Cal inhibits extracellular matrix (ECM)
degradation by suppressing the levels of MMP-2 and MMP-9
ECM degradation is crucial for tumor cell invasion,
suggesting that MMPs are required. To clarify whether MMP-2 and
MMP-9 were involved in the inhibition of invasion by Cal, the
expression levels of TPA-induced MMP-2 and MMP-9 affected by Cal
were investigated using western blot analysis. As shown in Fig. 3C, the relative activities of MMP-2
increased between 45.48 and 100%, compared with the TPA-only group.
Cal inhibited the TPA-induced MMP-2 activities to 80.64, 57.40 and
48.14% following exposure to 20, 30 and 40 µM Cal,
respectively for 24 h. Similar results were observed for MMP-9, in
which MMP-9 activity increased between 46.13 and 100% following TPA
stimulation, and decreased to 74.68, 65.47 and 58.10% with exposure
to Cal at 20, 30 and 40 µM, respectively, compared with TPA
treatment only (Fig. 3D). These
results suggested that Cal suppressed MMP-2 and MMP-9 to prevent
ECM degradation and inhibit metastasis of the A549 cells.
Effect of Cal on the levels of PKC-α and
ERK1/2 level in A549 cells
In order to further investigate the underlying
mechanism, the effects of Cal on the expression levels of PKC-α and
ERK1/2 were detected using western blot analysis. As shown in
Fig. 3E and F, a significant
increase in the expression of PKC-α following TPA induction, and
suppression of PKC-α following Cal treatment, were observed, in a
dose-dependent manner, in the A549 cells. The expression of PKC-α
without TPA induction was 44.99%, compared with the A549 cells
treated with TPA only, and the levels reduced to 66.30, 60.34 and
52.37% when the TPA-induced cells were exposed to 20, 30 and 40
µM Cal, respectively (P<0.01). The levels of p-ERK levels
were 60.74% without TPA treatment and, following treatment with
TPA, the phosphorylation of ERK1/2 increased significantly. These
increased p-ERK levels were decreased to 79.27, 69.46 and 54.78% by
20, 30 and 40 µM Cal, respectively, in a dose-dependent
manner (Fig. 3G and H; P<0.01).
The results of the PKC-α and ERK1/2 analyses demonstrated that Cal
inhibited the activation of PKC-α and the expression of
p-ERK1/2.
Cal inhibits invasion and migration
through the suppression of PKC-α in A549 cells
To further verify whether Cal inhibited A549 cell
migration and invasion through the PKC-α pathway, the A549 cells
were pretreated with the PKC-α inhibitor, AEB071 (0.1 µM)
for 30 min, and then stimulated with 80 nM TPA in the presence or
absence of Cal (30 µM) for 24 h. As shown in Fig. 4A and B, the relative levels of
PKC-α were reduced to 70.63 and 68.73% when treated with Cal or
AEB071 alone, respectively, following induction by TPA. When the
TPA-induced A549 cells were exposed to Cal combined with AEB071,
the PKC-α level decreased significantly to 42.36%.
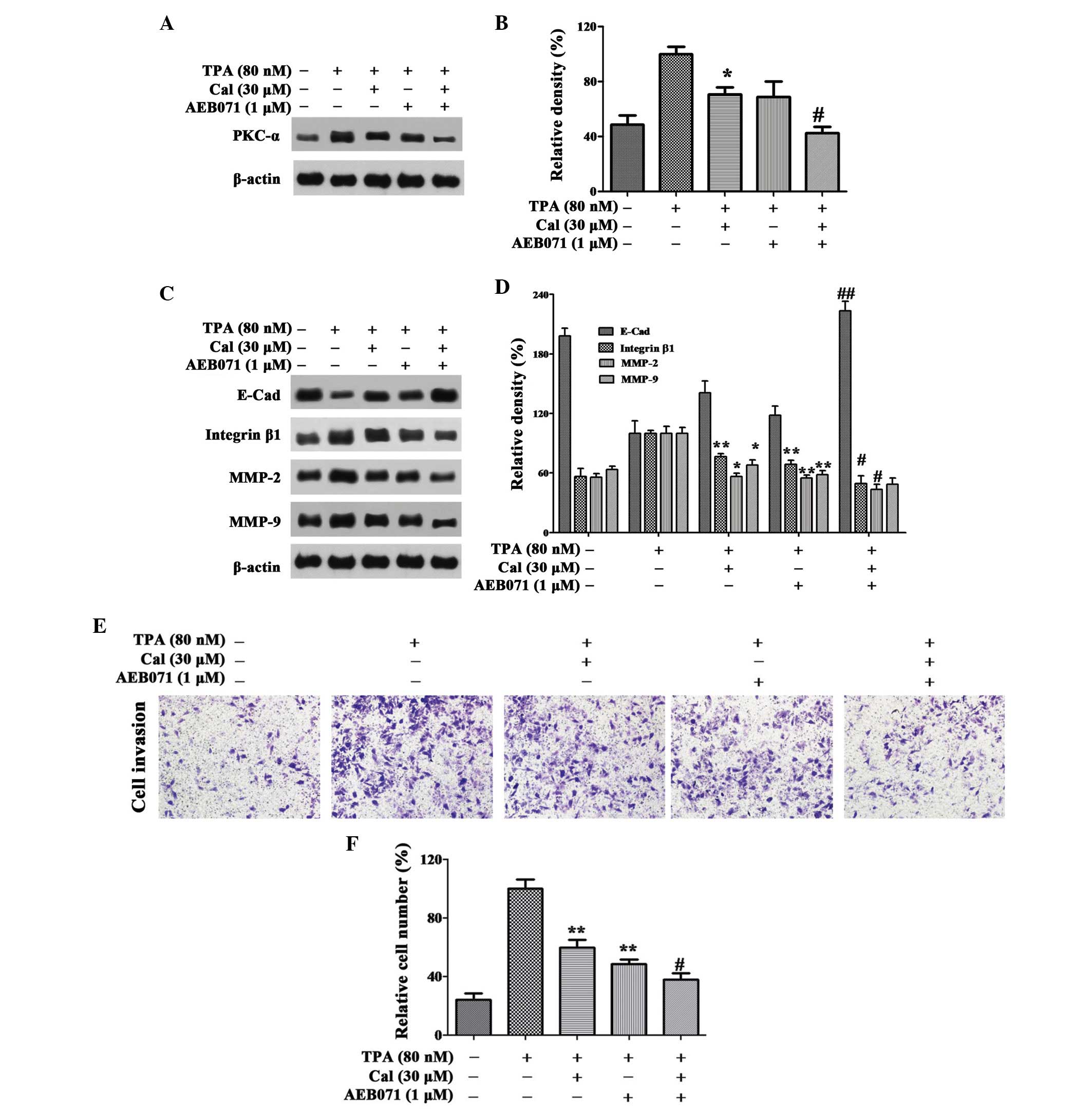 | Figure 4Cal inhibits invasion by suppressing
PKC-α. (A and B) Cells were pretreated with AEB071 (0.1 µM)
for 30 min and then incubated in the presence or absence of Cal (30
µM) for 24 h. The A549 cells were then subjected to western
blotting to analyze the protein levels of PKC-α and (C and D)
E-Cad, integrin β1, MMP-2 and MMP-9. (E) Cells were pretreated with
AEB071 (0.1 µM) for 30 min and then incubated in the
presence or absence of Cal (30 µM) for 24 h. Cellular
invasiveness was measured using a Transwell invasion assay and an
IX73 microscope following staining with crystal violet
(magnification, ×100). (F) Invasion rate is expressed as a
percentage of the TPA-induced group. Values are presented as the
mean ± standard deviation of three independent experiments,
performed in triplicate. *P≤0.05 and
**P≤0.01, vs. TPA-induced group. #P≤0.05 and
##P≤0.01, vs. Cal+ group. Cal, calycosin;
TPA, phorbol-12-myristate-13-acetate; PCK, protein kinase C; E-Cad,
E-cadherin; MMP, matrix metalloproteinase. |
As shown in Fig. 4C and
D, AEB071 and Cal increased the levels of E-Cad reduced by TPA.
Significant increases were observed with their co-treatment.
Treatment with AEB071 or Cal alone reduced the expression levels of
integrin β1, MMP-2 and MMP-9. There were significant decreases in
the expression levels of these proteins following co-treatment of
AEB071 and Cal, compared with their treatment alone.
In the Transwell invasion experiment, when the A549
cells were treated with Cal or AEB071 alone, the cell invasion
ability was decreased to 59.78 and 48.60%, compared with the
TPA-induced group. The relative percentage of permeated cells was
reduced to 37.92% when treated with Cal combined with AEB071
(P<0.01; Fig. 4E and F). The
results suggested that Cal inhibited the invasion of A549 cells by
downregulating the expression levels of integrin β1, MMP-2 and
MMP-9, and elevating the expression of E-Cad, via suppression of
the PKC-α pathway.
Cal inhibits invasion and migration of
A549 cells through reducing the activation of ERK1/2
phosphorylation
In order to investigate whether Cal affected the
downregulation of ERK1/2 phosphorylation, the ERK1/2 inhibitor,
PD98059 (25 µM), was used in the present study, and the
protein expression levels of ERK1/2 and p-ERK1/2 in the A549 cells
were detected using western blot analysis. The relative expression
level of p-ERK1/2 was reduced to 51.42% and 38.73% following
treatment with Cal or PD98059 alone, respectively, and the
expression level was 26.13% following treatment with Cal and
PD98059 combined (Fig. 5A and
B).
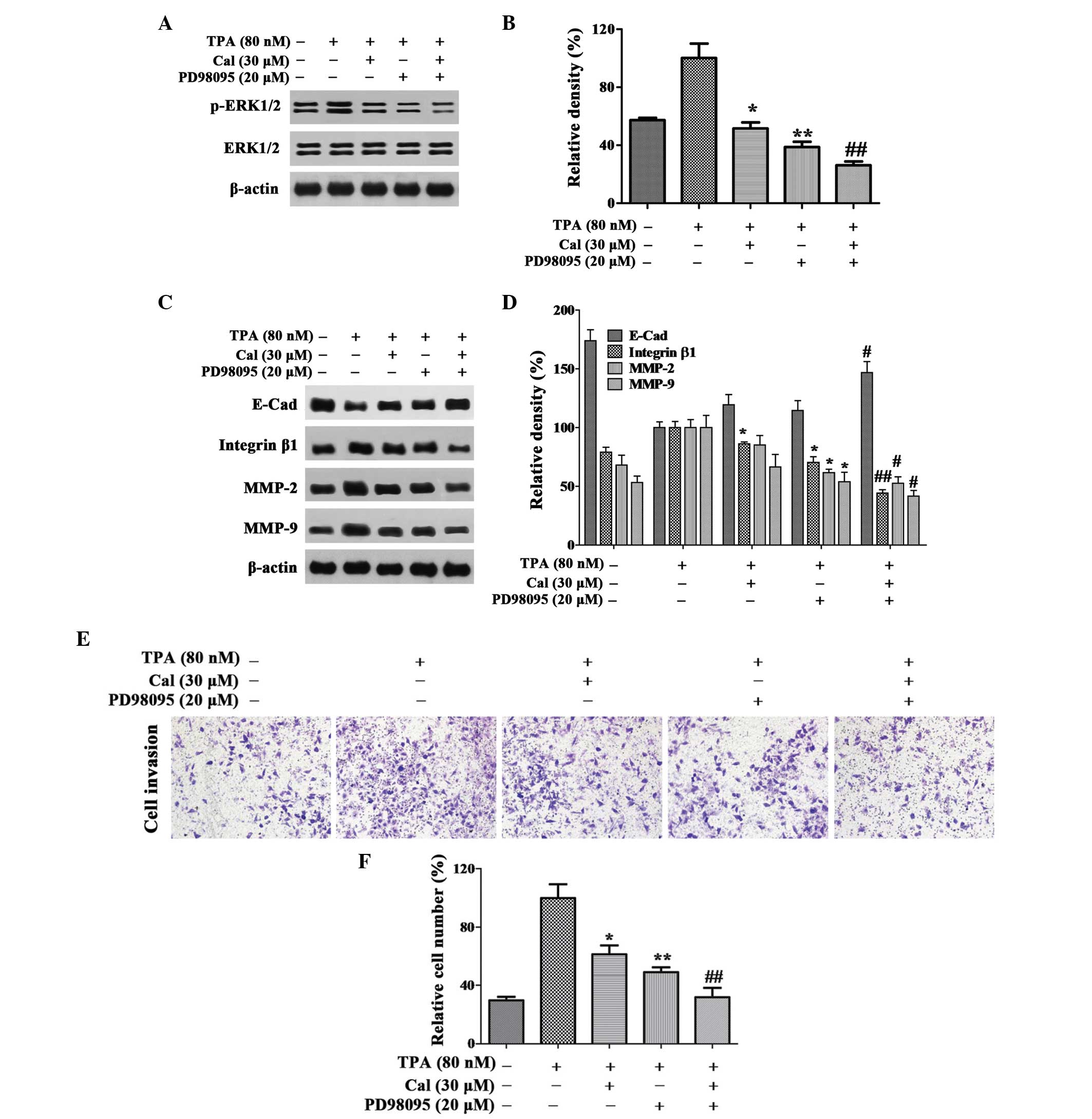 | Figure 5Cal inhibits invasion by suppressing
ERK1/2 phosphorylation. (A and B) Cells were pretreated with
PD98059 (20 µM) for 30 min and then incubated in the
presence or absence of Cal (30 µM) for 24 h. A549 cells were
then subjected to western blotting to analyze the protein levels of
p-ERK1/2 and ERK1/2 and (C and D) E-Cad, integrin β1, MMP-2 and
MMP-9. (E) Cells were pretreated with PD98059 (20 µM) for 30
min and then incubated in the presence or absence of Cal (30
µM) for 24 h. Cell invasiveness was measured using a
Transwell invasion assay and an IX73 microscope following staining
with crystal violet (magnification, ×100). (F) Invasion rate is
expressed as a percentage of the TPA-induced group. Values are
presented as the mean ± standard deviation of three independent
experiments, performed in triplicate. *P≤0.05 and
**P≤0.01, vs. TPA-induced group; #P≤0.05 and
##P≤0.01, vs. Cal+ group. Cal, calycosin;
TPA, phorbol-12-myristate-13-acetate; PCK, protein kinase C; p-ERK,
phosphorylated extracellular signal-regulated kinase; E-Cad,
E-cadherin; MMP, matrix metalloproteinase. |
As shown in Fig. 5C and
D, the expression levels of integrin β1, MMP-2 and MMP-9 were
decreased markedly following co-treatment with PD98059 and Cal.
Notably, these expression levels were lower than those observed
following treatment with either Cal or PD98059 alone. In the
TPA-induced A549 cells, a significant increase in the expression of
E-Cad was observed in the Cal and PD98059 co-treatment group,
compared with treatment with either alone.
The results of the invasion assay indicated that the
relative percentage of permeated cells were significantly inhibited
by Cal and PD98059 co-treatment to 31.84%, compared with 61.29 and
48.80%, respectively, following treatment with Cal or PD98059 alone
(P<0.01; Fig. 5E and F). These
results suggested that Cal inhibited the invasion of A549 cells by
downregulating the expression levels of integrin β1, MMP-2 and
MMP-9, and elevating the expression of E-Cad via suppression of the
ERK1/2 pathway.
Discussion
Lung cancer invasion and metastasis are the major
reasons for the failure of surgery and radiotherapy to cure disease
and for relapse following systemic therapy (30). Therefore, chemical materials, which
have an effect on the treatment of metastasis have an important
impact on lung cancer mortality rates. Studies have demonstrated
that Cal reduces AGE-induced macrophage migration and adhesion to
human umbilical vein endothelial cells (31,32).
However, the antimetastatic effect and the underlying mechanisms in
A549 cells remain to be elucidated. The present study revealed that
Cal inhibited the migration and invasion of A549 lung cancer cells
via suppression of the PKC-α/ERK 1/2 signaling pathways to regulate
the expression levels of migration and invasion-associated
proteins, including E-Cad, integrin β1 and MMPs.
Cancer cell proliferation and apoptosis are
physiologic processes, which are important in tumor development
(33). In clinical cancer therapy,
cell proliferation inhibition and apoptosis induction in tumor
tissue remain the optimum treatment strategy (34). Cal has been reported to inhibit
proliferation and induce apoptosis in osteosarcoma and MCF-7 cells
(35,36). In the present study, Cal inhibited
the proliferation of A549 cells in a dose-dependent manner. The
percentage of apoptotic cells, determined using Annexin V-FITC/PI
staining revealed that, following Cal treatment for 24 h, the
proportion of apoptotic cells increased markedly.
Cancer cell migration and invasion are associated
with adhesion, not only with the cell, but also with the ECM
(37). The levels of proteins,
including E-Cad and integrin β1 are associated with the adhesion
ability of cancer cells. E-Cad is a key protein involved in
cell-cell adhesion, and loss of E-Cad can reduce cell adhesion in
the tissue and promote invasion and metastasis in several types of
epithelial tumor (38,39). The level of E-Cad can be suppressed
by TPA in Caco-2 cells and results in the disassembly of adherin
junctions (40). Integrin
β1-mediated adhesion, migration and metastasis are induced by
activating intracellular signaling pathways, including the ERK and
phosphoinositide 3-kinase signaling pathways (41). Integrin β1 is involved in lung
cancer invasion and metastasis and can be inhibited by suppressing
the ERK1/2 signaling pathway (42). In the present study, Cal
significantly inhibited A549 cell migration stimulated by TPA via
upregulating the expression of E-Cad and downregulating the
expression of integrin β1.
Cell invasion requires proteolysis of ECM components
and transmigration through the ECM (43). ECM degradation allows cancer cells
to invade into blood or lymphatic system and spread to distant
tissues and organs. In these processes, the expression of
proteolytic enzymes, including, MMPs and particularly MMP-2 and
MMP-9 are crucial for ECM degradation (44). The overexpression of MMP-2 and
MMP-9 induced by TPA result in a significant increase in invasion
and may be suppressed by the PKC α/ERK/NF-κB pathway (45). Treatment with Cal significantly
inhibited cell adhesion and invasion by downregulating the
expression levels of MMP-2 and MMP-9, according to the results of
the present study.
The expression levels of the E-Cad, integrin β1,
MMP-2 and MMP-9 proteinases can be regulated by multiple signaling
pathways, including PKC-α (46).
Overexpression of PKC-α, stimulated by TPA, can promote tumor cell
metastasis via upregulating the expression of MMPs and
downregulating the expression of E-cad (47,48).
The reduction in the stimulated expression of PKC-α by TPA with
inhibitors may reduce the motility and invasion of A549 cells, as
well as the expression levels of MMPs (49). In present study, when exposed to
Cal, the overexpression of PKC-α induced by TPA decreased in a
dose-dependent manner. AEB071, an effective PKC-α inhibitor, can be
used to verify whether PKC-α was involved in cancer cell migration
and invasion (50). In the present
study, treatment with AEB071 significantly inhibited migration and
invasion by reducing the protein expression levels of integrin β1,
MMP-2 and MMP-9, and increasing the protein expression of E-Cad.
The results suggested that the Cal inhibited the invasion of A549
cells by downregulating integrin β1, MMP-2 and MMP-9, and elevating
E-Cad, via suppression of the PKC-α pathway.
The overexpression of PKC-α can activate downstream
signaling pathway, including ERK1/2, and alter the expression of
E-Cad, integrin β1, MMP-2 and MMP-9, which leads to tumor migration
and invasion (51). Chemicals
inhibiting ERK1/2 phosphorylation may restrain cancer cell
migration and invasion (52). The
phosphorylation of ERK1/2 in cells treated with Cal was
significantly reduced, compared with the TPA-induced cells in the
present study. The ERK1/2 inhibitor, PD98059, is an effective
method to verify whether the ERK1/2 signaling is pathway involved
in cancer cell invasion (53).
Combined with PD98059, Cal significantly suppressed migration and
invasion, downregulated the protein expression levels of integrin
β1, MMP-2 and MMP-9, and increased the expression of E-Cad in the
present study. Therefore, Cal inhibited the invasion of the A549
cells by downregulating the expression levels of integrin β1, MMP-2
and MMP-9, and elevating the expression of E-Cad, via suppression
of the ERK1/2 pathway.
In conclusion, the results of the present study
demonstrated that Cal inhibited the proliferation, invasion and
migration of A549 cells by suppressing the PKC-α/ERK1/2 signaling
pathway. The results offer novel insight into the molecular
mechanisms of Cal in lung cancer therapy. Therefore, Cal may be a
useful compound for the inhibition of metastasis in lung
cancer.
Acknowledgments
This study is supported by the Suzhou Science and
Technology Program (grant. no. ZXY2012009) and the College Graduate
Research and Innovation Project of Jiangsu Province (grant. no.
CXLX13_599).
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeSantis CE, Lin CC, Mariotto AB, Siegel
RL, Stein KD, Kramer JL, Alteri R, Robbins AS and Jemal A: Cancer
treatment and survivorship statistics. CA Cancer J Clin.
64:252–271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shiono S, Kanauchi N, Yanagawa N, Abiko M
and Sato T: Stage II–IV lung cancer cases with lymphovascular
invasion relapse within 2 years after surgery. Gen Thorac
Cardiovasc Surg. 62:112–118. 2014. View Article : Google Scholar
|
|
4
|
Lai CS, Boshoff C, Falzon M and Lee SM:
Complete response to erlotinib treatment in brain metastases from
recurrent NSCLC. Thorax. 61(91)2006.PubMed/NCBI
|
|
5
|
Huo XW, Li SN, Shi TT, Suo AL, Ruan ZP and
Yao Y: Tripartite motif 16 inhibits epithelial-mesenchymal
transition and metastasis by downregulating sonic hedgehog pathway
in non-small cell lung cancer cells. BBRC. 460(1021)2015.
|
|
6
|
Jo E, Park SJ, Choi YS, Jeon WK and Kim
BC: Kaempferol suppresses transforming growth factor-β1-induced
epithelial-to-mesenchymal transition and migration of A549 lung
cancer cells by inhibiting Akt1-mediated phosphorylation of Smad3
at threonine-179. Neoplasia. 17(525)2015. View Article : Google Scholar
|
|
7
|
Liu ZL, Zhu WR, Zhou WC, Ying HF, Zheng L,
Guo YB, Chen JX and Shen XH: Traditional Chinese medicinal herbs
combined with epidermal growth factor receptor tyrosine kinase
inhibitor for advanced non-small cell lung cancer: A systematic
review and meta-analysis. J Integr Med. 12:346–358. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang XB, Wu WY, Long SQ, Deng H and Pan
ZQ: Effect of gefitinib plus Chinese herbal medicine (CHM) in
patients with advanced non-small-cell lung cancer: a retrospective
case-control study. Complement Ther Med. 22:1010–1018. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou ZY, Xu L, Li HG, Tian JH, Jiao LJ,
You SF, Han ZF, Jiang Y, Guo HR and Liu H: Chemotherapy in
conjunction with traditional Chinese medicine for survival of
elderly patients with advanced non-small-cell lung cancer: protocol
for a randomized double-blind controlled trial. J Integr Med.
12:175–181. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ma X, Zhang T, Wei Y, Tu P, Chen Y and Ito
Y: Preparative isolation and purification of calycosin from
Astragalus membranaceus Bge. var. mongholicus (Bge.) Hsiao by
high-speed counter-current chromatography. J Chromatogr A.
962:243–247. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gao J, Liu ZJ, Chen T and Zhao D:
Pharmaceutical properties of calycosin, the major bioactive
isoflavonoid in the dry root extract of Radix astragali. Pharm
Biol. 52:1217–1222. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang D, Wang S, Zhu L, Tian Y, Wang H,
Zhuang Y, Li Y and Wang D: Profiling of hepatocellular carcinoma
cell cycle regulating genes targeted by calycosin. Biomed Res Int.
2013(317926)2013.
|
|
13
|
Wang Y, Dong X, Li Z, Wang W, Tian J and
Chen J: Downregulated RASD1 and upregulated miR-375 are involved in
protective effects of calycosin on cerebral ischemia/reperfusion
rats. J Neurol Sci. 339:144–148. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li W, Sun YN, Yan XT, Yang SY, Kim S, Lee
YM, Koh YS and Kim YH: Flavonoids from Astragalus membranaceus and
their inhibitory effects on LPS-stimulated pro-inflammatory
cytokine production in bone marrow-derived dendritic cells. Arch
Pharm Res. 37:186–192. 2014. View Article : Google Scholar
|
|
15
|
Liu Y, He J, Chen X, Li J, Shen M, Yu W,
Yang Y and Xiao Z: The Proapoptotic Effect of Formononetin in Human
Osteosarcoma Cells: Involvement of Inactivation of ERK and Akt
Pathways. Cell Physiol Biochem. 34:637–645. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tian J, Duan YX, Bei CY and Chen J:
Calycosin induces apoptosis by upregulation of RASD1 in human
breast cancer cells MCF-7. Horm Metab Res. 45:593–598. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang KP: Role of protein kinase C in
cellular regulation. Biofactors. 2:171–178. 1990.PubMed/NCBI
|
|
18
|
Konopatskaya O and Poole AW: Protein
kinase Calpha: Disease regulator and therapeutic target. Trends
Pharmacol Sci. 31:8–14. 2010. View Article : Google Scholar :
|
|
19
|
Hwang YP, Yun HJ, Kim HG, Han EH, Choi JH,
Chung YC and Jeong HG: Suppression of
phorbol-12-my-ristate-13-acetate-induced tumor cell invasion by
piperine via the inhibition of PKCα/ERK1/2-dependent matrix
metallopro-teinase-9 expression. Toxicol Lett. 203:9–19. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin CW, Shen SC, Chien CC, Yang LY, Shia
LT and Chen YC: 12-O-tetradecanoylphorbol-13-acetate-induced
invasion/migration of glioblastoma cells through activating
PKCalpha/ERK/NF-kappaB-dependent MMP-9 expression. J Cell Physiol.
225:472–481. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang MY, Hsu LS, Peng CH, Shi YS, Wu CH
and Wang CJ: Polyphenol-rich extracts from Solanum nigrum
attenuated PKC alpha-mediated migration and invasion of
hepatocellular carcinoma cells. J Agric Food Chem. 58:5806–5814.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lau GT, Huang H, Lin SM and Leung LK:
Butein downregulates phorbol 12-myristate 13-acetate-induced COX-2
transcriptional activity in cancerous and non-cancerous breast
cells. Eur J Pharmacol. 648:24–30. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Roskoski R Jr: ERK1/2 MAP kinases:
Structure, function and regulation. Pharmacol Res. 66:105–143.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou X, Liu Y, You J, Zhang H, Zhang X and
Ye L: Myosin light-chain kinase contributes to the proliferation
and migration of breast cancer cells through cross-talk with
activated ERK1/2. Cancer Lett. 270:312–327. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang JY, Li S, Li ZH, Zhang ZJ, Hu G,
Cheang LC, Alex D, Hoi MP, Kwan YW, Chan SW, et al: Calycosin
promotes angiogenesis involving estrogen receptor and
mitogen-activated protein kinase (MAPK) signaling pathway in
zebrafish and HUVEC. PLoS One. 5:e118222010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen J, Hou R, Zhang X, Ye Y, Wang Y and
Tian J: Calycosin suppresses breast cancer cell growth via
ERβ-dependent regulation of IGF-1R, p38 MAPK and PI3K/Akt pathways.
PLoS One. 9:e912452014. View Article : Google Scholar
|
|
27
|
Liao YC, Shih YW, Chao CH, Lee XY and
Chiang TA: Involvement of the ERK signaling pathway in fisetin
reduces invasion and migration in the human lung cancer cell line
A549. J Agric Food Chem. 57:8933–8941. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shieh JM, Chiang TA, Chang WT, Chao CH,
Lee YC, Huang GY, Shih YX and Shih YW: Plumbagin inhibits
TPA-induced MMP-2 and u-PA expressions by reducing binding
activities of NF-kappaB and AP-1 via ERK signaling pathway in A549
human lung cancer cells. Mol Cell Biochem. 335:181–193. 2010.
View Article : Google Scholar
|
|
29
|
Liu B, Wang G, Yang J, Pan X, Yang Z and
Zang L: Berberine inhibits human hepatoma cell invasion without
cytotoxicity in healthy hepatocytes. PLoS One. 6:e214162011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Noda M and Takahashi C: Recklessness as a
hallmark of aggressive cancer. Cancer Sci. 98:1659–1665. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu Y, Feng L, Wang S, Zhu Q, Zheng Z,
Xiang P, He B and Tang D: Calycosin protects HUVECs from advanced
glycation end products-induced macrophage infiltration. J
Ethnopharmacol. 137:359–370. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu Y, Feng L, Wang S, Zhu Q, Lin J, Lou C,
Xiang P, He B, Zheng Z, Tang D and Zuo G: Phytoestrogen
calycosin-7-O-β-D-glucop yranoside ameliorates advanced glycation
end products-induced HUVEC damage. J Cell Biochem. 112:2953–2965.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lokeshwar BL, Selzer MG, Zhu BQ, Block NL
and Golub LM: Inhibition of cell proliferation, invasion, tumor
growth and metastasis by an oral non-antimicrobial tetracycline
analog (COL-3) in a metastatic prostate cancer model. Int J Cancer.
98:297–309. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hu C, Yi C, Hao Z, Cao S, Li H, Shao X,
Zhang J, Qiao T and Fan D: The effect of somatostatin and SSTR3 on
proliferation and apoptosis of gastric cancer cells. Cancer Biol
Ther. 3:726–730. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qiu R, Ma G, Zheng C, Qiu X, Li X, Li X,
Mo J, Li Z, Liu Y, Mo L, et al: Antineoplastic effect of calycosin
on osteosarcoma through inducing apoptosis showing in vitro and in
vivo investigations. Exp Mol Pathol. 97:17–22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen J, Zhao X, Ye Y, Wang Y and Tian J:
Estrogen receptor beta-mediated proliferative inhibition and
apoptosis in human breast cancer by calycosin and formononetin.
Cell Physiol Biochem. 32:1790–1797. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Trepat X, Chen Z and Jacobson K: Cell
migration. Compr Physiol. 2:2369–2392. 2012.
|
|
38
|
Canel M, Serrels A, Frame MC and Brunton
VG: E-cadherin-integrin crosstalk in cancer invasion and
metastasis. J Cell Sci. 126:393–401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rodriguez-Teja M, Gronau JH, Minamidate A,
Darby S, Gaughan L, Robson C, Mauri F, Waxman J and Sturge J:
Survival outcome and EMT suppression mediated by a lectin domain
interaction of endo180 and CD147. Mol Cancer Res. 13:538–547. 2015.
View Article : Google Scholar
|
|
40
|
Barbosa LA, Goto-Silva L, Redondo PA,
Oliveira S, Montesano G, De Souza W and Morgado-Díaz JA:
TPA-induced signal transduction: A link between PKC and EGFR
signaling modulates the assembly of intercellular junctions in
Caco-2 cells. Cell Tissue Res. 312:319–331. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hou M, Cui J, Liu J, Liu F, Jiang R, Liu
K, Wang Y, Yin L, Liu W and Yu B: Angiopoietin-like 4 confers
resistance to hypoxia/serum deprivation-induced apoptosis through
PI3K/Akt and ERK1/2 signaling pathways in mesenchymal stem cells.
PLoS One. 9:e858082014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang H, Wu C, Wan S, Zhang H, Zhou S and
Liu G: Shikonin attenuates lung cancer cell adhesion to
extracellular matrix and metastasis by inhibiting integrin β1
expression and the ERK1/2 signaling pathway. Toxicology.
308:104–112. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Willis AL, Sabeh F, Li XY and Weiss SJ:
Extracellular matrix determinants and the regulation of cancer cell
invasion stratagems. J Microsc. 251:250–260. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen K, Zhang S, Ji Y, Li J, An P, Ren H,
Liang R, Yang J and Li Z: Baicalein inhibits the invasion and
metastatic capabilities of hepatocellular carcinoma cells via
downregulation of the ERK pathway. PLoS One. 8:e729272013.
View Article : Google Scholar
|
|
45
|
Shi MD, Shih YW, Lee YS, Cheng YF and Tsai
LY: Suppression of 12-O-tetradecanoylphorbol-13-acetate-induced
MCF-7 breast adenocarcinoma cells invasion/migration by α-tomatine
through activating PKC α/ERK/NF-κB-dependent MMP-2/MMP-9
expressions. Cell Biochem Biophys. 66:161–174. 2013. View Article : Google Scholar
|
|
46
|
Wang Y, Yang H, Liu H, Huang J and Song X:
Effect of staurosporine on the mobility and invasiveness of lung
adenocarcinoma A549 cells: An in vitro study. BMC Cancer.
9:174–186. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Park SH, Kim JH, Lee DH, Kang JW, Song, Oh
SR and Yoon DY: Luteolin 8-C-β-fucopyranoside inhibits invasion and
suppresses TPA-induced MMP-9 and IL-8 via ERK/AP-1 and ERK/NF-κB
signaling in MCF-7 breast cancer cells. Biochimie. 951:2082–2090.
2013. View Article : Google Scholar
|
|
48
|
Zucchini-Pascal N, Peyre L and Rahmani R:
Crosstalk between beta-catenin and snail in the induction of
epithelial to mesenchymal transition in hepatocarcinoma: Role of
the ERK1/2 pathway. Int J Mol Sci. 1410:20768–20792. 2013.
View Article : Google Scholar
|
|
49
|
Chen CC, Chen JJ and Chou CY: Protein
kinase calpha but not p44/42 mitogen-activated protein kinase, p38,
or c-Jun NH(2)-terminal kinase is required for intercellular
adhesion molecule-1 expression mediated by interleukin-1beta:
Involvement of sequential activation of tyrosine kinase, nuclear
factor-kappaB-inducing kinase and IkappaB kinase 2. Mol Pharmacol.
58:1479–1489. 2000.PubMed/NCBI
|
|
50
|
Capsoni F, Ongari AM, Reali E, Bosè F and
Altomare GF: The protein kinase C inhibitor AEB071 (sotrastaurin)
modulates migration and superoxide anion production by human
neutrophils in vitro. Int J Immunopathol Pharmacol. 25:617–626.
2012.PubMed/NCBI
|
|
51
|
Cheng X, Gu J, Zhang M, Yuan J, Zhao B,
Jiang J and Jia X: Astragaloside IV inhibits migration and invasion
in human lung cancer A549 cells via regulating PKC-α-ERK1/2-NF-κB
pathway. Int Immunopharmacol. 23:304–313. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ren W, Liu Y, Wan S, Fei C, Wang W, Chen
Y, Zhang Z, Wang T, Wang J, Zhou L, et al: BMP9 inhibits
proliferation and metastasis of HER2-positive SK-BR-3 breast cancer
cells through ERK1/2 and PI3K/AKT pathways. PLoS One. 9:e968162014.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Karroum A, Mirshahi P, Benabbou N, Faussat
AM, Soria J, Therwath A, Mirshahi M and Hatmi M: Matrix
metallo-proteinase-9 is required for tubular network formation and
migration of resistant breast cancer cells MCF-7 through PKC and
ERK1/2 signalling pathways. Cancer Lett. 295:242–251. 2010.
View Article : Google Scholar : PubMed/NCBI
|














