Introduction
Osteoporosis is characterized by the progressive
loss of bone mass and the micro-architectural deterioration of bone
tissue, resulting in bone fragility and an increased risk of
fracture (1). Gallium is a group
IIIA metal and was first identified in 1875 by Paul-Émile Lecoq de
Boisbaudran in France (2). Gallium
has been demonstrated to be efficacious in the treatment of a
number of diverse disorders that are characterized by accelerated
bone loss, including cancer-associated hypercalcemia, bone
metastases, Paget's disease, myeloma and fatal cage-layer
osteoporosis (2–9). However, the adverse effects
associated with gallium limits its use therapeutically for the
treatment of osteoporosis (10).
Osteoblasts are key cells involved in bone
remodeling. Osteoblasts orchestrate bone remodeling via the
expression of receptor activator of nuclear factor-κB (NF-κB)
ligand (RANKL) in response to osteoclast-stimulating hormones and
cytokines, including parathyroid hormone, tumor necrosis factor and
interleukin-1, however, in addition they protect the skeleton by
secreting osteoprotegerin (OPG) (11). Osteoclasts are important cells in
the process of bone resorption, and are predominantly regulated by
RANKL and OPG, which are secreted by osteoblasts (12). Therefore, the relative
concentrations of RANKL and OPG in bone are key determinants of
bone mass and strength. OPG protects bone from excessive resorption
by binding to RANKL and preventing it from binding to receptor
activator of NF-κB (RANK) (13).
Thus, it is suggested that the balance between RANKL and OPG serves
an important role in the homeostasis of bone metabolism.
Considering the important role of OPG and RANKL in
the regulation of osteoclasts and the inhibitory effects of gallium
nitrate (GaN) on ovariectomized (OVX)-induced bone loss (5), the current study hypothesized that
GaN may regulate the expression of OPG and RANKL in osteoblasts.
Therefore, the aim of the present study was to investigate whether
GaN is able to reduce RANKL and stimulate the expression of OPG in
osteoblasts in vivo and in vitro, and thereby inhibit
the differentiation of osteoclasts and prevent bone loss in OVX
rats. The current study aimed to provide novel insight into the
mechanisms of the effect of GaN on osteoporosis.
Materials and methods
Chemicals, experimental animals and
treatments
GaN was purchased from BetaPharma Co., Ltd.
(Shanghai, China). A total of 45 adult Sprague-Dawley female rats
(Experimental Animal Center, Shengjing Hospital of China Medical
University, Shenyang, China) at 8–10 weeks of age and 180–200 g in
weight were used. Rats were randomly divided into three groups (15
rats/group), all to receive a supplement diet. The rats of two of
the groups were to become OVX rats and those in the remaining group
underwent a sham operation (control). Within 1 week of their
arrival, rats were anesthe-tized with an intraperitoneal injection
of 10% chloral hydrate (3.0 ml/kg; Tianjin Kermel Chemical Reagent
Co., Ltd., Tianjin, China) and underwent either bilateral
ovariectomy or a sham operation. Animals were kept separately in
stainless-steel cages under controlled laboratory conditions
(22–25°C, 40–60% relative humidity, 13 h light/11 h dark cycle)
with standard rat chow and water available ad libitum for 2
months. Following this, group 1 (n=15, sham) and group 2 (n=15,
OVX) were treated with the vehicle (HyClone™ phosphate-buffered
saline; Thermo Fisher Scientific, Inc., Waltham, MA, USA) by
intraperitoneal injection, and group 3 (n=15, OVX + GaN) was
treated with GaN injected intraperitoneally (120 µg/kg/day)
for 8 weeks.
All animal experiments were performed in accordance
with the international standards for animal experimentation
(14). All procedures were
reviewed and approved by the Ethical Committee of China Medical
University (Shenyang, China).
Measurements of bone mineral density
(BMD) using dual-energy X-ray absorptiometry
Dual energy X-ray absorp-tiometry (XR-600; Norland
Medical Systems, Inc., Tustin, CA, USA) was used to scan the left
tibia of all rats to determine the level of BMD under chloral
hydrate (3.0 ml/kg) anesthesia (for ~20 min, once per month.
Preparation of tissue
At the time of sacrifice, blood was collected from
the dorsal aorta under ether anesthesia (Tianjin Kermel Chemical
Reagent Co., Ltd.). Briefly, after the blood was collected, the
rats were sacrificed by administering an overdose of the
anesthetic. Following centrifugation at 1,800 × g at 4°C for 10
min, serum was harvested and stored at −20°C until analysis. The
left tibiae were processed for histology. The right tibiae were
processed for immunohistochemistry. The left femurs were processed
for reverse transcription-quantitative polymerase chain reaction
(RT-qPCR). The right femurs were processed for western blot
analysis. All bone tissues were immediately frozen at −80°C.
Bone histomorphometry
The tibia tissues were fixed in 4% formaldehyde
(Tianjin Kermel Chemical Reagent Co., Ltd.) for 16 h at 4°C for
histological evaluation. Following fixation, tibiae were
decalcified in 10% ethylene diamine tetraacetic acid (Tianjin
Kermel Chemical Reagent Co., Ltd.), dehydrated, embedded in
paraffin (Beyotime Institute of Biotechnology, Shanghai, China),
cut to 5 µm sections and stained with hematoxylin and eosin
(H&E; Beyotime Institute of Biotechnology). The tibial analysis
was conducted in the proximal metaphysis beginning adjacent to the
epiphyseal growth plate. The callus total area, callus bony area
and cartilage area were measured using Image-Pro Plus software,
version 6.0 (Media Cybernetics, Inc., Rockville, MD, USA).
Immunohistochemistry
For immunohistochemistry, sections were incubated
overnight at 4°C with a polyclonal goat anti-rat OPG (cat. no.
orb11189) and polyclonal goat anti-rat RANKL (cat. no. orb11190;
1:100; Biorbyt Ltd., Cambridge, UK). Goat serum (Solarbio, Beijing,
China) was used as the blocking agent. The primary antibodies were
detected by incubation (30 min at 37°C) with an anti-goat IgG
secondary antibody conjugated with horseradish peroxidase (HRP;
cat. no. A0227, 1:200; Beyotime Institute of Biotechnology, Haimen,
China). Antibody complexes were visualized using a digital
microscope (DP73; Olympus, Tokyo, Japan) with 3,3-diaminobenzine
solution containing hydrogen peroxide (Beyotime Institute of
Biotechnology) and then counterstained using hematoxylin. The
integrated optical density was measured using Image-Pro Plus
software, version 6.0 (Media Cybernetics, Inc.).
Osteoblast cell culture
Osteoblasts were isolated from the calvariae of four
male, 1-day old Sprague-Dawley rats (Experimental Animal Center,
Shengjing Hospital of China Medical University) by sequential 0.25%
trypsin and digestion with 0.1% type II collagenase (HyClone™;
Thermo Fisher Scientific, Inc.). Cells released in the second and
third digests were pooled and grown in HyClone™ Dulbecco's modified
Eagle's medium-low glucose medium supplemented with 10% fetal
bovine serum (FBS), 100 IU/ml penicillin and 100 µg/ml
streptomycin (HyClone™; Thermo Fisher Scientific, Inc.) at 37°C in
humidified 5% CO2 air. The cells were regularly
trypsinized and subcultured to prevent cell confluence.
Osteoblast cytotoxicity test of GaN
The cytotoxicity of GaN against osteoblasts was
investigated using a Cell Counting Kit-8 assay (CCK-8; Beyotime
Institute of Biotechnology). In brief, cells were seeded into
96-well flat-bottomed plates at a density of 5×103
cells/well and then placed in serum-starved conditions for a
further 6 h. Subsequently, cells were treated with GaN at
increasing concentrations (0, 10−11, 10−10,
10−9, 10−8, 10−7, 10−6
and 10−5 mol/l) in the presence of 10% FBS for 24 h.
Following the 24 h incubation, 10 µl CCK-8 dye was added to
each well and incubated for 1 h according to the manufacturer's
instructions. The absorbance of each well was measured at 450 nm
using a microplate reader (Synergy H4; BioTek Instruments, Inc.,
Winooski, VT, USA).
Enzyme-linked immunosorbent assay
(ELISA)
Primary osteoblasts were serum starved for 12 h,
followed by GaN (10−9 mol/l) stimulation for 24 h. The
supernatants of the osteoblasts were then collected. For in
vivo and in vitro experiments, the supernatants of the
osteoblasts and the serum from the rats were analyzed to measure
the concentration of OPG and RANKL using the OPG and RANKL ELISA
kits respectively (R&D Systems Inc., Minneapolis, USA),
according to the manufacturer's instructions.
RNA extraction and RT-qPCR
The left femurs were placed in liquid nitrogen, bone
chips were collected and gently mashed using a mallet. The bone
tissues were then ground into a powder in liquid nitrogen using a
pestle. For the in vivo and in vitro experiments, the
total RNA was extracted from the pulverized bone powder and cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
instructions. The RNA concentration was determined by measuring
absorbance at 260 nm, and 1 µg total RNA was transcribed to
cDNA using the PrimerScript™ RT Reagent kit with gDNA Eraser
(Takara Biotechnology Co., Ltd., Dalian, China) according to the
manufacturer's instructions. The primers used were as follows: OPG,
forward 5′-GAC CCC AGA GCG AAA CAC G-3′ and reverse 5′-GGC ACA GCA
AAC CTG AAG AA-3′; RANKL, forward 5′-CAT CGG GTT CCC ATA AAG-3′ and
reverse 5′-GAA GCA AAT GTT GGC GTA-3′; β-actin, forward 5′-GGA GAT
TAC TGC CCT GGC TCC TAG C-3′ and reverse 5′-GGC CGG ACT CAT CGT ACT
CCT GCT T-3′. RT-qPCR was conducted using the Exicycler™ 96
(Bioneer Corporation, Daejeon, Korea) using the following cycling
conditions: 95°C denaturation step for 10 min followed by 40 cycles
of 95°C for 10 sec, 60°C annealing for 20 sec and extension at 72°C
for 30 sec. The detection of the fluorescent product was conducted
at the end of the 72°C extension period. The housekeeping gene
β-actin was used to normalize the quantities of the target genes.
Data were analyzed using the 2−ΔΔCq method (15), normalizing levels against those of
β-actin. To evaluate the mean gene expression level, RT-qPCR was
performed in triplicate for each sample.
Western blot analysis
The pulverized bone powder was homogenized in
radioimmunoprecipitation assay (RIPA) lysis buffer [50 mM Tris pH
7.4, 150 mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1%
sodium dodecyl sulfate (SDS); Beyotime Institute of Biotechnology,
Shanghai] overnight at 4°C. Cell pellets were lysed directly in the
culture bottles using the RIPA buffer. The lysates were collected
and then centrifuged at 16,000 × g for 30 min at 4°C. The
supernatant was then collected and the total protein levels were
measured using a bicinchoninic acid assay kit (Beyotime Institute
of Biotechnology). Equal amounts of protein (40 µg) were
separated by SDS-polyacrylamide gel electrophoresis and transferred
onto a polyvinylidene fluoride membrane at 4°C (Beyotime Institute
of Biotechnology, Shanghai). Following blocking in 5% fat-free milk
for 1 h, the blots were incubated with the following primary
antibodies: Anti-OPG (cat. no. orb11189; 1:200, Beyotime Institute
of Biotechnology), anti-RANKL (cat. no. orb11190; 1:500, Beyotime
Institute of Biotechnology) and anti-β-actin (cat. no. WL0001,
1:1,000; WanLei Life Sciences, Shenyang, China) at 4°C overnight.
Membranes were then washed and incubated with HRP-conjugated
secondary antibodies (anti-goat IgG; cat. no. A0277; 1:5,000) at
37°C for 2 h. Proteins were detected using enhanced
chemiluminescence (Synoptics Ltd., Cambridge, UK). Blots were
repeated at least three times for each condition. Following
development, the band intensities were quantified using
Gel-Pro-Analyzer software, version 4.0 (Media Cybernetics,
Inc.)
Statistical analysis
Values are presented as the mean ± standard
deviation and analyzed using the one-way analysis of variance and
Student's t-test in IBM SPSS 19.0 software (IBM SPSS, Armonk, MY,
USA). All experiments were repeated a minimum of three times, and
representative experiments are presented throughout. P<0.05 was
considered to indicate a statistically significant difference.
Results
BMD measurements
The average BMD of rats in each group throughout the
experimental period is presented in Fig. 1A. Following ovariectomy, the BMD of
animals was significantly reduced at 4 and 8 weeks (P<0.05), by
16.5 and 34.1%, respectively. However, the administration of GaN to
OVX rats significantly increased BMD at 4 and 8 weeks (P<0.05),
by 19.3 and 37.3%, respectively, compared with the OVX rats.
Bone histomorphometry analysis
To investigate the effect of GaN on OVX-induced bone
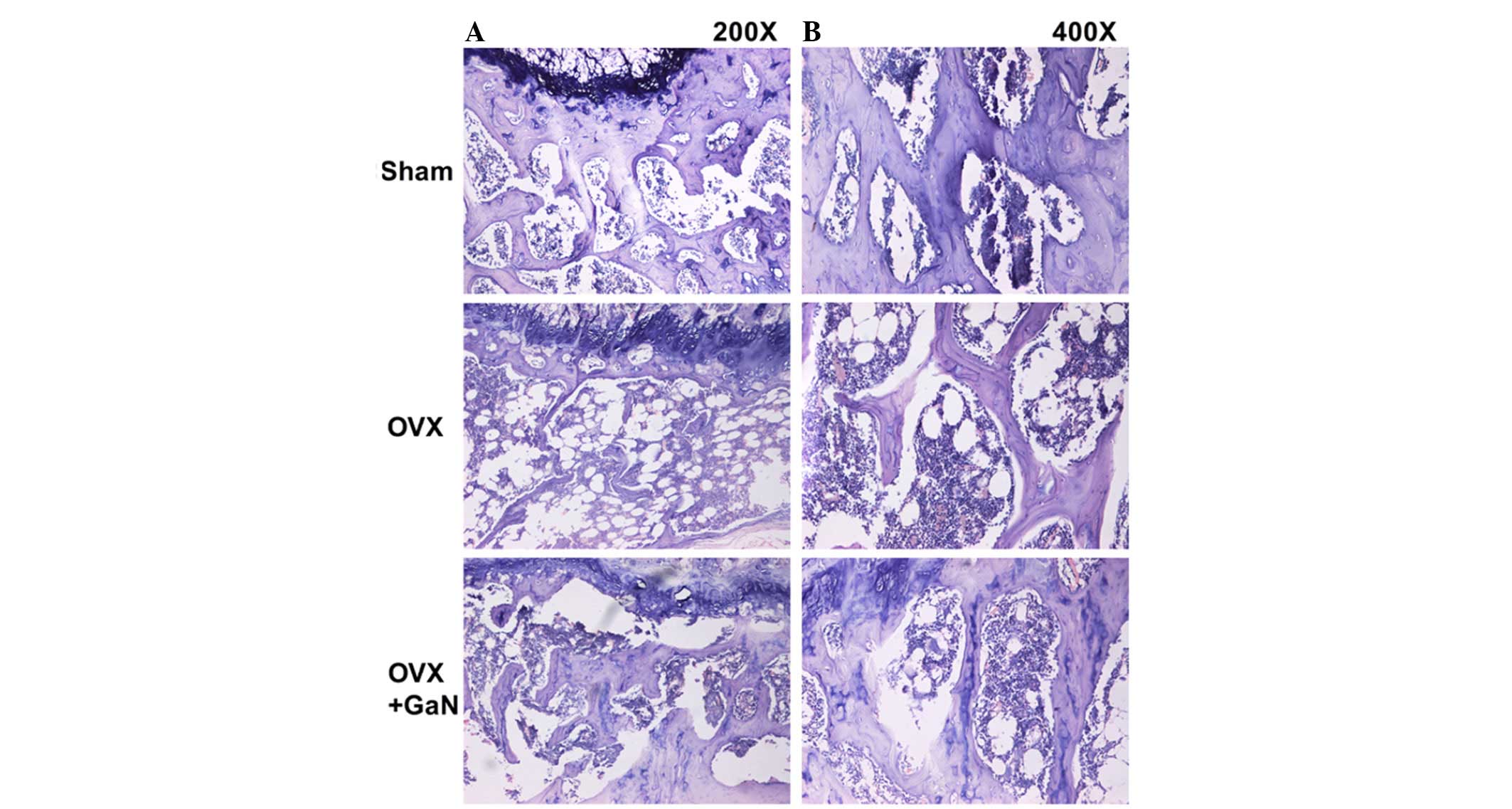
loss in vivo, H&E staining (Fig. 2) was conducted. The results of bone
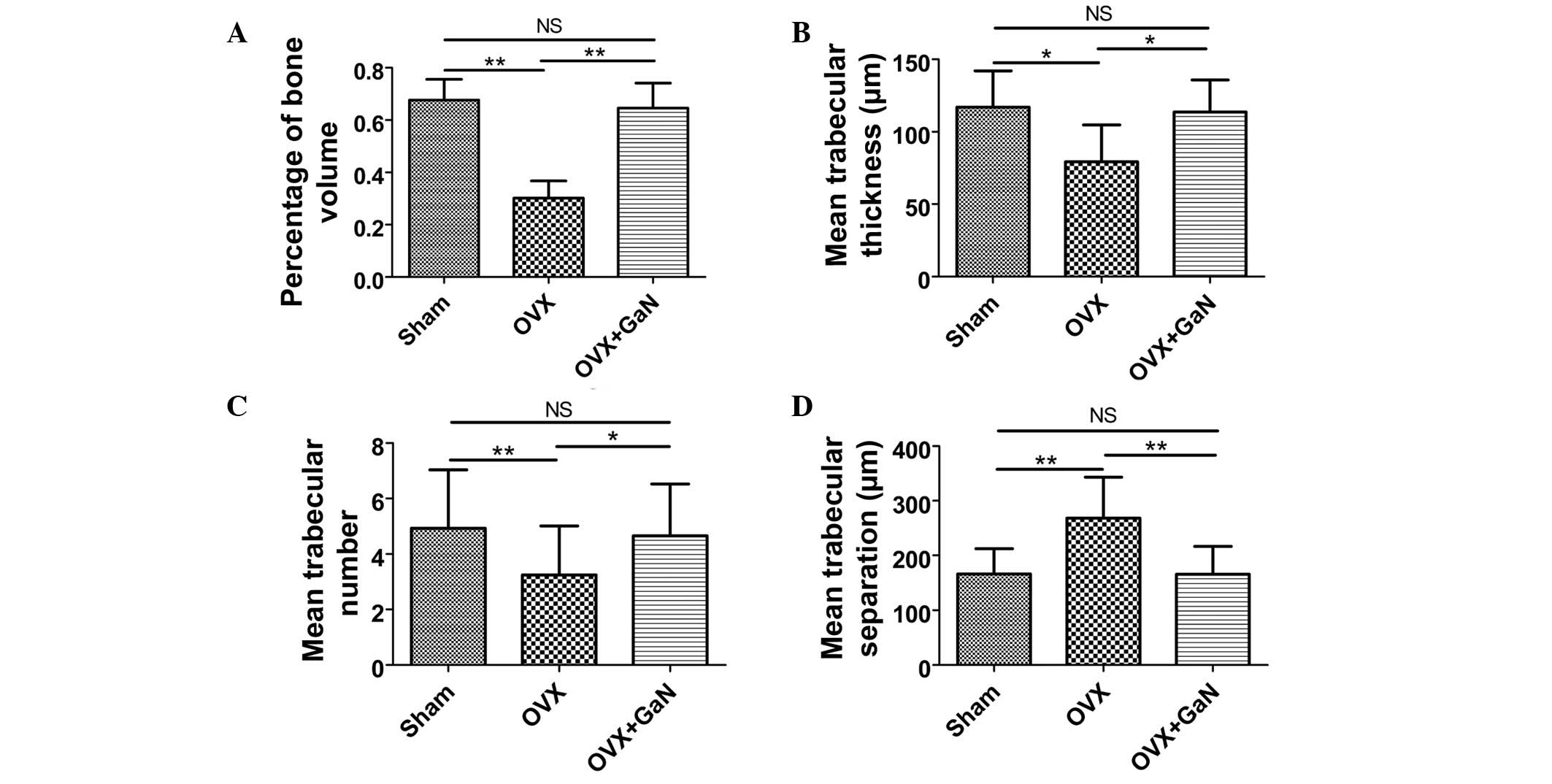
histomorphometry analysis were expressed as the percentage of bone
volume (BV), mean trabecular thickness (Tb.Th), mean trabecular
number (Tb.N) and mean trabecular separation (Tb.Sp). The BV in the
tibia of the OVX + GaN group increased by 40.9% (P<0.05)
compared with the OVX group (Fig.
3A). GaN significantly increased the BV in the tibia of OVX
rats. The Tb.Th of the GaN treatment group was significantly
increased by 43.3% (P<0.01) compared with the OVX group
(Fig. 3B). The Tb.N exhibited an
increase following GaN treatment of 43.4% (P<0.05) compared with
the OVX group (Fig. 3C). In
addition, treatment with GaN led to a reduction in the Tb.Sp by
38.2% (P<0.01) compared with the OVX treatment group (Fig. 3D).
Immunohistochemical assessment
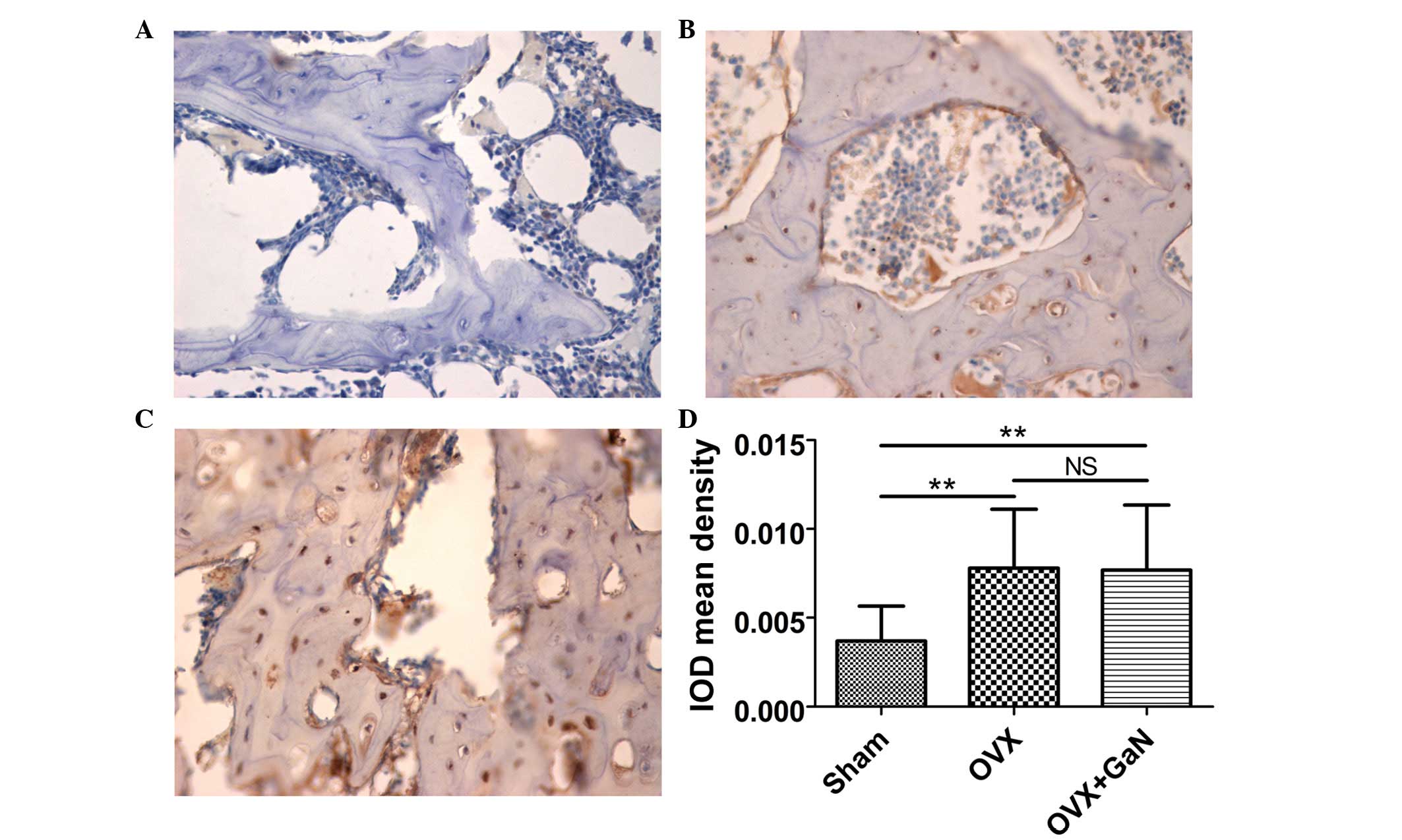
To confirm the preventative effect of GaN on
OVX-induced bone loss in vivo, immunohistochemical
assessment was conducted to examine the expression of OPG and RANKL
in rats. The results demonstrated that GaN is able to enhance the
expression of OPG (P<0.01) compared with the OVX group (Fig. 4), however without affecting the
expression levels of RANKL compared with the OVX treatment group
(Fig. 5).
Osteoblast cytotoxicity test
The optimal concentration of GaN was determined
using a CCK-8 assay. For the control, the concentration of GaN was
0 mol/l. The optical density values of the osteoblasts were
greatest following treatment with 10−9 mol/l GaN.
Therefore, this concentration was selected as the optimal
concentration of GaN for subsequent investigations (Fig. 1B).
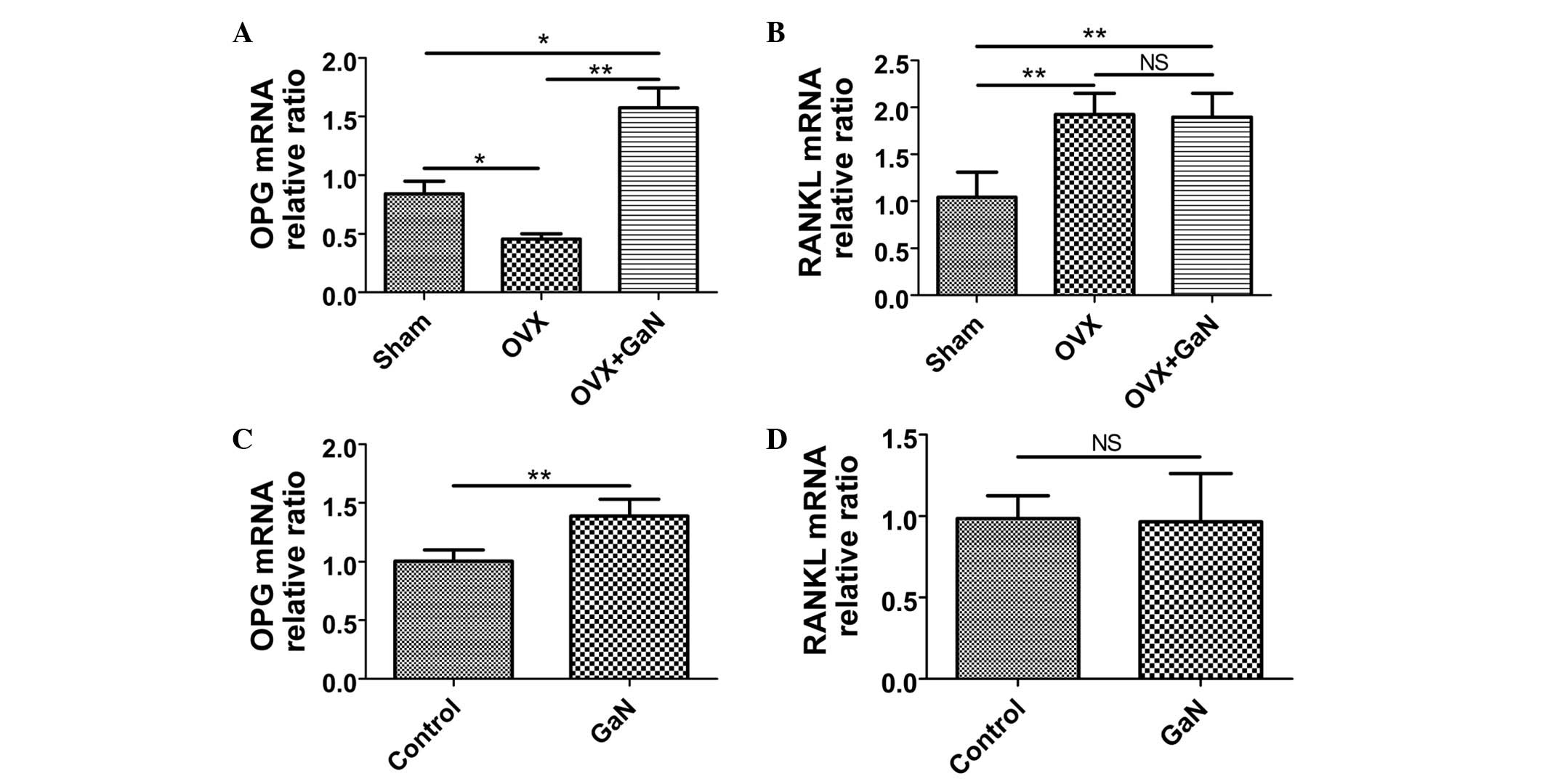
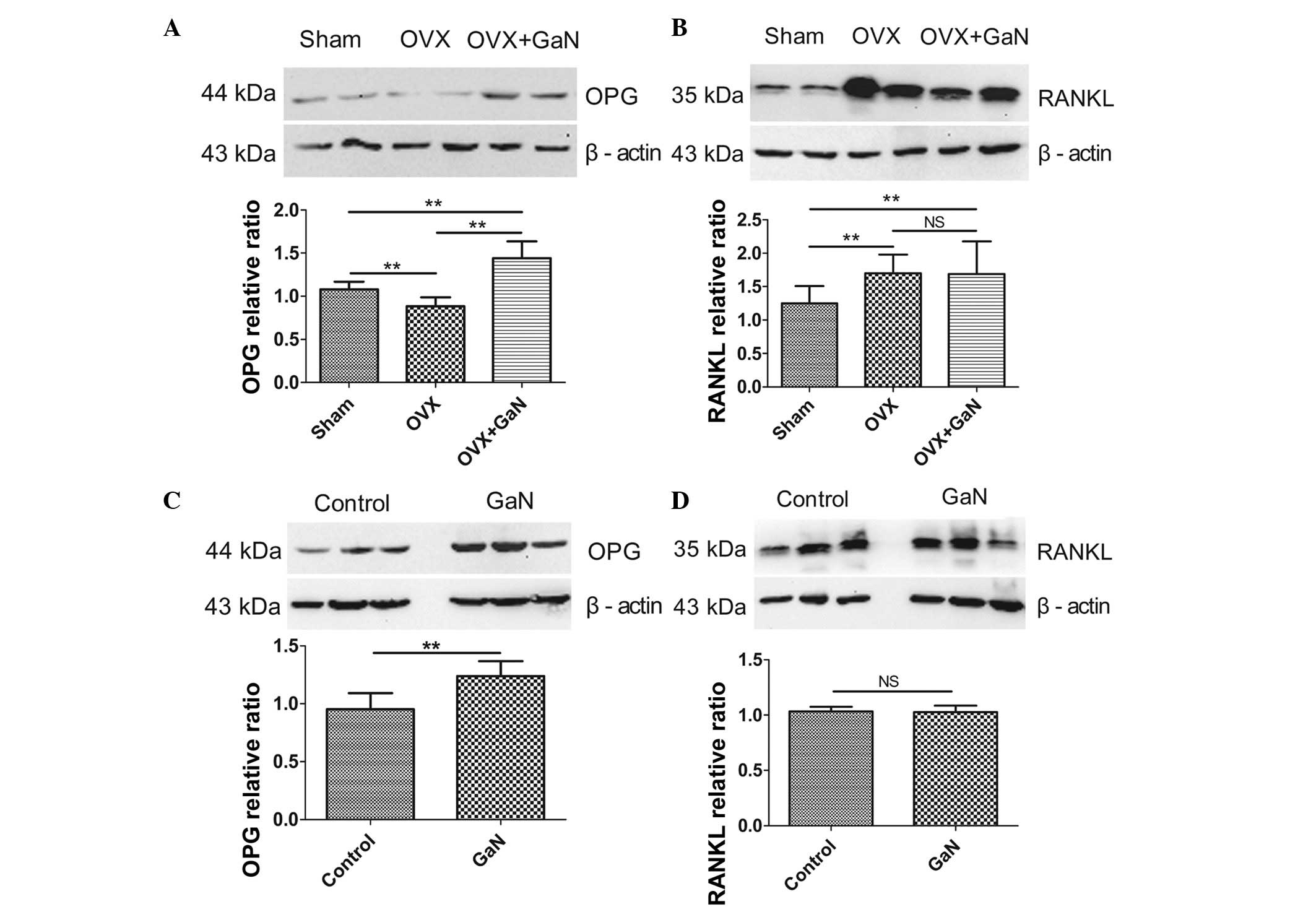
GaN affects the mRNA and protein
expression ratio of OPG/RANKL in vivo and in vitro
To investigate the effect of GaN on OPG and RANKL
expression in vivo and in vitro, OVX rats and
osteoblasts were treated with GaN, and the mRNA and protein levels
of OPG and RANKL in bone tissue were measured using RT-qPCR and
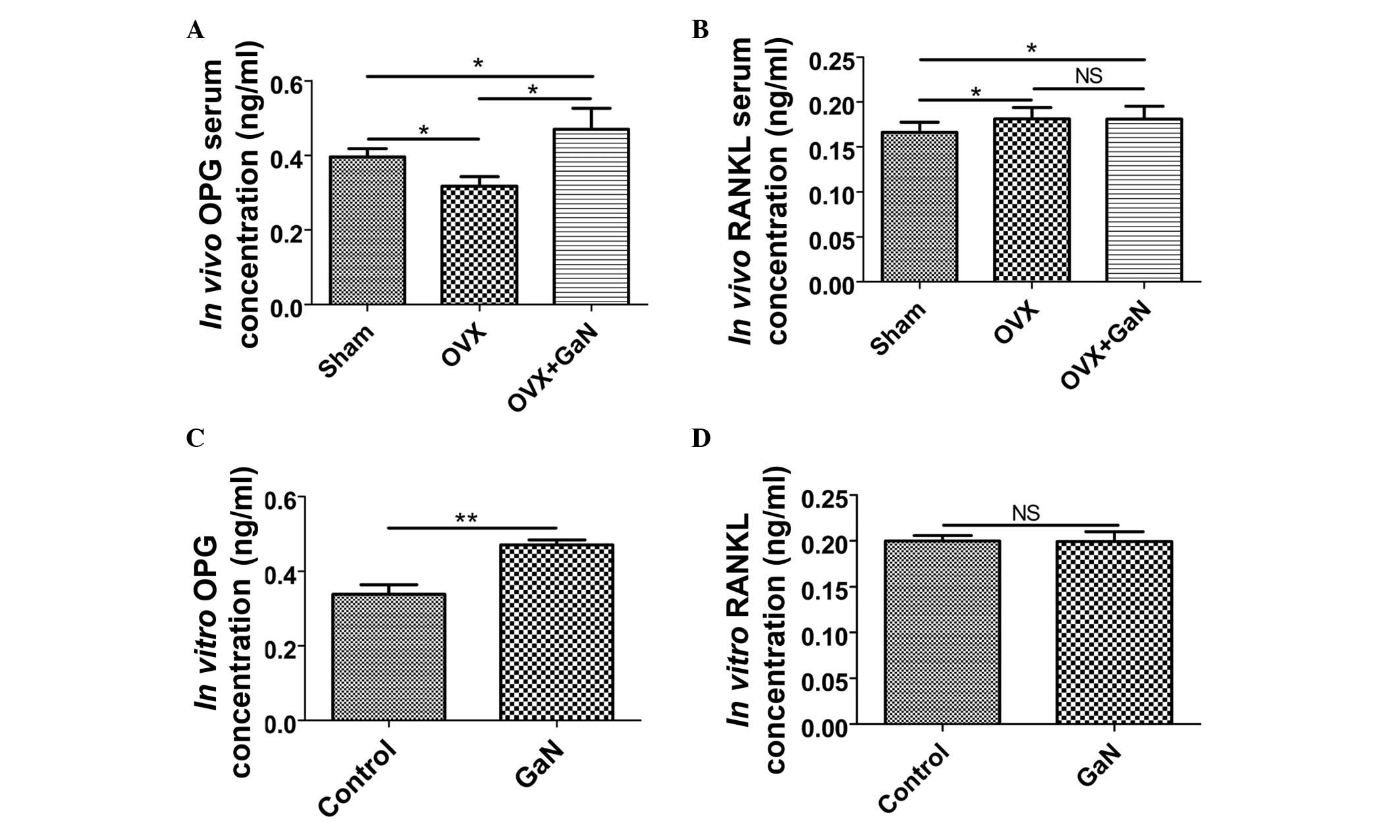
western blot analysis, respectively (Figs. 6 and 7). In addition, the expression levels of
OPG and RANKL in the supernatants of osteoblasts and the serum of
rats were measured using ELISA (Fig.
8). Together, the results indicated that GaN increased the
expression levels of OPG, however did not effect RANKL expression
in GaN-treated OVX rats and osteoblast cells. These data
demonstrate that GaN stimulates OPG however does not affect RANKL
expression in vivo and in vitro.
Discussion
Osteoporosis is the most common bone disorder in
aging populations and is an important public health issue, with
osteoporotic fractures having a key impact upon health, in terms of
acute and long term disability and economic cost (16). The World Health Organization
describes osteoporosis as a generalized metabolic disease
characterized by progressive loss of the elements of bone tissue
and a simultaneous deterioration of skeletal microarchitecture
(17), leading to bone fragility
and resulting in an increased risk of fractures. Epidemiological
data worldwide have consistently demonstrated that the annual
incidence of fragility fractures is increasing, in particular in
ageing populations (18–23). Furthermore, awareness of
osteoporosis is increasing, with this multifactorial disease
regarded as a serious public health issue.
Elemental gallium, a group IIIA metal, alters the
mineral, matrix and cellular properties of bone (24,25).
In animal experiments, gallium has been demonstrated to affect OVX
osteopenic rats by reducing serum mineral levels and increasing
bone mineral content (26). X-ray
diffraction analysis of bone powder from gallium-treated rats
demonstrated alterations characteristic of an increase in the size
or crystalline perfection of hydroxyapatite minerals (27). The current study demonstrated that
GaN treatment resulted in an increase in tibia trabecular bone
parameters, BMD and bone strength in OVX rats. The results of
histomorphometric analysis indicate that GaN administration
increased the BV and Tb.N, whilst reducing the Tb.Sp in the
proximal tibia. These results are in accordance with previous
studies which documented the effect of gallium on osteoporosis in
OVX rats following short-term treatment (16,28).
However, the adverse effects of gallium have limited its use as a
therapeutic agent for the treatment of osteoporosis (10). A common adverse effect is
hypocalcemia, which on occasion is sufficiently severe to result in
transient tetany (29,30). Yeast may be used as an element
carrier that is able to convert inorganic elements to organic
species and thereby reduce element-associated toxicity (31–33).
Previous studies have demonstrated that yeast-incorporated gallium
not only reduces gallium-associated toxicity, however additionally
maintains its therapeutic effect on improving bone loss and
promoting fracture healing in OVX female rats (16,26,28).
Bone formation and resorption are normally in
physiological balance, maintaining a stable bone mass. At the
cellular level, the beneficial effects of gallium are based on a
dual effect on osteoblasts and osteoclasts, which control the
process of bone remodeling. A previous report by Verron et
al (34) indicates that
gallium reduces osteoclastic resorption, differentiation and
formation in vitro without affecting the viability or
activity of primary and MC3T3-E1 osteoblasts. Osteoclasts are key
cells in bone resorption, and their differentiation is
predominantly regulated by RANKL and OPG. As such, the balance
between RANKL and OPG serves a significant role in the homeostasis
of bone metabolism (35–39). OPG protects bone from excessive
resorption by binding to RANKL and preventing it from binding to
RANK. Therefore, the relative concentration of RANKL and OPG in
bone is a major determinant of bone mass and strength (13,40,41).
The evidence from previous studies led to the current study, which
investigated the mechanisms underlying this regulation using an
in vivo experimental model of established osteoporosis and
an in vitro cell culture system. The present study
demonstrated the important role of GaN in the regulation of the
OPG/RANKL axis and the inhibition of osteoclastogenesis in
vivo and in vitro. These data support the view that GaN
inhibits differentiation of osteoclasts by stimulating OPG without
affecting RANKL production in osteoblasts, which may indicate the
potential mechanism of GaN in the prevention of post-menopausal
osteoporosis.
In conclusion, the current study demonstrates that
GaN is able to counteract bone loss in an experimental model of
established osteoporosis. In addition, the data have demonstrated
that GaN stimulates the synthesis of OPG without affecting RANKL
expression in osteoblasts, resulting in an increase in the
OPG/RANKL ratio and a reduction in osteoclast differentiation in
vivo and in vitro. These results suggest that GaN may
upregulate the secretion of OPG in osteoblasts, and subsequently
prevent bone loss and osteoporosis.
Acknowledgments
The current study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81370981
and 81300714).
References
|
1
|
Peck WA: Consensus development conference:
Diagnosis, prophylaxis, and treatment of osteoporosis. Am J Med.
94:646–650. 1993. View Article : Google Scholar
|
|
2
|
Warrell RP Jr, Bockman RS, Coonley CJ,
Isaacs M and Staszewski H: Gallium nitrate inhibits calcium
resorption from bone and is effective treatment for cancer-related
hypercalcemia. J Clin Invest. 73:1487–1490. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Warrell RP Jr, Skelos A, Alcock NW and
Bockman RS: Gallium nitrate for acute treatment of cancer-related
hypercalcemia: Clinicopharmacological and dose response analysis.
Cancer Res. 46:4208–4212. 1986.PubMed/NCBI
|
|
4
|
Warrell RP Jr, Israel R, Frisone M, Snyder
T, Gaynor JJ and Bockman RS: Gallium nitrate for acute treatment of
cancer-related hypercalcemia. A randomized, double-blind comparison
to calcitonin. Ann Intern Med. 108:669–674. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Warrell RP Jr: Gallium nitrate for the
treatment of bone metastases. Cancer. 80(Suppl): 1680–1685. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Warrell RP Jr, Bosco B, Weinerman S,
Levine B, Lane J and Bockman RS: Gallium nitrate for advanced Paget
disease of bone: Effectiveness and dose-response analysis. Ann
Intern Med. 113:847–851. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Niesvizky R: Gallium nitrate in multiple
myeloma: Prolonged survival in a cohort of patients with
advanced-stage disease. Semin Oncol. 30(Suppl 5): 20–24. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li C, Jiang Z and Liu X: Biochemical
mechanism of gallium on prevention of fatal cage-layer
osteoporosis. Biol Trace Elem Res. 134:195–202. 2010. View Article : Google Scholar
|
|
9
|
Chen X and Wang C: Activity of gallium on
prevention of fatal cage-layer osteoporosis. Biol Trace Elem Res.
132:129–135. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Foster BJ, Clagett-Carr K, Hoth D and
Leyland-Jones B: Gallium nitrate: The second metal with clinical
activity. Cancer Treat Rep. 70:1311–1319. 1986.PubMed/NCBI
|
|
11
|
Simonet WS, Lacey DL, Dunstan CR, Kelley
M, Chang MS, Lüthy R, Nguyen HQ, Wooden S, Bennett L, Boone T, et
al: Osteoprotegerin: A novel secreted protein involved in the
regulation of bone density. Cell. 89:309–319. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Teitelbaum SL: Osteoclasts, integrins, and
osteoporosis. J Bone Miner Metab. 18:344–349. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Theoleyre S, Wittrant Y, Tat SK, Fortun Y,
Redini F and Heymann D: The molecular triad OPG/RANK/RANKL:
Involvement in the orchestration of pathophysiological bone
remodeling. Cytokine Growth Factor Rev. 15:457–475. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
ICLA recommendations for the specification
of the animals, the husbandry, and the techniques used in animal
experimentation. International Committee on Laboratory Animals -
Secretariat. Anat Anz. 145:413–414. 1979.
|
|
15
|
Liss B: Improved quantitative real-time
RT-PCR for expression profiling of individual cells. Nucleic Acids
Res. 30e892002.
|
|
16
|
Ma Z and Fu Q: Therapeutic effect of
organic gallium on ovariectomized osteopenic rats by decreased
serum minerals and increased bone mineral content. Biol Trace Elem
Res. 133:342–349. 2010. View Article : Google Scholar
|
|
17
|
Goodfellow LR, Earl S, Cooper C and Harvey
NC: Maternal diet, behaviour and offspring skeletal health. Int J
Environ Res Public Health. 7:1760–1772. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kanis JA and McCloskey EV: Epidemiology of
vertebral osteoporosis. Bone. 13(Suppl 2): S1–S10. 1992.PubMed/NCBI
|
|
19
|
Melton LJ III, Lane AW, Cooper C, Eastell
R, O'Fallon WM and Riggs BL: Prevalence and incidence of vertebral
deformities. Osteoporos Int. 3:113–119. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
van Staa TP, Dennison EM, Leufkens HG and
Cooper C: Epidemiology of fractures in England and Wales. Bone.
29:517–522. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Van der Klift M, De Laet CE, McCloskey EV,
Hofman A and Pols HA: The incidence of vertebral fractures in men
and women: The Rotterdam Study. J Bone Miner Res. 17:1051–1056.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Felsenberg D, Silman AJ, Lunt M, Armbrecht
G, Ismail AA, Finn JD, Cockerill WC, Banzer D, Benevolenskaya LI,
Bhalla A, et al European Prospective Osteoporosis Study (EPOS)
Group: Incidence of vertebral fracture in europe: Results from the
European Prospective Osteoporosis Study (EPOS). J Bone Miner Res.
17:716–724. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lord SR: Hip fractures: Changing patterns
in hospital bed use in NSW between 1979 and 1990. Aust N Z J Surg.
63:352–355. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Repo MA, Bockman RS, Betts F, Boskey AL,
Alcock NW and Warrell RP Jr: Effect of gallium on bone mineral
properties. Calcif Tissue Int. 43:300–306. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bockman RS, Boskey AL, Blumenthal NC,
Alcock NW and Warrell RP Jr: Gallium increases bone calcium and
crystallite perfection of hydroxyapatite. Calcif Tissue Int.
39:376–381. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bockman RS, Boskey AL, Blumenthal NC,
Alcock NW and Warrell RP Jr: Gallium increases bone calcium and
crystallite perfection of hydroxyapatite. Calcif Tissue Int.
39:376–381. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma Z and Fu Q: Comparison of the
therapeutic effects of yeast-incorporated gallium with those of
inorganic gallium on ovariectomized osteopenic rats. Biol Trace
Elem Res. 134:280–287. 2010. View Article : Google Scholar
|
|
28
|
Pei Y and Fu Q: Yeast-incorporated gallium
promotes fracture healing by increasing callus bony area and
improving trabecular microstructure on ovariectomized osteopenic
rats. Biol Trace Elem Res. 141:207–215. 2011. View Article : Google Scholar
|
|
29
|
Bedikian AY, Valdivieso M, Bodey GP,
Burgess MA, Benjamin RS, Hall S and Freireich EJ: Phase I clinical
studies with gallium nitrate. Cancer Treat Rep. 62:1449–1453.
1978.PubMed/NCBI
|
|
30
|
Krakoff IH, Newman RA and Goldberg RS:
Clinical toxicologic and pharmacologic studies of gallium nitrate.
Cancer. 44:1722–1727. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu J, Zhang B, He X, Zhang P and Chai Z:
Selection of a high-biomass, chromium-rich yeast strain and
optimization of cultivation conditions. J Ind Microbiol Biotechnol.
27:195–198. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Han C, Yuan J, Wang Y and Li L:
Hypoglycemic activity of fermented mushroom of Coprinus comatus
rich in vanadium. J Trace Elem Med Biol. 20:191–196. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Han C, Cui B and Wang Y: Vanadium uptake
by biomass of Coprinus comatus and their effect on hyperglycemic
mice. Biol Trace Elem Res. 124:35–39. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Verron E, Masson M, Khoshniat S, Duplomb
L, Wittrant Y, Baud'huin M, Badran Z, Bujoli B, Janvier P, Scimeca
JC, et al: Gallium modulates osteoclastic bone resorption in vitro
without affecting osteoblasts. Br J Pharmacol. 159:1681–1692. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yasuda H, Shima N, Nakagawa N, Yamaguchi
K, Kinosaki M, Mochizuki S, Tomoyasu A, Yano K, Goto M, Murakami A,
et al: Osteoclast differentiation factor is a ligand for
osteopro-tegerin/osteoclastogenesis-inhibitory factor and is
identical to TRANCE/RANKL. Proc Natl Acad Sci USA. 95:3597–3602.
1998. View Article : Google Scholar
|
|
36
|
Lacey DL, Timms E, Tan HL, Kelley MJ,
Dunstan CR, Burgess T, Elliott R, Colombero A, Elliott G, Scully S,
et al: Osteoprotegerin ligand is a cytokine that regulates
osteoclast differentiation and activation. Cell. 93:165–176. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Anderson DM, Maraskovsky E, Billingsley
WL, Dougall WC, Tometsko ME, Roux ER, Teepe MC, DuBose RF, Cosman D
and Galibert L: A homologue of the TNF receptor and its ligand
enhance T-cell growth and dendritic-cell function. Nature.
390:175–179. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wong BR, Rho J, Arron J, Robinson E,
Orlinick J, Chao M, Kalachikov S, Cayani E, Bartlett FS III,
Frankel WN, et al: TRANCE is a novel ligand of the tumor necrosis
factor receptor family that activates c-Jun N-terminal kinase in T
cells. J Biol Chem. 272:25190–25194. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
American Society for Bone and Mineral
Research President's Committee on Nomenclature: Proposed standard
nomenclature for new tumor necrosis factor family members involved
in the regulation of bone resorption. J Bone Miner Res.
15:2293–2296. 2000. View Article : Google Scholar
|
|
40
|
Li Y, Toraldo G, Li A, Yang X, Zhang H,
Qian WP and Weitzmann MN: B cells and T cells are critical for the
preservation of bone homeostasis and attainment of peak bone mass
in vivo. Blood. 109:3839–3848. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hofbauer LC and Schoppet M: Clinical
implications of the osteoprotegerin/RANKL/RANK system for bone and
vascular diseases. JAMA. 292:490–495. 2004. View Article : Google Scholar : PubMed/NCBI
|