Introduction
Obesity, specifically the enlargement of visceral
adiposity, is considered the major risk factor for the development
of insulin resistance, a characteristic feature of diabetes and
metabolic syndrome (1), which is
often characterized by nonalcoholic fatty liver disease (NAFLD)
(2). The metabolic pathways
leading to hepatic steatosis include enhanced non-esterified fatty
acid released from adipose, increased de novo lipogenesis
(DNL), decreased β-oxidation and reduced very low density
lipoprotein (VLDL) export (3).
Adipocytes posses the full complements of enzymes
and regulatory factors required to execute DNL and lipolysis, and
the two tightly controlled biochemical processes determine the rate
of lipid metabolism (4).
Sterol-regulatory-element-binding protein-1c (SREBP 1c), a key
regulator for lipid metabolism that is involved in adipocyte
differentiation, is expressed at high levels in the adipose tissue
and stimulates the expression of several lipogenic genes, including
FAS, acetyl-CoA carboxylase (ACC), stearyl-CoA desaturase 1 (SCD 1)
and lipoprotein lipase (LPL) (4).
LPL is the rate-limiting enzyme for the import of triglyceride
(TG)-derived fatty acids from VLDLs or chylomicrons for storage by
the adipose tissue (5). TG
synthesis and storage in the adipose tissue are important in
maintaining metabolic homeostasis (6). The first committed step in TG
synthesis via the glycerol phosphate pathway is mediated by
glycerol-3-phosphate acyltransferase (GPAT) enzymes (7). An additional fatty acid is
subsequently transferred to lysophosphatidic acid by the family of
1-acylglycerol-3-phosphate acyltransferase (AGPAT) enzymes to
produce phosphatidate, which serves as a precursor of acidic
phospholipids or diacylglycerol (DAG) (8). The DAG is converted to TG through the
action of diacylglycerol acyltransferase (DGAT) enzymes (9).
The hormonal regulation of lipolysis in adipocytes
provides a main switch between lipid storage and lipid mobilization
in response to dietary requirements. Hormone-sensitive lipase (HSL)
is activated and translocated to the lipid droplet surface, where
it interacts with specific lipid droplet proteins, including
perilipin and fat-specific protein-27 (FSP-27), which are regulated
by peroxisome proliferator-activated receptor (PPAR)γ (10). Of note, HSL works in concert with
other lipases, including adipose triglyceride lipase (ATGL), to
maximize the lipolytic output (11). Among the factors that contribute to
enhanced lipolysis associated with obesity, tumor necrosis factor
(TNF)α and adipocyte size appear to be the most relevant. TNFα,
secreted from the macrophages and adipocytes within the adipose
tissue of obese humans and animals, chronically stimulates
lipolysis (12). Monocyte
chemoattractant protein (MCP-1) is produced at high levels in obese
fat pads and, therefore, attracts a higher number of macrophages
(13). Taken together, obesity,
particularly the excess accumulation of fat in intra-abdominal
depots, causes and/or exacerbates metabolic disorders,
independently and in association with other diseases (14). Obesity is associated with an
increased risk of hepatic steatosis, and the incidence of steatosis
is correlated with the degree of obesity (15).
Fructus xanthii (FX), termed Cang-Erzi in
Chinese Pin Yin, was first recorded in Qian Jin dietetic therapy,
and is commonly used as a traditional Chinese medicine for treating
sinusitis and headache due to rheumatism (16). Data from animal experiments have
confirmed that FX possesses antioxidant, antinociceptive and
anti-inflammatory properties, and can protect pancreatic β-cells
form cytokine-induced damage (17,18).
In our previous study, it was observed that FX attenuates
HFD-induced heptic steatosis, suppresses fatty acid β-oxidation and
upregulates the expression levels of inflammatory genes in the
liver (19). Steatosis is the
result of ectopic lipid accumulation in the liver, and contributes
to liver-specific diseases (20).
Adipose tissue is the lipid storage organ, thus the dysfunctional
lipid storage in adipocytes is a sentinel event in the progression
toward metabolic disorder in HFD-induced obesity (21). The forced expansion of adipose
tissue prevents metabolic disease, despite gross obesity (22), which supports the hypothesis that
lipid 'spill-over' from fat promotes metabolic disease by fostering
ectopic lipid deposition (21).
The dysfunction of adipose tissue may trigger or exacerbate lipid
accumulation in the liver. The present study aimed to investigate
the role of FX in the regulation of lipid metabolism and storage in
EF in order to determine the role of FX in NAFLD.
Materials and methods
Preparation of FX
The raw material of FX was provided by the
Department of Pharmacy of the First Affiliated Hospital of Xiamen
University, (Xiamen, China). The herbs were immersed in a 10X
volume of water (V/V) for 30 mins, and then decocted twice at
boiling temperature for 2 h. The decocted liquids were collected,
filtrated, concentrated to 114 mg/ml, and were then placed in a
sterile bottle (30 ml) and stored in a refrigerator. Fingerprint
analysis of FX was performed using high performance liquid
chromatography from three different batches of FX preparation. All
chemicals were of analytical grades. Caffeic acid and chlorogenic
acid were purchased from Sigma-Aldrich (St. Louis, MO, USA). 3,
4-Dihydroxybenzoic acid, neochlorogenic acid, isochlorogenic acid
C, cynarin and 4-dicaffenolyquinic acid were obtained from
Sigma-Aldrich.
HPLC separation was performed on C18 column (250×4.6
mm3; 5 µm). The mobile phase consisted of solvent
A (methanol) and solvent B (water containing 0.2% formic acid). The
gradient elution program was as follows: 0–20 min, 2–5% A; 20–30
min, 5–8% A; 30–45 min, 8–15% A; 45–55 min, 15–25%; 55–60 min, 25%
A; 60–70 min, 25–35% A; 70–75 min, 35–38% A; 75–95 min, 38–65% A;
and 95–105 min. A was isocratic at 65%. The UV wavelength was set
at 260 nm, column temperature was kept at 25°C and the flow rate
was set at 1.0 ml/min. Three batches of FX were detected under the
same conditions. The results showed the same retention time and
similar peak area, indicating that the composition of the FX
decoction was consistent and reproducible (19).
Animals and experimental diets
All animal experiments were approved by the Animal
Care and Use Committee of Xiamen University. Fifty male
Sprague-Dawley rats (Shanghai Experimental Animal center, Chinese
Academy of Sciences), (age, 6 weeks; weight, 180–200 g) were housed
in standard cages in a temperature (22±1°C) and humidity
(50–60%)-controlled room, and maintained in a 12 h light/dark
cycle. All the rats were provided with unrestricted rodent chow and
water. Following acclimatization for 1 week, the rats were
separately fed a normal chow diet (NCD; control) or a high fat diet
(HFD) for 6 weeks. The animals were then subdivided into the
following four groups (n=10): NCD group; HFD group, in which
animals received equivalent volume of vehicle; HFD+FX (570
mg/kg/day); and HFD+FX (1,140 mg/kg/day) groups, in which the
animals in were treated with FX for another 6 weeks. At the end of
the treatment period, the rats were anesthetized with pentobarbital
(40 mg/kg body weight; Sigma-Aldrich); 5–7 ml blood was collected,
and EF samples were removed and stored at −80°C until analysis.
Following tissue harvest, the rats were sacrificed by
exsanguination while under anesthesia with pentobarbital (40 mg/g
body weight). The study was approved by the ethics committee of
Xiamen University (Xiamen, China; SYXK 2013-0006).
Free fatty acid (FFA) assay
The blood samples were centrifuged at 3,000 x g for
5 min, and the plasma was separated and transferred to Eppendorf
tubes for storage at −80°C prior to analysis. The concentration of
FFA in the plasma was measured using a Free Fatty Acid
Quantification kit (Abcam, Cambridge, UK).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from the subcutaneous fat
(SCF) and EF using an RNAsimple Total RNA kit (Tiangen Biotech Co.,
Ltd., Beijing, China), according to the manufacturer's protocol.
Isolated RNA was quantified using a NanoDrop ND-1000
spectrophotometer (NanoDrop Technologies; Thermo Fisher Scientific,
Inc., Waltham, MA, USA).
The reverse transcription of total RNA into cDNA was
performed using a FastQuanti RT kit with gDNAse (Tiangen Biotech
Co., Ltd.), according to the manufacturer's protocol. qPCR was
conducted in a total volume of 20 µl (2X 10 µl,
SuperReal PreMix, 50X 0.4 µl ROX Reference Dye, 2 µl
DNA template, 6 µl RNase-Free ddH2O, 0.8
µl forward primer and 0.8 µl reverse primer. The
reaction was performed as follows: 15 min at 95°C, 40 cycles of
95°C for 10 sec, 60°C for 20 sec, 72°C for 20 sec and 15 sec
incubation at 95°C. Gene expression analysis was performed using
the comparative quantification cycle (Cq) method (23). Amplification of the β-actin
sequence was performed in parallel and was used to normalize values
obtained for target genes. The primers (Shanghai Life Biotechnology
Co., Ltd., Guangzhou, China) used are listed in Table I.
 | Table IOligonucleotide primers used for
reverse transcription-quantitative polymerase chain reaction
analysis. |
Table I
Oligonucleotide primers used for
reverse transcription-quantitative polymerase chain reaction
analysis.
| Gene | Forward primer | Reverse primer |
|---|
| ChREBP |
5′-CTCGTGCAGGTCATCAAGAA-3′ |
5′-CAGCCCTCTTCATCTCCAAG-3′ |
| SREBP-1c |
5′-GGAGCCATGGATTGCACATT-3′ |
5′-CCTGTCTCACCCCCAGCATA-3′ |
| LXR |
5′-TCAGCATCTTCTCTGCAGACCGG-3′ |
5′-TCATTAGCATCCGTGGGAACA-3′ |
| PPAR-α |
5′-CTCGTGCAGGTCATCAAGAA-3′ |
5′-CAGCCCTCTTCATCTCCAAG-3′ |
| FAS |
5′-CACAGCATTCAGTCCTATCCACAGA-3′ |
5′-CACAGCCAACCAGATGCTTCA-3′ |
| ACC1 |
5′-GGACAGACTGATCGCAGAGAAAG-3′ |
5′-TGGAGAGCCCCACACACA-3′ |
| SCD-1 |
5′-CCTTAACCCTGAGATCCCGTAGA-3′ |
5′-AGCCCATAAAAGATTTCTGCAAA-3′ |
| CPT-1 |
5′-AATTGCAGTGGTATTTGAAGCTAAAA-3′ |
5′-GATATATTCTTCCCACCAGTCACTCA-3′ |
| ACO |
5′-GATTCAAGACAAAGCCGTCCAG-3′ |
5′-TCCACCAGAGCAACAGCATTG-3′ |
| ACOX1 |
5′-GTTGATCACGCACATCTTGGA-3′ |
5′-TCGTTCAGAATCAAGTTCTCAATTTC-3′ |
| C/EBP |
5′-TTACAACAGGCCAGGTTTCC-3′ |
5′-GGCTGGCGACATACAGTACA-3′; |
| PPARγ |
5′-ATTCTGGCCCACCAACTTCGG-3′ |
5′-TGGAAGCCTGATGCTTTATCCCCA-3′ |
| GPAT |
5′-TGATCAGCCAGGAGCAGCTG-3′ |
5′-AGACAGTATGTGGCACTCTC-3′ |
| AGPAT |
5′-GCATTTCAGGATCTCGTTCACA-3′ |
5′-ATCAACCCAACGAGAGCACTTT-3′ |
| DPAT |
5′-AGACTAGGAGGAGTGTGCAGGC-3′ |
5′-CGCTTCTTCCAAGGGAACTATG-3′ |
| ATGL |
5′-CACTTTAGCTCCAAGGATGA-3′ |
5′-TGGTTCAGTAGGCCATTCCT-3′ |
| HSL |
5′-CGCCTTACGGAGTCTATGC-3′ |
5′-TCTGATGGCTCTGAGTTGC-3′ |
| LPL |
5′-CAGCTGGGCCTAACTTTGAG-3′ |
5′-CCTCTCTGCAATCACACGAA-3′ |
| CD36 |
5′-CGGCGATGAGAAAGCAGAA-3′ |
5′-CAACCAGGCCCAGGAGC-3′ |
| perlipin |
5′-AGAGGAGACAGATGAGGAGGAAG-3′ |
5′-AGATGGTGTTCTGCAGAGTCTTC-3′ |
| FSP27 |
5′-CCTTTCCCAGAAGCCAACT-3′ |
5′-AGAGTCCAGCGGAGCATT-3′ |
| MCP1 |
5′-AATGGGTCCAGAAGTACATTAGAAA-3′ |
5′-GGTGCTGAAGTCCTTAGGGTTG-3′ |
| TNF-α |
5′-ACACCATGAGCACGGAAAGC-3′ |
5′-CCGCCACGAGCAGGAA-3′ |
| IL-1β |
5′-AATGGACAGAACATAAGCCAACA-3′ |
5′-CCCAAGGCCACAGGCAT-3′ |
| CD68 |
5′-TAGTTCTTTCTCCAGCAATTCACC-3′ |
5′-CCCGAAGTGTCCCTTGTCC-3′ |
| β-actin |
5′-CGGTCAGGTCATCACTATCG-3′ |
5′-TTCCATACCCAGGAAGGAAG-3 |
Quantitation of adipocyte size
The EF samples were fixed with 4% paraformaldehyde
(Sigma-Aldrich) and embedded in paraffin (Sigma-Aldrich). Standard
4 µm thick sections were stained with hematoxylin and eosin
(Sigma-Aldrich). Adipocyte size, which was determined as the
average adipocyte area (in µm2) was measured
using Image J software (National Institutes of Health, Bethesda,
MD, USA). Adipocyte sizes were measured from EF samples from 10
rats in each group.
Immunohistochemistry
Tissue sections were deparaffinized with xylene
(Sinopharm Chemical Reagent Co., Ltd., Shanghai, China) and
stepwise rehydrated with serial dilutions of ethanol. For epitope
retrieval, slides were incubated in 0.01 M citric acid buffer
(Sinopharm Chemical Reagent Co., Ltd.) at 100°C for 2 min.
Endogenous horseradish peroxidase activity was quenched by
incubation with 3% hydrogen peroxide Sinopharm Chemical Reagent
Co.,Ltd, Shanghai, China for 10 min. The tissue sections were then
blocked in goat serum and incubated with monoclonal rabbit anti-rat
primary antibody against F4/80 (1:100; cat. no. ab111101; Abcam)
diluted in blocking buffer at 4°C overnight. Incubation with
secondary antibodies was performed using an Ultrasensitive SP IHC
kit (Maixin, Fuzhou, China) and detected using 3,
3′-diaminobenzidine (Solarbio, Beijing, China). Each step was ended
by washing in buffer for 5 min twice. All sections were
counterstained with hematoxylin prior to dehydration and coverslip
placement. From each animal, more than five tissue sections,
including representative sections, were analyzed. The images were
captured by a Leica microscope (DMI3000B, Leica Microsystems, Inc.,
Buffalo Grave, IL, USA).
Statistical analysis
In the present study, all results are presented as
the mean ± standard error of the mean. Statistical analyses were
conducted using GraphPad Prism 5 (GraphPad Software Inc., La Jolla,
CA, USA) Individual differences between the HFD and FX group were
assessed using one-way analysis of variance or Student's t-test,
when applicable. P<0.05 was considered to indicate a
statistically significant difference.
Results
FX decreases serum levels of FFA
The HFD group were found to have increased serum
concentrations of FFA, which were reversed by FX treatment at low
and high doses (P<0.05) as shown in Fig. 1, suggesting a decrease in the
release of FFA from the adipose tissues.
FX increases adipocyte size in the
EF
Metabolic improvements are generally associated with
a reduction in average fat mass and cell size (24). In the present study, histological
examination of EF revealed that the average adipocyte size in the
HFD group was markedly increased relative to that of the NCD
adipocytes, the number of adipocytes in the HFD group was lower
than that in the NCD group, and a high dose of FX increased
adipocyte number (Fig. 2A and B).
Quantitative assessment of adipocyte size distribution clearly
demonstrated this shift toward smaller size adipocytes areas in the
high dose FX group (Fig. 2C). The
AUC of the adipocyte size showed that FX treatment at a high dose
increased the AUC of 1,000–25,000 µm2, and
decreased the AUC of 25,000–50,000 µm2 and
>50,000 µm2 (P>0.05; Fig. 2D).
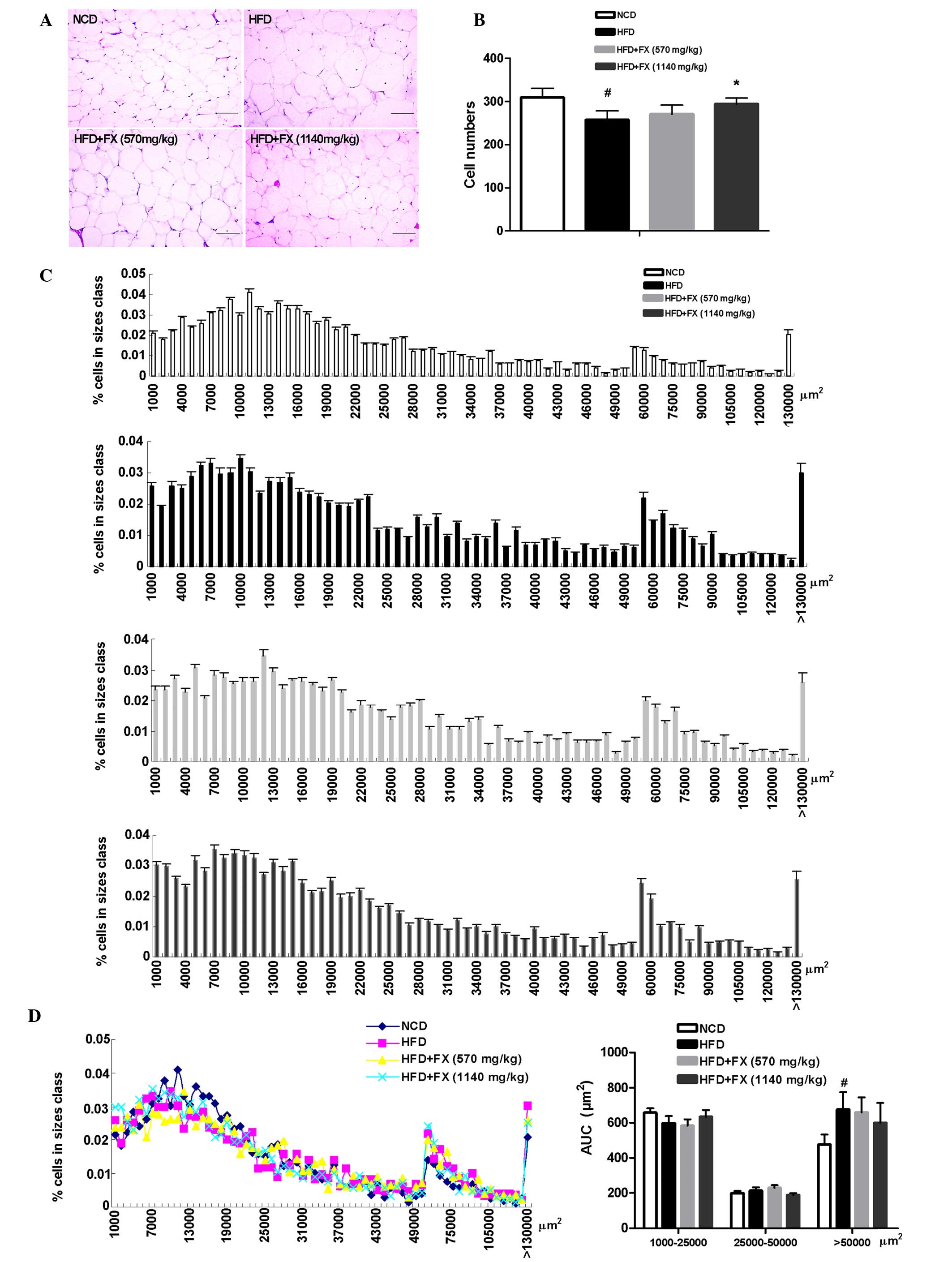 | Figure 2Effect of FX on epididymal adipose
tissue mass. (A) Histological analysis of epididymal adipose
tissue. Scale bar, 200 µm (B) Adipocyte numbers. (C)
Distribution of adipocyte sizes. (D) Different size and AUC of
adipocytes: Small, 1000–25000 µm2; medium,
25,000–50,000 µm2; and large, >50,000
µm2. Data are presented as the mean ± standard
error of the mean (n=10). #P<0.05, vs. NCD group;
*P<0.05, vs. HFD group. FX, Fructus xanthii;
NCD, normal chow diet; HFD, high fat diet. AUC, area under the
curve. |
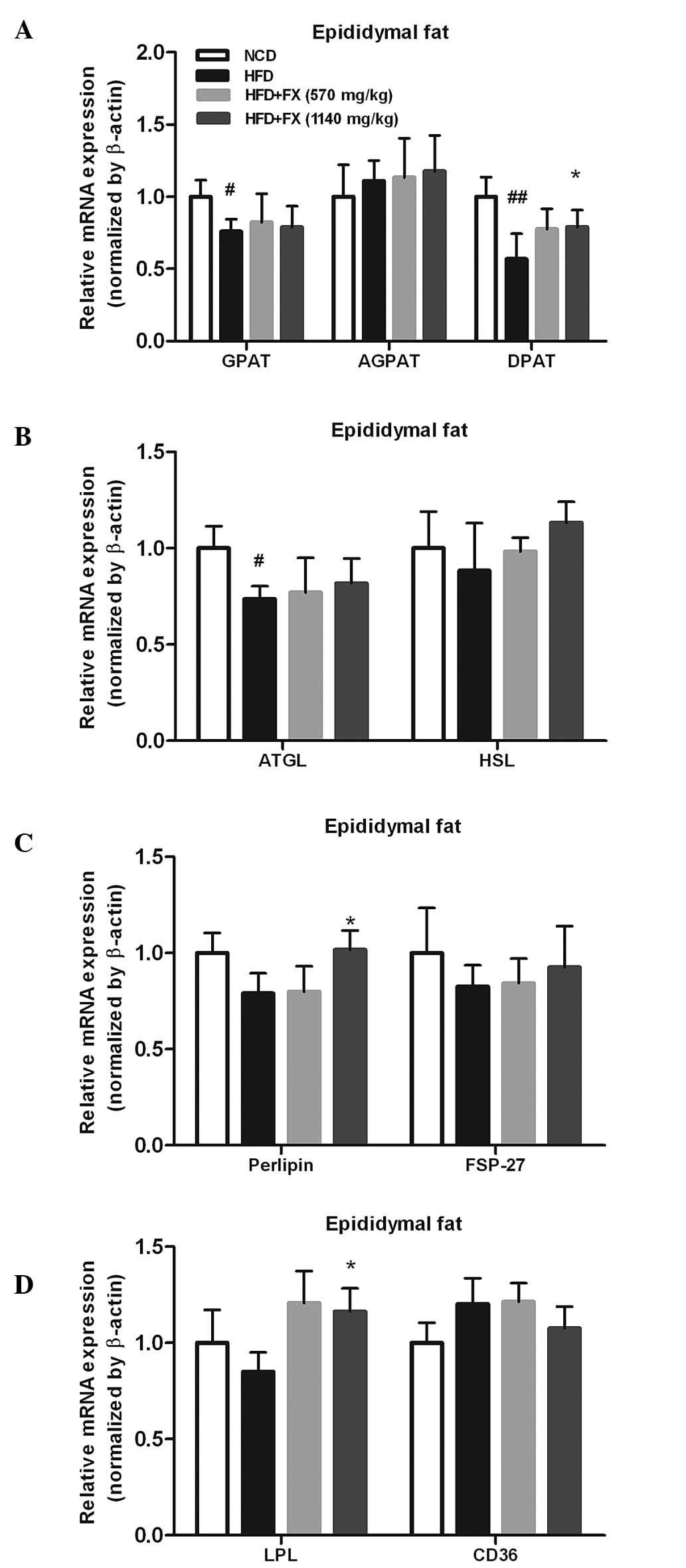
Effect of FX on lipid metabolism in the
EF
The increased release of FFA from adipose tissue has
long been linked to lipogenesis and steatosis in the liver
(25). In the EF, a HFD increased
the mRNA expression levels of carbohydrate-responsive
element-binding protein (ChREBP) and SCD1 (P<0.05), and
decreased the levels of SREBP 1c, FAS and acetyl-CoA carboxylase
(ACC1; P<0.05), which were essentially reversed by FX.
Furthermore, FX at a high dose increased the gene expression level
of SCD1 significantly (P<0.05), as shown in Fig. 3. However, treatment with FX had no
effect on the expression of fatty acid oxidation-associated genes
(carnitine palmitoyltransferase 1, acyl-CoA oxidase and acyl-CoA
oxidase 1) or the transcription factor regulating their activities,
PPARα. These results demonstrated a role of FX in promoting
lipogenesis in EF.
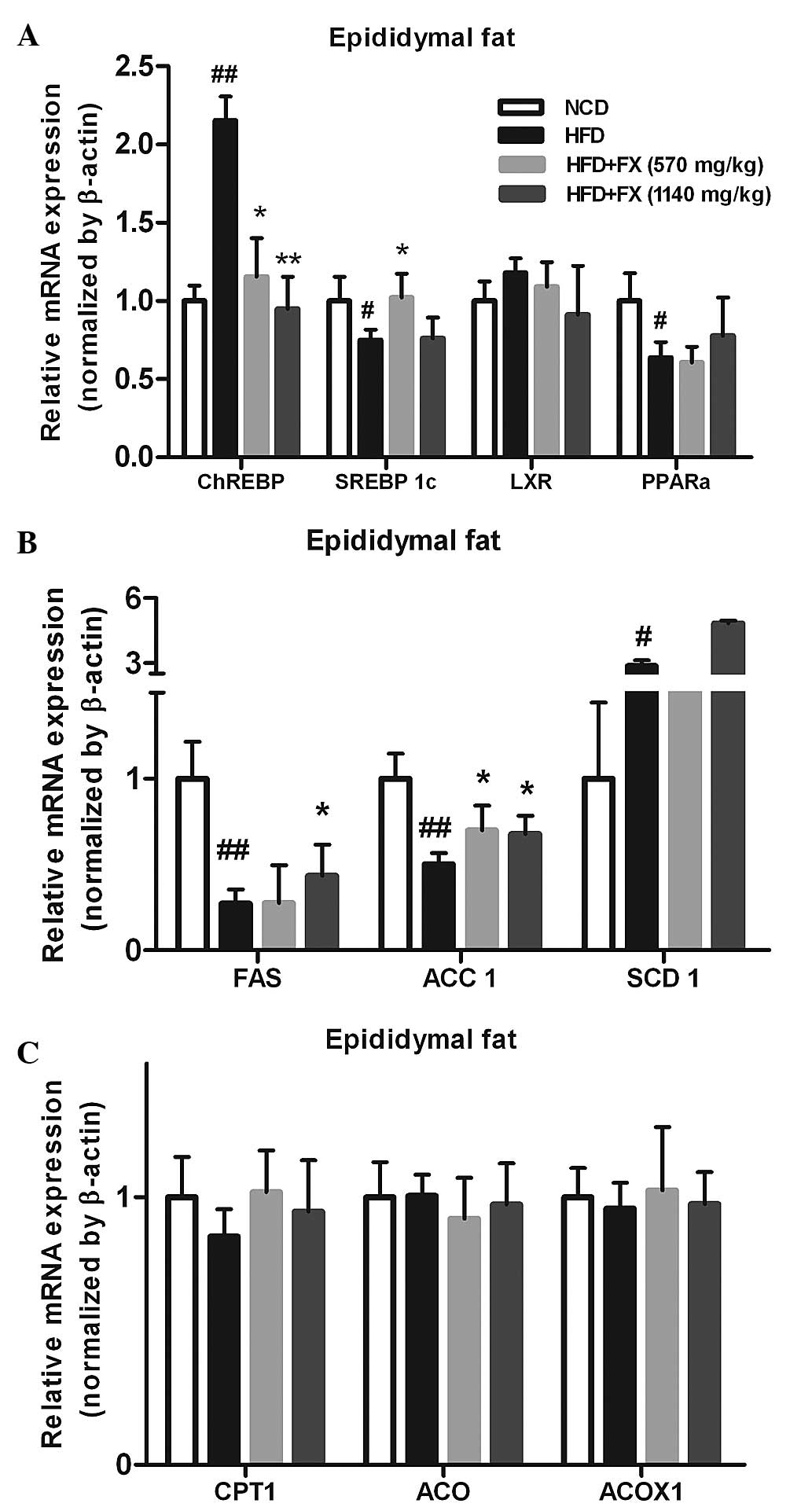 | Figure 3Effect of FX on the expression levels
of lipogenic genes in the EF. (A) Expression levels of lipogenic
genes; (B) Expression levels of antioxidant genes; (C) Expression
levels of lipolytic genes. Data are presented as the mean ±
standard error of the mean (n=10). #P<0.05 and
##P<0.01, vs. NCD group; *P<0.05 and
**P<0.01, vs. HFD group. FX, Fructus xanthii;
NCD, normal chow diet; HFD, high fat diet; ChREBPl,
carbohydrate-responsive element-binding protein; SREBP1c,
sterol-regulatory-element-binding protein-1c; LXR, liver X
receptor; PPARα, peroxisome proliferator-activated receptor α;
ACC1, acetyl-CoA carboxylase; SCD1, stearyl-CoA desaturase 1; CPT1,
carnitine palmitoyltransferase 1; ACP, acylacyl carrier protein;
ACOX, acyl-CoA oxidase; ACOX1, acyl-CoA oxidase 1. |
FX switches TG hydrolysis to TG
storage
The expression of the TG synthesis-associated genes,
GPAT and DPAT, in the HFD group were significantly lower, compared
with those in the NCD group (P<0.05). However, a high dose of FX
supplementation significantly increased the mRNA expression of DPAT
(P<0.05). No differences were observed in the level of AGPAT
among the groups, as shown in Fig.
4A. The mRNA levels of TG lipolysis-associated genes (ATGL and
HSL) treated by FX did not differ from the levels in the HFD group,
however,he level of ATGL in the HFD group was lower than that in
the NCD group (P<0.05), as shown in Fig. 4B.
In addition, the expression of perilipin, involved
in the lipolytic process, was significantly increased (P<0.05),
whereas, FSP-27, another lipid droplet-associated protein, was not
modified by FX (Fig. 4C). Fatty
acid uptake and transport are important functions for adipose fat
storage, and the fatty acid import and transport-associated genes
(LPL and CD36) were marginally affected by HFD and NCD. High
dose-FX treatment elevated the expression of LPL significantly
(P<0.05; Fig. 4D).
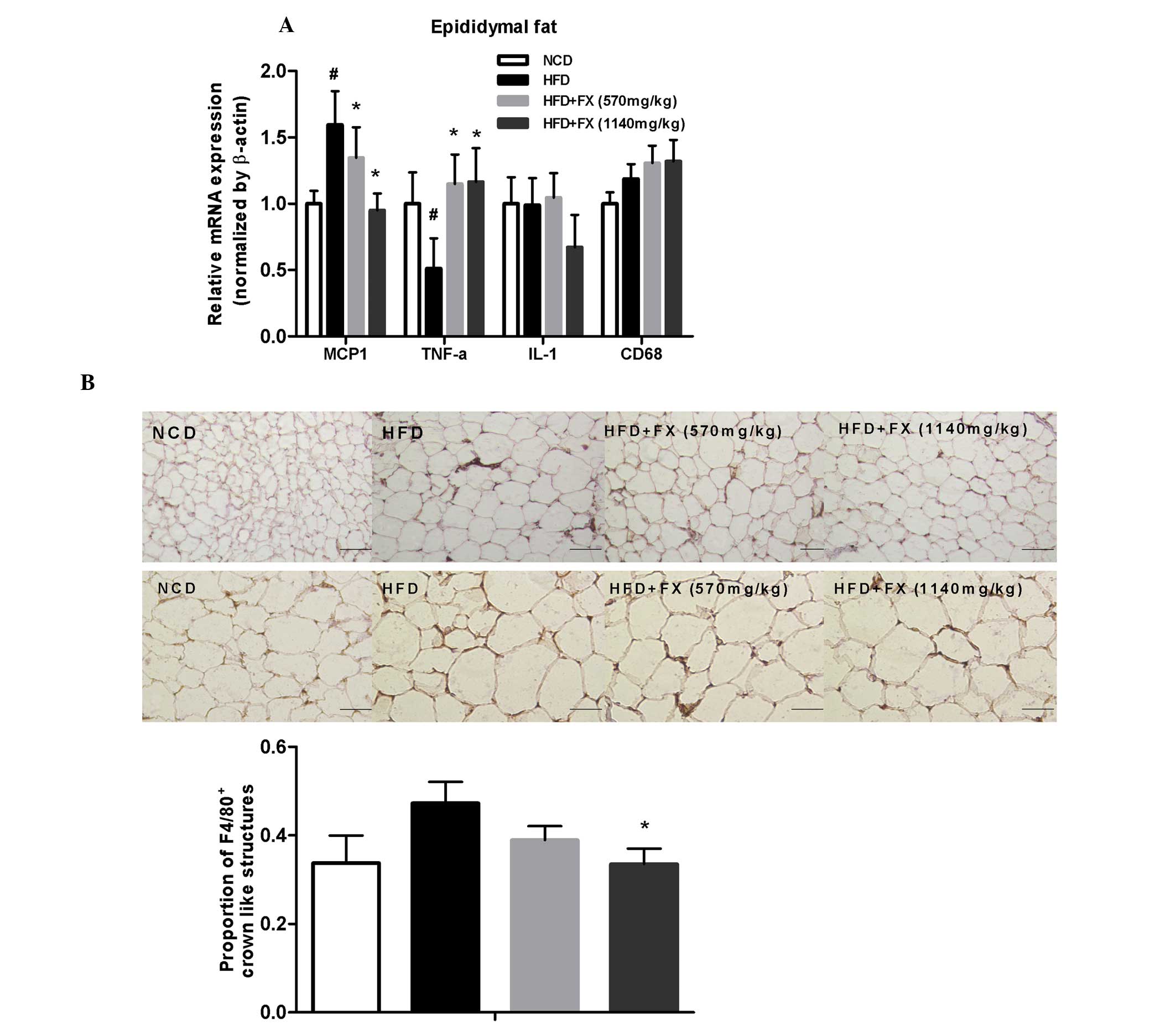
FX decreases inflammation in EF
Compared with the NCD group, genes of inflammatory
factors, including MCP1 and CD68, were upregulated in the HFD
group, whereas the expression of TNF-α was decreased. FX decreased
the expression of MCP1 in a dose-dependent manner (P<0.05), and
increased the expression of TNF-α significantly (P<0.05), as
shown in Fig. 5A.
Macrophages are stained brown. In the NCD group,
F4/80+ staining were uniformly small, dispersed, and
rarely seen in aggregates. The fraction of F4/80+
staining was increased and in formed aggregates in the HFD group.
FX treatment at a high dose decreased the degree of
F4/80+ staining, as shown in Fig. 5B. Immunohistochemical analysis
showed the total number of F4/80+ crown-like structures
were higher in the HFD group, compared with the NCD group, although
without statistical significance. FX treatment at a high dose
suppressed the number of F4/80+ crown-like structures
(P<0.05), as shown in Fig.
5B.
Discussion
Our previous study demonstrated that FX decreases
fatty acid synthesis and lipid accumulation in the liver, thus
attenuating hepatic steatosis in HFD rats via inhibiting
lipogenesis and promoting lipolysis mechanisms, which are
associated with the inhibition of ChREBP and SREBP 1c, and the
induction of PPARα. This suggested the potential application of FX
in treating fatty liver diseases (19). In the present study, it was
demonstrated that FX increased the synthesis of fatty acid and TG,
thus decreasing the circulating FFA levels, suggesting that FX is
involved in addressing the abnormality of FFA in the circulation,
which is executed by promoting the storage of the excess fat,
rather than the elimination of added fat.
Significant evidence has indicated the ectopic
deposition of adipose tissue with respect to its enhanced risk for
metabolic complications (21).
Adipose tissue stores TG in a positive energy balance condition.
Disorder in adipose fat storage function may cause excess fatty
acid influx into the liver, leading to steatosis (26). Adipose metabolism acts to buffer
nutrient excess in promoting lipid storage with increased
adipogenesis and lipogenesis (27). The quantity of TG accumulation in
adipose tissue results from the balance of several metabolic
pathways, which take place in the tissue, including DNL, fatty acid
uptake from circulating TG and lipid mobilization (28).
In the present study, HFD induced hepatic steatosis
and high levels of circulating FFA (Fig. 1). Elevated circulating FFA
concentration is associated with the accumulation of TG in the
liver. Ectopic lipid deposition in the liver (fatty liver) reflects
a diminished capacity of adipocytes to store fatty acids, either
due to impaired lipogenesis or a diminished capacity for
hypertrophy or proliferation (29).
Evidence suggests that one factor, which links
excess caloric intake and a positive energy balance with insulin
resistance is the inability to appropriately expand the adipose
tissue (22). To examine whether
the EF is associated with the amelioration of HFD-induced metabolic
dysfunction, the present study examined morphological changes, and
the sizes and numbers of adipocytes in the EF. HFD-induced fat
accumulation in EF was significantly ameliorated, and adipocyte
size was markedly reduced in the FX-treated group, in which the
distribution of adipocyte size was shifted to smaller sized
adipocytes (Fig. 2), showing that
FX reduces the burden of HFD, predominantly due to appropriate
extended EF, thus reducing fatty acid release from the EF overflux
into the liver, and slowed the development of fatty liver.
No significant difference in EF weight was observed
between the HFD and FX-treated groups (data not shown). It has
become evident that, in addition to absolute fat quantity,
qualitative aspects of adipose tissue function and cellular
composition have an important effect on the systemic metabolic
phenotype (22). Therefore,
determined RT-qPCR was used to determine the expression levels of
relevant genes involved in lipogenic transcription factors, fatty
acid synthesis and esterification, oxidation, TG synthesis
enzymes.
DNL in white adipose tissue is considered to occur
to a similar extent in the liver (30). In the EF, following HFD, the levels
of ChREBP and SCD1 were increased significantly, and FX normalized
the high expression level of ChREBP induced by HFD; with the effect
of FX at two doses on the level of SCD1 showed the opposite.
However, the expression of SREBP 1c was reduced by HFD, together
with its target enzymes, FAS and ACC1. FX treatment elevated the
expression levels of the three genes significantly, suggesting
that, during HFD, lipogenesis of adipose tissue is disordered and
causes liver ectopoic lipid deposition (19), and that FX treatment increased
fatty acid synthesis, which are critical substrates for the
biosynthesis of TG (Fig. 3). These
data demonstrated that HFD disturbed the major lipid metabolic
pathways in adipose tissue, and FX had regulatory effects suitable
to the local situation. Above all, a concomitant increase in the
rate of fatty acid synthesis compensated for the increase in fatty
acid utilization in EF, preventing an increase in circulating FFA
concentrations.
On examination of the genes involved in catalyzing
the steps of TG synthesis, the effect of FX on DGAT was determined,
which converts diacylglycerol to TG. However, FX had no effect on
the expression levels of GPAT or AGPAT. The expression levels of
ATGL and HSL, which are two major TG hydrolases responsible for
adipose lipolysis, were not significantly affected by FX.
Furthermore, the expression of LPL following treatment with FX was
increased, compared with that of the HFD group; this indicated a
clear effect of FX on fatty acid uptake from the circulating TG
(Fig. 4). These above data showed
that FX transported more fatty acids from the circulation, did not
increase the hydrolysis of TG in adipose tissue, and marginally
increased the lipogenesis of TG. Therefore, adipose tissue in the
FX-treated group buffered HFD-induced TG overproduction, which
attenuated liver lipid deposition.
Within a certain range, adipose tissue extension is
more conducive to increased fat storage. In addition to this, the
number of necrotic adipocytes positively correlates with average
adipocyte size in obese mice and other mouse models of adipocyte
hypertrophy (31). The death of
hypertrophic adipocytes facilitates the infiltration of
macrophages, which further perpetuates adipose inflammation and
insulin resistance (32). In the
present study, high levels of MCP1 and low levels of TNF-α were
observed in the HFD group, and despite the controversial results,
treatment with FX was shown to attenuate the HFD-induced
inflammatory changes. Macrophage infiltration in the EF of the HFD
group was marked, indicated by F4/80+ staining in brown,
however, treatment with FX decreased the staining to differing
degrees (Fig. 5). These results
suggested that FX limited the inflammatory infiltration, inhibited
the vicious cycle of increased inflammation and dysfunction, and
maintained the function of EF.
In conclusion, adipose lipid homeostasis is
basically dependent on two major functions: TG/fatty acid uptake
and lipolysis/fatty acid release (33). FX supplementation reversed NAFLD in
association with a reduction in the number of circulating FFA,
conveying more FFA from the circulation, and partially increasing
the expression levels of the fatty acid and TG synthesis-associated
genes in the EF. These data suggest that FX ameliorated HFD-induced
lipid dyshomeostasis at the adipose tissue-liver axis.
Acknowledgments
This study was supported by the Xiamen Science and
Technology Bureau (Xiamen Research Platform for Systems Biology of
Metabolic Disease; grant no. 3502Z20100001); the National Natural
Science Foundation to Dr Shuyu Yang (grant no. 30973912), Dr Xuejun
Li (grant no. 81073113), and Dr Suhuan Liu (grant no. 81270901);
and the education scientific research project of young teachers in
Fujian Province (grant no. JA15306)
References
|
1
|
Handschin C and Spiegelman BM: The role of
exercise and PGC1 alpha in inflammation and chronic disease.
Nature. 454:463–469. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Duval C, Thissen U, Keshtkar S, Accart B,
Stienstra R, Boekschoten MV, Roskams T, Kersten S and Müller M:
Adipose tissue dysfunction signals progression of hepatic steatosis
towards nonalcoholic steatohepatitis in C57Bl/6 mice. Diabetes.
59:3181–3191. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Postic C and Girard J: The role of the
lipogenic pathway in the development of hepatic steatosis. Diabetes
Metab. 34(6 Pt 2): 643–648. 2008. View Article : Google Scholar
|
|
4
|
Seo JB, Moo HM, Kim WS, Lee YS, Jeong HW,
Yoo EJ, Ham J, Kang H, Park MG, Steffensen KR, et al: Activated
liver X receptors stimulate adipocyte differentiation through
induction of peroxisome proliferator-activated receptor gamma
expression. Mol Cell Biol. 24:3430–3444. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marcelin G and Chua S Jr: Contributions of
adipocyte lipid metabolism to body fat content and implications for
the treatment of obesity. Curr Opin Pharmacol. 10:588–593. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Takeuchi K and Reue K: Biochemistry,
physiology and genetics of GPAT, AGPAT and lipin enzymes in
triglyceride synthesis. Am J Physiol Endocrinol Metab.
296:E1195–E1209. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gimeno RE and Cao J: Thematic review
series: Glycerolipids. Mammalian glycerol-3-phosphate
acyltransferases: New genes for an old activity. J Lipid Res.
49:2079–2088. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shindou H, Hishikawa D, Harayama T, Yuki K
and Shimizu T: Recent progress on acyl-CoA: Lysophospholipid
acyltransferase research. J Lipid Res. 50(Suppl): S46–S51. 2009.
View Article : Google Scholar :
|
|
9
|
Yen CL, Stone SJ, Koliwad S, Harris C and
Farese RV Jr: Thematic review series: Glycerolipids. DGAT enzymes
and triacylglycerol biosynthesis. J Lipid Res. 49:2283–2301. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gong J, Sun Z, Wu L, Xu W, Schieber N, Xu
D, Shui G, Yang H, Parton RG and Li P: Fsp27 promotes lipid droplet
growth by lipid exchange and transfer at lipid droplet contact
sites. J Cell Biol. 195:953–963. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rodriguez-Cuenca S, Carobbio S, Velagapudi
VR, Barbarroja N, Moreno-Navarrete JM, Tinahones FJ, Fernandez-Real
JM, Orešic M and Vidal-Puig A: Peroxisome proliferator-activated
receptor γ-dependent regulation of lipolytic nodes and metabolic
flexibility. Mol Cell Biol. 32:1555–1565. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wellen KE and Hotamisligil GS:
Obesity-induced inflammatory changes in adipose tissue. J Clin
Invest. 112:1785–1788. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sartipy P and Loskutoff DJ: Monocyte
chemoattractant protein 1 in obesity and insulin resistance. Proc
Natl Acad Sci USA. 100:7265–7270. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kopelman PG: Obesity as a medical problem.
Nature. 404:635–643. 2000.PubMed/NCBI
|
|
15
|
Wanless LR and Lentz JS: Fatty liver
hepatitis and obesity: An autopsy study with analysis of risk
factors. Hepatology. 12:1106–1110. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ma P and Li H: The research development of
Fructus xanthii. Chinese Herbal Medicine. 30:634–636. 1999.
|
|
17
|
Huang MH, Wang BS, Chiu CS, Amagaya S,
Hsieh WT, Huang SS, Shie PH and Huang GJ: Antioxidant,
antinociceptive and anti-inflammatory activities of Xanthii Fructus
extract. J Ethnopharmacol. 135:545–552. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Song MY, Kim EK, Lee HJ, Park JW, Ryu DG,
Kwon KB and Park BH: Fructus Xanthii extract protects against
cytokine-induced damage in pancreatic beta-cells through
suppression of NF-kappaB activation. Int J Mol Med. 23:547–553.
2009.PubMed/NCBI
|
|
19
|
Li XM, Li ZP, Xue M, Ou ZM, Liu M, Yang M,
Liu S, Yang S and Li X: Fructus Xanthii attenuates hepatic
steatosis in rats fed on high-fat diet. Plos one. 8:e614992013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kato K, Takamura T, Takeshita Y, Ryu Y,
Misu H, Ota T, Tokuyama K, Nagasaka S, Matsuhisa M, Matsui O and
Kaneko S: Ectopic fat accumulation and distant organ-specific
insulin resistance in Japanese people with nonalcoholic fatty liver
disease. PLoS One. 9:e921702014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lument CN and Saltiel AR: Inflammatory
links between obesity and metabolic disease. J Clin Invest.
121:2111–2117. 2011. View
Article : Google Scholar
|
|
22
|
Kim JY, van de Wall E, Laplante M, Azzara
A, Trujillo ME, Hofmann SM, Schraw T, Durand JL, Li H, Li G, et al:
Obesity-associated improvements in metabolic profile through
expansion of adipose tissue. J Clin Invest. 117:2621–2637. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2[−Delta Delta C(T)] Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
Ginsberg HN, Zhang YL and Hernandez-Ono A:
Metabolic syndrome: Focus on dyslipidemia. Obesity (Silver Spring).
14(Suppl 1): S41–S49. 2006. View Article : Google Scholar
|
|
25
|
Sun X, Tang Y, Tan X, Li Q, Zhong W, Sun
X, Jia W, McClain CJ and Zhou Z: Activation of peroxisome
proliferator-activated receptor-γ by rosiglitazone improves lipid
homeostasis at the adipose tissue-liver axis in ethanol-fed mice.
Am J Physiol Gastrointest Liver Physiol. 302:G548–G557. 2012.
View Article : Google Scholar
|
|
26
|
Khan T, Muise ES, Iyengar P, Wang ZV,
Chandalia M, Abate N, Zhang BB, Bonaldo P, Chua S and Scherer PE:
Metabolic dysregulation and adipose tissue fibrosis: Role of
collage VI. Mol Cell Biol. 29:1575–1591. 2009. View Article : Google Scholar :
|
|
27
|
Hofbauer KG: Molecular patheays to
obesity. Int J Obes Relat Metab Disord. 26(Suppl 2): S18–S27. 2002.
View Article : Google Scholar
|
|
28
|
Alberdi G, Rodríguez V, Miranda J,
Macarulla M, Arias N, Andrés-Lacueva C and Portillo MP: Changes in
white adipose tissue metabolism induced by resveratrol in rats.
Nutr Metab (Lond). 8:292011. View Article : Google Scholar
|
|
29
|
Wang M, Grayburn P, Chen S, Ravazzola M,
Orci L and Unger RH: Adipogenic capacity and the susceptibility to
type 2 diabetes and metabolic syndrome. Proc Natl Acad Sci USA.
105:6139–6144. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Letexier D, Pinteur C, Large V, Fréring V
and Beylot M: Comparison of the expression and activity of the
lipogenic pathway in human and rat adipose tissue. J Lipid Res.
44:2127–2134. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Strissel KJ, Stancheva Z, Miyoshi H,
Perfield JW II, DeFuria J, Jlick Z, Greenberg AS and Obin MS:
Adipocyte death, adipose tissue remodeling and obesity
complications. Diabetes. 56:2910–2918. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang HR and Zhang CH: Adipose 'talk' to
distant organs to regulate insulin sensitivity and vascular
function. Obesity (Silver Spring). 18:2071–2076. 2010. View Article : Google Scholar
|
|
33
|
Large V, Peroni O, Letexier D, Ray H and
Beylot M: Metabolism of lipids in human white adipocyte. Diabetes
Metab. 30:294–309. 2004. View Article : Google Scholar : PubMed/NCBI
|