Introduction
Bone cancer pain is a type of severe and chronic
pain that affects the quality of life in cancer patients; however,
no breakthrough regarding the elucidation of underlying mechanisms
and the development of therapeutics for bone cancer pain has been
achieved (1). Studies have
indicated that nitric oxide (NO) is an important neurotransmitter
contributing to the development and maintenance of central and
peripheral sensitization in inflammatory (2) and neuropathic pain (3).
NO is synthesized from L-arginine via a family of
nitric oxide synthases (NOS), which are key enzymes in NO
biosynthesis and comprise neuronal NOS (nNOS), endothelial NOS
(eNOS) and inducible NOS (iNOS). The pathophysiological functions
of NO are regulated by the expression and activity of these
isoforms (4,5). nNOS, expressed in the nervous system,
produces NO in neurons and has been shown to contribute to spinal
nociceptive processing in several pain models (6–12).
iNOS, which is not constitutively expressed and must be induced to
be synthesized, is abundant in a variety of cell types, including
glial cells, macrophages, chondrocytes and neutrophils (13). Previous studies demonstrated that
iNOS is also involved in the mechanisms of inflammatory,
neuropathic pain (5,14) as well as herpetic allodynia
(15).
Bone cancer pain is thought to have inflammatory,
neuropathic and tumorigenic components. A previous study by our
group has shown that NR2B-containing N-methyl-d-aspartate (NMDA) receptor has a
critical role in spinal nociceptive processing during bone cancer
pain (16). Activation of NMDA
receptors allows Ca2+ to enter neurons. Increased
intracellular Ca2+ triggers a cascade of events that
includes activation of nNOS, leading to the production of NO in the
spinal cord (17). Another
previous study by our group also demonstrated that upregulation of
NR2B and nNOS expression in the spinal dorsal horn contributes to
hyperalgesia induced by chronic compression of dorsal root ganglia
(18). However, the role of NOS in
the development of bone cancer pain has remained elusive.
The present study used a murine model of
osteosar-coma-associated bone cancer pain, to explore whether NOS
in the spinal L3-L5 segments is responsible for the development of
bone cancer pain and to determine the efficacy of
NG-monomethyl-l-arginine (L-NMMA)
administration in alleviating bone cancer pain.
Materials and methods
Animals
Experiments were approved by the Animal Care and Use
Committee of the Medical School of Nanjing University (Nanjing,
China) and were in accordance with the guidelines for the use of
laboratory animals (19). The
numbers of animals and their suffering were minimized in all cases.
Male C3H/HeJ mice (n=100; weight, 20–22 g; 4–6 weeks old),
purchased from the Model Animal Research Center of Nanjing
University (Nanjing, China), were housed in a
temperature-controlled (21±1°C) room with a 12-h light/dark cycle
with access to food and water ad libitum. Each group used to
analyze behavior contained 8 mice, and each group in PCR or
immunohisto-chemistry study contained 5 mice. The mice subjected to
the behavioral studies were not processed for
PCR/immunohistochemistry, and were euthanized after the last
behavioral test.
Cell culture and tumor-cell
inoculation
The NCTC 2472 osteosarcoma cell line (no. 2087787;
American Type Culture Collection, Manassas, VA, USA) was used in
the present study. The cells were incubated in NCTC 135 medium
(Sigma-Aldrich, St. Louis, USA) containing 10% horse serum (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) in a humidified
atmosphere containing 5% CO2 at 37°C.
The mouse model of bone cancer was established
according to the method of previous studies by our and another
group (16,20). Briefly, 1% pentobarbital sodium (50
mg/kg; Sigma-Aldrich) in normal saline [Baxter Healthcare (Tianjin)
Co. Ltd., Tianjin, China] was administered to the mice by
intraperitoneal injection and the right femur condyle was
perforated with a 30-gauge needle (Jiangsu Zhengkang Medical
Equipment Co., Ltd., Changzhou, China). Subsequently,
2×105 NCTC 2472 cells in 20 µl α-minimum
essential medium (Thermo Fisher Scientific, Inc.) were injected
into the intra-medullary space of the femur. In the sham group, the
medium contained no cells. Subsequently, the injection hole was
sealed with dental amalgam (AT&M Biomaterials Co., Ltd.,
Beijing, China), followed by copious irrigation with normal saline
and closing of the wound.
In the interaction studies, mice were intrathecally
administered 50 µg L-NMMA (Cayman Chemical Company, Ann
Arbor, Michigan, USA) dissolved in artificial cerebrospinal fluid
(10 µg/µl; 5 µl per mouse; Harvard Apparatus,
Holliston, MA, USA) at post-operative day 14, at which
pain-associated behavior was observed. Mice in the sham group
received vehicle treatment. Intrathecal injections were performed
free-hand between the L5 and L6 lumbar space in unanesthetized male
mice according to the method of Hylden and Wilcox (21). Pain-associated behavior was
assessed prior to, as well as at 2, 12 and 24 h after L-NMMA
administration.
The mice assigned for behavioral tests were
sacrificed after the last measurement. The mice were deeply
anesthetized with pentobarbital (50 mg/kg, intraperitoneally) and
then sacrificed by cervical dislocation. The mice assigned for
Reverse-transcription quantitative polymerase chain reaction
(RT-qPCR) were individually sacrificed at post-operative days 7, 10
or 14, and the spinal cord segments were immediately removed. The
mice assigned for immunohistochemical staining were also deeply
anesthetized with pentobarbital individually at post-operative days
7, 10 or 14, and then transcardially perfused with saline followed
by 4% paraformaldehyde (PFA; Beijing Dingguo Changsheng
Biotechnology Co., Ltd., Beijing, China).
Assessment of pain-associated
behavior
Withdrawal thresholds as well as latency to
mechanical and thermal stimulation of mice were examined prior to
surgery as well as at post-operative days 3, 5, 7, 10 and 14. Mice
were allowed to acclimatize for at least 30 min prior to each
test.
Mechanical allodynia
The paw withdrawal mechanical threshold (PWMT) was
assessed according to the method by Chaplan et al (22), which was also used in a previous
study by our group (16). Briefly,
a set of von Frey filaments (0.16, 0.4, 0.6, 1.0, 1.4 and 2.0g;
Stoelting, Wood Dale, IL, USA) was used to vertically push against
the plantar surface of the right hind paw. Each mouse was tested
five times per stimulus force. The lightest von Frey filament that
evoked three or more brisk withdrawal or paw flinching reactions
was regarded as the PWMT.
Thermal hyperalgesia
The paw withdrawal thermal latency (PWTL) was
measured according to a method of a previous study by our group
(23) using a radiant thermal
stimulator (BME410A; Institute of Biological Medicine, Academy of
Medical Science, Tianjin, China) that focused onto the plantar
surface of the hind paw. The PWTL was defined as the latency of the
mice to lift or lick their hind paw. Each mouse was tested five
times with a 5-min interval. A cut-off time of 20 sec was used to
avoid tissue damage. The mean PWTL was obtained from the latter
three stimuli.
RT-qPCR
Following the sacrifice of the mice, the L3-L5
lumbar spinal cord segments were immediately harvested, frozen in
liquid nitrogen and stored at -80°C. The spinal tissues were
homogenized prior to storage by grinding in liquid nitrogen with a
small mortar. Total RNA was isolated and purified using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's instructions. RT was performed using a Moloney
murine leukemia virus reverse transcriptase kit (Promega Corp.,
Madison, WI, USA). The generated cDNA was then used as a template
for PCR amplification with Taq DNA polymerase and ROX reference dye
provided by the SYBR Premix Ex Taq kit (DRR041A; Takara, Dalian,
China) using StepOnePlus RT-PCR system (Applied Biosystems; Thermo
Fisher Scientific, Inc.). β-actin, nNOS and iNOS were amplified
using specific oligonucleotide primers: forward,
5′-AGGAGCAAGGAGGCCATATT-3′ and reverse, 5′-AACACACCAGCATCCTCCTC-3′
for nNOS; forward, 5′-TGATGTGCTGCCTCTGGTCT-3′ and reverse,
5′-ACTTCCTCCAGGATGTTGTA-3′ for iNOS; and forward,
5′-GAGACCTTCAACACCCCAGC-3′ and reverse, 5′-CACAGAGTACTTGCGCTCAG-3′
for β-actin (GenScript, Piscataway, NJ, USA). β-actin was used as
internal standard. PCR amplification was performed using the
following thermocycling conditions: 94°C for 5 min, followed by 35
cycles of 94°C for 30 sec, 66°C (or 62°C for iNOS and β-actin) for
30 sec and 72°C for 30 sec, and a final elongation at 72°C for 10
min. The relative expression was calculated using the ΔΔCq method
(24) and optimized with a
standard curve to confirm specificity. Each sample (5 µl)
was electrophoresed on 2% agarose gel (Biowest, Nuaillé, France)
with ethidium bromide (Biomatik Corporation, Cambridge, ON, Canada)
and the intensity of each band was analyzed using a gel imaging
analytical system (UVP GDS 8000; UVP, Upland, CA, USA). The sample
without reverse transcriptase was used as a negative control and
showed no detectable band.
Immunohistochemical staining
To localize and assess the expression of nNOS and
iNOS in the spinal cord during bone cancer development, mice in the
tumor and sham groups were transcardially perfused with saline,
followed by freshly prepared 4% paraformaldehyde at post-operative
days 7, 10 and 14 (n=15 per group). The L3-L5 lumbar spinal cord
segments were removed, fixed in 4% PFA overnight at 4°C and then
embedded in paraffin for sectioning (5 µm; five mice per
group and ten sections from each mouse). The sections were dewaxed
in xylene (Nanjing Chemical Reagent Co., Ltd., Nanjing, China) for
10 mins and hydrated with a graded ethanol series (100, 95, 90, 85,
80 and 70%; Nanjing Chemical Reagent Co., Ltd.) for 3 mins each, at
room temperature. The sections were washed in double distilled
water (Nanjing Chemical Reagent Co., Ltd.) for 5 mins and boiled in
0.1 mol/l sodium citrate buffer (pH 6.0; Wuhan Boster Biological
Technology, Ltd., Wuhan, China) for 20 min prior to exposure to 3%
H2O2 (Shanghai Shenggong Biology Engineering
Technology Service, Ltd., Shanghai, China) for 10 min to bleach
endogenous peroxidases. Immunostaining was performed with rabbit
primary polyclonal antibodies against nNOS (1:10,000;
Sigma-Aldrich; SAB4502010) or rabbit primary polyclonal anti-iNOS
(1:800; Sigma-Aldrich; SAB4502011) for 30 min at 37°C, followed by
washing with PBS 3 times for 10 mins. They were then incubated with
a horseradish peroxidase-labeled goat anti-rabbit IgG (heavy and
light chain; Beyotime Institute of Biotechnology; A0208) secondary
antibody at 4°C overnight. The sections were then washed with PBS 3
times for 10 mins and visualized with diaminobenzidine (Beyotime
Institute of Biotechnology, Haimen, China) and observed under a
light microscope (Olympus DP11; Olympus, Tokyo, Japan). Optical
density of the images was analyzed using Image-Pro Plus 6.0
analysis software (Media Cybernetics, Inc., Rockville, MD, USA).
The mean optical density was obtained by averaging the values from
five sections.
Statistical analysis
Values are expressed as the mean ± standard
deviation. Animals were assigned to various treatment groups in a
randomized way. Repeated-measures analysis of variance (ANOVA) was
performed to determine overall differences at each time-point for
PWMT and PWTL. One-way ANOVA followed by a least-significant
differences post-hoc test was used to determine differences
in the mRNA levels of nNOS and iNOS among all experimental groups.
All analyses were performed using SPSS 13.0 software (SPSS, Inc.,
Chicago, IL, USA) and P<0.05 was considered to indicate a
statistically significant difference.
Results
Pain-associated behavior in tumor-bearing
mice
The PWMT and PWTL of mice in the tumor group prior
to the operation were not significantly different when compared
with those of the mice in the sham group. The right hind limb of
the mice in the tumor group displayed a significant decrease in the
PWMT at post-operative day 3 (P<0.05), and this was also
observed in the mice in the sham group. At post-operative day 5,
the PWMT of the two groups reclined to the basal level (Fig. 1A). Subsequently, the PWMT of mice
in the tumor group showed a further decline again at day 7, which
continuously decreased until the end of the experiment on
post-operative day 14 (0.48±0.25 g).
The mice in the tumor group showed a marked decrease
in PWTL at post-operative day 3, which recovered to basal levels at
days 5 and 7. Until day 7, similar trends were observed in the sham
group mice. However, the PWMT of mice in the tumor group decreased
again at post-operative day 10 and further declined to 10.5±3.2 sec
at day 14, while that in the sham group remained constant (Fig. 1B). Of note, significant decreases
in the PWTL at post-operative days 7–10 and in the PWMT at days 10
and 14 were observed in the tumor group when compared with those in
the sham group or the time-point prior to surgery, indicating the
development of marked bone cancer-associated pain.
nNOS and iNOS mRNA levels are increased
in the spinal cords of tumor-bearing mice
The mRNA levels of nNOS were significantly increased
in the spinal cord of mice in the tumor group at post-operative day
10 when compared with those in the same mice at day 7 (P<0.05;
Fig. 2A) as well as compared with
those in the sham group at day 7, while no significant differences
were present within the sham group at these time-points.
Furthermore, the mRNA levels of iNOS were significantly increased
in the spinal cord of mice in the tumor group at post-operative
days 10 and 14 when compared with those in the sham group at the
same time-points (P<0.05), or compared with those in the tumor
group at day 7 (Fig. 2B). However,
no significant differences in the mRNA levels of iNOS were observed
within the sham group at days 7–10.
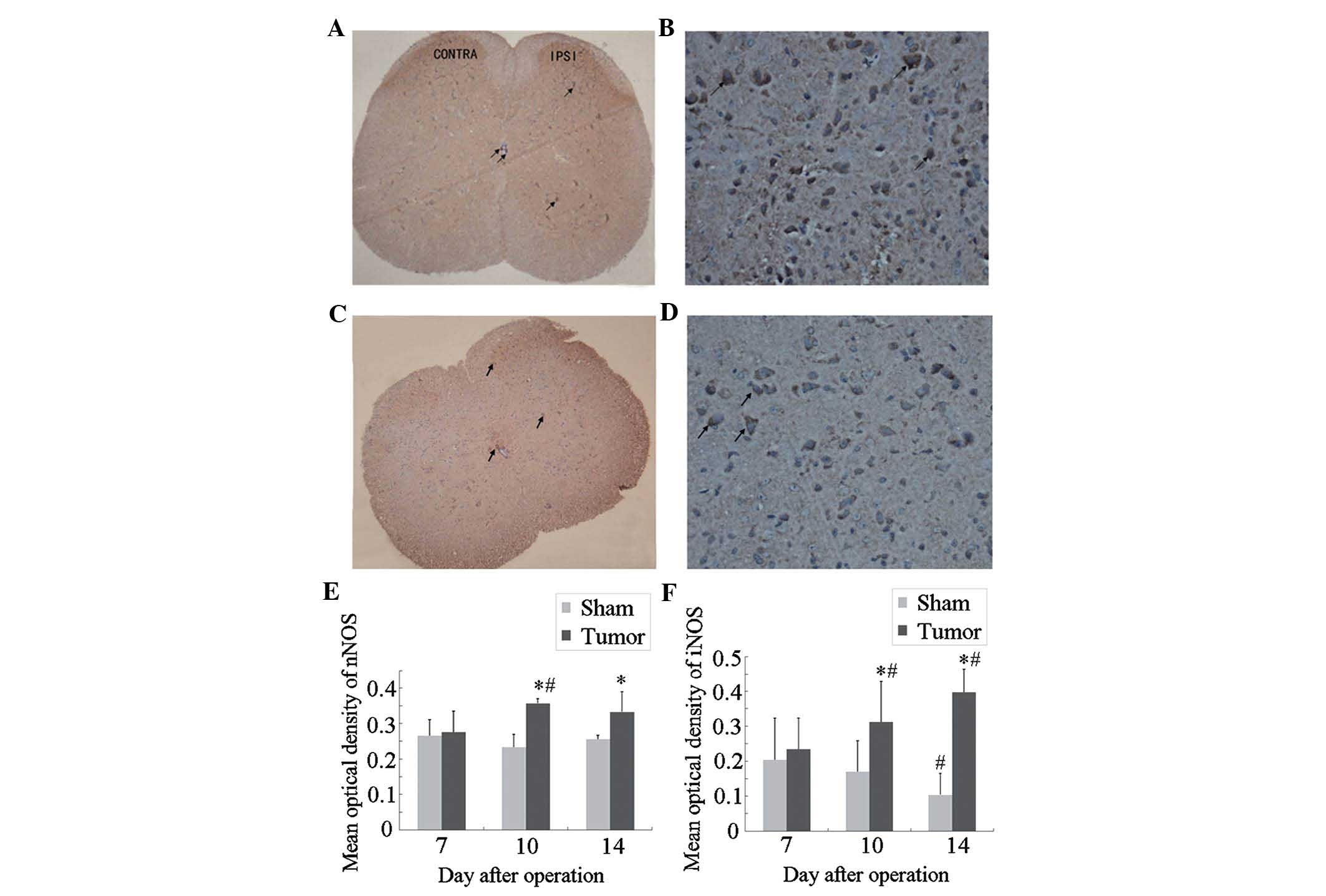
Immunocytochemical localization and
expression of nNOS and iNOS in the spinal cord of tumor-bearing
mice
Next, an immunohistochemical study was performed to
assess the localization and expression levels of nNOS and iNOS in
the spinal cord during bone cancer development. The results showed
that neurons positive for nNOS (Fig.
3A and B) and iNOS (Fig. 3C and
D) were mainly located in the superficial dorsal horn (laminae
I–II), which is involved in the elaboration of nociceptive stimuli,
and around the central canal (lamina X) of the L3–L5 spinal cord
(17). A small number of
iNOS-positive neurons were observed in the ventral horn and central
canal of the spinal cord.
At post-operative day 7, no significant differences
in nNOS or iNOS expression were present between the sham and tumor
groups (Fig. 3E and F). Compared
with those in the sham group, the expression of nNOS and iNOS in
the tumor group was significantly increased at post-operative day
10 and further increased at day 14. In the mice of the sham group,
the expression of iNOS at day 14 was significantly decreased as
compared with that at day 7, while no significant differences in
nNOS expression were observed within the sham group at different
time-points.
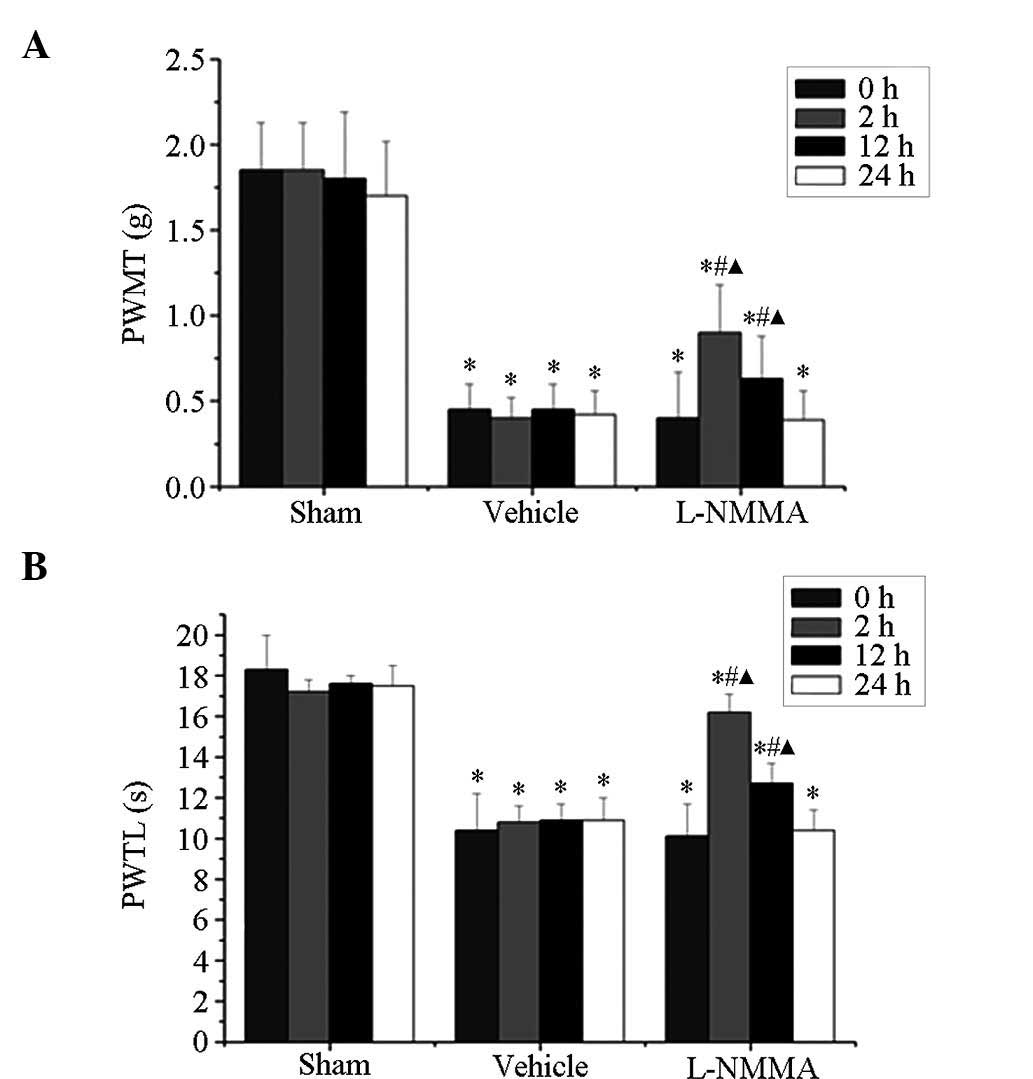
L-NMMA alleviates bone cancer pain in
mice
At post-operative day 14, mice in the tumor group
showed a marked decline in PWMT and PWTL. Of note, this decrease
was attenuated by intrathecal administration of 50 µg L-NMMA
(Fig. 4). The PWMT was
significantly increased at 2 h (0.90±0.28 vs. 0.40±0.27; P<0.05)
and 12 h (0.63±0.25 vs. 0.40±0.27; P<0.05) following
administration of L-NMMA (Fig.
4A). Similarly, the PWTL was also markedly increased at 2 h
(16.2±0.9 vs. 10.1±2.3; P<0.05) and 12 h (12.7±1.0 vs. 10.1±2.3;
P<0.05) following administration of L-NMMA (Fig. 4B).
Discussion
The present study using a murine model of bone
cancer pain following a method of a previous study by our group
(16) to demonstrate that bone
cancer-induced mechanical allodynia and thermal hyperalgesia are
accompanied with an upregulation of nNOS and iNOS in the lumbar
spinal cord. In addition, the present study found that NOS
inhibitor L-NMMA alleviated cancer-induced mechanical and thermal
hyperalgesia, suggesting that spinal nNOS and iNOS may participate
in the development of bone cancer pain.
While the mRNA levels of nNOS and iNOS in the lumbar
spinal cord of mice in the tumor group were increased at day 10
after tumor inoculation, the up-regulation of iNOS was sustained
until day 14 following surgery, at which the pain-associated
behavior of the mice in the tumor group were prominent and stable.
The cells with immunoreactivity for nNOS and iNOS were mainly
located in neurons present at the dorsal horn of the spinal cord
(laminae I–II), which is an area involved in the elaboration of
nociceptive stimuli, and in a few cells in the ventral parts and
around the central canal (17).
It has been reported that the mRNA levels of nNOS in
the lumbar spinal cord were increased at day one following surgical
inflammation (25). Another study
by Tang et al (5) observed
a low but constitutive expression of iNOS at the mRNA and protein
level in the rat spinal cord under non-stimulated conditions. They
also reported that constitutively expressed spinal iNOS mediates
tissue injury and inflammation-induced hyperalgesia, and that
intrathecal administration of a highly selective iNOS inhibitor
produced a dose-dependent inhibition of hyperalgesia. The iNOS gene
is induced by bacterial lipopolysaccharide or classical
pro-inflammatory cytokines, including tumor necrosis factor α. iNOS
produces a larger amount of NO over a longer time period compared
with the other two constitutive enzymes, nNOS and eNOS (26). Substantial evidence has been
provided for the link between the upregulation of spinal iNOS and
inflammatory and neuropathic pain, suggesting a potential
therapeutic value of iNOS inhibitors for such conditions (15,27).
In addition, Pedersen et al (28) reported that an increased expression
of iNOS in the dorsal horn was associated with long-term
potentiation that contributed to central hyperalgesia. Consistent
with a previous study (29), the
results of the present study suggested that nNOS contributed to the
initial NO release to facilitate nociceptive processing, while iNOS
possibly participated in the persistent nocicep-tive responses.
This observed difference in the time-course of the expression of
nNOS and iNOS suggests a distinct contribution of NOSs to the
mechanisms of bone cancer pain development.
L-NMMA, which is a non-selective inhibitor of NOS
that can act on all three isoforms of NOS, has been proved to exert
anti-nociceptive effects in several models of peripheral
inflammation and nerve injury. Intradermal administration (30) or intrathecal injection (31) of L-NMMA has been shown to reduce,
or in certain cases, to completely reverse inflammatory
hyperalgesia. In addition, intrathecal injection of L-NMMA is able
to reduce the mechanical allodynia evoked by nerve injury (32). Furthermore, a randomized crossover
trial demonstrated the analgesic effects of L-NMMA in patients with
chronic tension-type headache (33). Consistent with these studies, the
present study found that the mechanical and thermal hyperalgesia at
post-operative day 14 induced by bone cancer was alleviated at 2
and 12 h after intrathecal injection of L-NMMA, while this effect
disappeared at 24 h after L-NMMA administration.
In conclusion, the present study demonstrated that
spinal NOS may contribute to nociceptive signal processing during
central sensitization in the development of bone cancer pain. As a
NOS inhibitor, L-NMMA alleviated the cancer-induced mechanical
allodynia and thermal hyperalgesia, suggesting its prospective
application in bone cancer-associated pain. Further study is
required to determine the underlying mechanism of NOS-associated
bone cancer pain.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (nos. 81070892, 81171048,
81171047, 81371207, 81300950 and 81300951), the Natural Science
Foundation of Jiangsu Province (no. BK2010105) and a grant from the
Department of Health of Jiangsu Province of China (nos. XK201140
and RC2011006).
References
|
1
|
Colvin L and Fallon M: Challenges in
cancer pain management-bone pain. Eur J Cancer. 44:1083–1090. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Miyamoto T, Dubin AE, Petrus MJ and
Patapoutian A: TRPV1 and TRPA1 mediate peripheral nitric
oxide-induced nociception in mice. PLoS One. 4:e75962009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Naik AK, Tandan SK, Kumar D and
Dudhgaonkar SP: Nitric oxide and its modulators in chronic
constriction injury-induced neuropathic pain in rats. Eur J
Pharmacol. 530:59–69. 2006. View Article : Google Scholar
|
|
4
|
Guan Y, Yaster M, Raja SN and Tao YX:
Genetic knockout and pharmacologic inhibition of neuronal nitric
oxide synthase attenuate nerve injury-induced mechanical
hypersensitivity in mice. Mol Pain. 3:292007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tang Q, Svensson CI, Fitzsimmons B, Webb
M, Yaksh TL and Hua XY: Inhibition of spinal constitutive NOS-2 by
1400W attenuates tissue injury and inflammation-induced
hyperalgesia and spinal p38 activation. Eur J Neurosci.
25:2964–2972. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Choi JI, Kim WM, Lee HG, Kim YO and Yoon
MH: Role of neuronal nitric oxide synthase in the antiallodynic
effects of intrathecal EGCG in a neuropathic pain rat model.
Neurosci Lett. 510:53–57. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lam HH, Hanley DF, Trapp BD, Saito S, Raja
S, Dawson TM and Yamaguchi H: Induction of spinal cord neuronal
nitric oxide synthase (NOS) after formalin injection in the rat
hind paw. Neurosci Lett. 210:201–204. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chacur M, Matos RJ, Alves AS, Rodrigues
AC, Gutierrez V, Cury Y and Britto LR: Participation of neuronal
nitric oxide synthase in experimental neuropathic pain induced by
sciatic nerve transection. Braz J Med Biol Res. 43:367–376. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee JS, Zhang Y and Ro JY: Involvement of
neuronal, inducible and endothelial nitric oxide synthases in
capsaicin-induced muscle hypersensitivity. Eur J Pain. 13:924–928.
2009. View Article : Google Scholar
|
|
10
|
Handy RL and Moore PK: Effects of
selective inhibitors of neuronal nitric oxide synthase on
carrageenan-induced mechanical and thermal hyperalgesia.
Neuropharmacology. 37:37–43. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tanabe M, Nagatani Y, Saitoh K, Takasu K
and Ono H: Pharmacological assessments of nitric oxide synthase
isoforms and downstream diversity of NO signaling in the
maintenance of thermal and mechanical hypersensitivity after
peripheral nerve injury in mice. Neuropharmacology. 56:702–708.
2009. View Article : Google Scholar
|
|
12
|
Chu YC, Guan Y, Skinner J, Raja SN, Johns
RA and Tao YX: Effect of genetic knockout or pharmacologic
inhibition of neuronal nitric oxide synthase on complete Freund's
adjuvant-induced persistent pain. Pain. 119:113–123. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Toda N, Kishioka S, Hatano Y and Toda H:
Modulation of opioid actions by nitric oxide signaling.
Anesthesiology. 110:166–181. 2009. View Article : Google Scholar
|
|
14
|
De Alba J, Clayton NM, Collins SD, Colthup
P, Chessell I and Knowles RG: GW274150, a novel and highly
selective inhibitor of the inducible isoform of nitric oxide
synthase (iNOS), shows analgesic effects in rat models of
inflammatory and neuropathic pain. Pain. 120:170–181. 2006.
View Article : Google Scholar
|
|
15
|
Sasaki A, Mabuchi T, Serizawa K, Takasaki
I, Andoh T, Shiraki K, Ito S and Kuraishi Y: Different roles of
nitric oxide synthase-1 and -2 between herpetic and postherpetic
allodynia in mice. Neuroscience. 150:459–466. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gu X, Zhang J, Ma Z, Wang J, Zhou X, Jin
Y, Xia X, Gao Q and Mei F: The role of N-methyl-D-aspartate
receptor subunit NR2B in spinal cord in cancer pain. Eur J Pain.
14:496–502. 2010. View Article : Google Scholar
|
|
17
|
Meller ST and Gebhart GF: Nitric oxide
(NO) and nociceptive processing in the spinal cord. Pain.
52:127–136. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ma ZL, Zhang W, Gu XP, Yang WS and Zeng
YM: Effects of intrathecal injection of prednisolone acetate on
expression of NR2B subunit and nNOS in spinal cord of rats after
chronic compression of dorsal root ganglia. Ann Clin Lab Sci.
37:349–355. 2007.PubMed/NCBI
|
|
19
|
Zimmermann M: Ethical guidelines for
investigations of experimental pain in conscious animals. Pain.
16:109–110. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schwei MJ, Honore P, Rogers SD,
Salak-Johnson JL, Finke MP, Ramnaraine ML, Clohisy DR and Mantyh
PW: Neurochemical and cellular reorganization of the spinal cord in
a murine model of bone cancer pain. J Neurosci. 19:10886–10897.
1999.PubMed/NCBI
|
|
21
|
Hylden JL and Wilcox GL: Intrathecal
morphine in mice: A new technique. Eur J Pharmacol. 67:313–316.
1980. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chaplan SR, Bach FW, Pogrel JW, Chung JM
and Yaksh TL: Quantitative assessment of tactile allodynia in the
rat paw. J Neurosci Methods. 53:55–63. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hargreaves K, Dubner R, Brown F, Flores C
and Joris J: A new and sensitive method for measuring thermal
nociception in cutaneous hyperalgesia. Pain. 32:77–88. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
25
|
Dolan S, Kelly JG, Huan M and Nolan AM:
Transient up-regulation of spinal cyclooxygenase-2 and neuronal
nitric oxide synthase following surgical inflammation.
Anesthesiology. 98:170–180. 2003. View Article : Google Scholar
|
|
26
|
Conti A, Miscusi M, Cardali S, Germanò A,
Suzuki H, Cuzzocrea S and Tomasello F: Nitric oxide in the injured
spinal cord: Synthases cross-talk, oxidative stress and
inflammation. Brain Res Rev. 54:205–218. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Martucci C, Trovato AE, Costa B, Borsani
E, Franchi S, Magnaghi V, Panerai AE, Rodella LF, Valsecchi AE,
Sacerdote P and Colleoni M: The purinergic antagonist PPADS reduces
pain related behaviours and interleukin-1 beta, interleukin-6, iNOS
and nNOS overproduction in central and peripheral nervous system
after peripheral neuropathy in mice. Pain. 137:81–95. 2008.
View Article : Google Scholar
|
|
28
|
Pedersen LM, Jacobsen LM, Mollerup S and
Gjerstad J: Spinal cord long-term potentiation (LTP) is associated
with increased dorsal horn gene expression of IL-1beta, GDNF and
iNOS. Eur J Pain. 14:255–260. 2010. View Article : Google Scholar
|
|
29
|
Guhring H, Tegeder I, Lötsch J, Pahl A,
Werner U, Reeh PW, Rehse K, Brune K and Geisslinger G: Role of
nitric oxide in zymosan induced paw inflammation and thermal
hyperalgesia. Inflamm Res. 50:83–88. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Aley KO, McCarter G and Levine JD: Nitric
oxide signaling in pain and nociceptor sensitization in the rat. J
Neurosci. 18:7008–7014. 1998.PubMed/NCBI
|
|
31
|
Malmberg AB and Yaksh TL: Spinal nitric
oxide synthesis inhibition blocks NMDA-induced thermal hyperalgesia
and produces antinociception in the formalin test in rats. Pain.
54:291–300. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yonehara N, Kudo C and Kamisaki Y:
Involvement of NMDA-nitric oxide pathways in the development of
tactile hypersensitivity evoked by the loose-ligation of inferior
alveolar nerves in rats. Brain Res. 963:232–243. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ashina M, Lassen LH, Bendtsen L, Jensen R
and Olesen J: Effect of inhibition of nitric oxide synthase on
chronic tension-type headache: A randomised crossover trial.
Lancet. 353:287–289. 1999. View Article : Google Scholar : PubMed/NCBI
|