Introduction
Hepatocellular carcinoma (HCC) is the third leading
cause of cancer-related mortality worldwide and the principal cause
of fatality among patients with cirrhosis (1). Despite recent advances in cancer
treatment with respect to surgery, chemotherapy and biologics, the
majority of cases of HCC remain incurable once they have become
metastatic and are associated with a poor prognosis.
Hepatocarcinogenesis has often been described as a multistep
process involving a number of genetic alterations, eventually
leading to the malignant transformation of hepatocytes (2). The advances in suitable therapies for
the purpose of increasing survival rate are limited as the
pathophysiological mechanisms causing HCC have not been fully
elucidated. Revealing the pathological mechanism underlying HCC
development and progression is indispensable for developing
effective therapeutic agents.
The human transcriptome comprises not only large
numbers of protein-coding messenger RNAs (mRNAs), but also a large
set of non-protein coding transcripts that have structural,
regulatory or unknown functions (3). Novel evidence shows that lncRNAs
(long non-coding RNAs) exhibit important regulatory roles in
tumorigenesis and cancer progression (4). For example, HOTAIR levels have been
shown to be higher in tumor tissues than the adjacent normal
tissue, and overexpression of HOTAIR correlates with the presence
of metastasis. In addition, patients with high HOTAIR expression
levels have been shown to have a poor prognosis (5). Forced expression of HOTAIR in cancer
cells leads to altered histone H3 lysine27 methylation and gene
expression, and increased metastasis (6). Maternally expressed gene 3 (MEG3)
expression is markedly decreased in several primary human tumors
and tumor cell lines by promoter hypermethylation (7). MEG3 is associated with meningioma
progression by inducing the accumulation of p53 protein and
selectively regulating p53 target gene expression. In addition,
overexpression of MEG3 inhibits cancer cell proliferation and
colony formation (8).
The long noncoding RNA, small nucleolar RNA host
gene-growth arrest-specific 5 (GAS5), was originally identified via
subtractive cDNA cloning of genes which are preferentially
expressed in growth-arrested cells (9). GAS5 comprises 12 exons and encodes
ten box C/D within its introns (10). The open reading frame encoded
within GAS5 exons is short and thought not to encode a functional
protein. This, together with the presence of a conserved
5′-terminal oligopyrimidine tract (5′-TOP), explains why GAS5-a
ncRNA accumulates upon growth arrest in a range of species
(11). Several studies have
implicated GAS5 ncRNA in other types of cancer (12,13).
GAS5 and/or its snoRNAs have been shown to be aberrantly expressed
in breast cancer, head and neck squamous cell carcinoma (14–16)
and glioblastoma multiforme (17),
where tumor expression is reduced, and non-small-cell lung cancer,
where overexpression of the snoRNAs U44, U76 and U78 occurs
(18). However, the mechanisms
underlying GAS5 regulation of cancer cell proliferation in HCC
remain unclear.
Based on the above findings, the present study
analyzed the expression of GAS5 in HCC tissues and adjacent normal
tissues, and its effect on cell proliferation in HCC cells. It was
demonstrated that GAS5 levels are significantly decreased in HCC
tissues, and decreased expression of GAS5 indicates a poor
prognosis in patients with HCC. In addition, GAS5 was shown to
suppress hepatoma cell proliferation in a vimentin-dependent
manner.
Materials and methods
Tissue samples, cell lines and clinical
data collection
A total of 50 patients were analyzed in this study.
The patients underwent HCC resection at the Department of General
Surgery at the Zhongnan Hospital of Wuhan University (Wuhan, China)
between February 2009 and March 2010. The study was approved by the
ethics committee of Zhongnan Hospital of Wuhan University (Wuhan,
China). Written informed consent was obtained from all patients.
The diagnosis of HCC was histopathologically confirmed. No patients
received preoperative treatment. Tumor and corresponding adjacent
normal tissues were selected for each case. Normal human liver
tissues were obtained >5 cm from the edge of the cancerous
region. Resected tissue samples were immediately frozen in liquid
nitrogen and stored at −80°C until RNA extraction, or fixed in 4%
paraformaldehyde for paraffin sectioning. Clinicopathological data
was collected for all subjects including details of age, gender and
HCC features, such as tumor size, differentiation, hepatitis B
virus (HBV) infection, liver cirrhosis, serum α-fetoprotein (AFP)
levels, tumor number and the presence of portal vein tumor thrombus
(PVTT). Patient follow up was performed by telephone interview and
questionnaire every 3 months until April 30, 2015. Overall survival
(OS) was calculated from the date of the initial surgery to the
date of death. Patients who succumbed to diseases not directly
related to HCC were excluded from the study.
The following cell lines were used in this study:
L02 normal human liver cells, and Huh7, Hep3B, HepG2, QGY-7701,
MHCC97L and HCCLM9he human hepatoma cell lines. All cell lines were
obtained from the Cell Bank of the Chinese Academy of Sciences
(Shanghai, China), where they were characterized by mycoplasma
detection, DNA fingerprinting, isozyme detection and determination
of cell viability. All cells were cultured in the recommended media
supplemented with 10% (v/v) fetal bovine serum (FBS; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), and 100 U/ml penicillin
(Beyotime Institute of Biotechnology, Shanghai, China) and
streptomycin (Beyotime Institute of Biotechnology) at 37°C in an
incubator with a 5% CO2 atmosphere (Thermo Fisher
Scientific, Inc.).
Ethics statement
Patient samples were collected and stored in the
Zhongnan Hospital of Wuhan University tumor bank. Written informed
consent was obtained from all of the patients. All procedures were
approved by the Research Ethics Committee of Wuhan University.
Patient data and samples were treated according to the ethical and
legal standards adopted by the Declaration of Helsinki 2013.
RNA preparation, semiquantitative-reverse
transcription and reverse transcription quantitative polymerase
chain reaction (RT-qPCR)
The expression level of GAS5 was first examined by
semiquantitative PCR in tissue from 6 patients with HCC. The
expression of GAS5 was then analyzed by RT-qPCR in tissue from all
50 patients with HCC patients. Total RNA was extracted from tissues
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific Inc.)
according to the manufacturer's protocol. Quantification of total
RNA was performed with a Nanodrop™ spectrophotometer (Thermo Fisher
Scientific Inc.) at 260 and 280 nm. Only samples with an A260/A280
ratio between 1.8 and 2.0 were utilized for further analysis. Total
RNA (5 µg) was used to synthesize first-strand cDNA using
random primers and SuperScript II reverse transcriptase
(Invitrogen; Thermo Fisher Scientific Inc.) according to the
manufacturer's protocol. The 20 µl RT reactions were
performed using a PrimeScript RT reagent kit (Takara Bio Inc.,
Wuhan, China) and incubated for 30 min at 37°C and 5 sec at 85°C,
and then maintained at 4°C for 1% agarose gel electrophoresis. For
qPCR, 2 µl diluted RT products were combined with 10
µl of 2X SYBR Master mix (Toyobo Co., Ltd. Osaka, Japan), 1
µl forward and reverse primers (10 µM; Table I; Beijing View Solid Biotechnology
(Beijing, China), and 6 µl nuclease-free water in a final
volume of 20 µl according to the manufacturer's
instructions. Amplification was performed with the iQ5 quantitative
PCR system (Bio-Rad Laboratories Inc., Hercules, CA, USA), under
the following conditions: 95°C for 30 sec, followed by 40 cycles at
95°C for 15 sec, and 60°C for 30 sec, and 72°C for 30 sec. qPCR was
conducted in triplicate, including non-template controls.
Amplification of the appropriate product was confirmed by melting
curve analysis following amplification. Relative expression of GAS5
and vimentin were calculated using the comparative cycle
quantification (Cq) (2−ΔΔCq) method (19) with β-actin as the endogenous
control to normalize the data. PCR products were run on agarose
gels to determine size, and dissociation curves were subsequently
utilized to examine the specificity of the RT-qPCR assay.
 | Table IDNA and RNA sequences of primers. |
Table I
DNA and RNA sequences of primers.
| Primer name | Sequence
(5′-3′) | bp |
|---|
| GAS5-F |
CAACTTGCCTGGACCAGCTT | 126 |
| GAS5-R |
TCAAGCCGACTCTCCATACC | |
| β-actin-F |
AGCGAGCATCCCCCAAAGTT | 285 |
| β-actin-R |
GGGCACGAAGGCTCATCATT | |
| Vimentin-F |
GCAGGAGGCAGAAGAATGGT | 140 |
| Vimentin-R |
CCACTTCACAGGTGAGGGAC | |
| E-cadherin-F |
CTTGCGGAAGTCAGTTCAGA | 216 |
| E-cadherin-R |
CACCGTGAACGTGTAGCTCT | |
|
Vimentin-siRNA-F |
UCACGAUGACCUUGAAUAA | |
|
Vimentin-siRNA-R |
GAGGGAAACUAAUCUGGAU | |
In situ hybridization (ISH)
ISH was used to detect GAS5 in HCC clinical
specimens. A digoxigenin-UTP-labeled antisense RNA probe (Beijing
View Solid Biotechnology) was derived from 234 to 478 nt of GAS5 by
in vitro transcription using the DIG RNA Labeling kit (Roche
Diagnostics, Indianapolis, IN, USA). The digoxigenin-UTP labeled
sense RNA probe derived from 234 to 478 nt of GAS5 was employed as
a negative control. ISH was performed using the ISH kit (Boster
Bio-Engineering Company, Wuhan, China). The ISH-staining regions
were reviewed and scored by two pathologists, the score standard
for the staining intensity was as follows: 0, negative; 1, weak; 2,
medium; and 3, strong. The proportion of GAS5-positive cells was
scored as: 0, 0%; 1, 1–25%; 2, 26–50%; 3, 51–75%; and 4, 76–100%.
The total scores were calculated by combining two scores and ranged
from 0 to 7. Total scores of ≥3 were defined as the high-expression
group (positive group). This scoring method was simple and
reproducible. Results were highly concordant between the two
independent pathologists.
Immunohistochemistry
Vimentin and E-cadherin expression in primary tumor
tissues and adjacent non-tumor tissues were examined using IHC.
Paraffin-embedded blocks containing tumor tissues or non-tumor
tissues and >70% primary tumor tissue were selected for IHC
staining. Paraffin sections were cut to 4 µm, mounted on
silanized slides and the preserved at 4°C. The paraffin sections
were dewaxed, rehydrated and blocked with 0.3%
H2O2. Tissue antigens were retrieved with a
microwave oven at 95°C for 25 min and cooled to room temperature in
10 mmol sodium citrate buffer (pH 6.0). Each slide was washed with
phosphate-buffered saline (PBS) and incubated overnight at 4°C with
anti-vimentin (1:100, cat. no. 5741S, Cell Signaling Technologies,
Inc.) or anti-E-cadherin (1:100, cat. no. 3195S, Cell Signaling
Technologies, Inc.). Primary antibodies were diluted with
background-reducing components (Dako, Glostrup, Denmark). Slides
were then incubated with goat-anti-mouse IgG-horseradish peroxidase
(HRP) (cat. no. BA1050; 1:5,000; Wuhan Boster Biological
Technology, Ltd., Wuhan, China) or goat-anti-rabbit IgG-HRP
(1:5,000; cat. no. BA1055; Wuhan Boster Biological Technology,
Ltd.). Slides were stained for 2 min with diaminobenzidine
tetrahydrochloride and then counterstained with hematoxylin. Tissue
treated with antibody dilution solution was used as a negative
control. All controls yielded satisfactory results. An FV1000
confocal laser-scanning microscope (Olympus, Tokyo, Japan) was used
to examine the tissues. The total immunostaining score was
calculated as the sum of the positive percentage and the staining
intensity of the stained cells, which ranged from 0 to 6. The
percent positivity was scored as follows: 0, 0–25%; 1, 26–50%; 2,
51–75%; and 3, ≥75%. The staining intensity was scored as: 0, no
staining; 1, weakly stained; 2, moderately stained; and 3, strongly
stained. The results of the immunohistochemical staining were
scored by two experienced pathologists, who were blinded to the
clinical data. A negative expression of protein was defined as a
total score ≤3, and positive expression was defined as a total
score ≥4.
Transfection assays
The GAS5 sequence (Gene-bank: NR_002578.2) was
synthesized according to the full length GAS5 sequence (based on
the GAS5 sequence) and then cloned into a pcDNA3.1 vector
(Invitrogen; Thermo Fisher Scientific Inc.). The GAS5-pcDNA3.1 or
empty vector was transfected into hepatoma cells. Transfection was
conducted using LipJetTM in vitro DNA and siRNA Transfection
kit (SignaGen Laboratories, Gaithersburg, MD, USA) according to the
manufacturer's protocol. All transfections were performed using
Lipofectamine 2000 reagent (Invitrogen; Thermo Fisher Scientific
Inc.) according to the manufacturer's protocol. The empty pcDNA3.1
vector was used as the control. All transfection experiments were
performed at least three times.
Fluorescent immunocytochemistry
Hepatoma cells were fixed with 4% paraformaldehyde
[China Sinopharm International (Shanghai), Co., Ltd.] for 20 min
and then permeabilized with 0.5% Triton X-100 [China Sinopharm
International (Shanghai), Co., Ltd.] for 10 min. The fixed cells
were blocked with 3% FBS for 30 min. For specific detection of
vimentin protein (Cell Signaling Technologies, Inc., Boston, MA,
USA), the cells were incubated with polyclonal rabbit anti-rat
vimentin antibody (1:100, CST, USA) overnight at 4°C. The samples
were incubated with Cy3-labeled goat anti-rabbit IgG (1:100,
eBioscience, Inc., San Diego, CA, USA) in PBS for 1 h at 37°C.
Then, the cells were washed twice with PBS and incubated with 4′,
6-diamidino-2-phenylindole (DAPI; BD Biosciences, La Jolla, CA,
USA) for 5 min. The images were analyzed using fluorescence
microscopy (Nikon Eclipse 80i, Tokyo, Japan). Identical
illumination and camera settings were used within each dataset.
Cell proliferation assay
Cell proliferation assays were performed using Cell
Counting kit-8 (Dojindo Molecular Technologies Inc., Kumamoto,
Japan), according to the manufacturer's protocol. Human hepatoma
cells were plated in 24-well plates in triplicate at a density of
2-5×104 cells per well and cultured in growth medium.
Cells were then treated with pcDNA3.1, empty vector or
vimentin-small interfering (si)RNA (Beijing View Solid
Biotechnology) and the numbers of cells per well were measured by
the (450 nm) at the indicated time points. The plasmid
GAS5-pcDNA3.1 (Beijing View Solid Biotechnology) was constructed by
introducing a KpnIXhoI fragment containing the GAS5
cDNA into the same sites in pcDNA3.1.
Bromodeoxyuridine (Brdu) assay
Hepatoma cells were fixed in 4% paraformaldehyde for
20 min, cells were then washed three times in PBS, dyed with BrdU
(BD Biosciences) for 30 min, washed again in PBS three times, and
treated with DAPI for 5 min. The images were analyzed using
fluorescence microscopy (Nikon Eclipse 80i). Identical illumination
and camera settings were used within each dataset.
Flow cytometric analysis
Hepatoma cells (2-5×105) treated with
GAS5-pcDNA3.1 or negative control (NC) were plated in 6-well
plates. After 48 h incubation the cells were harvested by
trypsinization. The cultures were double stained with incubated
with Annexin V and propidium iodide [China Sinopharm International
(Shanghai), Co., Ltd.] for 30 min in the dark. Cultures were
collected and analyzed for cell apoptosis using a flow cytometer
(FACScan; BD Biosciences Franklin Lakes, NJ, USA) equipped with
CellQuest 3.3 software. Cells were categorized as early apoptotic
cells, late apoptotic cells, dead cells, or viable cells. The ratio
of early apoptotic cells was compared with that in the controls
from each experiment.
Cell invasion assays
After transfection for 24 h, cells in serum-free
media were seeded into the upper chamber of a Transwell apparatus
(BD Biosciences). for invasion assays with Matrigel (Sigma-Aldrich,
St. Louis, MO, USA). The lower chambers were filled with media
containing 10% FBS. After 24 h of incubation at 37°C in a 5%
CO2 atmosphere, the cells which had invaded through the
membrane were fixed in 4% paraformaldehyde and stained with 0.1%
crystal violet (Sigma-Aldrich). The cells on the lower surface were
photographed (Olympus; IX73) and three random fields were counted
using a confocal laser-scanning microscope (Olympus FV1000). Three
independent experiments were performed.
Western blot analysis
Western blot analysis to assess vimentin and
E-cadherin protein expression was performed as previously described
(20). Glyceraldehyde 3-phosphate
dehydrogenase served as a control. Mouse monoclonal anti-GAPDH
(cat. no. KM9002; 1:5,000; Sungene, Tianjin, China), rabbit
monoclonal anti-vimentin (cat. no. 5741S; 1:1,000; Cell Signaling
Technology, Inc.) and rabbit monoclonal E-cadherin (cat. no. 3195S;
1:1,000; Cell Signaling Technology, Inc.) primary antibodies were
used. Goat anti-mouse IgG-HRP (cat. no. BA1050; 1:5,000; Boster
Bio-Engineering Company) and goat anti-rabbit IgG-HRP (cat. no.
BA1055; 1:5,000, Boster Bio-Engineering Company).
Statistical analysis
All statistical analyses were performed using SPSS
software (SPSS, Inc., Chicago, IL, USA). The differences between
groups were tested using Student's t-test. Survival curves were
calculated using Kaplan-Meier and log-rank tests. The effects of
variables on survival were determined by univariate and
multivariate Cox proportional hazards modeling. The correlation
between GAS5 and vimentin was analyzed by Spearman's rank
correlation. Two-sided P-values were calculated. P<0.05 was
considered to indicate a statistically significant difference.
Results
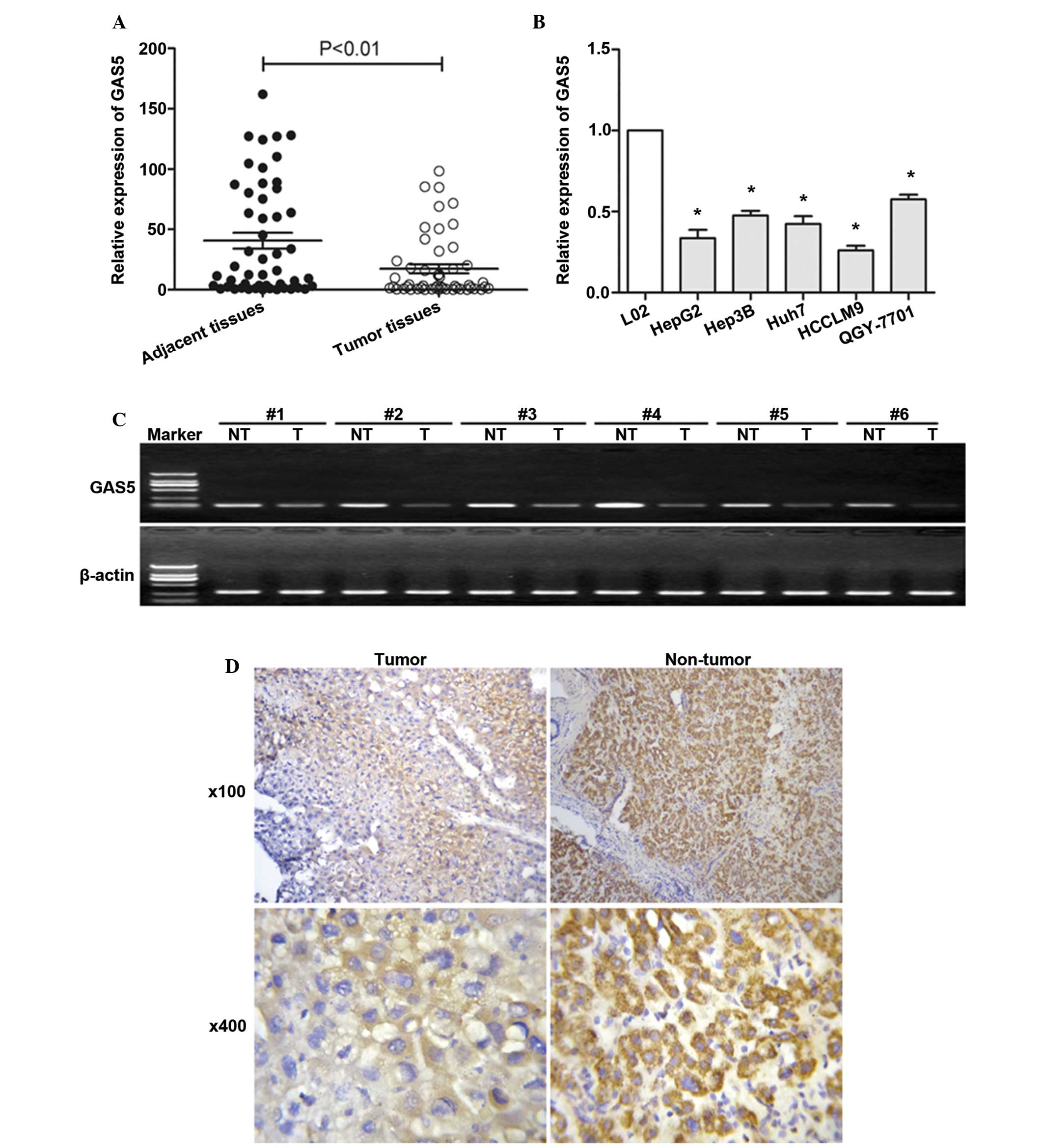
GAS5 is significantly downregulated in
HCC tissues and hepatoma cells
To investigate whether GAS5 regulates
hepatocarcinogenesis, the expression level of GAS5 was examined in
HCC and adjacent normal tissues (Fig.
1). The expression level of GAS5 was first examined by
semiquantitative RT-PCR in tissue from 6 patients with HCC. GAS5
was downregulated in HCC tissues compared with adjacent non-tumor
tissues (Fig. 1C). The expression
of GAS5 was then investigated by RT-qPCR in tissue from all 50
patients with HCC. Fig. 1A showed
significantly lower GAS5 expression in HCC tissues. Fig. 1A and C show that GAS5 is
significantly downregulated in HCC tissues compared with adjacent
normal tissues. The mean value of GAS5 expression in normal tissues
was 40.74 but the mean value of GAS5 expression in HCC tissues is
17.35. GAS5 expression levels were also assessed in hepatoma cells,
as shown in Fig. 1B, GAS5 was
observed to be downregulated in hepatoma cells compared with normal
liver cells. An ISH assay (Fig.
1D) further confirmed that GAS5 was significantly downregulated
in HCC tissues compared with adjacent normal tissues.
GAS5 expression and clinicopathological
factors in HCC
As shown in Table
II, in order to assess the correlation between GAS5 expression
and clinicopathological data, tumor tissues were divided into the
low-expression group (mean expression value 1.24, n=25) and the
high-expression group (mean expression value 33.45, n=25), based on
the mean expression level of all tumor tissues (mean expression
value, 2.98). Clinicopathological factors were analyzed in the high
and low GAS5 expression groups. The patients with low GAS5 (n=25)
exhibited a poorer histologic grade (P<0.010) and increased
presence of PVTT (P=0.001) than the high GAS5 expression group
(n=25). However, there was no significant correlation between GAS5
expression and other clinicopathologic features, such as age,
gender, HBV infection, liver cirrhosis, serum AFP and tumor number
(P>0.05).
| Table IICorrelation between GAS5 expression
and clinicopathologic parameters of HCC. |
Table II
Correlation between GAS5 expression
and clinicopathologic parameters of HCC.
| Parameter | Number of
cases | GAS5 expression
| P-value |
|---|
| Low (n=25) | High (n=25) |
|---|
| Age (years) | | | | 0.869 |
| ≥65 | 32 | 17 | 15 | |
| <65 | 18 | 8 | 10 | |
| Gender | | | | 0.747 |
| Male | 37 | 18 | 19 | |
| Female | 13 | 7 | 6 | |
| Tumor size
(cm) | | | | 0.774 |
| ≥5 | 29 | 15 | 14 | |
| <5 | 21 | 10 | 11 | |
| Tumor
differentiation | | | | <0.010 |
| Well or
moderate | 24 | 5 | 19 | |
| Poor | 26 | 20 | 6 | |
| HBV infection | | | | 0.239 |
| Yes | 32 | 14 | 18 | |
| No | 18 | 11 | 7 | |
| Liver
cirrhosis | | | | 0.569 |
| Yes | 22 | 10 | 12 | |
| No | 28 | 15 | 13 | |
| Serum AFP
(µg/l) | | | | 0.544 |
| ≥400 | 34 | 16 | 18 | |
| <400 | 16 | 9 | 7 | |
| Tumor number | | | | 0.157 |
| Singular | 40 | 22 | 18 | |
| Multifocal | 10 | 3 | 7 | |
| PVTT | | | | 0.001 |
| Yes | 15 | 13 | 2 | |
| No | 35 | 12 | 23 | |
Low expression of GAS5 indicates a poor
prognosis in HCC
To determine the factors responsible for patient
survival, univariate and multivariate analysis were performed. GAS5
expression levels were obtained from the RT-qPCR data of the cohort
of 50 patients mentioned above. Univariate analysis of overall
survival revealed that GAS5 expression (P=0.000), PVTT (P=0.008),
and differentiation (P=0.003) were prognostic indicators (Table III). Multivariate analysis showed
that GAS5 expression was an independent prognostic indicator for
overall survival (P<0.001; Fig.
2A). No significant associations were found with age, gender,
HBV infection, liver cirrhosis, serum AFP and tumor number
(Table III). Furthermore, high
expression of GAS5 was found to be associated with lower rates of
OS (P=0.017; log-rank test; Fig.
2B). However, no significant associations were found between OS
and age, gender, HBV infection, liver cirrhosis, serum AFP and
tumor number.
| Table IIIUnivariate and multivariate analysis
using the Cox proportional hazard regression model for overall
survival. |
Table III
Univariate and multivariate analysis
using the Cox proportional hazard regression model for overall
survival.
| Variable |
Univariate
(P-value) |
Multivariate
(P-value) |
|---|
| Age | 0.943 | |
| Gender | 0.574 | |
| Tumor size | 0.891 | |
| Histologic
grade | 0.003 | 0.023 |
| HBV infection | 0.121 | |
| Liver
cirrhosis | 0.385 | |
| Serum AFP | 0.270 | |
| Tumor number | 0.476 | |
| PVTT | 0.008 | 0.031 |
| GAS5
expression | 0.000 | <0.001 |
GAS5 regulates cell apoptosis and cell
proliferation in vitro
To investigate the role of GAS5 in the regulation of
cell proliferation, cell apoptosis and the ability of cell
invasion, the hepatoma cells were treated with GAS5-pcDNA3.1.
RT-qPCR was used to observe the expression of GAS5 (Fig. 3A). Overexpression of GAS5 inhibited
HCCLM9 proliferation (Fig. 3B and
C) and promoted HCCLM9 apoptosis (Fig. 3D and E). In addition,
overexpression of GAS5 suppressed the invasion ability of HCCLM9
(Fig. 3F and G). The above data
demonstrated that GAS5 is key in hepatocarcinogenesis.
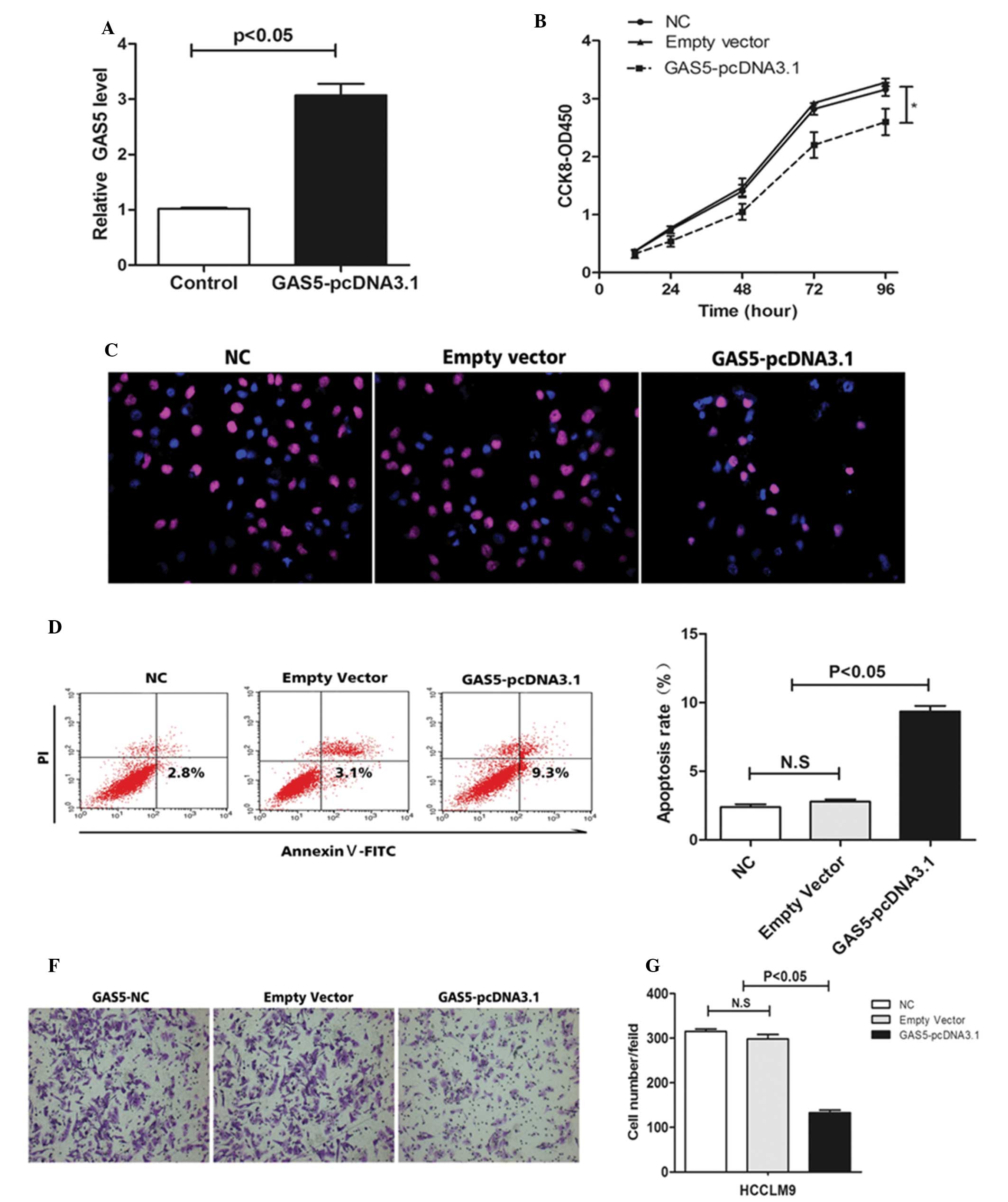 | Figure 3Effect of GAS5 on hepatoma cell
growth, invasion and apoptosis in vitro. (A) The relative
expression level of GAS5 in HCCLM9 cells, transfected with empty
vector or GAS5-pcDNA3.1, was tested by reverse
transcription-quantitative polymerase chain reaction. (B) At 48 h
after transfection, a cell counting kit-8 assay was performed to
determine the proliferation of HCCLM9 cells. (C) At 48 h after
transfection, a Brdu assay indicated that overexpression of GAS5
inhibited HCCLM9 proliferation (magnification, ×400). (D and E) At
48 h after transfection, cell apoptosis of HCCLM9 was analyzed by
flow cytometry. (F and G) At 48 h after transfection a cell
invasion assay was performed to investigate the ability of invasion
of HCCLM9. The results show data from at least three independent
experiments, expressed as the mean ± standard deviation.
*P<0.05. NC, negative control; GAS5, growth-arrest
specific 5; Brdu, bromodeoxyuridine; N.S. not significant. |
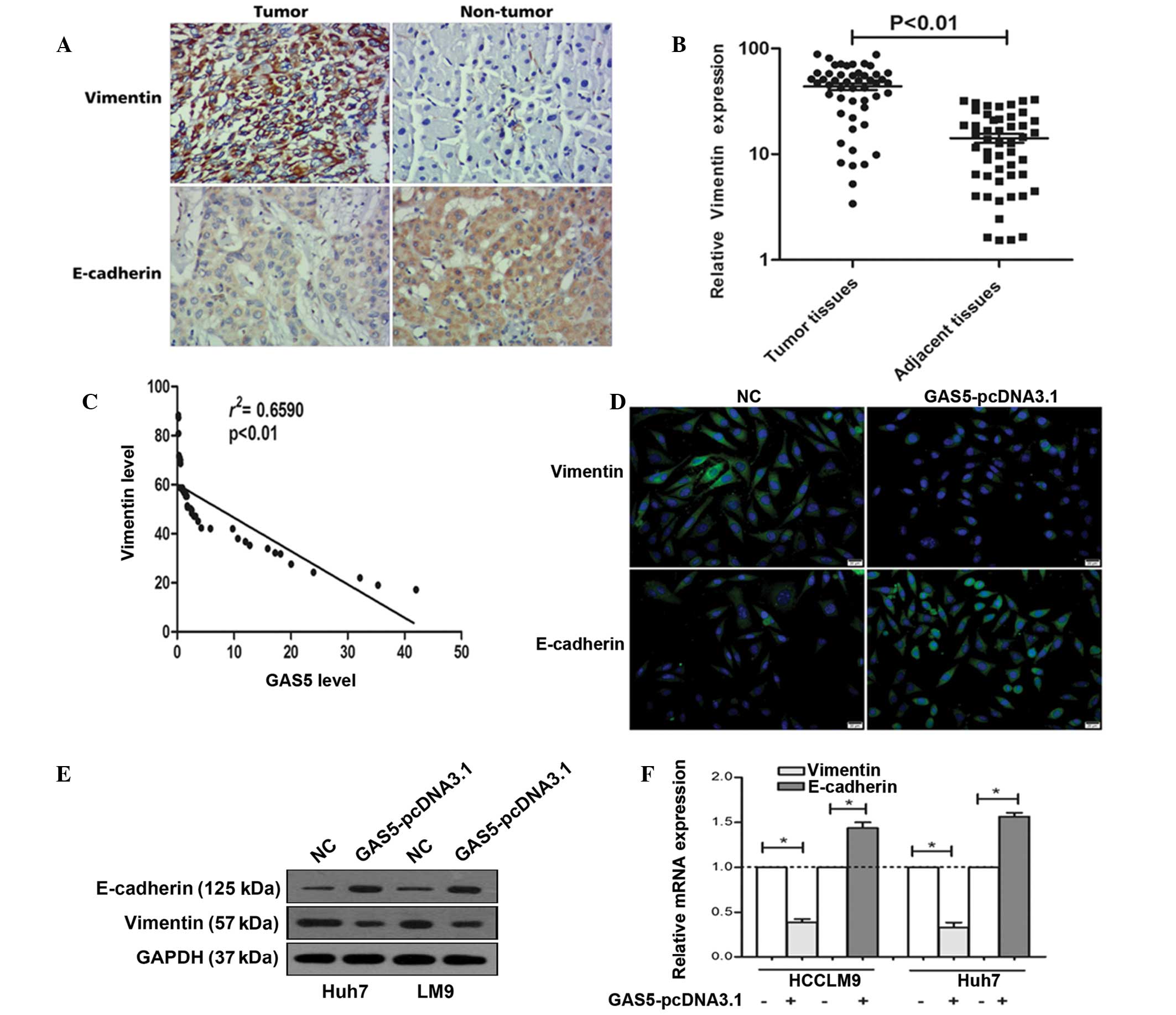
Expression of vimentin and E-cadherin in
HCC
Epithelial-mesenchymal transition (EMT) has been
shown to be of critical importance in the early events of tumor
cell metastatic dissemination as cells become more motile and
acquire invasive potential (21).
It was previously found that GAS5 suppressed the invasion ability
of tumor cells. Thus, the present study determined the expression
of vimentin and E-cadherin (marker protein of EMT) (22). As shown in Fig. 4A, immunohistochemistry indicated
that vimentin was significantly upregulated in tumor tissues, while
E-cadherin was downregulated in tumor tissues. In addition, RT-qPCR
confirmed the above phenomenon (Fig.
4B).
GAS5 negatively regulates vimentin
expression in vitro
To investigate the correlation between GAS5 and
vimentin, Spearman's rank correlation analysis was used. As shown
in Fig. 4C a significant negative
correlation is observed between the GAS5 levels and the vimentin
levels in HCC tissues (r2=0.6590, P<0.01). In
addition, fluorescent immunocytochemistry indicated that
overexpression of GAS5 decreased vimentin expression and increased
E-cadherin expression (Fig. 4D).
Western blot analysis demonstrated that the vimentin protein level
is downregulated and E-cadherin is upregulated in Huh7 and HCCLM9
after GAS5 overexpression (Fig.
4E). Figure 4F shows that
overexpression of GAS5 significantly decreased the vimentin mRNA
levels and increased E-cadherin mRNA in Huh7 and HCCLM9 cells.
These data indicated that GAS5 regulates hepatoma cell
proliferation and invasion by the regulation of vimentin.
GAS5 inhibits cell proliferation by
regulating vimentin
GAS5 overexpression increased hepatoma cell
proliferation and a significant negative correlation was observed
between GAS5 and vimentin. Thus, it was hypothesized that the role
of GAS5 in regulating hepatoma cell proliferation is mediated by
vimentin. Figure 5A shows that
vimentin knockdown partially strengthened the ability of
GAS5-pcDNA3.1-inhibited cell proliferation in HCCLM9 cells.
Vimentin inhibition also strengthened the ability of
GAS5-pcDNA3.1-inhibited cell proliferation in Huh7 cells (Fig. 5B). These data confirmed that GAS5
suppressed hepatoma cell progression, at least in part, by
regulating vimentin expression.
Discussion
In recent years, genome-wide surveys have revealed
that 98% of the total human genome can be transcribed, including
certain short or lncRNAs with numbered or no protein-coding
capacity (23–25). Up to now, studies have identified a
huge number of lncRNAs involved in the development and progress of
human diseases, including cancer (21,26).
LncRNAs can function as regulators of tumor suppressor or oncogene
expression and may be one of the 'cutt' that lead to oncogenesis
(27).
To date, there is increasing evidence that suggests
that certain lncRNAs can be identified as biomarkers for the
prognosis of tumor therapeutic targets in human cancer (28). GAS5 is an lncRNA involved in the
regulation of cell cycles. GAS5 is key in normal growth arrest in
T-cell and non-transformed lymphocytes. GAS5 inhibition suppresses
cell apoptosis and maintains a more rapid cell cycle, whereas GAS5
overexpression results in an increase in cell apoptosis and a
reduction in the rate of progression through the cell cycle,
revealing that GAS5 is required for normal growth arrest (14). Recently, studies have identified a
novel role of GAS5 in the regulation of tumorigenesis (29). GAS5 was downregulated in a number
of tumor types, such as breast cancer, head and renal cell
carcinoma, prostate cancer, and glioblastoma (30–32).
These observations indicate that GAS5 may function as a tumor
suppressor in human tumor progression. However, the molecular
mechanisms underlying GAS5 regulation of cancer cell proliferation
in HCC remain unclear.
EMT has been shown to be of critical importance in
the early events of tumor cell metastatic dissemination during
which cells become more motile and exhibit invasive potential
(33). EMT is not just a mechanism
that forms fibroblast-like cells; it is a process that results in
cancer cell migration, invasion, and metastasis (34,35).
Various biomarkers have been screened to indicate the EMT process,
including the loss of E-cadherin, zona occludens-1 and cytokeratin,
and the upregulation of matrix metalloproteinase, fibronectin,
α-smooth muscle actin, vimentin, snail and slug (35). Vimentin is a 57 kDa, type III
intermediate filament that is found in mesenchymal cells of various
types of tissues during their developmental stages. Its function is
to maintain cell and tissue integrity. Vimentin is associated with
tumor invasion and a poor prognosis in a number of types of
cancers, including breast cancer, prostate cancer, melanoma and
lung cancer, and serves as a potential target for cancer therapy
(36,37).
The current study verified that the expression level
of GAS5 is significantly downregulated in HCC tissues compared with
adjacent normal controls. Decreased expression of GAS5 was
associated closely with the tumor differentiation (P<0.010) and
PVTT (P=0.001). The downregulated expression of GAS5 was associated
with poor prognosis of HCC. GAS5 overexpression decreases hepatoma
cell proliferation and invasion. In addition, GAS5 overexpression
promotes the apoptosis of hepatoma cells. It was further
demonstrated that GAS5 is involved in the EMT of HCC cells.
Overexpression of GAS5 downgegulated the vimentin level and the
upregulated E-cadherin level in hepatoma cells. A significant
negative correlation was observed between GAS5 and the vimentin
level in vivo. Notably, knockdown of vimentin partially
increased GAS5-pcDNA3.1-induced inhibition of cell
proliferation.
These data suggest an important role of GAS5 in the
molecular etiology of HCC and implicate the potential application
of GAS5 in HCC therapy.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant nos. 81472268/H1617 and
81272692/H1617).
References
|
1
|
Venook AP, Papandreou C, Furuse J and de
Guevara LL: The incidence and epidemiology of hepatocellular
carcinoma: A global and regional perspective. Oncologist. 15(Suppl
4): S5–S13. 2010. View Article : Google Scholar
|
|
2
|
El-Serag HB and Rudolph KL: Hepatocel lu
la r carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ørom UA, Derrien T, Beringer M, Gumireddy
K, Gardini A, Bussotti G, Lai F, Zytnicki M, Notredame C, Huang Q,
et al: Long noncoding RNAs with enhancer-like function in human
cells. Cell. 143:46–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gibb EA, Brown CJ and Lam WL: The
functional role of long non-coding RNA in human carcinomas. Mol
Cancer. 10(38)2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kogo R, Shimamura T, Mimori K, Kawahara K,
Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, et al:
Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin
modification and is associated with poor prognosis in colorectal
cancers. Cancer Res. 71:6320–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou Y, Zhang X and Klibanski A: MEG3
noncoding RNA: A tumor suppressor. J Mol Endocrinol. 48:R45–R53.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang X, Gejman R, Mahta A, Zhong Y, Rice
KA, Zhou Y, Cheunsuchon P, Louis DN and Klibanski A: Maternally
expressed gene 3, an imprinted noncoding RNA gene, is associated
with meningioma pathogenesis and progression. Cancer Res.
70:2350–2358. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schneider C, King RM and Philipson L:
Genes specifically expressed at growth arrest of mammalian cells.
Cell. 54:787–793. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Smith CM and Steitz JA: Classification of
gas5 as a multi-small-nucleolar-RNA (snoRNA) host gene and a member
of the 5′-terminal oligopyrimidine gene family reveals common
features of snoRNA host genes. Mol Cell Biol. 18:6897–6909. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Williams GT and Farzaneh F: Are snoRNAs
and snoRNA host genes new players in cancer? Nat Rev Cancer.
12:84–88. 2012.PubMed/NCBI
|
|
12
|
Yu X and Li Z: Long non-coding RNA growth
arrest-specific transcript 5 in tumor biology. Oncol Lett.
10:1953–1958. 2015.PubMed/NCBI
|
|
13
|
Glover AR, Zhao JT, Ip JC, Lee JC,
Robinson BG, Gill AJ, Soon PS and Sidhu SB: Long noncoding RNA
profiles of adrenocortical cancer can be used to predict
recurrence. Endocr Relat Cancer. 22:99–109. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mourtada-Maarabouni M, Pickard MR, Hedge
VL, Farzaneh F and Williams GT: GAS5, a non-protein-coding RNA,
controls apoptosis and is downregulated in breast cancer. Oncogene.
28:195–208. 2009. View Article : Google Scholar
|
|
15
|
Liao J, Yu L, Mei Y, Guarnera M, Shen J,
Li R, Liu Z and Jiang F: Small nucleolar RNA signatures as
biomarkers for non-small-cell lung cancer. Mol Cancer. 9(198)2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gee HE, Buffa FM, Camps C, Ramachandran A,
Leek R, Taylor M, Patil M, Sheldon H, Betts G, Homer J, et al: The
small-nucleolar RNAs commonly used for microRNA normalisation
correlate with tumour pathology and prognosis. Br J Cancer.
104:1168–1177. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee J, Kotliarova S, Kotliarov Y, Li A, Su
Q, Donin NM, Pastorino S, Purow BW, Christopher N, Zhang W, et al:
Tumor stem cells derived from glioblastomas cultured in bFGF and
EGF more closely mirror the phenotype and genotype of primary
tumors than do serum-cultured cell lines. Cancer Cell. 9:391–403.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liao J, Yu L, Mei Y, Guarnera M, Shen J,
Li R, Liu Z and Jiang F: Small nucleolar RNA signatures as
biomarkers for non-small-cell lung cancer. Mol Cancer. 9(198)2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yuan Q, Loya K, Rani B, Möbus S,
Balakrishnan A, Lamle J, Cathomen T, Vogel A, Manns MP, Ott M,
Cantz T and Sharma AD: MicroRNA-221 overexpression accelerates
hepatocyte proliferation during liver regeneration. Hepatology.
57:299–310. 2013. View Article : Google Scholar
|
|
20
|
Kapinas K, Kessler C, Ricks T, Gronowicz G
and Delany AM: miR-29 modulates Wnt signaling in human osteoblasts
through a positive feedback loop. J Biol Chem. 285:25221–25231.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takahashi K, Yan I, Haga H and Patel T:
Long noncoding RNA in liver diseases. Hepatology. 60:744–753. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu L, Ye H, Huang G, Luo F, Liu Y, Liu Y,
Yang X, Shen J, Liu Q and Zhang J: Long noncoding RNA GAS5
suppresses the migration and invasion of hepatocellular carcinoma
cells via miR-21. Tumour Biol. Sept 24–2015.Epub ahead of
print.
|
|
23
|
ENCODE Project Consortium; Birney E,
Stamatoyannopoulos JA, Dutta A, Guigó R, Gingeras TR, Margulies EH,
Weng Z, Snyder M, Dermitzakis ET, et al: Identification and
analysis of functional elements in 1% of the human genome by the
ENCODE pilot project. Nature. 447:799–816. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Amaral PP, Dinger ME, Mercer TR and
Mattick JS: The eukaryotic genome as an RNA machine. Science.
319:1787–1789. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Louro R, Smirnova AS and Verjovski-Almeida
S: Long intronic noncoding RNA transcription: Expression noise or
expression choice? Genomics. 93:291–298. 2009. View Article : Google Scholar
|
|
26
|
Zhang EB, Yin DD, Sun M, Kong R, Liu XH,
You LH, Han L, Xia R, Wang KM, Yang JS, et al: P53-regulated long
non-coding RNA TUG1 affects cell proliferation in human non-small
cell lung cancer, partly through epigenetically regulating HOXB7
expression. Cell Death Dis. 5:e12432014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Taft RJ, Pang KC, Mercer TR, Dinger M and
Mattick JS: Non-coding RNAs: Regulators of disease. J Pathol.
220:126–139. 2010. View Article : Google Scholar
|
|
28
|
Ying L, Chen Q, Wang Y, Zhou Z, Huang Y
and Qiu F: Upregulated MALAT-1 contributes to bladder cancer cell
migration by inducing epithelial-to-mesenchymal transition. Mol
Biosyst. 8:2289–2294. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tani H, Torimura M and Akimitsu N: The RNA
degradation pathway regulates the function of GAS5 a non-coding RNA
in mammalian cells. PLoS One. 8:e556842013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Qiao HP, Gao WS, Huo JX and Yang ZS: Long
non-coding RNA GAS5 functions as a tumor suppressor in renal cell
carcinoma. Asian Pac J Cancer Prev. 14:1077–1082. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cao S, Liu W, Li F, Zhao W and Qin C:
Decreased expression of lncRNA GAS5 predicts a poor prognosis in
cervical cancer. Int J Clin Exp Pathol. 7:6776–6783.
2014.PubMed/NCBI
|
|
32
|
Pickard MR, Mourtada-Maarabouni M and
Williams GT: Long non-coding RNA GAS5 regulates apoptosis in
prostate cancer cell lines. Biochim Biophys Acta. 1832:1613–1623.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zeisberg M and Neilson EG: Biomarkers for
epithelial-mesenchymal transitions. J Clin Invest. 119:1429–1437.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Satelli A and Li S: Vimentin in cancer and
its potential as a molecular target for cancer therapy. Cell Mol
Life Sci. 68:3033–3046. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lehtinen L, Ketola K, Makela R, Mäkelä R,
Mpindi JP, Viitala M, Kallioniemi O and Iljin K: High-throughput
RNAi screening for novel modulators of vimentin expression
identifies MTHFD2 as a regulator of breast cancer cell migration
and invasion. Oncotarget. 4:48–63. 2013. View Article : Google Scholar : PubMed/NCBI
|