Introduction
Chronic obstructive pulmonary disease (COPD) is
characterized by reduced airflow, which is typically progressive,
and associated with an abnormal inflammatory response of the lungs
to noxious particles and gases. The pathogenesis of COPD involves
several processes, including pulmonary and systemic inflammation,
protease/anti-protease imbalance, vascular and extracellular matrix
(ECM) remodeling, and tissue repair (1,2).
Previous research reported that increased levels of macrophages,
neutrophils and cytotoxic (CD8+) T lymphocytes were
recruited into airway walls during COPD; additionally, numerous
proinflammatory mediators and cytokines were observed in the airway
(1,3,4).
Nuclear factor (NF)-κB is a nuclear transcription factor. Following
an inflammatory stimulus, phosphorylation of inhibitor of κB (IκB)
triggers its degradation and the translocation of NF-κB to the
nucleus, where it induces the expression of a variety of genes,
including cytokines [interleukin (IL)-1β, IL-6 and tumor necrosis
factor (TNF)-α], enzymes [cyclooxygenase-2 and matrix
metal-loproteinases (MMPs)], adhesion molecules and acute phase
proteins (5). IL-6 and TNF-α are
critical proinflammatory factors, and MMPs are closely associated
with lung injury and repair in patients with COPD. The functions of
these inflammatory factors and MMPs are dependent on each other.
The expression and activity of MMP-9 (also termed gelatinase B) can
be induced by TNF-α, with its induction potentially associated with
the NF-κB-mediated signal transduction pathway. TNF-α-induced MMP-9
activity is suppressed by 18β-glycyrrhetinic acid via inhibition of
the Akt-dependent NF-κB pathway (6). Resveratrol
(trans-3,4′,5-trihydroxys-tilbene), a natural polyphenol, is
present in various plants, nuts and fruits. It is particularly
abundant in grapes and red wine. Numerous previous studies have
demonstrated that resveratrol has a wide range of pharmacological
properties; for example, there is considerable evidence
demonstrating that resveratrol has anti-inflammatory properties
(1,7,8),
with one study observing that resveratrol attenuates
lipopolysaccharide (LPS)-induced acute kidney injury by suppressing
inflammation driven by macrophages (9). Resveratrol also has a therapeutic
effect on house dust mite-induced mouse allergic asthma and its
mechanism of action may be associated with reducing the production
of spleen tyrosine kinase protein (10). Additionally, resveratrol has been
demonstrated to protect against brain injury by downregulation of
MMP-9 (11). An isoflavone
compound, genistein (4′,5,7-trihydroxyisoflavone), has been
previously shown to act as a broad-spectrum tyrosine kinase
inhibitor. Genistein possesses anti-inflammatory properties and,
therefore, may be important in modulating mammalian inflammation
pathways to inhibit certain types of chronic disease (12–14).
Previous research revealed that genistein inhibits the activation
of NF-κB and the production of TNF-α in patients with asthma
(15). Although these natural
compounds, including resveratrol and genistein, may reduce chronic
inflammation, their action on the inflammatory mediators NF-κB,
TNF-α and MMP-9 in lymphocytes from patients with COPD has not been
fully elucidated. The present study aimed to observe the effects of
resveratrol and genistein on the expression levels of NF-κB, TNF-α
and MMP-9 in patients with COPD.
Materials and methods
Patients
Patients with COPD (n=34; age, 71.8±9.0 years) were
recruited from the Department of Gerontal Respiratory Medicine of
the First Hospital of Lanzhou University (Lanzhou, China). No
patients had received any medication for 24 h or any steroids for 2
weeks prior to collection of the 10 ml blood sample by venepuncture
into a heparinized tube for immediate use. The following criteria
were required for inclusion in the present study: i) COPD confirmed
by clinical history and abnormal lung function according to the
criteria established by the Global Initiative for Chronic
Obstructive Lung Disease guidelines (16); ii) COPD diagnosis ≥2 years with ≥2
exacerbations; and iii) no long-term maintenance therapy, with the
exception of the inhalation of a short-acting bronchodilator as
required. Patients with a history or diagnosis of asthma, lung
cancer, bronchiectasis or severe heart disease were excluded from
the study. Healthy subjects were defined as having normal lung
function and no hypertension, diabetes mellitus, heart disease or
similar conditions. The age and gender distribution of healthy
subjects (n=30) was similar to those of the patients with COPD
(Table I). According to the
ethical guidelines of the Helsinki Declaration, informed consent
was obtained from all participants and was approved by the Ethical
Committee of the First Hospital of Lanzhou University.
 | Table IDemographic and clinical
characteristics of patients with COPD (n=34) and healthy subjects
(n=30). |
Table I
Demographic and clinical
characteristics of patients with COPD (n=34) and healthy subjects
(n=30).
| Characteristic | COPD patients | Healthy subjects |
|---|
| Age, years | 71.8±9.0 | 67.8±8.2 |
| Gender (male/female),
n | 23/11 | 20/10 |
| Smokers, n | 20 | 17 |
| FEV1,
% | 52.16±24.07a | 104.10±14.73 |
|
FEV1/FVC, % | 53.93±13.66a | 81.03±7.00 |
Lymphocyte preparation and experimental
groups
Lymphocytes were isolated using Ficoll-Hypaque
separation. Briefly, full blood was layered on Ficoll-Hypaque
cushions (Beijing Solarbio Science and Technology Co., Ltd.,
Beijing, China) and centrifuged for 20 min at 1,000 × g.
Lymphocytes were collected from the interface, washed twice with
phosphate-buffered saline (PBS; Beijing Solarbio Science and
Technology Co., Ltd.) and suspended in complete RPMI-1640 medium
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented
with 10% fetal calf serum (Thermo Fisher Scientific, Inc.), 2 mM
L-glutamine (Beijing Solarbio Science and Technology Co., Ltd.),
100 IU/ml penicillin and 100 µg/ml streptomycin (North China
Pharmaceutical Group Corporation, Shijiazhuang, China), at a
concentration of 2×106 cells/ml. The viability of
lymphocytes was determined using trypan blue (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) exclusion assay
(17). Viable lymphocytes were
seeded into 24-well culture plates and were maintained in a
humidified incubator at 37°C and a 5% CO2 atmosphere.
Lymphocytes from patients with COPD and healthy subjects were
randomly divided into four groups: Untreated control group,
dexamethasone (DEX; Jiangsu Lianshui Pharmaceutical Co., Ltd.,
Lianshui, China; 0.5 µmol/l), resveratrol (12.5
µmol/l; Sigma-Aldrich, St. Louis, MO, USA) and genistein (25
µmol/l; Sigma-Aldrich) groups. After 1 h of treatment, 100
µl lymphocytes were smeared onto slides for NF-κB
immunocytochemistry staining. After 48 h, the suspension was
centrifuged at 400 × g for 10 min and the supernatant of the
lymphocytes was collected, and stored at −80°C for use in TNF-α and
MMP-9 assays.
Concentration-effect curves
A kinetic study of the effects of resveratrol and
genistein on the proportion of NF-κB-positive cells, and the
secretion of TNF-α and MMP-9 in lymphocytes was performed.
Lymphocytes from patients with COPD were treated with 0, 3.125,
6.25, 12.5, 25 and 50 µmol/l resveratrol, and 0, 6.25, 12.5,
25, 50 and 100 µmol/l genistein for 1 h for NF-κB
immunocytochemistry staining, and 48 h for TNF-α and MMP-9 assays.
Concentration-effect curves of NF-κB-positive cell proportion, and
TNF-α and MMP-9 concentration following resveratrol and genistein
treatment were produced.
Immunocytochemistry
Lymphocytes (100 µl at 2×106
density) were smeared on slides and fixed in 4% paraformaldehyde
(Beijing Solarbio Science and Technology Co., Ltd.) for 30 min.
Slides were then washed twice for 3 min in PBS and soaked in 3%
H2O2 (1:50 dilution; Beijing Haiderun
Pharmaceutical Co., Ltd., Beijing, China) for 30 min to inactivate
endogenous peroxidase. After 3 further 3-min washes in PBS, the
slides were incubated with 50 µl goat serum (Beijing
Solarbio Science and Technology Co., Ltd.) for 20 min at room
temperature to block non-specific binding. Rabbit monoclonal NF-κB
p65 antibody (50 µl; 1:100; Wuhan Boster Biological
Technology Ltd., Wuhan, China; cat. no. PB0321) was then added to
the cells and incubated overnight at 4°C. Slides were washed 3
times in PBS for 3 min and then incubated with monoclonal
biotinylated goat anti-rabbit immunoglobulin G (Wuhan Boster
Biological Technology Ltd.; cat. no. BA1003) for 20 min at 37°C.
Slides were washed 3 times in PBS for 3 min and incubated with 50
µl streptavidin-peroxidase (SABC kit; Wuhan Boster
Biological Technology Ltd.) for 20 min at 37°C. The slides were
washed 4 times in PBS for 5 min prior to addition of the peroxidase
substrate (100 µl 3,3′-diaminobenzidine) for 10 min at room
temperature. The slides were washed in distilled water for 3 min.
Finally, the slides were dried naturally, then cleared in xylene
(Tianjin Fu Chen Chemical Reagents Factory, Tianjin, China) and
mounted in gelatin (Amresco, LLC, Solon, OH, USA). Negative reagent
controls were stained in parallel with the primary anti-NF-κB p65
antibody, and blocked by incubation overnight at 4°C with a
specific blocking peptide (Wuhan Boster Biological Technology Ltd.,
Wuhan, China). For each sample, two slides were scored and ≥500
cells were counted randomly on each slide using an Olympus DP71
(Olympus Corporation, Tokyo, Japan). NF-κB p65-positive cells were
defined as staining brown in the cellular nucleus and used to
calculate the percentage of NF-κB-positive lymphocytes.
Radioimmunoassay
The concentration level of TNF-α in the lymphocyte
supernatant was measured using a radioimmunoassay kit (Beijing
Chemclin Biotech Co., Ltd., Beijing, China), according to the
manufacturer's protocol.
Enzyme-linked immunosorbent assay
(ELISA)
The concentration level of MMP-9 in the lymphocyte
supernatant was measured using an ELISA kit (Wuhan Boster
Biological Technology Ltd.), according to the manufacturer's
protocol.
Statistical analysis
The data are presented as the mean ± standard
deviation and were analyzed using SPSS software version 14.0 (SPSS,
Inc., Chicago, IL, USA). Statistical analysis was conducted using
Student's t-test for the independent samples in two groups,
analysis of variance and least significant difference test for the
comparison of multiple groups and the Pearson linear correlation
was used for correlation analysis. P<0.05 was considered to
indicate a statistically significant difference.
Results
NF-κB, TNF-α and MMP-9 signaling pathways
are significantly activated in patients with COPD
Compared with healthy subjects, the proportion of
NF-κB-positive cells, and the concentrations of TNF-α and MMP-9 in
the supernatant were significantly increased in COPD lymphocytes
(Table II). Additionally, there
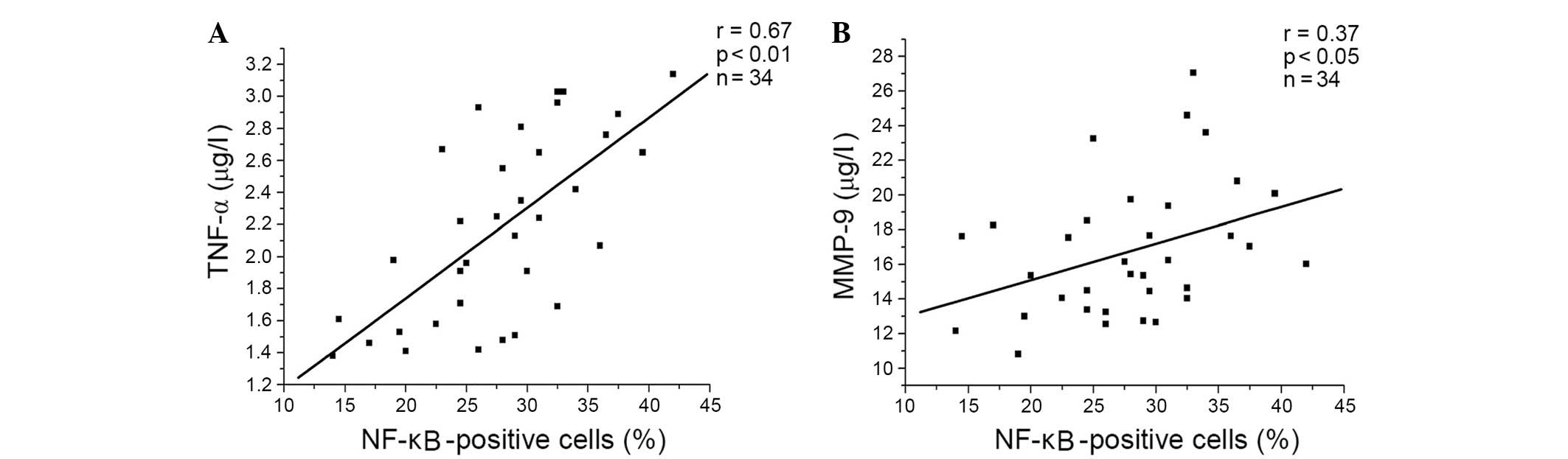
was a significant positive correlation between the percentage of
NF-κB-positive cells, and the levels of TNF-α and MMP-9 in patients
with COPD (Fig. 1).
| Table IIPercentage of NF-κB-positive cells,
and the levels of TNF-α and MMP-9 in patients with COPD and healthy
subjects. |
Table II
Percentage of NF-κB-positive cells,
and the levels of TNF-α and MMP-9 in patients with COPD and healthy
subjects.
| Group | NF-κB-positive
cells, % | TNF-α,
µg/l | MMP-9,
µg/l |
|---|
| Healthy
subjects | 6.5±2.2 | 0.89±0.29 | 10.21±2.67 |
| COPD | 27.9±6.8a | 2.19±0.57a | 16.06±3.60a |
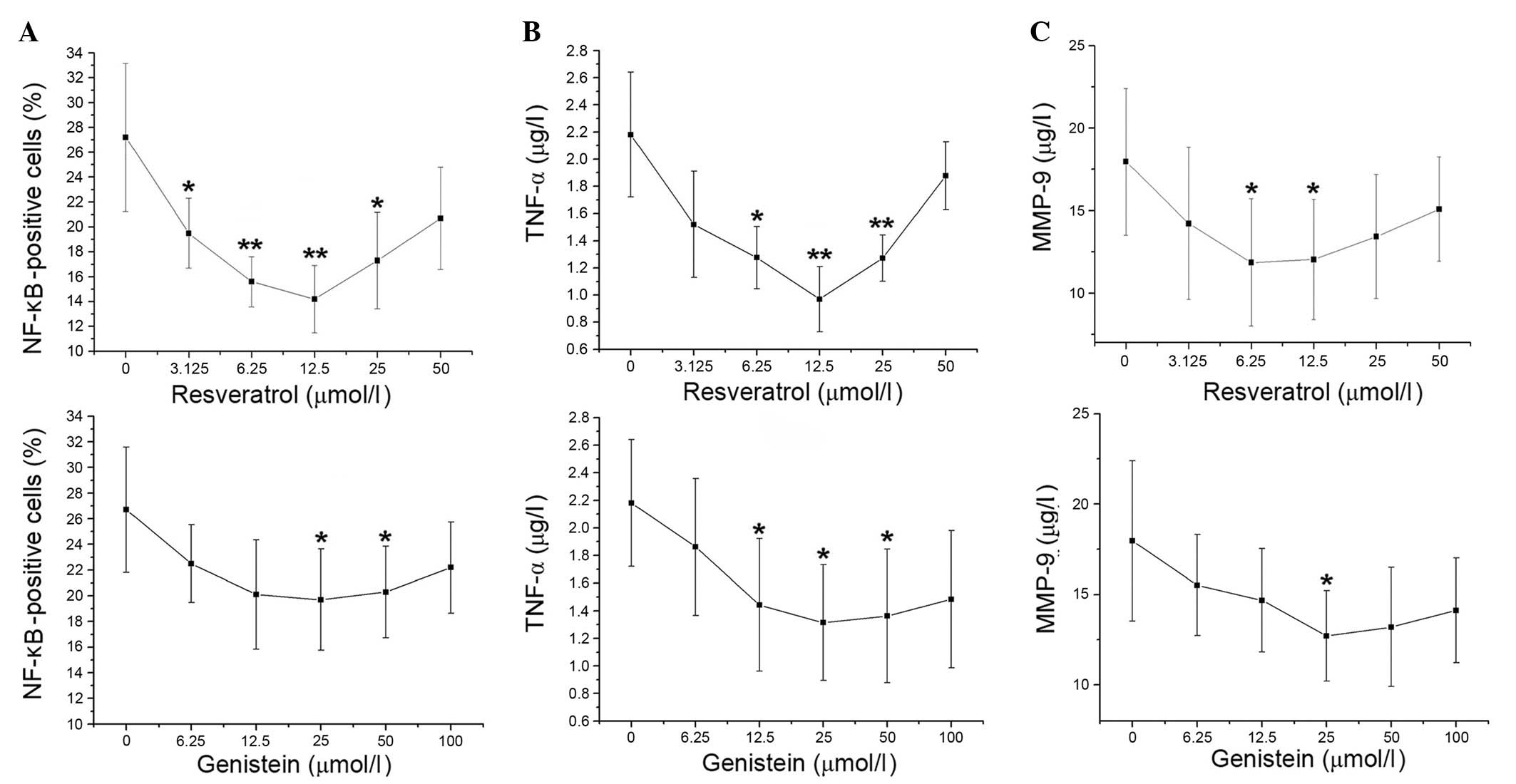
Concentration-effect curves of
resveratrol and genistein in patients with COPD
Compared with control treatment, the percentage of
NF-κB-positive cells decreased upon increasing concentrations of
resveratrol (3.125–12.5 µmol/l; P=0.003 and P= 0.002,
respectively). The proportion of NF-κB-positive cells then
gradually increased at 25 and 50 µmol/l resveratrol. The
percentage of NF-κB-positive cells was decreased at genistein
concentrations of 6.25–25 µmol/l, then increased at 50 and
100 µmol/l genistein. The proportion of NF-κB-positive cells
significantly decreased compared with the control at 25 and 50
µmol/l genistein (P=0.038 and P=0.046, respectively), with
the greatest reduction observed at 25 µmol/l (Fig. 2A). The concentration-effect curves
of TNF-α were similar to the percentage of NF-κB-positive cells in
the resveratrol and genistein-treated groups (Fig. 2B). Compared with the control, MMP-9
levels decreased with increasing concentrations of resveratrol
(3.125–6.25 µmol/l), with resveratrol causing the greatest
decrease to MMP-9 levels at 6.25 µmol/l (P= 0.049). MMP-9
levels stabilized at 12.5µmol/l resveratrol then marginally
increased at 25 and 50 µmol/l resveratrol. Additionally, the
levels of MMP-9 were gradually decreased by increasing genistein
treatment (6.25–25 µmol/l) compared with the control, with
the greatest reduction observed at 25 µmol/l genistein
(P=0.015). At higher genistein concentrations, the levels of MMP-9
rose slightly (Fig. 2C). Thus, the
optimal concentrations of resveratrol and genistein were 12.5 and
25 µmol/l, respectively.
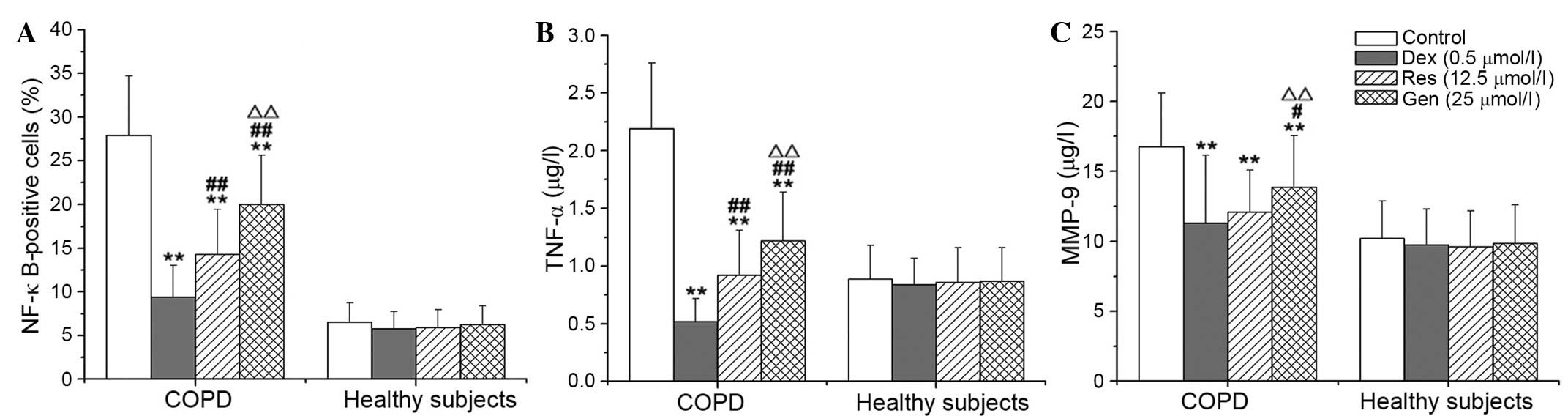
Resveratrol and genistein inhibit the
NF-κB, TNF-α and MMP-9 signaling pathways
Compared with no treatment, DEX, resveratrol and
genistein significantly reduced the percentage of NF-κB-positive
cells, and the concentration levels of TNF-α and MMP-9 in
lymphocytes from patients with COPD (P<0.001). The inhibitory
effects of resveratrol and genistein were not as potent as DEX, and
the effects of resveratrol were significantly greater than those of
genistein (P<0.001; Fig. 3).
DEX, resveratrol and genistein had no significant effect on NF-κB,
TNF-α or MMP-9 in the healthy control subjects. Additionally, the
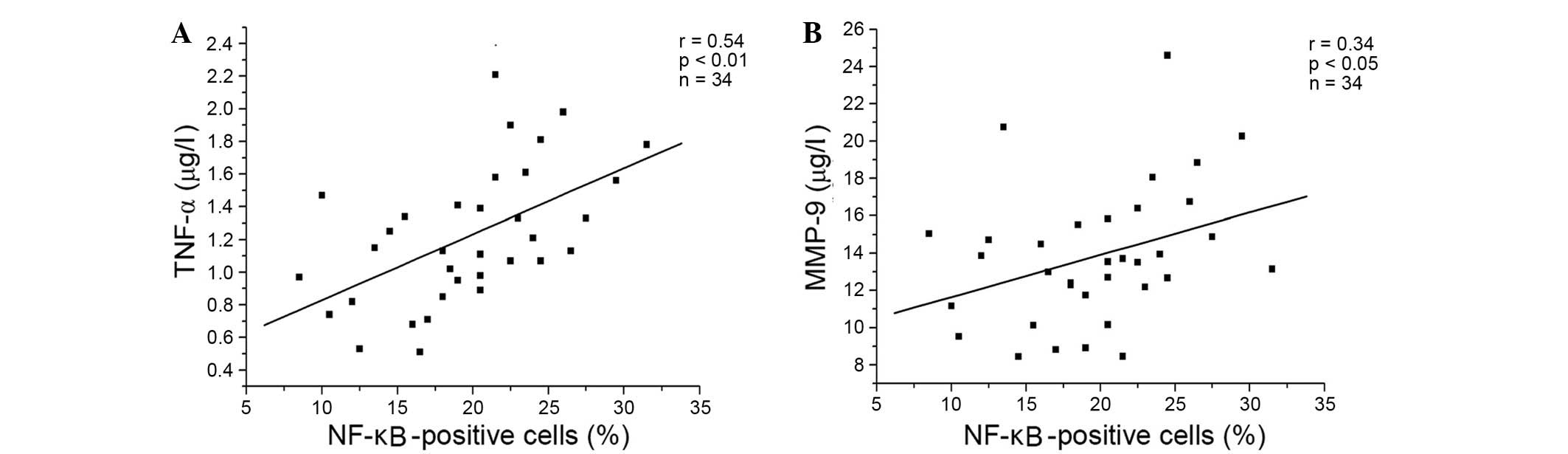
present study demonstrated positive correlations between the
percentage of NF-κB-positive cells, and the levels of TNF-α
(P<0.001) and MMP-9 (P=0.034) in the resveratrol-treated group
(Fig. 4A and B, respectively), and
the genistein-treated group (Fig. 5A
and B, respectively) in patients with COPD. A similar trend in
correlation curves was observed between Figs. 4 and 5 and Fig.
1.
Discussion
COPD is associated with chronic inflammation
predominantly affecting the lung parenchyma and peripheral airways,
resulting in largely irreversible and progressive airflow
limitation. This inflammation is characterized by increased levels
of alveolar macrophages, neutrophils and T lymphocytes, which are
recruited from the circulation. Oxidative stress is important in
driving this inflammation (1).
NF-κB is known to be critical in the regulation of proinflammatory
molecules during cellular responses, particularly TNF-α, IL-6 and
IL-8. When phosphorylated, IκB dissociates from the NF-κB-IκB
complex, resulting in the translocation of NF-κB from the cytoplasm
to the nucleus. Activation of NF-κB is regarded as an important
initial event in the airway inflammatory response to a variety of
stimuli, including infectious agents, toxins, cytokines, growth
factors and oxidant stress (1,5,18).
NF-κB may represent a link between inflammation and oxidative
stress in chronic inflammatory diseases (19). The data of the present study
demonstrated that the percentage of nuclear NF-κB-positive cells in
lymphocytes was significantly increased in COPD patients compared
with healthy controls. This indicates that translocation and
activation of NF-κB is upregulated in patients with COPD. The
expression of TNF-α, a proinflammatory cytokine, is regulated by
NF-κB and TNF-α activates NF-κB expression (12). TNF-α has an important amplifying
effect in COPD airway inflammation. Furthermore, circulating TNF-α
is partially associated with hypoxemia, and increased systemic
TNF-α levels have previously been demonstrated to be involved in
cachexia and skeletal muscle weakness in patients with COPD
(1). MMPs are a family of
zinc-dependent endopeptidases that are characterized by their
ability to degrade components of the ECM. Previous reports have
implicated MMPs in numerous physiological and pathophysiological
processes, including tissue remodeling, angiogenesis, and cell
differentiation and migration. Typically, the enzymatic activity of
MMPs is tightly controlled by the secretion of endogenous specific
tissue inhibitors of MMPs, however, a disruption of this balance is
observed in COPD and other active inflammatory diseases, resulting
in tissue injury (2,20). MMP-9 has previously been suggested
to be important in the process of COPD and associated with tissue
remodeling (21,22). The data of the current study also
demonstrated that the levels of TNF-α and MMP-9 were increased in
the lymphocytes of patients with COPD compared with healthy
controls, and there were positive correlations between the
percentage of NF-κB-positive cells, and the concentration of TNF-α
and MMP-9 in patients with COPD.
Resveratrol is a polyphenol present in various
fruits and vegetables, and is particularly abundant in grapes. The
root extract of the weed Polygonum cuspidatum, an important
constituent of Japanese and Chinese folk medicine, is also a
plentiful source of resveratrol. The anti-inflammatory and
antioxidant properties of resveratrol have been previously reported
in numerous studies. Knobloch et al (23) demonstrated that resveratrol
attenuates the release of inflammatory cytokines from human
bronchial smooth muscle cells exposed to lipoteichoic acid in COPD.
Birrel et al (24)
demonstrated that resveratrol has anti-inflammatory effects in rat
lungs challenged with LPS. Furthermore, resveratrol has been
demonstrated to be a potent inhibitor of TNF-α-induced activation
of NF-κB, and can suppress TNF-α-activated inflammatory pathways
in vitro and in vivo (12). The suppression of TNF-α has been
observed in normal and tumor cells. Resveratrol also inhibited
NF-κB activation induced by a wide variety of inflammatory agents.
The ability of resveratrol to block reactive oxygen species (ROS)
generation and lipid peroxidation induced by TNF-α may be
associated with its effects on transcription factors and associated
kinases. Furthermore, Lee and Kim (25) demonstrated that resveratrol reduces
MMP-9 activation through the inhibition of ROS, and the activation
of NF-κB and activator protein-1 in human fibrosarcoma cells.
Resveratrol also inhibited TNF-α-mediated MMP-9 expression and
invasion in human hepatocellular carcinoma cells (26). Knobloch et al (27) reported that resveratrol almost
completely reduces MMP-9 release from LPS-exposed alveolar
macrophages in patients with COPD. The findings of the present
study demonstrate that the activation of NF-κB, and the
hypersecretion of TNF-α and MMP-9 in lymphocytes from patients with
COPD were inhibited by resveratrol. Additionally, the current study
observed that there were positive correlations between the
percentage of NF-κB-positive cells, and the concentration levels of
TNF-α and MMP-9 following resveratrol treatment of lymphocytes from
patients with COPD. This indicates that resveratrol inhibition of
TNF-α and MMP-9 may be involved in NF-κB deactivation.
Genistein, the major isoflavone present in soy,
scavenges free radicals, inhibits tyrosine kinases and possesses
anti-inflammatory properties. Previous research demonstrated that
increased consumption of genistein is associated with improved lung
function in patients with asthma (28). Additionally, dietary soy isoflavone
supplementation reduces eosinophil leukotriene C4 synthesis and
eosinophilic airway inflammation in patients with asthma (29). In a novel finding, Hirayama et
al (30) reported that there
was a significant positive association between lung function and
the intake of isoflavones, including genistein and daidzein. In
this previous study, total isoflavone, genistein and daidzein were
significantly associated with decreased risk of COPD and
breathlessness. Thus, it is likely that isoflavones and their
aglycones act as anti-inflammatory agents in the lung and may
protect against tobacco carcinogens. Gupta et al (12) reviewed these polyphenols,
suggesting that they may protect against pro-inflammatory diseases
by downregulation of TNF-α and other proinflammatory biomarkers.
Other previous findings indicate that genistein suppresses
TNF-α-induced inflammation by inhibiting the ROS/Akt/NF-κB pathway
and promoting AMP-activated protein kinase activation (31,32).
Additionally, Wu et al (33) reported that IL-1β-induced
upregulation of proMMP-9 was inhibited by genistein in rat brain
astrocytes. The data of the present study demonstrated that
genistein inhibits the translocation of NF-κB to the nucleus, and
decreases the hypersecretion of TNF-α and MMP-9 in lymphocytes from
patients with COPD. Additionally, the current study observed that
there were positive correlations between the percentage of
NF-κB-positive cells, and the levels of TNF-α and MMP-9 following
genistein treatment of lymphocytes from patients with COPD. The
inhibitory effects of resveratrol and genistein on NF-κB
localization, and TNF-α and MMP-9 levels were not as effective as
DEX treatment (a commonly used anti-inflammatory positive control),
and genistein was not as effective as resveratrol. The inhibitory
actions of resveratrol and genistein on TNF-α were greater that the
effects on MMP-9. It is possible that the anti-inflammatory
activity of resveratrol and genistein may be more potent than the
effects on airway remodeling in patients with COPD.
In conclusion, the present study demonstrated that
the translocation of NF-κB from the cytoplasm to the nucleus, and
the secretion of TNF-α and MMP-9 were increased in lymphocytes from
patients with COPD compared with healthy controls. Additionally,
resveratrol and genistein inhibited the translocation of NF-κB, and
decreased TNF-α and MMP-9 concentration levels. Furthermore, the
inhibitory actions of resveratrol and genistein on TNF-α were
greater than the effects on MMP-9. The results of the present study
suggest that resveratrol and genistein may be potential drug
candidates for the treatment of COPD.
Acknowledgments
The present study was supported by The Gansu Healthy
Foundation (no. GSWST2010-05) and Gansu Science and Technology
Support Program (no. 144FKCA062).
References
|
1
|
Barnes PJ: Cellular and molecular
mechanisms of chronic obstructive pulmonary disease. Clin Chest
Med. 35:71–86. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Eurlings IM, Dentener MA, Mercken EM, de
Cabo R, Bracke KR, Vernooy JH, Wouters EF and Reynaert NL: A
comparative study of matrix remodeling in chronic models for COPD;
mechanistic insights into the role of TNF-α. Am J Physiol Lung Cell
Mol Physiol. 307:L557–L565. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mohan D, Gale NS, McEniery CM, Bolton CE,
Cockcroft JR, MacNee W, Fuld J, Lomas DA, Calverley PM, Shale DJ,
et al ERICA Consortium: Evaluating the role of inflammation in
chronic airways disease: The ERICA study. COPD. 11:552–559. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kersul AL, Iglesias A, Ríos Á, Noguera A,
Forteza A, Serra E, Agustí A and Cosío BG: Molecular mechanisms of
inflammation during exacerbations of chronic obstructive pulmonary
disease. Arch Bronconeumol. 47:176–183. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Barnes PJ and Karin M: Nuclear
factor-kappaB: A pivotal transcription factor in chronic
inflammatory diseases. N Engl J Med. 336:1066–1071. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jayasooriya RG, Dilshara MG, Park SR, Choi
YH, Hyun JW, Chang WY and Kim GY: 18β-Glycyrrhetinic acid
suppresses TNF-α induced matrix metalloproteinase-9 and vascular
endothelial growth factor by suppressing the Akt-dependent NF-κB
pathway. Toxicol In Vitro. 28:751–758. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tung BT, Rodríguez-Bies E, Talero E,
Gamero-Estévez E, Motilva V, Navas P and López-Lluch G:
Anti-inflammatory effect of resveratrol in old mice liver. Exp
Gerontol. 64:1–7. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Park EJ and Pezzuto JM: The pharmacology
of resveratrol in animals and humans. Biochim Biophys Acta.
1852.1071–1113. 2015.
|
|
9
|
Chen L, Yang S, Zumbrun EE, Guan H,
Nagarkatti PS and Nagarkatti M: Resveratrol attenuates
lipopolysaccharide-induced acute kidney injury by suppressing
inflammation driven by macrophages. Mol Nutr Food Res. 59:853–864.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen J, Zhou H, Wang J, Zhang B, Liu F,
Huang J, Li J, Lin J, Bai J and Liu R: Therapeutic effects of
resveratrol in a mouse model of HDM-induced allergic asthma. Int
Immunopharmacol. 25:43–48. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shao AW, Wu HJ, Chen S, Ammar AB, Zhang JM
and Hong Y: Resveratrol attenuates early brain injury after
subarachnoid hemorrhage through inhibition of NF-κB-dependent
inflammatory/MMP-9 pathway. CNS Neurosci Ther. 20:182–185. 2014.
View Article : Google Scholar
|
|
12
|
Gupta SC, Tyagi AK, Deshmukh-Taskar P,
Hinojosa M, Prasad S and Aggarwal BB: Downregulation of tumor
necrosis factor and other proinflammatory biomarkers by
polyphenols. Arch Biochem Biophys. 559:91–99. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gao F, Wei D, Bian T, Xie P, Zou J, Mu H,
Li J, Lin J, Bai J and Liu R: Genistein attenuated allergic airway
inflammation by modulating the transcription factors T-bet, GATA-3
and STAT-6 in a murine model of asthma. Pharmacology. 89:229–236.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Dong J, He P, Li W, Zhang Q, Li N
and Sun T: Genistein inhibit cytokines or growth factor-induced
proliferation and transformation phenotype in fibroblast-like
synoviocytes of rheumatoid arthritis. Inflammation. 35:377–387.
2012. View Article : Google Scholar
|
|
15
|
Liu XJ, Zhao J and Gu XY: The effects of
genistein and puerarin on the activation of nuclear factor-kappaB
and the production of tumor necrosis factor-alpha in asthma
patients. Pharmazie. 65:127–131. 2010.PubMed/NCBI
|
|
16
|
Global Initiative for Chronic Obstructive
Lung Disease(GOLD): Global Strategy for the Diagnosis, Management,
and Prevention of Chronic Obstructive Pulmonary Disease. http://www.goldcopd.org.
Accessed December 31, 2011.
|
|
17
|
Martin-Piedra MA, Garzon I, Oliveira AC,
Alfonso-Rodriguez CA, Carriel V, Scionti G and Alaminos M: Cell
viability and proliferation capability of long-term human dental
pulp stem cell cultures. Cytotherapy. 16:266–277. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Singh R, Mackay AJ, Patel AR, Garcha DS,
Kowlessar BS, Brill SE, Donnelly LE, Barnes PJ, Donaldson GC and
Wedzicha JA: Inflammatory thresholds and the species-specific
effects of colonising bacteria in stable chronic obstructive
pulmonary disease. Respir Res. 15:1142014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fischer BM, Voynow JA and Ghio AJ: COPD:
Balancing oxidants and antioxidants. Int J Chron Obstruct Pulmon
Dis. 10:261–276. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hirota N and Martin JG: Mechanisms of
airway remodeling. Chest. 144:1026–1032. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yao H, Hwang JW, Sundar IK, Friedman AE,
McBurney MW, Guarente L, Gu W, Kinnula VL and Rahman I: SIRT1
redresses the imbalance of tissue inhibitor of matrix
metalloproteinase-1 and matrix metalloproteinase-9 in the
development of mouse emphysema and human COPD. Am J Physiol Lung
Cell Mol Physiol. 305:L615–L624. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gross NJ: Novel antiinflammatory therapies
for COPD. Chest. 142:1300–1307. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Knobloch J, Wahl C, Feldmann M, Jungck D,
Strauch J, Stoelben E and Koch A: Resveratrol attenuates the
release of inflammatory cytokines from human bronchial smooth
muscle cells exposed to lipoteichoic acid in chronic obstructive
pulmonary disease. Basic Clin Pharmacol Toxicol. 114:202–209. 2014.
View Article : Google Scholar
|
|
24
|
Birrell MA, McCluskie K, Wong S, Donnelly
LE, Barnes PJ and Belvisi MG: Resveratrol, an extract of red wine,
inhibits lipopolysaccharide induced airway neutrophilia and
inflammatory mediators through an NF-kappaB-independent mechanism.
FASEB J. 19:840–841. 2005.PubMed/NCBI
|
|
25
|
Lee SJ and Kim M: Resveratrol with
antioxidant activity inhibits matrix metalloproteinase via
modulation of SIRT1 in human fibrosarcoma cells. Life Sci.
88:465–472. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu H, Pan C, Zhao S, Wang Z, Zhang H and
Wu W: Resveratrol inhibits tumor necrosis factor-alpha-mediated
matrix metalloproteinase-9 expression and invasion of human
hepatocellular carcinoma cells. Biomed Pharmacother. 62:366–372.
2008. View Article : Google Scholar
|
|
27
|
Knobloch J, Hag H, Jungck D, Urban K and
Koch A: Resveratrol impairs the release of steroid-resistant
cytokines from bacterial endotoxin-exposed alveolar macrophages in
chronic obstructive pulmonary disease. Basic Clin Pharmacol
Toxicol. 109:138–143. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bime C, Wei CY, Holbrook J, Smith LJ and
Wise RA: Association of dietary soy genistein intake with lung
function and asthma control: A post-hoc analysis of patients
enrolled in a prospective multicentre clinical trial. Prim Care
Respir J. 21:398–404. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kalhan R, Smith LJ, Nlend MC, Nair A,
Hixon JL and Sporn PH: A mechanism of benefit of soy genistein in
asthma: Inhibition of eosinophil p38-dependent leukotriene
synthesis. Clin Exp Allergy. 38:103–112. 2008.
|
|
30
|
Hirayama F, Lee AH, Binns CW, Hiramatsu N,
Mori M and Nishimura K: Dietary intake of isoflavones and
polyunsaturated fatty acids associated with lung function,
breathlessness and the prevalence of chronic obstructive pulmonary
disease: Possible protective effect of traditional Japanese diet.
Mol Nutr Food Res. 54:909–917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li J, Li J, Yue Y, Hu Y, Cheng W, Liu R,
Pan X and Zhang P: Genistein suppresses tumor necrosis factor
α-induced inflammation via modulating reactive oxygen
species/Akt/nuclear factor κB and adenosine monophosphate-activated
protein kinase signal pathways in human synoviocyte MH7A cells.
Drug Des Devel Ther. 8:315–323. 2014. View Article : Google Scholar
|
|
32
|
Jeong JW, Lee HH, Han MH, Kim GY, Kim WJ
and Choi YH: Anti-inflammatory effects of genistein via suppression
of the toll-like receptor 4-mediated signaling pathway in
lipopolysaccharide-stimulated BV2 microglia. Chem Biol Interact.
212:30–39. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu CY, Hsieh HL, Sun CC, Tseng CP and Yang
CM: IL-1 beta induces proMMP-9 expression via c-Src-dependent
PDGFR/PI3K/Akt/p300 cascade in rat brain astrocytes. J Neurochem.
105:1499–1512. 2008. View Article : Google Scholar : PubMed/NCBI
|