Introduction
Graves' disease (GD) is an autoimmune disease of the
thyroid gland, which is characterized by hyperthyroidism, diffuse
goiter and Graves' ophthalmopathy (GO) (1). It has been reported that up to 50% of
patients with GD develop the ocular complication, GO (2,3). The
principle characteristics of GO include upper eyelid retraction,
soft tissue swelling, proptosis, strabismus, erythema of
periorbital tissues, and compressive optic neuropathy, and certain
patients with GO suffer from inflammation, diplopia, intense pain,
and compressive optic neuropathy or corneal ulceration, which
threaten vision (1,4).
Although the pathogenesis of GO remains to be fully
elucidated, it is widely accepted that the occurrence of this
disease is associated with the abnormal secretion of inflammatory
cytokines (1,5,6).
These overexpressed inflammatory cytokines promote the infiltration
of thyroid lymphocytic and the activation of B cells, which result
in the production of autoimmune antibodies against thyroid antigens
and contribute to the pathogenesis of GO. It has been demonstrated
that, when stimulated by proinflammatory cytokines, orbital
fibroblasts from patients with GO can produce excess
glycosaminoglycans and inflammatory cytokines, including
interleukin (IL)-6, and IL-8 (7–9). The
expression levels of intercellular adhesion molecule-1 (ICAM-1)
(10,11) and cyclooxygenase (COX)-2 (12,13)
have also been found to upregulated in the orbital connective
tissues of patients with GO. Currently, glucocorticoids are used as
the first-line treatment for GO due to their marked
anti-inflammatory and immunosuppressive effects. However, although
glucocorticoids are effective in a substantial number of patients
with GO, they have several long-term side effects, including
hypertension, diabetes and osteoporosis (14). Therefore, it is essential to
investigate novel therapies for the management of GO.
Traditional medicines offer an abundance of
plant-derived remedies to identify novel lead molecules for the
development of novel drugs. Celastrol is a pentacyclic
triterpenoid, which was originally isolated from Thunder God Vine
root. Celastrol has been demonstrated to exert potent inhibitory
action on tumorigenesis. Several studies have reported that
celastrol inhibits the proliferation of a variety of tumor cells
and suppresses tumor initiation, promotion and metastasis in
various cancer models in vivo (15,16).
In addition, celastrol has potent anti-inflammatory effects, and
the efficacy of celastrol as an anti-inflammatory drug has been
examined in several diseases, including rheumatoid arthritis
(17,18), allergic asthma (19), systemic lupus erythematosus
(20) and skin inflammation
(21). The nuclear factor (NF)-κB
signaling pathway is well integrated with other signaling pathways,
and is important in a number of diseases, including cancer and
inflammatory diseases (22,23).
Several studies have revealed that celastrol is an inhibitor of the
NF-κB signaling pathway, and that the mutation of cysteine 179 in
the activation loop of inhibitor of κB (IκB) kinase β (IKKβ)
eliminates sensitivity towards to celastrol, suggesting that
celastrol suppresses NF-κB activation by targeting cysteine 179 in
the IKK (24,25).
In the present study, the effect of celastrol on
IL-1β-induced inflammation was examined in orbital fibroblasts from
patients with GO. It was found that celastrol significantly
attenuated the expression levels of IL-6, IL-8, COX-2 and ICAM-1,
and inhibited the IL-1β-induced increases in the expression levels
of IL-6, IL-8, ICAM-1 and COX-2. It was also demonstrated that the
level of prostaglandin E (PGE)2 in the orbital fibroblasts induced
by IL-1β was suppressed by celastrol. Further investigation
revealed that celastrol suppressed IL-1β-induced inflammatory
responses in the orbital fibroblasts through inhibiting the
activation of NF-κB activation. Taken together, the results of the
present study suggested that celastrol attenuated the IL-1β-induced
pro-inflammatory pathway in orbital fibroblasts from patients with
GO, which was associated with the suppression of NF-κB
activation.
Materials and methods
Reagents
Celastrol was obtained from Sigma-Aldrich (St.
Louis, MO, USA). Dulbecco's modified Eagle's medium (DMEM) and
fetal bovine serum (FBS) were obtained from Life Technologies
(Grand Island, NY, USA). Penicillin, and gentamycin were purchased
from Amresco, Inc. (Framingham, MA, USA). The Cell Counting Kit-8
(CCK-8) assay kit was obtained from Dojindo Laboratories (Kumamoto,
Japan). The BAY-11-7082 and Annexin V-fluorescein isothiocyanate
(FITC) apoptosis detection kit were purchased from Beyotime
Institute of Biotechnology (Shanghai, China). IL-6, IL-8, IL-10 and
PGE2 ELISA Duoset kits, and recombinant human IL-1β were purchased
from R&D Systems, Inc. (Minneapolis, MN, USA).
Cell culture
Orbital fibroblasts were cultured from adipose
connective tissues, which were obtained from four patients with GO
(two male, two female) and severe proptosis associated with
increased orbital fat volume during a process of surgical
decompression. The control tissues were obtained from two patients
with no history of GO or autoimmune thyroid disease, and were
collected during the course of upper lid blepharoplasties from 2
individuals (one male, one female). The mean age of all subjects
was 58 years. The protocol for obtaining orbital adipose connective
tissue was approved by the Institutional Review Board of Longhua
Hospital (Shanghai, China), and written informed consent was
obtained from all patients.
GO orbital tissues samples were minced and plated
directly into culture dishes. The cells were maintained in DMEM
containing 10% FBS, penicillin (100 U/ml) and gentamycin (20
mg/ml), in a humidified 5% CO2 incubator at 37°C. When
the fibroblasts had grow to 80% confluence, the cell culture medium
was removed and the cells were washed with phosphate-buffered
saline (PBS). The fibroblasts were then passaged serially by
treatment with trypsin (Sigma-Aldrich). The cell culture medium was
replaced every 2 days, and cells between the third and seventh
passage were used for the subsequent examinations.
Cell viability assays
Cell viability was assayed using the CCK-8 according
to the manufacturer's protocol. Briefly, 100 µl cells were
seeded onto 96-well plates (1×104 cells/ml) for 24 h,
following which the cells were treated with, or without, 1
µM celastrol for 24 h at 37°C. Subsequently, 10 µl of
the CCK-8 solution was added to each well of the plate, followed by
1 h incubation at 37°C. The optical density (OD) was measured at
450 nm using a microplate reader (Multiskan MK3; Thermo Fisher
Scientific GmbH., Darmstadt, Germany). The cell inhibitory rate was
calculated according to the following equation: Cell inhibitory
rate = [1 − (OD experiment − OD blank) / (OD control − OD blank)] ×
100%. All experiments were performed in triplicate and repeated
three times independently.
Apoptosis assays
Apoptosis assays were performed according to the
manufacturer's protocols. Briefly, the cells in the logarithmic
growth phase were collected and washed with isotonic PBS, following
which 1×106 cells were seeded into 6-well cell culture
plates. After 24 h, the cell cultures were removed, and the cells
were incubated with serum-free DMEM, with or without 1 µM
celastrol, for another 24 h at 37°C. The cells were then digested
with trypsin and collected by centrifugation at 300 × g for 10 min
at 4°C. The cells were washed with ice-cold PBS, and resuspended
gently with 195 µl annexin V-FITC binding buffer and 5
µl annexin V-FITC, following which 10 µl propidium
iodide (PI) solutions were added. The mixture was incubated in the
dark at room temperature for 15 min. Cytometric analysis was
performed using a FACS Calibur flow cytometer (BD Biosciences,
Franklin Lakes, NJ, USA). Data acquisition and analysis were
performed using the WinMDI 2.9 computer program (BD
Biosciences).
Western blot analysis
The cells were collected and washed with ice-cold
PBS, following which the cells were centrifuged at 300 × g for 5
min at 4°C, and the supernatant was removed. The cells were lysed
with radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology) at 4°C for 20 min. The lysates were
centrifuged for 10 min at 12,000 × g at 4°C, and the supernatant
was collected. The protein concentration was determined using a
Bradford assay (BioRad Laboratories, Inc., Hercules, CA, USA). A
total of 30–50 µg proteins were subjected to sodium dodecyl
sulfate polyacrylamide gel electrophoresis on 10% (w/v) gels
(Beyotime Institute of Biotechnology), and were then
electrophoretically transferred onto a polyvinylidene fluoride
membrane (EMD Millipore, Billerica, MA, USA). Following blocking
with blocking buffer (Beyotime Institute of Biotechnology) for 1 h
at room temperature, the membrane was incubated with the indicated
primary antibodies overnight at 4°C. This was followed by
incubation in horseradish peroxidase (HRP)-conjugated corresponding
secondary antibodies for 1 h at room temperature. Positive signals
were visualized using ECL Advanced Solution (Bioworld Technology,
Inc., St. Louis Park, MN, USA). Actin was used as a loading
control. The primary antibodies used in the present study were as
follows: Rabbit polyclonal ICAM-1 (1:1,000; Cell Signaling
Technology, Inc., Danvers, MA, USA; cat. no. 4915), rabbit
monoclonal COX-2 (1:1,000; Cell Signaling Technology, Inc.; cat.
no. 12282) and rabbit monoclonal β-actin (1:1,000; Cell Signaling
Technology, Inc.; cat. no. 8457).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNAs were isolated from the orbital
fibroblasts using TRIzol reagent (Thermo Fisher Scientific, Inc.).
The total RNAs were reverse transcribed into cDNA using Reverse
Transcriptase M-MLV (Takara Bio, Inc., Otsu, Japan) and were
amplified using SYBR Green Master mix (Takara Bio, Inc.). The mRNA
expression was analyzed using an ABI 7500 Real-Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). Relative gene
expression levels were obtained following normalization with
β-actin. The thermocycling conditions used were as follows: 95°C
for 20 sec; 40 cycles of 95°C for 20 sec, 60°C for 30 sec and 72°C
for 30 sec. All reactions were run in triplicate. The primer
sequences used were as follows: IL-6, forward
5′-ATGAACTCCTTCTCCACAAG -3′ and reverse 5′-TGTCAATTCGTTCTGAAGAG-3′
(26); IL-8, forward
5′-GTGCAGTTTTGCCAAGGAGT-3′ and reverse 5′-TAATTTCTGTGTTGGCGCAG-3′
(26); IL-10, forward
5′-CTTCGAGATCTCCGAGATGCCTTC-3′ reverse
5′-ATTCTTCACCTGCTCCACGGCCTT-3′ (27); ICAM-1, forward
5′-CTCAGTCAGTGTGACCGCAGA-3′ and reverse
5′-CCCTTCTGAGACCTCTGGCTTC-3′ (28); COX-2, forward
5′-GCTCAAACATGATGTTTGCATTG-3′ and reverse 5′-GCTGGCCCTCGCTTATGA-3′
(29); and β-actin,
forward-TCACCCACACTGTGCCCAT-3′ and reverse
5′-TCCTTAATGTCACGCACGATTT-3′ (29). The 2−ΔΔCq method was
used to quantify the results (30).
ELISA
The orbital fibroblasts (1×106) were
seeded into 6-well cell culture plates and, after 24 h, the cell
culture medium was replaced with DMEM containing 1% FBS, and 10
ng/ml IL-1β was added, with or without 1 µM celastrol.
Following 24 h of incubation, the supernatants from the cell
cultures were collected, and the concentrations of IL-6, IL-8,
IL-10 and PEG-2 were determined using an ELISA kit, according to
the manufacturer's protocol. The absorbance was measured at 450 nm
using a microplate reader (Molecular Devices LLC, Sunnyvale, CA,
USA).
Luciferase assays
For the luciferase assays, HEK 293T cells (Cell Bank
of the Chinese Academy of Sciences, Shanghai, China) were seeded
into a 24-well plate at a density of 3×104 cells/well.
After 24 h at 37°C, HEK 293T cells were transfected with 200 ng
firefly luciferase reporter gene construct (per well) and 1 ng
pRL-SV40 Renilla luciferase constructs (per well) for
normalization, using cotransfection with 2.4 µl
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.). At
24 h post-transfection, the cells were stimulated with 100 ng/ml
lipopolysaccharide (Sigma-Aldrich), with or without 1 µM
celastrol, 4 h. Cells were subsequently collected and luciferase
activity was measured with the Dual-Luciferase® Reporter
(DLR™) assay system (Promega, Madison, WI, USA).
Statistical analysis
All experiments were performed at least three times
and the results are presented as the mean ± standard error of the
mean. Student's t-test was used to compare two independent
groups using SPSS 16.0 software (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effect of celastrol on the viability and
apoptosis of orbital fibroblasts
In the present study, orbital fibroblasts obtained
from normal controls or patients with GO were treated with
different concentrations of celastrol (0, 200, 400, 600, 800, 1, 2,
3, 4 and 5 µM) for 24 h, and cell viability was examined
using a CCK-8 assay. As shown in Fig.
1, exposure of the orbital fibroblasts from the GO and normal
groups to celastrol at concentrations ≤1 µM for 24 h led to
no significant decline in the numbers of living cells, whereas 2
µM celastrol decreased cell viability in the two groups to
85.02 and 88.94%, respectively (Fig.
1A). The results of the apoptosis assay also showed that
exposure of the cells to celastrol at 1 µM for 24 h did not
induce cell apoptosis (Fig. 1B).
Therefore, in the subsequent experiments, the cells were treated
with 1 µM celastrol for 24 h to further investigate the role
of celastrol in GO.
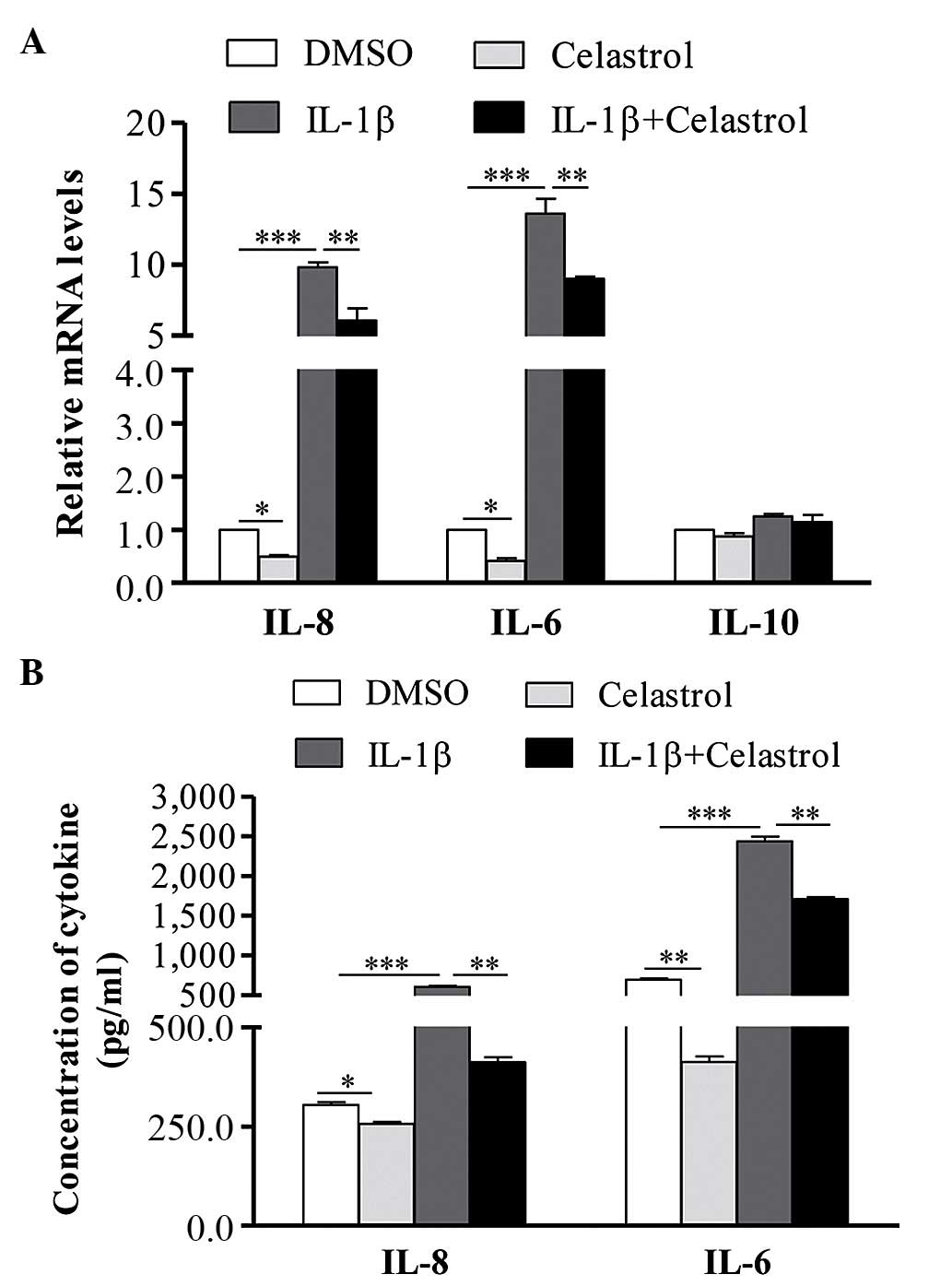
Effect of celastrol on the expression
levels of IL-1β-induced IL-6 and IL-8
As is already known, inflammation is critical in the
pathogenesis of GO, therefore, the present study examined the
expression levels of IL-6, IL-8 and IL-10 in IL-1β-induced GO cells
and normal cells, which were treated with or without celastrol. As
shown in Fig. 2A, following
treatment with IL-1β, the mRNA expression levels of IL-6 and IL-8
were significantly increased in the GO cells, whereas no change was
observed in the expression of IL-10. Celastrol was found to
decrease the mRNA expression levels of IL-6 and IL-8 in the
IL-1β-induced orbital fibroblasts. The results of the ELISA also
showed that, following stimulation with IL-1β, the levels of IL-6
and IL-8 in the orbital fibroblast supernatant were significantly
upregulated, and co-treatment of celastrol significantly attenuated
the IL-1β-induced expression of IL-6 and IL-8 (Fig. 2B).
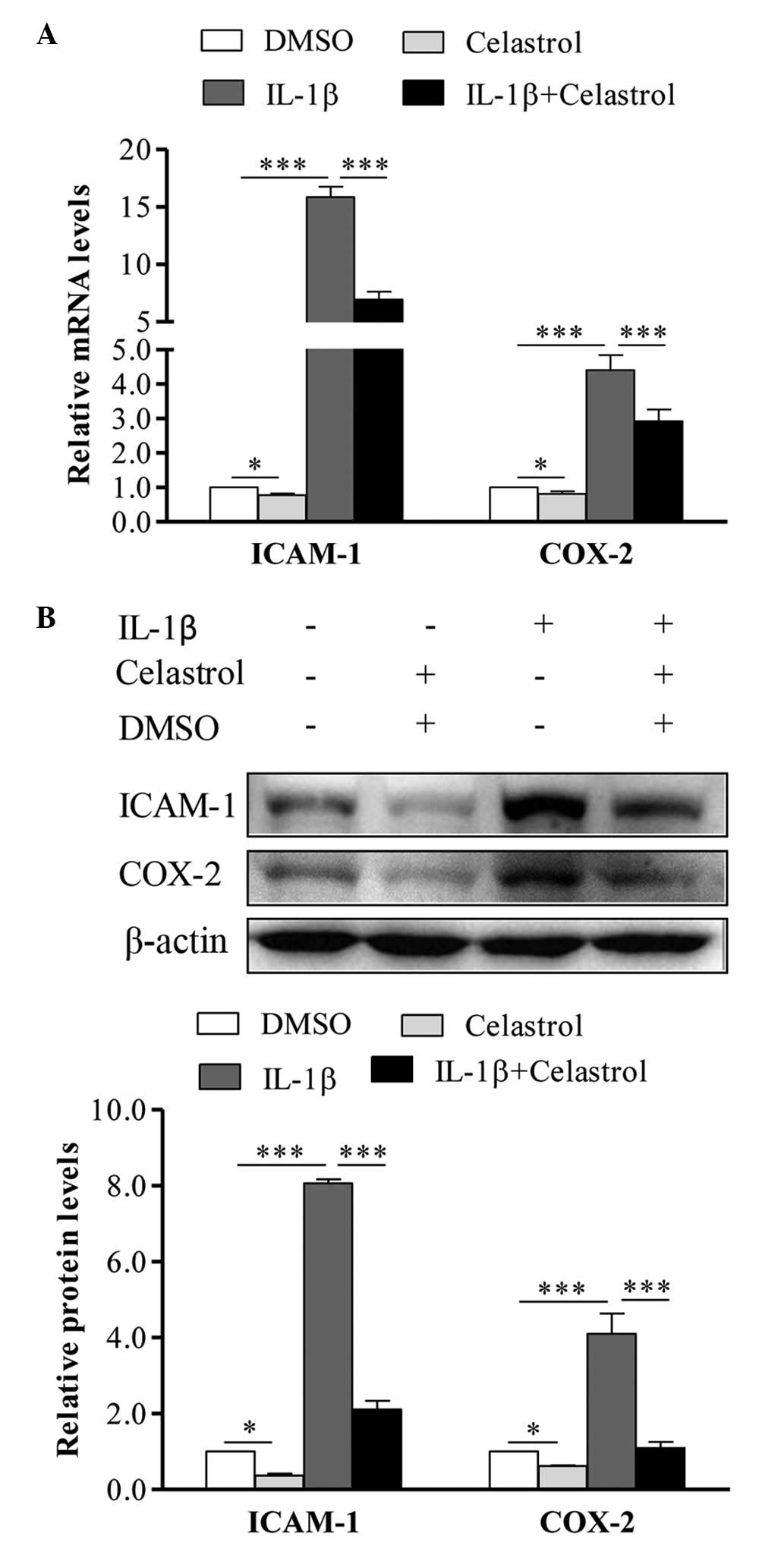
Effect of celastrol on the expression
levels of IL-1β-induced ICAM-1 and COX-2
To investigate the effect of celastrol on ICAM-1 and
COX-2, the GO cells were treated with 10 ng/ml IL-1β, with or
without 1 µM celastrol, for 24 h, following which the cells
were collected and subjected to RT-qPCR analysis. As shown in
Fig. 3A, in the IL-1β-induced
orbital fibroblasts, the mRNA expression levels of ICAM-1 and COX-2
were significantly increased, whereas treatment with celastrol
almost completely reversed the IL-1β-induced upregulation of ICAM-1
and COX-2. In addition, following treatment with IL-1β, the protein
expression levels of ICAM-1 and COX-2 were markedly enhanced, and
this was also depressed by celastrol (Fig. 3B).
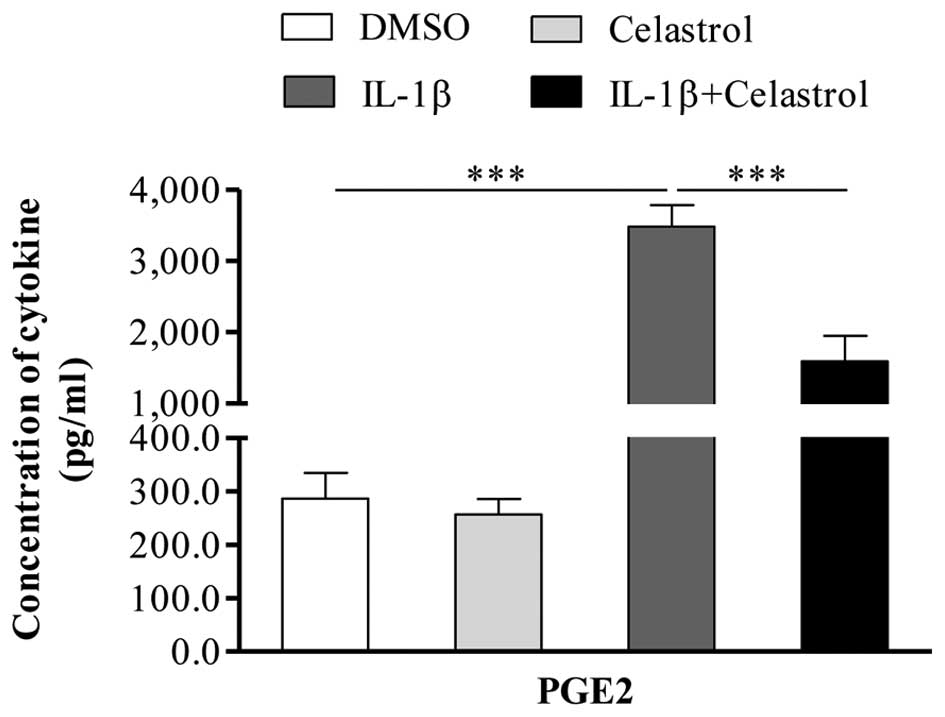
Effect of celastrol on IL-1β-induced PGE2
in GO orbital fibroblasts
PGE2 is important in modulating the inflammatory
process, and COX-2 is a key enzyme, which catalyzes the production
of PGE2. It has been suggested that the increase in PGE2 may be
attributed to the pathological inflammatory process of GO. As it
was found that the IL-1β-induced expression of COX-2 was depressed
by celastrol, the present study evaluated the effect of celastrol
on the IL-1β-induced expression of PGE2. Following treatment of the
GO orbital fibroblasts with 10 ng/ml IL-1β, with or without 1
µM celastrol for 24 h, the supernatants were analyzed using
ELISA to detect the production of PGE2. As shown in Fig. 4, IL-1β significantly induced the
production of PGE2 in the orbital fibroblasts, whereas co-treatment
with celastrol markedly attenuated the IL-1β-induced expression of
PGE2.
Effect of celastrol on the NF-κB
signaling pathway in GO orbital fibroblasts
The NF-κB signaling pathway is important in
regulating the production of several cytokines. In the cytoplasm,
NF-κB is arrested by IκB, and the activation of IKK phosphorylates
IκB, thereby releasing NF-κB, which translocates to the nucleus and
activates the transcription of response genes (31). It has been demonstrated that
celastrol is a potent inhibitor of NF-κB, therefore, the present
study examined whether celastrol exerts suppressive effects on
IL-1β-induced proinflammatory molecules through the inhibition of
NF-κB. As shown in Fig. 5A–C,
following treatment with IL-1β, the phosphorylation of IκBα was
significantly upregulated, whereas cotreatment with celastrol
significantly suppressed the IL-1β-induced phosphorylation of IκBα.
Pretreatment with the NF-κB inhibitor, BAY-11-7082, almost
completely inhibited the activation of NF-κB induced by IL-1β.
 | Figure 5Celastrol suppresses the
IL-1β-induced expression and activation of the NF-κB signaling
pathway in orbital fibroblasts. (A) Orbital fibroblasts from
patients with Graves' ophthalmopathy were stimulated with IL-1β (10
ng/ml), with or without 1 µM celastrol, for 24 h, Western
blot analysis was performed to determine the phosphorylation of
IκBa. (B) Luciferase activity in 293T cells treated with celastrol.
Cells were transfected with an NF-κB luciferase reporter vector.
After 24 h, the cells were stimulated with 10 ng/ml LPS with or
without 1 µM celastrol, for 24 h and then ysed for a
luciferase assay. (C) Orbital fibroblasts were pretreated with
BAY-11-7082 (2.5 µM) for 30 min, then stimulated with 10
ng/ml LPS, with or without 1 µM celastrol, for 24 h. Cells
were lysed for western blot analysis to detect the expression of
IκBa and phosphorylation of IκBa. Data is presented as the mean ±
standard error of the mean of three independent experiments.
**P<0.01; ***P<0.01. IL-1β,
interleukin-1β; LPS, lipopolysaccharide; NF-κB, nuclear factor-κB;
IκBα, inhibitor of κBα; p-, phosphorylated; DMSO, dimethyl
sulfoxide; BAY, BAY-11-7082. |
The effect of celastrol was also examined using an
NF-κB luciferase system in 293T cells. As shown in Fig. 5B, celastrol significantly inhibited
IL-1β-induced NF-κB activation, in a dose-dependent manner.
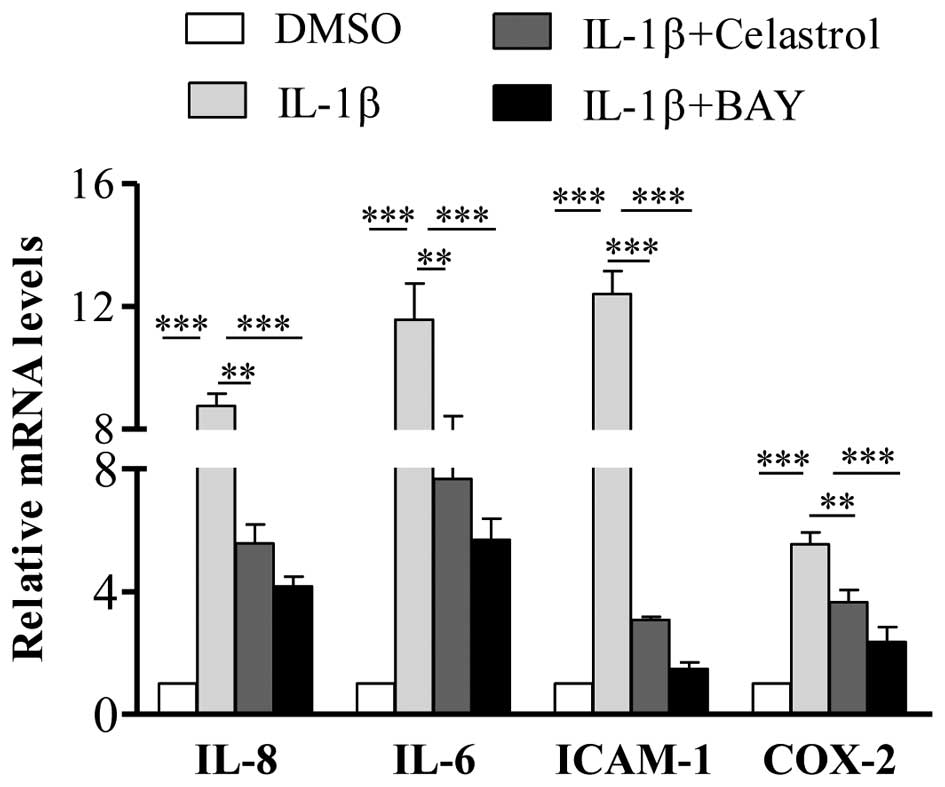
Celastrol suppresses the induction of
cytokines by IL-1β in orbital fibroblasts through inhibition of the
NF-κB signaling pathway
To further determine whether the IL-1β-induced
stimulation of proinflammatory gene expression was mediated by the
NF-κB-dependent pathway, the present study pretreated GO cells with
BAY-11-7082 (2.5 µM) for 30 min, following which IL-1β
and/or celastrol were added. Following incubation for 24 h, the
cells were harvested and subjected to RT-qPCR analysis. The results
showed that pre-incubation with BAY-11-7082 significantly decreased
the IL-1β-induced gene expression levels of IL-6, IL-8, ICAM-1 and
COX-2 (Fig. 6), which confirmed
activation of the NF-κB pathway as the mechanism underlying the
increased expression of these cytokines.
Discussion
Celastrol is generally used for the treatment of
inflammatory and autoimmune diseases, however, the role of
celastrol in the development of GO remains to be fully elucidated.
In the present study, it was found that treatment with celastrol
significantly attenuated inflammatory responses in IL-1β-induced
orbital fibroblasts from patients with GO through inhibiting the
activation of NF-κB. These results suggested that the use of
celastrol may offer potential in the management of GO.
GO is an autoimmune disease, which is characterized
by the infiltration of immune cells into the orbit and the
production of excess glycosaminoglycans and inflammatory cytokines,
which regulate the inflammatory response through recruiting and
activating inflammatory cells. It has been suggested that cytokines
are critical in the development of GO, as several cytokines have
been detected in orbital tissues from patients with GO, including
IL-1β, IL-6, IL-8, COX-2 and ICAM-1 (12,32).
It is well known that cytokines are produced predominantly by
immune cells, and several have suggested that orbital fibroblasts
are another important source of cytokines, which are critical in
initiating and maintaining inflammation (33), with accumulating evidence
suggesting that orbital fibroblasts are the autoimmune target and
effector cells in GO (34–36). IL-1β is an important member of the
IL-1 cytokine family, and mRNA expression levels of IL-1β have been
reported to be high in the orbital tissues of patients with GO
(37). IL-1β is involved in
mediating the inflammatory response, and it has been reported that
IL-1β induces several mediators that have been correlated with the
pathogenesis of GO, including IL-6 (38), IL-8 (39) and hyaluronic acid (40). In the present study, it was
demonstrated that celastrol significantly suppressed the production
of cytokines IL-6 and IL-8 in the orbital fibroblast induced by
IL-1β.
ICAM-1 was also induced by IL-1β in the orbital
fibroblasts, and the expression of ICAM-1 has been reported to be
involved in the migration of lymphocytes to inflammatory sites in
the orbit (41). The induction of
COX-2 is considered to be critical to the inflammatory response in
patients with GO. Orbital fibroblasts from the patients with GO
treated by IL-1β produced high levels of COX-2, and there is a
positive correlation between the expression of COX-2 and the
increasing severity of orbital disease (12). All these results suggested a
possible association between the expression levels of ICAM-1 and
COX2, and orbital inflammation in GO. Thus, the downregulation of
these cytokines may result in decreased recruitment of leukocyte
subsets into orbital fibroblasts. Pre-treatment of the orbital
fibroblasts with celastrol had a potent inhibitory effect on the
levels of IL-1β-induced ICAM-1 and COX-2 in the IL-1β-induced
orbital fibroblasts. Together with the data described above, the
results of the present study demonstrated that celastrol inhibited
the production of the IL-6, IL-8, ICAM-1 and COX2 cytokines in
orbital fibroblasts induced by IL-1β, thereby suppressing the
inflammatory response.
NF-κB is a central transcription factor, which is
well established as a regulator in mediating inflammatory and
innate immune responses. NF-κB may be activated by various factors,
including the IL-1 cytokine (42).
NF-κB is important in regulating cell proliferation and cell
survival. In the inactive state, NF-κB is located in the cytoplasm,
bound to the inhibitory protein, IκBα. Following stimulation, the
IKK complex is activated, which results in the phosphorylation and
subsequent degradation of IκBα, leading to the release of NF-κB and
its translocation to the nucleus, and activation of the
transcription of target genes (43). Previous experiments have confirmed
that the upregulation of COX-2 in GO is due to the activation of
NF-κB, and treatment with NF-κB inhibitor almost completely
suppresses IL-1β-induced COX-2 in orbital fibroblast (32). Therefore, the effective inhibition
of NF-κB may be one of the therapeutic targets in GO.
Celastrol is a pharmacologically active compound,
which possesses a broad rage of biological activities and is
generally used for the treatment of inflammatory and autoimmune
diseases. Although several studies have demonstrated that celastrol
offers therapeutic potential in a number of inflammatory-associated
diseases in vivo and in vitro (17,18,44),
its effects have not been investigated previously in GO. The
application of celastrol has been controversial due to its
toxicity. The present study showed that treatment with 1 µM
celastrol exerted no clear cytotoxic effects on the orbital
fibroblast, and did not induce a significant level of apoptosis.
This suggests that celastrol has realistic potential in clinical
application. Celastrol is considered an inhibitor of NF-κB, and
several studies have demonstrated potent inhibitory effects on
NF-κB in various types of cell (45,46).
Of note, in the present study, celastrol was found to significantly
suppress the production of cytokines induced by IL-1β in orbital
fibroblasts, and the levels of PGE2 in the IL-1β-induced orbital
fibroblasts was also inhibited by celastrol.
The results of the present study suggested that
celastrol attenuated the IL-1β-induced pro-inflammatory pathway in
orbital fibroblasts from patients with GO, which was associated
with the suppression of NF-κB. The present study was the first, to
the best of our knowledge, to evaluate the anti-inflammatory
effects of celastrol on orbital fibroblasts in patients with GO,
and the results suggested that celastrol may be efficient in the
treatment of GO, in terms of attenuating the inflammatory
process.
Acknowledgments
This study was supported by The National Natural
Science Foundation of China (grant nos. 81373617, 81072793 and
30772800) and the Longhua Medical Project (grant no. LYTD-10).
References
|
1
|
Prabhakar BS, Bahn RS and Smith TJ:
Current perspective on the pathogenesis of Graves' disease and
ophthalmopathy. Endocr Rev. 24:802–835. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bahn RS: Graves' ophthalmopathy. N Engl J
Med. 362:726–738. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Garrity JA and Bahn RS: Pathogenesis of
graves ophthalmopathy: Implications for prediction, prevention, and
treatment. Am J Ophthalmol. 142:147–153. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wiersinga WM and Bartalena L: Epidemiology
and prevention of Graves' ophthalmopathy. Thyroid. 12:855–860.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ajjan RA and Weetman AP: New understanding
of the role of cytokines in the pathogenesis of Graves'
ophthalmopathy. J Endocrinol Invest. 27:237–245. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kazim M, Goldberg RA and Smith TJ:
Insights into the pathogenesis of thyroid-associated orbitopathy:
Evolving rationale for therapy. Arch Ophthalmol. 120:380–386. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Smith TJ: Orbital fibroblasts exhibit a
novel pattern of responses to proinflammatory cytokines: Potential
basis for the pathogenesis of thyroid-associated ophthalmopathy.
Thyroid. 12:197–203. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee WM, Paik JS, Cho WK, Oh EH, Lee SB and
Yang SW: Rapamycin enhances TNF-α-induced secretion of IL-6 and
IL-8 through suppressing PDCD4 degradation in orbital fibroblasts.
Curr Eye Res. 38:699–706. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoon JS, Chae MK, Lee SY and Lee EJ:
Anti-inflammatory effect of quercetin in a whole orbital tissue
culture of Graves' orbitopathy. Br J Ophthalmol. 96:1117–1121.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Heufelder AE and Bahn RS: Elevated
expression in situ of selectin and immunoglobulin superfamily type
adhesion molecules in retroocular connective tissues from patients
with Graves' ophthalmopathy. Clin Exp Immunol. 91:381–389. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kahaly G, Hansen C, Felke B and Dienes HP:
Immunohistochemical staining of retrobulbar adipose tissue in
Graves' ophthalmopathy. Clin Immunol Immunopathol. 73:53–62. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Konuk EB, Konuk O, Misirlioglu M, Menevse
A and Unal M: Expression of cyclooxygenase-2 in orbital
fibroadipose connective tissues of Graves' ophthalmopathy patients.
Eur J Endocrinol. 155:681–685. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang HS, Cao HJ, Winn VD, Rezanka LJ,
Frobert Y, Evans CH, Sciaky D, Young DA and Smith TJ: Leukoregulin
induction of prostaglandin-endoperoxide H synthase-2 in human
orbital fibroblasts. An in vitro model for connective tissue
inflammation. J Biol Chem. 271:22718–22728. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bartalena L, Pinchera A and Marcocci C:
Management of Graves' ophthalmopathy: Reality and perspectives.
Endocr Rev. 21:168–199. 2000.PubMed/NCBI
|
|
15
|
Kannaiyan R, Shanmugam MK and Sethi G:
Molecular targets of celastrol derived from Thunder of God Vine:
Potential role in the treatment of inflammatory disorders and
cancer. Cancer Lett. 303:9–20. 2011. View Article : Google Scholar
|
|
16
|
Salminen A, Lehtonen M, Paimela T and
Kaarniranta K: Celastrol: Molecular targets of Thunder God Vine.
Biochem Biophys Res Commun. 394:439–442. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cascão R, Vidal B, Raquel H, Neves-Costa
A, Figueiredo N, Gupta V, Fonseca JE and Moita LF: Effective
treatment of rat adjuvant-induced arthritis by celastrol. Autoimmun
Rev. 11:856–862. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li H, Zhang YY, Tan HW, Jia YF and Li D:
Therapeutic effect of tripterine on adjuvant arthritis in rats. J
Ethnopharmacol. 118:479–484. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim DY, Park JW, Jeoung D and Ro JY:
Celastrol suppresses allergen-induced airway inflammation in a
mouse allergic asthma model. Eur J Pharmacol. 612:98–105. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li H, Zhang YY, Huang XY, Sun YN, Jia YF
and Li D: Beneficial effect of tripterine on systemic lupus
erythematosus induced by active chromatin in BALB/c mice. Eur J
Pharmacol. 512:231–237. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim DH, Shin EK, Kim YH, Lee BW, Jun JG,
Park JH and Kim JK: Suppression of inflammatory responses by
celastrol, a quinone methide triterpenoid isolated from Celastrus
regelii. Eur J Clin Invest. 39:819–827. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sen T, Dutta A and Chatterjee A:
Epigallocatechin-3-gallate (EGCG) downregulates gelatinase-B
(MMP-9) by involvement of FAK/ERK/NFkappaB and AP-1 in the human
breast cancer cell line MDA-MB-231. Anticancer Drugs. 21:632–644.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ghosh S, May MJ and Kopp EB: NF-kappa B
and Rel proteins: evolutionarily conserved mediators of immune
responses. Annu Rev Immunol. 16:225–260. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee JH, Koo TH, Yoon H, Jung HS, Jin HZ,
Lee K, Hong YS and Lee JJ: Inhibition of NF-kappa B activation
through targeting I kappa B kinase by celastrol, a quinone methide
triterpenoid. Biochem Pharmacol. 72:1311–1321. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sethi G, Ahn KS, Pandey MK and Aggarwal
BB: Celastrol, a novel triterpene, potentiates TNF-induced
apoptosis and suppresses invasion of tumor cells by inhibiting
NF-kappaB-regulated gene products and TAK1-mediated NF-kappaB
activation. Blood. 109:2727–2735. 2007.
|
|
26
|
Yiu WH, Wong DW, Chan LY, Leung JC, Chan
KW, Lan HY, Lai KN and Tang SC: Tissue kallikrein mediates
pro-inflammatory pathways and activation of protease-activated
receptor-4 in proximal tubular epithelial cells. PLoS One.
9:e888942014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liang C, Du W, Dong Q, Liu X, Li W, Wang Y
and Gao G: Expression levels and genetic polymorphisms of
interleukin-2 and interleukin-10 as biomarkers of Graves' disease.
Exp Ther Med. 9:925–930. 2015.PubMed/NCBI
|
|
28
|
Zhao LQ, Wei RL, Cheng JW, Cai JP and Li
Y: The expression of intercellular adhesion molecule-1 induced by
CD40-CD40 L ligand signaling in orbital fibroblasts in patients
with Graves' ophthalmopathy. Invest Ophthalmol Vis Sci.
51:4652–4660. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Choi YH, Back KO, Kim HJ, Lee SY and Kook
KH: Pirfenidone attenuates IL-1β-induced COX-2 and PGE2 production
in orbital fibroblasts through suppression of NF-κB activity. Exp
Eye Res. 113:1–8. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
31
|
Chen LF and Greene WC: Shaping the nuclear
action of NF-kappaB. Nat Rev Mol Cell Biol. 5:392–401. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yoon JS, Lee HJ, Choi SH, Chang EJ, Lee SY
and Lee EJ: Quercetin inhibits IL-1β-induced inflammation,
hyaluronan production and adipogenesis in orbital fibroblasts from
Graves' orbitopathy. PLoS One. 6:e262612011. View Article : Google Scholar
|
|
33
|
Smith TJ: Unique properties of orbital
connective tissue underlie its involvement in Graves' disease.
Minerva Endocrinol. 28:213–222. 2003.PubMed/NCBI
|
|
34
|
Smith TJ: Novel aspects of orbital
fibroblast pathology. J Endocrinol Invest. 27:246–253. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Smith RS, Smith TJ, Blieden TM and Phipps
RP: Fibroblasts as sentinel cells. Synthesis of chemokines and
regulation of inflammation. Am J Pathol. 151:317–322.
1997.PubMed/NCBI
|
|
36
|
Cao HJ, Wang HS, Zhang Y, Lin HY, Phipps
RP and Smith TJ: Activation of human orbital fibroblasts through
CD40 engagement results in a dramatic induction of hyaluronan
synthesis and prostaglandin endoperoxide H synthase-2 expression.
Insights into potential pathogenic mechanisms of thyroid-associated
ophthalmopathy. J Biol Chem. 273:29615–29625. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wakelkamp IM, Bakker O, Baldeschi L,
Wiersinga WM and Prummel MF: TSH-R expression and cytokine profile
in orbital tissue of active vs. Inactive Graves' ophthalmopathy
patients. Clin Endocrinol (Oxf). 58:280–287. 2003. View Article : Google Scholar
|
|
38
|
Chen B, Tsui S and Smith TJ: IL-1 beta
induces IL-6 expression in human orbital fibroblasts:
Identification of an anatomic-site specific phenotypic attribute
relevant to thyroid-associated ophthalmopathy. J Immunol.
175:1310–1319. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hwang CJ, Afifiyan N, Sand D, Naik V, Said
J, Pollock SJ, Chen B, Phipps RP, Goldberg RA, Smith TJ and Douglas
RS: Orbital fibroblasts from patients with thyroid-associated
ophthalmopathy overexpress CD40: CD154 hyperinduces IL-6, IL-8 and
MCP-1. Invest Ophthalmol Vis Sci. 50:2262–2268. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kaback LA and Smith TJ: Expression of
hyaluronan synthase messenger ribonucleic acids and their induction
by interleukin-1beta in human orbital fibroblasts: Potential
insight into the molecular pathogenesis of thyroid-associated
ophthalmopathy. J Clin Endocrinol Metab. 84:4079–4084.
1999.PubMed/NCBI
|
|
41
|
Sikorski EE, Hallmann R, Berg EL and
Butcher EC: The Peyer's patch high endothelial receptor for
lymphocytes, the mucosal vascular addressin, is induced on a murine
endothelial cell line by tumor necrosis factor-alpha and IL-1. J
Immunol. 151:5239–5250. 1993.PubMed/NCBI
|
|
42
|
Lie PP, Cheng CY and Mruk DD: The biology
of interleukin-1: Emerging concepts in the regulation of the actin
cytoskeleton and cell junction dynamics. Cell Mol Life Sci.
69:487–500. 2012. View Article : Google Scholar :
|
|
43
|
Tak PP and Firestein GS: NF-kappaB: A key
role in inflammatory diseases. J Clin Invest. 107:7–11. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kiaei M, Kipiani K, Petri S, Chen J,
Calingasan NY and Beal MF: Celastrol blocks neuronal cell death and
extends life in transgenic mouse model of amyotrophic lateral
sclerosis. Neuro-degener Dis. 2:246–254. 2005. View Article : Google Scholar
|
|
45
|
He D, Xu Q, Yan M, Zhang P, Zhou X, Zhang
Z, Duan W, Zhong L, Ye D and Chen W: The NF-kappa B inhibitor,
celastrol, could enhance the anti-cancer effect of gambogic acid on
oral squamous cell carcinoma. BMC Cancer. 9:3432009. View Article : Google Scholar :
|
|
46
|
Zhou LL, Lin ZX, Fung KP, Cheng CH, Che
CT, Zhao M, Wu SH and Zuo Z: Celastrol-induced apoptosis in human
HaCaT keratinocytes involves the inhibition of NF-κB activity. Eur
J Pharmacol. 670:399–408. 2011. View Article : Google Scholar : PubMed/NCBI
|