Introduction
The increasing prevalence of obesity has become a
major worldwide issue. Obesity is associated with high rates of
type 2 diabetes mellitus, cardiovascular disease and infertility
(1,2). The Nurses' Health study reported that
a high body mass index (BMI) value is associated with a high risk
of infertility in women (3).
Furthermore, obesity is an important factor involved in polycystic
ovary syndrome, a condition associated with anovulatory infertility
(1,4–6).
However, the molecular mechanism of the effect of obesity on
follicular development remains unclear.
In the mammalian ovary, follicular development is
regulated by the hypothalamus-pituitary-ovary axis and various
other signaling pathways (7). As a
vital transcription factor, forkhead box O3a (FoxO3a) negatively
regulates primordial follicle activation and early follicular
development (8,9). Previous investigations have indicated
that FoxO3a is highly expressed and localized to the nucleus during
the dormancy of primordial follicles, then is phosphorylated and
exported to the cytoplasm during follicular activation in mice
(10). However, the activity of
FoxO3a is suppressed in later-growing follicles, indicating that
the downregulation of FoxO3a in oocytes may be a prerequisite for
the initiation of oocyte growth during follicular activation
(11,12).
It is well-established that the bidirectional
communication between oocytes and granulose cells (GCs) is also
important for normal follicular development. Bone morphogenetic
protein 15 (BMP15) and the transforming growth factor-β family
members, are fundamental for the paracrine signaling between
oocytes and GCs that controls follicular development and cumulus
expansion (13–18). GCs are one of the target cells of
BMP15 (19,20). BMPs bind to their cell membrane
receptor and exert their effects through receptor-mediated
activation of Smad transcription factors (21). Previous studies have demonstrated
that BMP15 regulates Smad1/5/8 activity and regulates ovarian
follicular development (22–24).
Additionally, Liu et al (11,25)
reported that constitutively active FoxO3a expressed in oocytes
stimulated downregulation of BMP15 expression and suppressed Smad
pathway activation. Furthermore, the BMP15 gene promoter contains
FoxO3a binding sites (11,25). These previous studies indicated
that the regulation of FoxO3a during follicular development may be
mediated via the BMP15/Smad1/5/8 signaling pathway.
Previous studies have systematically reported that
maintaining optimal activity of Akt serine/threonine kinase 1 (Akt)
signaling is essential for normal ovarian function, which regulates
the balance between survival and activation of primordial
follicles, and also is important for proliferation and
differentiation of GCs (26–28).
FoxO3a is also a substrate of Akt, thus, the present study also
examined the expression of Akt ovaries. The present study was
designed to verify whether Akt/FoxO3a/Smad1/5/8 signaling is the
intracellular mechanism underlying the effect of high-fat
diet-induced obesity on the follicular development, to demonstrate
the importance of understanding the effect of abnormal metabolite
process in ovarian function and potentially elucidating novel
interventions for obese pregnant mothers.
Materials and methods
Animals
Four-week old female C57BL/6J mice (weighing ≥35 g)
were purchased from Wushi Experimental Animal Supply (Fuzhou,
China). The animals were maintained under a 14-h light/10-h dark
condition, at 25°C. The experimental protocol was approved in
accordance with the Guide for the Care and Use of Laboratory
Animals prepared by the Institutional Animal Care and Use Committee
(Fujian Normal University, Fuzhou, China).
Experimental design
The mice (n=60) were divided into two groups
(n=30/group) and fed either a high-fat diet (HFD; Research Diets,
Inc., New Brunswick, NJ, USA) or a normal control diet (Research
Diets, Inc.) for 15 or 20 weeks. HFD contained 20% protein, 20%
carbohydrate and 60% fat. The normal control (NC) diet contained
20% protein, 70% carbohydrate and 10% fat. Body weight was recorded
weekly. Following feeding with the different diets for 15 and 20
weeks, the BMI was determined. BMI was calculated using the
following formula: body weight/height2,
(kg/m2). Following feeding with the different diets for
15 or 20 weeks, the mice were sacrificed with 20% CO2,
followed by cervical dislocation, and the ovaries were dissected
from the mice. One ovary was examined morphology under a Leica
MZ16F stereomicroscope (Leica Microsystems, Inc., Buffalo Grove,
IL, USA) or via hematoxylin and eosin (H&E) staining, and the
other ovary was snap-frozen and used for immunoblotting.
H&E staining
Ovaries were collected from mice fed with the
different diets for 15 or 20 weeks, and then fixed with 4%
paraformaldehyde in phosphate-buffered saline. Ovaries were
dehydrated in alcohol and embedded with paraffin. Sections (10
µm) were stained with H&E and observed under a light
microscope.
Immunoblotting
Immunoblotting was performed as previously described
(2,29–34).
Briefly, ovaries from different experimental groups were sonicated
in lysis buffer containing a protease inhibitor cocktail
(Sigma-Aldrich, St. Louis, MO, USA). The proteins were quantified
using DC protein assay kit (Bio-Rad Laboratories, Inc., Hercules,
CA, USA). Equal amounts of protein (30–50 µg) and the
Precision Plus Protein Standards (Bio-Rad Laboratories, Inc.) were
resolved by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred onto polyvinylidene difluoride
membranes. Membranes were blocked in 5% nonfat milk for 45 min at
25°C and then incubated for 18 h at 4°C with the primary antibodies
at 1:1,000 in 5% non-fat milk, including Akt (cat. no. 8272),
phosphorylated (p)-Akt (cat. no. 4058), FoxO3a (cat. no. 9467),
p-FoxO3a (cat. no. 9464s), Smad1/5/8 (cat. no. 9517P) and
p-Smad1/5/8 (cat. no. 9511) (all from Cell Signaling Technology,
Inc., Danvers, MA, USA). To determine whether equivalent amounts of
protein were loaded among all samples, membranes were stripped and
incubated with a mouse antibody against β-actin (Abcam, Cambridge,
UK) to generate a signal for use as a loading control. Following
primary antibody incubation, the membranes were incubated with goat
anti-rabbit or goat anti-mouse secondary antibodies (1:10,000;
Jackson ImmunoResearch Laboratories, Inc., West Grove, PA, USA) at
25°C for 4 h. Using the Super Signal West Femto Maximum Sensitivity
Substrate kit (Thermo Fisher Scientific, Inc., Waltham, MA, USA),
chemiluminescence emitted from the bands was directly captured
using a UVP Bioimage EC3 system (UVP, Inc., Upland, CA, USA).
Densitometric analysis of chemiluminescence signals was performed
using VisionWorks LS software (version, EC3 600; UVP, Inc.). The
quantitative analysis of protein levels was calculated as the
densitometric value of phosphorylated protein level/the
densitometric value of total protein. All experiments were repeated
in triplicate with the use of independently prepared tissue
lysates.
Statistical analysis
Data are presented as the mean ± standard error.
Experiments were repeated at least three times, and ovary samples
for each replicate were from different female mice. Statistical
differences were determined by one-way analysis of variance with
Tukey's test used to determine the significance of the results
using Sigma Stat 3.5 software (Systat Software, Inc., San Jose, CA,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Animal model of HFD-induced obesity
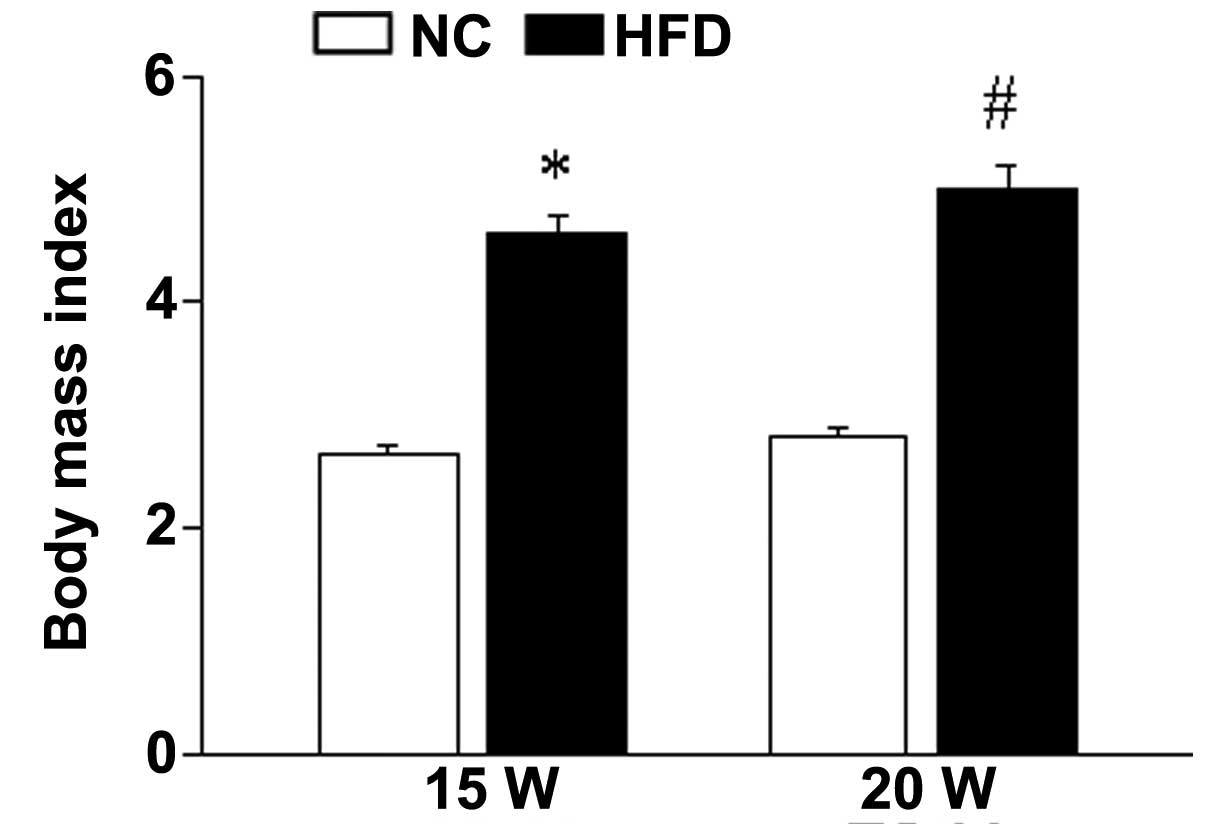
The mice received HFD or NC diet for 15 or 20 weeks.
After 15/20-week feeding, the BMI of the mice in the HFD group was
significantly increased compared with the NC group (15 weeks,
P<0.001; 20 weeks, P<0.001), indicating that the obese mouse
model has been successfully established (Fig. 1).
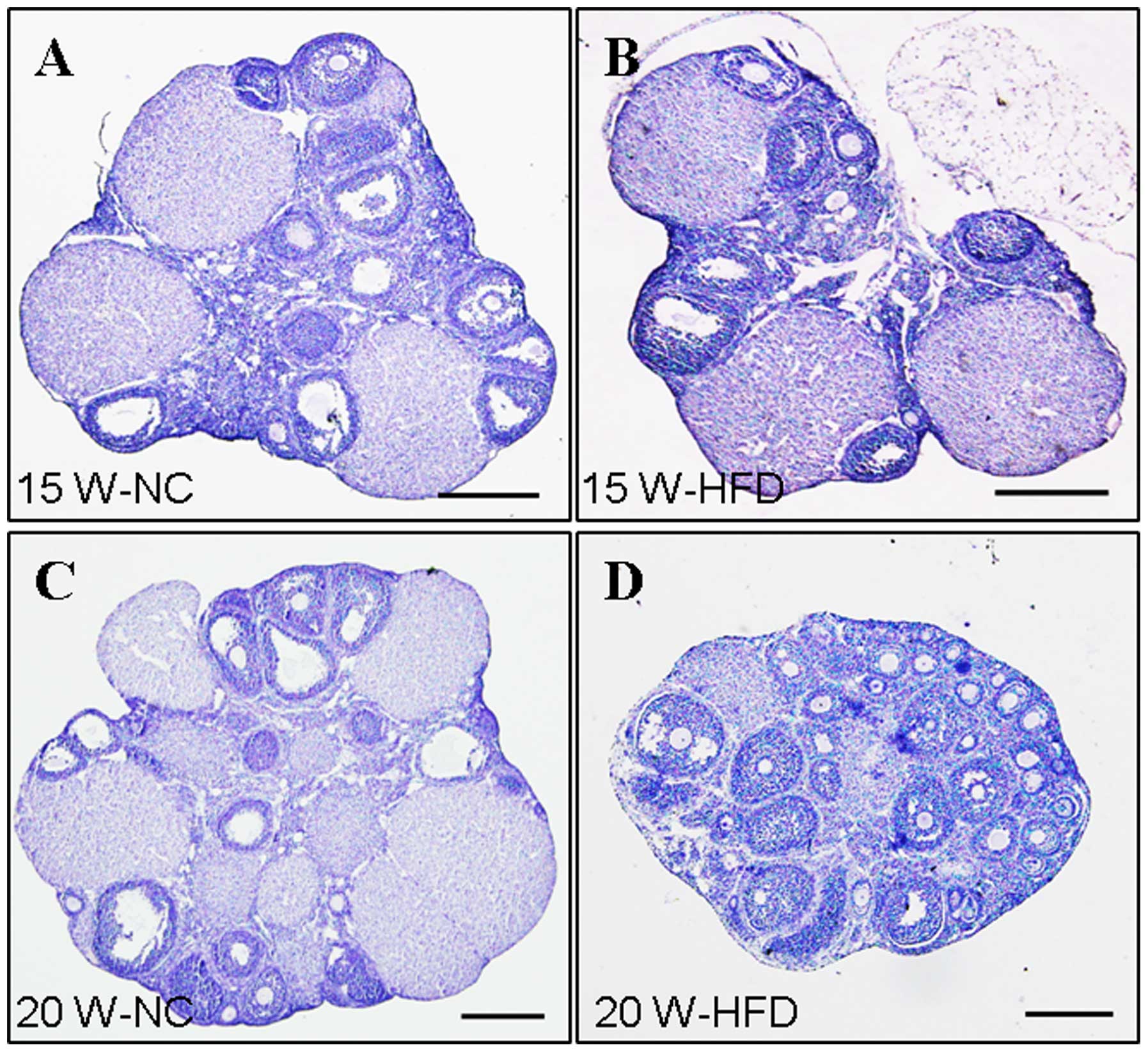
HFD-induced obesity causes abnormal
ovarian morphology
HFD-induced obesity had a negative effect on ovarian
function. The ovarian morphology in the HFD group (15 weeks) did
not exhibit obviously abnormal ovarian morphology compared with the
NC group (Fig. 2A and B). However,
when administered with HFD for 20 weeks, an abnormal morphological
effect of obesity on ovarian development was observed (Fig. 2C and D). H&E staining was
performed to observe the histology of the ovaries. The majority of
the follicles in the ovaries from the NC group had developed to the
later follicles stages or the corpora lutea stage (Fig. 3). By contrast, the mouse ovaries in
the HFD group exhibited hyaline appearance, were avascular and had
large numbers of cystic follicles stored under the tunica albuginea
(Fig. 2D). Additionally, the
majority of follicles in the ovaries from the HFD group (20 weeks)
were retarded at an early stage or were atretic (Fig. 3D). These findings suggested that
HFD-induced obesity exerts a detrimental effect on follicular
development.
HFD-induced obesity suppresses the
phosphorylation level of FoxO3a in the ovary
Compared with the follicles in NC group, the
majority of ovarian follicles were retarded at the early follicle
stage in the HFD group (Fig. 3),
indicating that the regulatory mechanisms underlying follicular
development are disrupted. FoxO3a is a major regulator of
primordial follicle activation and early follicular development. To
determine whether FoxO3a is involved in the effect of HFD-induced
obesity on follicular development, the expression level of p-FoxO3a
in the ovaries was examined. The protein level of p-FoxO3a was
significantly reduced in the ovaries from mice in the HFD group
compared with the NC group at 15 weeks (P<0.001) and 20 weeks
(P<0.001) (Fig. 4).
Additionally, the degree of reduction of p-FoxO3a in the ovaries
from mice fed with HFD for 20 weeks was greater than in the ovaries
from mice fed with HFD for 15 weeks; however, no difference in the
total protein levels were observed compared with β-actin (15 weeks,
P=0.002; 20 weeks, P=0.629; Fig.
4). These data suggest that HFD-induced obesity suppresses
FoxO3a phosphorylation in a time-dependent manner, leading to
increased FoxO3a activity and retarded follicular development.
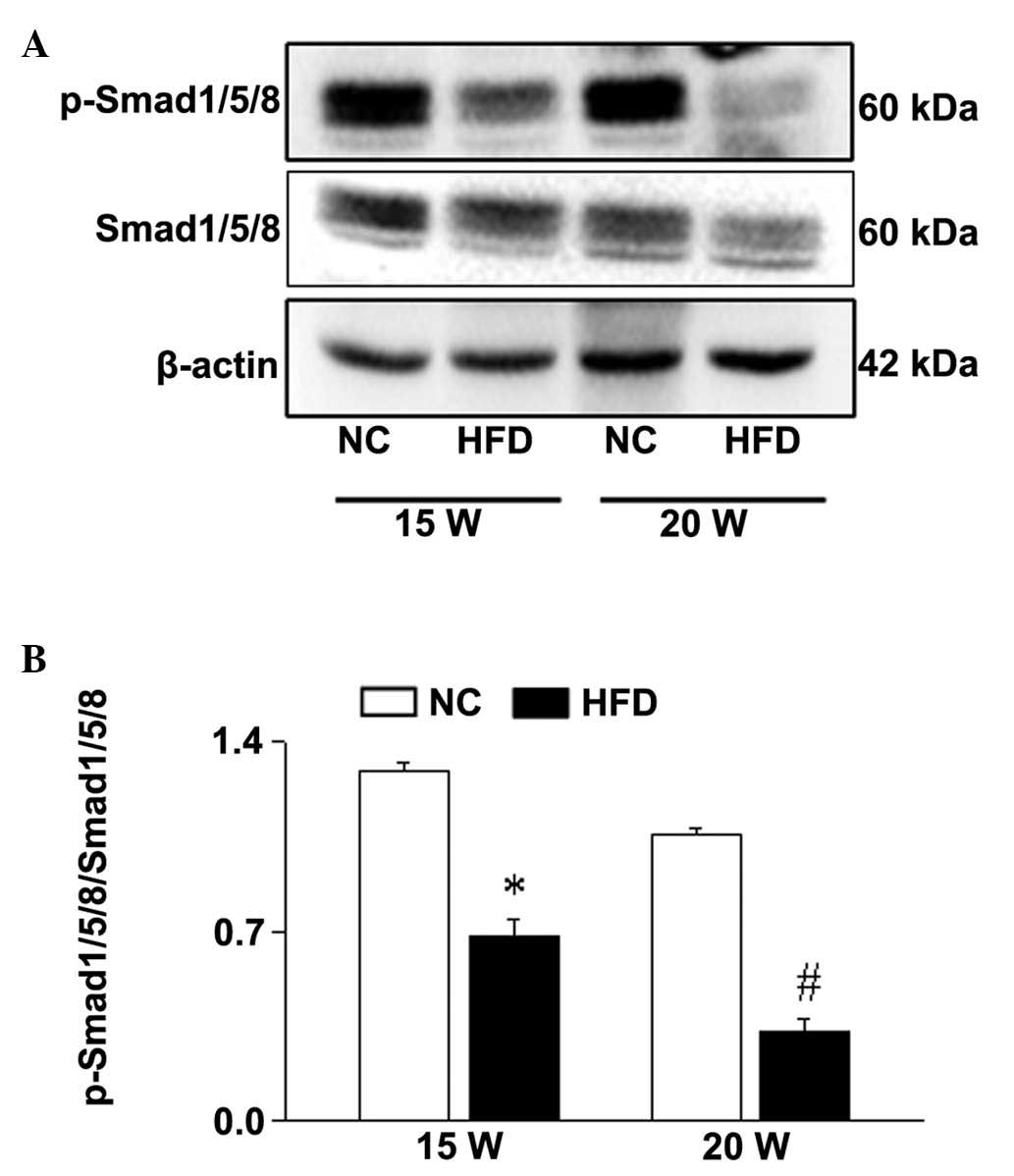
HFD-induced obesity represses Smad
signaling pathway activation in the ovary
BMP15 is specifically expressed in oocytes and is
important for the regulation of cell growth, development and
apoptosis (19,20). FoxO3a can bind the BMP15 gene
promoter to regulate its expression. The expression of p-Smad1/5/8,
the downstream effector of BMP15, was significantly reduced in the
ovaries from HFD mice compared with NC mice in a time-dependent
manner (15 weeks, P<0.001; 20 weeks; P<0.001; Fig. 5). Thus, these data indicate that
FoxO3a/Smad1/5/8 signaling may be involved in the effect of
HFD-induced obesity on follicular development.
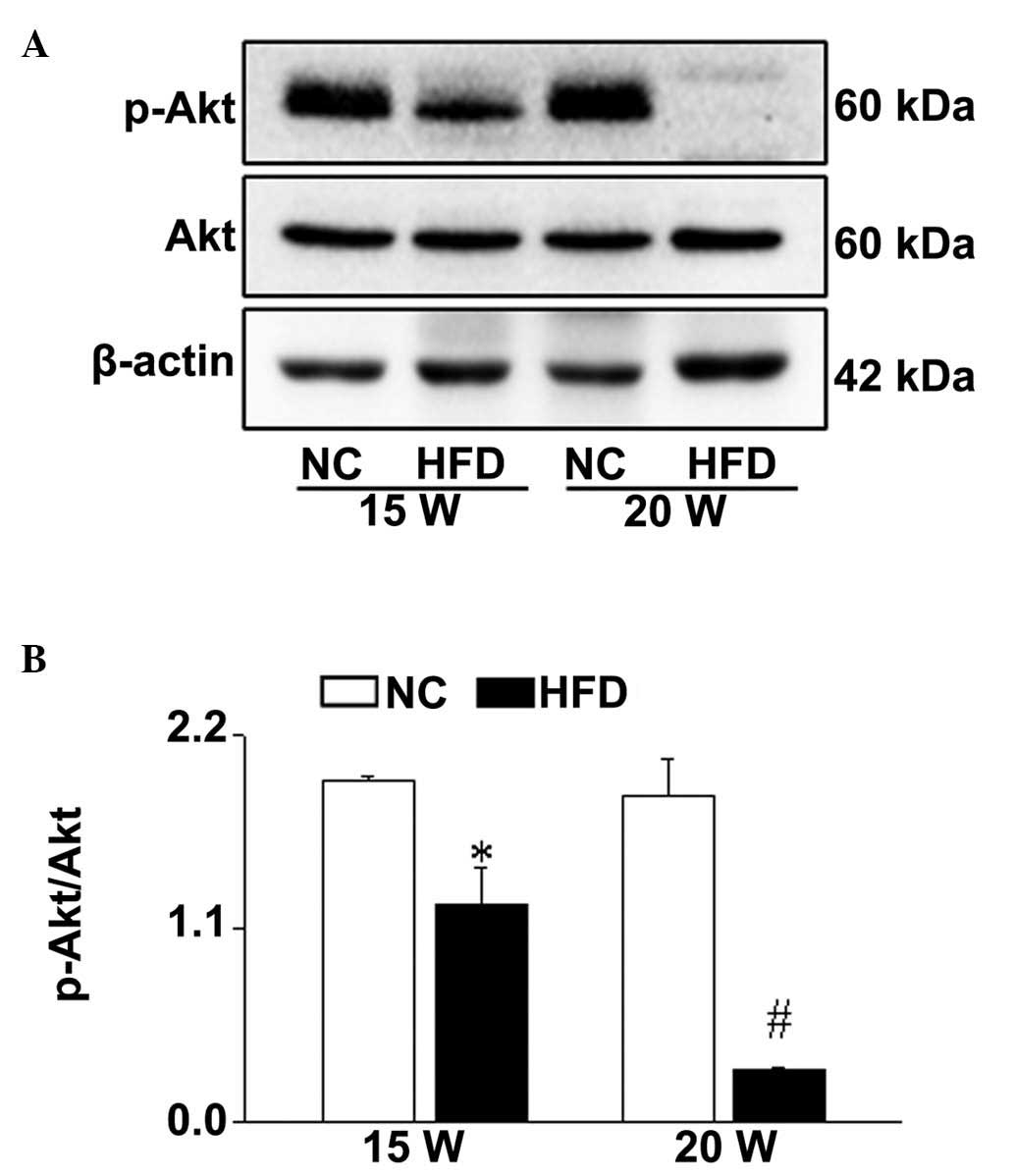
HFD-induced obesity downregulates Akt
signaling in the ovary
Akt, an upstream regulator of FoxO3a, is essential
for the adverse metabolic effects of insulin, which is the major
regulatory mechanism involved in follicular development (25). Thus, the effect of HFD-induced
obesity on Akt signaling during the follicular development was
investigated. The level of p-Akt was significantly reduced in the
ovaries from HFD group mice compared with the NC group (15 weeks,
P=0.038; 20 weeks, P<0.001; Fig.
6). In accordance with the expression level of p-FoxO3a, the
reduction of p-Akt expression in the ovaries from the mice fed with
HFD for 20 weeks was increased compared with the mice fed with HFD
for 15 weeks (Fig. 6). These data
suggested that HFD-induced obesity may cause abnormal follicular
development via impairing Akt/FoxO3a signaling in a time-dependent
manner during follicular development.
Discussion
The results of the present study demonstrated that
HFD-induced obesity observably inhibited follicular development in
the mouse ovary in a time-dependent manner. Furthermore, HFD
increased FoxO3a activity in the mouse ovaries, as indicated by
decreased p-FoxO3a levels. These results together indicated that
FoxO3a may be an important regulatory factor involved in the
ovarian follicular development of HFD-fed mice.
Obesity is currently the major factor that causes
type 2 diabetes, cardiac malformation and infertility (1). With the increasing epidemic of
obesity, a useful mouse model of obesity is essential for studying
the mechanisms that underlie the negative effects of obesity
negatively on the function of the ovary. The current study
established an obese mouse model using C57BL/6J female mice fed
with HFD for 15 or 20 weeks and focused on the molecular mechanism
underlying the effect of obesity on follicular development. The
present results demonstrated that HFD induced-obesity has
deleterious effect on the ovarian development in a time-dependent
manner. The follicular development in mice fed with HFD for 20
weeks was retarded at the early stage or were completely atretic.
Thus, it is important to investigate the mechanisms underlying
these processes.
In the mammalian ovary, follicular development is
regulated by the hypothalamus-pituitary-ovary axis, and various
signaling pathways (7). Ovarian
follicles develop from primordial follicles to primary follicles,
secondary follicles and antral follicles, which become a mature
follicle with a single oocyte enclosed by several layers of somatic
granulose cells (10). A
substantial process is involved in the regulation of follicular
development. In the ovary, primordial follicles are the source of
all developing follicles, which have three potential developmental
fates: i) To remain quiescent; ii) to be activated into the growing
follicle pool; or iii) to undergo death directly from the dormant
state (35–37). Consequently, the balance among
these three fates of the primordial follicle determines the length
of female reproductive life (27,37,38).
Additionally, the bidirectional communication between oocytes and
GCs is also important for normal follicular development.
As a vital transcription factor, FoxO3a negatively
regulates primordial follicle activation and early follicular
development (8,9). It has been previously demonstrated
that phosphorylation by phosphoinositide 3 kinase (PI3K)/Akt
controls a shuttling system that modulates FoxO3a nucleocytoplasmic
translocation and, thus, its activity. Under normal conditions,
FoxO3a is located in the nucleus and activates the expression of
genes associated with cell death/apoptosis (39). Following stimulation, Akt
phosphorylates FoxO3a, leading to the association of FoxO3a with
14-3-3 proteins, retention of FoxO3a in the cytoplasm and
inhibition of FoxO3a-dependent transcriptional activity (25,39).
Regarding follicular development, FoxO3a may activate the
expression of certain genes that maintain follicle quiescence. A
number of elegant genetic mouse experiments have provided insight
into the role of FoxO3a in oocyte quiescence. FoxO3a−/−
mice exhibited global activation of primordial follicles and
age-dependent infertility (8,40).
Oocyte-specific ablation of phosphatase and tensin homolog (a
negative regulator of PI3Ks) led to PI3K-induced Akt activation,
and thus, phosphorylated FoxO3a, suppressing its activity, and
consequently triggering a phenotype in oocytes equivalent to that
in mice lacking FoxO3a (41,42).
The reduced phosphorylation of FoxO3a observed in the present study
would maintain FoxO3a localization in the nuclei, which prevents
the activation of follicle development.
Previous studies have also demonstrated that
constitutively active FoxO3a suppressed Smad pathway activation via
downregulation of BMP15 expression in the ovaries from ZP3-FoxO3a
Tg mice (11). BMP15 is a growth
factor specifically secreted from oocytes, which is important for
promoting follicular development (13–18).
GC is one of the target cells of BMP15 (19,20).
Otsuka et al (43)
demonstrated that BMP15 controls the expression of plasma protein-A
in granulose cells coordinating with follicle stimulating hormone,
guaranteeing the development of the advantage follicle.
Additionally, BMP15 and growth differentiation factor-9 are
required for cholesterol synthesis and metabolism in cumulus cells,
adversely affecting the development of cumulus oocyte complexes
(22). These findings indicated
that FoxO3a/BMP15 signaling may be an important mechanism that
regulates follicular development. Notably, previous studies have
demonstrated that BMP15 regulates Smad1/5/8 activity and its
regulatory effect on ovarian follicular development (22–24).
Thus, the present study investigated Smad1/5/8 activity and
demonstrated that HFD significantly reduced the level of
p-Smad1/5/8. These studies suggest that HFD-induced obesity may
impair FoxO3a/Smad1/5/8 signaling and lead to abnormal follicular
development. The present data and previous results reinforce the
importance of bidirectional communication between oocytes and
somatic cells during oocyte and follicle development, and also
reveal that the signaling pathway and hypothalamus-pituitary-ovary
axis synergistically regulate follicular development.
PI3K/Akt signaling is one of the classic signaling
pathways in mammalian cells. Following stimulation, PI3K is
activated and then phosphorylates the third position hydroxyl group
of the inositol ring of phosphatidylinositol, resulting in the
production of second messengers of inositol lipid substances,
including PIP2 and PIP3 (44,45).
Akt, also known as protein kinase B, is a direct downstream target
of PI3K (46,47). Upon binding to the pleckstrin
homology domains in the N-terminus in the cell membrane with PIP3,
Akt is phosphorylated and activated by the catalysis of
3-phosphoinositide-dependent kinases (48). Activation of Akt, in turn,
activates downstream signaling molecules, including FoxO family
transcription factors, glycogen synthase kinase 3, Bcl-2 family
proteins and mammalian target rapamycin, to regulate physiological
processes, including cell proliferation, differentiation and
apoptosis (10).
Additionally, studies using a transgenic mouse model
with constitutively active FoxO3a expressed in oocytes demonstrated
that over-expression of FoxO3a impaired oocyte growth and
folliculogenesis, leading to female infertility (11). John et al (12) reported that as the downstream of
PI3K/Akt, hyperphosphorylation of FoxO3a induced over-activation of
primordial follicules (8,26,28).
As previous studies have systematically reported that maintaining
optimal activities of PI3K/Akt signaling pathway is essential for
normal ovarian function, which not only regulates the balance
between survival and activation of primordial follicles, but also
is vital for the proliferation and differentiation of GCs (26–28).
These studies suggested that regulation of the optimal expression
level of FoxO3a is important spatially and temporally during
follicular development. The data from the present study
demonstrated that HFD-induced obesity impairs Akt signaling and
induces over-activation of FoxO3a in a time-dependent manner,
implying that Akt/FoxO3a signaling is a critical signaling pathway
underlying HFD-induced obesity resulting in abnormal follicular
development.
In summary, the current study revealed the molecular
regulatory mechanisms underlying the effect of HFD-induced obesity
on follicular development, highlighting the importance of
Akt/FoxO3a/Smad1/5/8 signaling during regulation of follicular
development. The results from this study may provide a theoretical
basis for clinical treatment of infertility caused by obesity. As
the regulation of biological processes is complicated, further
investigation of the effect of HFD-induced obesity on the
regulation of hormones and inflammatory factors during ovarian
follicular development is required.
Acknowledgments
This work was supported by the National Natural
Science Foundation of China (grant nos. 31101032 and 31271255),
Program for New Century Excellent Talents in University of Ministry
of Education of China (grant no. NCET-120614), Doctoral Foundation
of the Ministry of Education in China (grant no. 20113503120002)
and Fujian Provincial Science and Technology Projects of the
Department of Education (grant no. JB14041).
References
|
1
|
Akamine EH, Marçal AC, Camporez JP,
Hoshida MS, Caperuto LC, Bevilacqua E and Carvalho CR: Obesity
induced by high-fat diet promotes insulin resistance in the ovary.
J Endocrinol. 206:65–74. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wu Y, Wang F, Fu M, Wang C, Quon MJ and
Yang P: Cellular stress, excessive apoptosis and the effect of
metformin in a mouse model of type 2 diabetic embryopathy.
Diabetes. 64:2526–2536. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wamae CN: Advances in the diagnosis of
human lymphatic filariases: A review. East Afr Med J. 71:171–182.
1994.PubMed/NCBI
|
|
4
|
Metwally M, Li TC and Ledger WL: The
impact of obesity on female reproductive function. Obes Rev.
8:515–523. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Legro RS: Evaluation and Treatment of
Polycystic Ovary Syndrome. Practice. 17:82–85. 2000.
|
|
6
|
Pasquali R and Casimirri F: The impact of
obesity on hyperandrogenism and polycystic ovary syndrome in
premenopausal women. Clin Endocrinol (Oxf). 39:1–16. 1993.
View Article : Google Scholar
|
|
7
|
Richards JS and Pangas SA: The ovary:
Basic biology and clinical implications. Journal Clin Invest.
120:963–972. 2010. View
Article : Google Scholar
|
|
8
|
Castrillon DH, Miao L, Kollipara R, Horner
JW and DePinho RA: Suppression of ovarian follicle activation in
mice by the transcription factor Foxo3a. Science. 301:215–218.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nteeba J, Ross JW, Perfield JW II and
Keating AF: High fat diet induced obesity alters ovarian
phosphatidylinositol-3 kinase signaling gene expression. Reprod
Toxicol. 42:68–77. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Uhlenhaut NH and Treier M: Forkhead
transcription factors in ovarian function. Reproduction.
142:489–495. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu L, Rajareddy S, Reddy P, Du C,
Jagarlamudi K, Shen Y, Gunnarsson D, Selstam G, Boman K and Liu K:
Infertility caused by retardation of follicular development in mice
with oocyte-specific expression of Foxo3a. Development.
134:199–209. 2007. View Article : Google Scholar
|
|
12
|
John GB, Shirley LJ, Gallardo TD and
Castrillon DH: Specificity of the requirement for Foxo3 in
primordial follicle activation. Reproduction. 133:855–863. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dong J, Albertini DF, Nishimori K, Kumar
TR, Lu N and Matzuk MM: Growth differentiation factor-9 is required
during early ovarian folliculogenesis. Nature. 383:531–535. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Elvin JA, Clark AT, Wang P, Wolfman NM and
Matzuk MM: Paracrine actions of growth differentiation factor-9 in
the mammalian ovary. Mol Endocrinol. 13:1035–1048. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Elvin JA, Yan C, Wang P, Nishimori K and
Matzuk MM: Molecular characterization of the follicle defects in
the growth differentiation factor 9-deficient ovary. Mol
Endocrinol. 13:1018–1034. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Vitt UA, McGee EA, Hayashi M and Hsueh AJ:
In vivo treatment with GDF-9 stimulates primordial and primary
follicle progression and theca cell marker CYP17 in ovaries of
immature rats. Endocrinology. 141:3814–3820. 2000.PubMed/NCBI
|
|
17
|
Vitt UA, Hayashi M, Klein C and Hsueh AJ:
Growth differentiation factor-9 stimulates proliferation but
suppresses the follicle-stimulating hormone-induced differentiation
of cultured granulosa cells from small antral and preovulatory rat
follicles. Biol Reprod. 62:370–377. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sánchez F and Smitz J: Molecular control
of oogenesis. Biochim Biophys Acta. 1822:1896–1912. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Paulini F and Melo EO: The role of
oocyte-secreted factors GDF9 and BMP15 in follicular development
and oogenesis. Reprod Domest Anim. 46:354–361. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gilchrist RB, Lane M and Thompson JG:
Oocyte-secreted factors: Regulators of cumulus cell function and
oocyte quality. Hum Reprod Update. 14:159–177. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
McCulley DJ, Kang JO, Martin JF and Black
BL: BMP4 is required in the anterior heart field and its
derivatives for endocardial cushion remodeling, outflow tract
septation, and semilunar valve development. Dev Dyn. 237:3200–3209.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Su YQ, Sugiura K, Wigglesworth K, O'Brien
MJ, Affourtit JP, Pangas SA, Matzuk MM and Eppig JJ: Oocyte
regulation of metabolic cooperativity between mouse cumulus cells
and oocytes: BMP15 and GDF9 control cholesterol biosynthesis in
cumulus cells. Development. 135:111–121. 2008. View Article : Google Scholar
|
|
23
|
Sugiura K, Su YQ, Diaz FJ, Pangas SA,
Sharma S, Wigglesworth K, O'Brien MJ, Matzuk MM, Shimasaki S and
Eppig JJ: Oocyte-derived BMP15 and FGFs cooperate to promote
glycolysis in cumulus cells. Development. 134:2593–2603. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu YQ, Chen LY, Zhang ZH and Wang ZC:
Effects of phosphatidylinositol-3 kinase/protein kinase b/bone
morphogenetic protein-15 pathway on the follicular development in
the mammalian ovary. Zhongguo Yi Xue Ke Xue Yuan Xue Bao.
35:224–228. 2013.In Chinese. PubMed/NCBI
|
|
25
|
Liu H, Luo LL, Qian YS, Fu YC, Sui XX,
Geng YJ, Huang DN, Gao ST and Zhang RL: FOXO3a is involved in the
apoptosis of naked oocytes and oocytes of primordial follicles from
neonatal rat ovaries. Biochem Biophys Res Commun. 381:722–727.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zheng W, Nagaraju G, Liu Z and Liu K:
Functional roles of the phosphatidylinositol 3-kinases (PI3Ks)
signaling in the mammalian ovary. Mol Cell Endocrinol. 356:24–30.
2012. View Article : Google Scholar
|
|
27
|
Adhikari D and Liu K: Molecular mechanisms
underlying the activation of mammalian primordial follicles. Endocr
Rev. 30:438–464. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Reddy KK, Lefkove B, Chen LB, Govindarajan
B, Carracedo A, Velasco G, Carrillo CO, Bhandarkar SS, Owens MJ,
Mechta-Grigoriou F and Arbiser JL: The antidepressant sertraline
downregulates Akt and has activity against melanoma cells. Cell
Melanoma Res. 21:451–456. 2008. View Article : Google Scholar
|
|
29
|
Yang P, Li X, Xu C, Eckert RL, Reece EA,
Zielke HR and Wang F: Maternal hyperglycemia activates an
ASK1-FoxO3a-caspase 8 pathway that leads to embryonic neural tube
defects. Sci Signal. 6:ra742013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu Y, Wang F, Reece EA and Yang P:
Curcumin ameliorates high glucose-induced neural tube defects by
suppressing cellular stress and apoptosis. AM J Obstet Gynecol.
212(802): e801–e808. 2015. View Article : Google Scholar
|
|
31
|
Li X, Xu M, Pitzer AL, Xia M, Boini KM, Li
PL and Zhang Y: Control of autophagy maturation by acid
sphingomyelinase in mouse coronary arterial smooth muscle cells:
Protective role in atherosclerosis. J Mol Med Berl. 92:473–485.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wei YM, Li X, Xu M, Abais JM, Chen Y,
Riebling CR, Boini KM, Li PL and Zhang Y: Enhancement of autophagy
by simvastatin through inhibition of Rac1-mTOR signaling pathway in
coronary arterial myocytes. Cell Physiol Biochem. 31:925–937. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li X, Gulbins E and Zhang Y: Oxidative
stress triggers Ca-dependent lysosome trafficking and activation of
acid sphingomyelinase. Cell Physiol Biochem. 30:815–826. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang Z, Pang X, Tang Z, Yin D and Wang Z:
Overexpression of hypoxia-inducible factor prolyl hydoxylase-2
attenuates hypoxia-induced vascular endothelial growth factor
expression in luteal cells. Mol Med Rep. 12:3809–3814.
2015.PubMed/NCBI
|
|
35
|
McGee EA and Hsueh AJ: Initial and cyclic
recruitment of ovarian follicles. Endocr Rev. 21:200–214.
2000.PubMed/NCBI
|
|
36
|
Hansen KR, Knowlton NS, Thyer AC,
Charleston JS, Soules MR and Klein NA: A new model of reproductive
aging: The decline in ovarian non-growing follicle number from
birth to menopause. Hum Reprod. 23:699–708. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Broekmans FJ, Knauff EA, te Velde ER,
Macklon NS and Fauser BC: Female reproductive ageing: Current
knowledge and future trends. Endocrinol Metab. 18:58–65. 2007.
|
|
38
|
Faddy MJ and Gosden RG: A model conforming
the decline in follicle numbers to the age of menopause in women.
Hum Reprod. 11:1484–1486. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tzivion G, Dobson M and Ramakrishnan G:
FoxO transcription factors; Regulation by AKT and 14-3-3 proteins.
Biochim Biophys Acta. 1813:1938–1945. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Reddy P, Zheng W and Liu K: Mechanisms
maintaining the dormancy and survival of mammalian primordial
follicles. Endocrinol Metab. 21:96–103. 2010.
|
|
41
|
Reddy P, Liu L, Adhikari D, Jagarlamudi K,
Rajareddy S, Shen Y, Du C, Tang W, Hämäläinen T, Peng SL, et al:
Oocyte-specific deletion of Pten causes premature activation of the
primordial follicle pool. Science. 319:611–613. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
John GB, Gallardo TD, Shirley LJ and
Castrillon DH: Foxo3 is a PI3K-dependent molecular switch
controlling the initiation of oocyte growth. Dev Biol. 321:197–204.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Otsuka F, Yamamoto S, Erickson GF and
Shimasaki S: Bone morphogenetic protein-15 inhibits
follicle-stimulating hormone (FSH) action by suppressing FSH
receptor expression. J Biol Chem. 276:11387–11392. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wymann MP and Pirola L: Structure and
function of phosphoinositide 3-kinases. Biochim Biophys Acta.
1436:127–150. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Whitman M, Downes CP, Keeler M, Keller T
and Cantley L: Type I phosphatidylinositol kinase makes a novel
inositol phospholipid, phosphatidylinositol-3-phosphate. Nature.
332:644–646. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Burgering BM and Coffer PJ: Protein kinase
B (c-Akt) in phosphatidylinositol-3-OH kinase signal transduction.
Nature. 376:599–602. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Franke TF, Kaplan DR, Cantley LC and Toker
A: Direct regulation of the Akt proto-oncogene product by
phosphatidylinositol-3,4-bisphosphate. Science. 275:665–668. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang XL, Xu J, Zhang XH, Qiu BY, Peng L,
Zhang M and Gan HT: PI3K/Akt signaling pathway is involved in the
pathogenesis of ulcerative colitis. Inflamm Res. 60:727–734. 2011.
View Article : Google Scholar : PubMed/NCBI
|