Introduction
Atrial fibrillation (AF) is the most common
sustained arrhythmia in clinical practice (1). It often occurs concomitantly with
other cardiovascular diseases, including hypertension, congestive
heart failure, coronary artery disease, and valvular heart disease
(2,3). Rheumatic mitral valve disease (RMVD),
a major cardiovascular disease in developing countries affected by
rheumatic fever, is a major clinical risk factor for AF (4,5).
Development of effective therapies and preventative strategies are
crucial for the control of AF-associated morbidity and mortality.
However, Currently, medical interventions for AF are relatively
limited, as the precise mechanisms of AF have not been completely
elucidated. Therefore, novel methods to probe the underlying
mechanisms of AF and potential novel mechanism-based therapeutic
strategies are required (6).
The field of cardiovascular proteomics has shed new
light on understanding the regulatory mechanisms of AF (7). Progression from paroxysmal AF to
persistent, and then to permanent AF involves complex changes in
gene expression, and subsequent changes in protein expression and
activity (8). Proteomic techniques
are a powerful tool to evaluate global protein changes in diseased
hearts and discover novel proteins, diagnostic biomarkers, and
potential drug targets for the development of novel therapeutic
agents (7). Two-dimensional
fluorescence difference gel electrophoresis (2-D DIGE) can provide
an overview of the proteome during disease (9). Previous use of proteomic methods have
identified differentially expressed proteins associated with AF
(AF-associated proteins) in several animal models (10,11)
and human heart tissue (12–16).
Genomic (8,17), morphological (18,19),
and electrophysiological (20–22)
differences have been observed between the right atrium (RA) and
left atrium (LA). This suggests that different mechanisms regulate
the RA and LA in AF (23). Thus,
it is unsurprising that the AF-associated proteins of the RA may
differ from those of the LA. However, the majority AF proteomics
studies have primarily focused on the RA. Thus, potential
differences in AF-associated proteins between RA and LA remain
unknown. Given the complexity of AF progression, a better
understanding of the differences in AF-associated proteins between
RA and LA may elucidate novel cardioprotective strategies.
Thus, the aim of the present study was to perform
2-D DIGE proteomics analysis to compare potential differences of
AF-associated proteins in the right atrial appendage (RAA) and left
atrial appendage (LAA) from patients with RMVD that were either in
sinus rhythm (SR) or AF.
Materials and methods
Ethics approval
Approval for this study was obtained from the Human
Ethics Committee at the First Affiliated Hospital of Sun Yat-Sen
University. The investigation complied with the principles that
govern the use of human tissues outlined in the Declaration of
Helsinki. All patients provided informed consent prior to
participation in the study.
Human tissue preparation
Human tissue preparation was performed as previously
described (8). Briefly, RAA and
LAA tissues were obtained from the same patient as surgical
biopsies at the time of the mitral valve replacement surgery (RAA
from cannulation site and LAA during ligation). Tissues were
immediately snap frozen in liquid nitrogen, and stored at −80°C
prior to use. Tissue samples from the RAA and LAA were obtained
from 18 patients with RMVD. The patients in the SR group (n=8) had
no history of AF. The patients in the AF group (n=10) presented
with a documented history of arrhythmias for >6 months prior to
surgery. The diagnosis of AF was reached by evaluating medical
records and 12-lead electrocardiogram findings. Patients with SR
also had no history of using antiarrhythmic drugs and were screened
to ensure that they had never experienced AF. Routine pre-operative
color Doppler echocardiography was performed on all patients.
Pre-operative functional status was recorded according to New York
Heart Association (NYHA) classifications.
Preparation of protein extracts
Protein extracts were isolated from tissues based on
previously published protocols (24). Briefly, tissues (approximately 0.1
g) were homogenized in lysis buffer [7 M urea, 2 M thiourea, 30 mM
Tris, 4% (w/v) CHAPS cell lysis buffer, 40 mM dithiothreitol (DTT),
0.6 mM phenylmethylsulfonyl fluoride] and the supernatant was
collected following centrifugation (2,000 × g, 4°C, 40 min).
To remove non-protein material from the extract and determine the
final protein concentration, the 2-D Clean-up kit (GE Healthcare
Life Sciences, Chalfont, UK) and 2-D Quant kit (GE Healthcare Life
Sciences) were used according to manufacturer's instructions.
Protein labeling with CyDye DIGE
fluorophores
Protein extracts were labeled with three CyDye DIGE
fluorophores, Cy2, Cy3 and Cy5 (GE Healthcare Life Sciences), for
2-D DIGE technology, according to the manufacturer's protocols.
Briefly, equal amounts of protein from the same group were pooled
and divided into four equal portions (50 µg each). The latter
portions were labeled with 400 pmol Cy3 or Cy5 according to the
experimental design. A pooled sample consisting of equal amounts of
all samples was used as the pooled internal standard and labeled
with Cy2. After incubating samples on ice for 30 min in the dark,
the labeling reaction was stopped with 10 mM lysine. For each gel,
Cy2-, Cy3- and Cy5-labeled proteins (50 µg each) were mixed and
calibrated to 450 µl with rehydration buffer [7 M urea, 2 M
thiourea, 4% (w/v) CHAPS cell lysis buffer, 40 mM DTT, 1% IPG
buffer (pH 4–7), 0.002% (w/v) bromophenol blue].
2-D electrophoresis
2-D electrophoresis was performed as previously
described (25). The labeled
protein mixture in each gel was applied to ImmobilineDryStrip
strips (24 cm, pH 4–7; GE Healthcare Life Sciences). Isoelectric
focusing (IEF) was performed with an EttanIPGphor II apparatus (GE
Healthcare Life Sciences) using the follow steps: 30 V for 12 h,
500 V for 1 h, 1,000 V for 1 h, and 10,000 V for up to a total of
85,000 V/h. Following IEF, the proteins were reduced and alkylated
by successive 15 min treatments with equilibration buffer
containing 2% (w/v) DTT followed by 2.5% (w/v) iodoacetamide.
Proteins were then resolved on 12.5% SDS-PAGE gels using an
EttanDALTsix instrument (GE Healthcare Life Sciences). In order to
facilitate mass spectrometry analysis, 500 µg of unlabeled pool
protein sample was run in parallel on a preparative gel and stained
by Deep Purple staining (GE Healthcare Life Sciences) according to
the manufacturer's instructions.
Gel image acquisition and
analysis
Gel images were acquired on a Typhoon 9400 scanner
(GE Healthcare Life Sciences) and analyzed using DeCyder software
(version 6.0; GE Healthcare Life Sciences) as previously described
(26,27). Briefly, Cy2-, Cy3- and Cy5-labeled
images of each gel were scanned with excitation/emission
wavelengths of 488/520, 532/580 and 633/670 nm, respectively.
Following CyDye labeling, signals were imaged and the gels were
stained using Deep Purple total protein stain (GE Healthcare Life
Sciences) according to standard protocols and scanned with
excitation/emission wavelengths of 532/560 nm. The 2-D DIGE gel
images were subsequently analyzed with DeCyder software. Protein
expression patterns for SR-RAA were compared with AF-RAA, and
SR-LAA were compared with AF-LAA. Ratios of proteins that increased
or decreased >1.5-fold (t-test, P<0.05) were considered
significant changes (9). The
corresponding protein spots were also selected in the stained
preparative gel for spot picking.
Spot picking and enzymatic
digestion
Selected protein spots in the preparative gels were
automatically picked and handled in an Ettan Spot Handling
Workstation (GE Healthcare Life Sciences) for the preparation of
the protein sample for matrix-assisted laser desorption/ionization
time-of-flight mass spectrometer/mass spectrometry (MALDI-TOF/TOF
MS) analysis. In the automated procedure, the selected protein
spots were picked, washed with 15 mM NH4HCO3
and 50% v/v methanol, then digested with 20 ng/ml trypsin
(sequencing grade; Promega Corporation, Madison, WI, USA) in 20 mM
NH4HCO3 for 2 h at 37°C. Tryptic peptides
were extracted with 50% v/v acetonitrile (ACN) and 0.5% v/v
trifluoroacetic acid (TFA), and dissolved in 5 mg/ml
R-cyano-4-hydroxycinnamic acid (GE Healthcare Life Sciences) matrix
in 50% (v/v) ACN and 0.1% (v/v) TFA. Finally, samples were spotted
on the mass spectrometry sample plate.
MALDI-TOF/TOF MS analysis and database
searching
Mass spectrometry analysis was performed using an
Utraflex III MALDI-TOF/TOF MS (Bruker Corporation, Ettlingen,
Germany) operating in a positive ion reflector mode. Monoisotopic
peak masses were acquired in a mass range of 700–4,000 Da, with a
signal-to-noise ratio >200. Four of the most intense ion
signals, excluding common trypsin autolysis peaks and matrix ion
signals, were automatically selected as precursors for mass
spectrometer/mass spectrometry (MS/MS) acquisition. Peptide mass
fingerprint (PMF) combined with MS/MS spectra was blasted against
the NCBInr database using the Biotools software (version 3.2;
Bruker Corporation) and MASCOT version 2.2 (Matrix Science, Inc.,
Boston, MA, USA). Search parameters were set as follows: Homo
sapiens, trypsin cleavage, one missed cleavage allowed,
carbamidomethylation as fixed modification, oxidation of
methionines allowed as variable modification, peptide mass
tolerance at 50 ppm, and fragment tolerance at 0.5 Da. The criteria
for successfully identifying a protein was as follows: Ion score
confidence interval (%) for PMF and MS/MS data >95%, peptide
count (hit) ≥4, and at least two peptides of distinct sequence were
identified in MS/MS analysis.
Protein categorization and
protein-protein interaction prediction
Protein categorization and protein-protein
interaction predictions were performed as previously described
(9). Briefly, the identified
proteins were classified, according to the Protein Analysis Through
Evolutionary Relationships (PANTHER) system (http://www.pantherdb.org), which classified genes and
proteins by their function. The PANTHER ontology, a highly
controlled vocabulary (ontology terms) by molecular function,
biological processes and molecular pathways, was employed to
categorize proteins into families and subfamilies with shared
function. The prediction of protein-protein interactions was
performed by utilizing the Search Tool for the Retrieval of
Interacting Genes/Proteins (STRING) system (http://www.string-db.org). STRING is a database of
known and predicted protein interactions that include direct
(physical) and indirect (functional) associations.
Western blot analysis
Western blot analysis was performed as previously
described (28). Briefly, protein
lysates were prepared from human tissues, separated on SDS-PAGE,
transferred to a polyvinylidene difluoride membrane, and subjected
to immunoblotting at 4°C overnight with the following antibodies:
Anti-aldo-keto reductase family 1 member B10 (AKR1B10; diluted
1:10,000; polyclonal antibody; cat. no. S1646; Epitomics,
Burlingame, CA, USA), anti-crystallin α B (CRYAB; diluted 1:2000;
polyclonal antibody; cat. no. PAB7394; Abnova Corporation, Taipei,
Taiwan), anti-annexin 4 (ANAX4; diluted 1:800; polyclonal antibody;
cat. no. H00000307-B01P; Abnova Corporation), anti-G protein
subunit α o1 (GNAO1; diluted 1:10,000; polyclonal antibody; cat.
no. S3107; Epitomics), anti-ribonuclease H1 (RNase H1; diluted
1:800; monoclonal antibody; cat. no. ab56560; Abcam, Cambridge, MA,
USA), anti-moesin (MSN; diluted 1:2,000; polyclonal antibody; cat.
no. PAB7062; Abnova Corporation), anti-nestin (NES; diluted
1:6,000; polyclonal antibody; cat. no. PAB12375; Abnova
Corporation) and anti-transferrin (diluted 1:8,000; polyclonal
antibody; cat. no. S0860; Epitomics), anti-osteoglycin (diluted
1:500; monoclonal antibody; cat. no. 5518-1; Epitomics). β-actin
was assessed as a loading control (diluted 1:20,000; monoclonal
antibody; cat. no. 60008-1-Ig; ProteinTech Group, Inc., Chicago,
IL, USA). After washing, membranes were incubated with horseradish
peroxidase (HRP)-conjugated secondary antibodies (goat anti-mouse,
cat. no. SA00001-1; goat anti-rabbit, cat. no. SA00001-2; or rabbit
anti-goat, cat. no. SA00001-4; all from ProteinTech Group, Inc.)
and visualized using an enhanced chemiluminescence western blot
detection system (Immobilon Western Chemiluminescent HRP substrate;
Merck Millipore, Darmstadt, Germany) on X-ray film (Kodak,
Rochester, NY, USA). All the proteins were detected on the same
blot. For certain protein with molecular weights very close to each
other, the blots were stripped and reprobed using Restore™ PLUS
Western Blot Stripping Buffer(Thermo Fisher Scientific Inc.,
Waltham, MA, USA) according to the manufacturer's instructions.
Images were scanned and analyzed using Kodak Molecular Imaging
software (version 5.0). Non-specific binding was subtracted, and
the band signals were expressed as relative protein amounts
compared with β-actin protein levels.
Statistical analysis
Data of clinical characteristics of patients and
western blot quantification are presented as the mean ± standard
deviation. Student's t-test was used for statistical comparison of
the data and to calculate significant differences in the relative
abundance of individual protein spots between the two groups during
2-D DIGE analysis. The SPSS 16.0 software package (SPSS, Inc.,
Chicago, IL, USA) was used to conduct the statistical analyses and
P<0.05 (two-tailed) was considered to indicate a statistically
significant difference.
Results
Clinical characteristics of the
patients with SR and AF
Tissue from RAA and LAA was obtained from each
patient. No significant differences in terms of age, gender or NYHA
class were observed between the SR and AF groups. Pre-operative
color Doppler echocardiography demonstrated that the LA size of the
patients with AF was significantly increased compared with the SR
patients. However, no differences in the left ventricular
end-diastolic diameter and left ventricular ejection fraction were
observed between the groups (Table
I).
 | Table I.Clinical characteristics of the
patients with SR and AF. |
Table I.
Clinical characteristics of the
patients with SR and AF.
| Characteristic | SR (n=8) | AF (n=10) |
|---|
| Gender
(male/female) | 5/3 | 5/5 |
| Age (years) | 50.16±6.88 | 51.42±7.12 |
| LA size (mm) | 43.31±3.23 |
57.65±5.08a |
| LVEDD (mm) | 48.35±5.15 | 50.13±3.07 |
| LVEF (%) | 61.34±5.66 | 58.71±3.81 |
| NYHA class |
|
|
| II | 6 | 7 |
|
III | 2 | 3 |
2-D DIGE analysis and MALDI-TOF/TOF MS
identification
Following 2-D DIGE analysis, a total of 2,813 spots
were observed to be well matched across all gels using the DeCyder
software analysis. The 2-D DIGE gel images are presented in
Fig. 1. Following visual review,
61 protein spots of high abundance that exhibited significantly
altered expression (AF-RAA vs. SR-RAA and AF-LAA vs. SR-LAA) were
selected for MALDI-TOF/TOF MS analysis. A total of 32
differentially expressed proteins were identified, including 15
that were upregulated and 17 that were downregulated in the AF-RAA
tissues compared with the SR-RAA tissues (Table II). Additionally, a total of 31
differentially expressed proteins, including 13 that were
upregulated and 18 that were downregulated in the AF-LAA tissues
compared with the SR-LAA tissues (Table II).
 | Figure 1.Analysis of RAA and LAA samples by
2-D DIGE analysis. Proteins extracted from AF (AF-RAA or AF-LAA)
and SR (SR-RAA or SR-LAA) groups were labeled with Cy3 and Cy5,
respectively. An internal standard comprised of proteins pooled
from a combination of the AF-RAA, AF-LAA, SR-RAA and SR-LAA groups
was labeled with Cy2 and included in all gels. The labeled AF and
SR samples were mixed and separated on a 2-D DIGE gel (isoelectric
focusing on pH 4–7 strips followed by 12.5% second dimension
SDS-PAGE). Green spots represent downregulated proteins, and red
spots represent upregulated proteins in AF samples compared with SR
samples. The yellow arrows represent the identified proteins that
exhibited significantly differential expression in the AF and SR
samples. (A) AF-RAA vs. SR-RAA samples. (B) AF-LAA vs. SR-LAA
samples. RAA, right atrial appendage; LAA, left atrial appendage;
2-D DIGE, two-dimensional fluorescence difference gel
electrophoresis; AF, atrial fibrillation; SR, sinus rhythm; MW,
molecular weight. |
| Table II.Differential proteins identified by
mass spectrometry following 2-D DIGE of the RAA and LAA
samples. |
Table II.
Differential proteins identified by
mass spectrometry following 2-D DIGE of the RAA and LAA
samples.
|
|
|
| Av.
ratiob | t-test |
|
|---|
| Accession
no.a | MW (kDa) | pI | RAAc | LAAd | RAAc | LAAd | Protein name |
|---|
| In RAA and LAA
(n=17) |
|
|
|
|
|
|
|
|
gi|88699 | 2.4 | 11.55 | 1.7 | 2.36 | 0.0065 | 0.013 | T-cell receptor β
chain J region (clone HBP22)-human (fragment) |
|
gi|7669550 | 124 | 5.5 | 1.56 | 1.5 | 0.0012 | 0.0044 | Vinculin isoform
meta-VCL |
|
gi|5669804 | 72 | 9.1 | −1.76 | −1.83 | 0.016 | 0.00039 | Polycystin-2-like
protein |
|
gi|42543698 | 42 | 6.62 | 1.57 | 1.5 | 0.038 | 0.011 | Chain A, The
Crystal Structure Of The Human Hsp70 Atpase Domain |
|
gi|41350923 | 110 | 5.85 | 1.61 | 2.06 | 0.011 | 0.0017 | Collagen, type VI,
α 2 |
|
gi|386973 | 61 | 5.3 | −1.88 | −1.87 | 0.022 | 0.00066 | β-Myosin heavy
chain |
|
gi|386972 | 62 | 5.3 | −1.88 | −1.91 | 0.01 | 0.0019 | α-Myosin heavy
chain |
|
gi|386970 | 71 | 5.32 | −1.79 | −1.94 | 0.019 | 0.00019 | Myosin heavy chain
β-subunit |
|
gi|33150528e | 34 | 5.33 | −1.75 | −2.01 | 0.012 | 0.0079 | Osteoglycin |
|
gi|223468663e | 36 | 7.67 | −1.68 | −2.02 | 0.026 | 0.0026 | Aldo-ketoreductase
family 1 member B10 |
|
gi|19908424 | 54 | 5.21 | 1.99 | 1.94 | 0.036 | 0.038 | Mutant desmin |
|
gi|189617e | 36 | 5.65 | 1.55 | 1.73 | 0.021 | 0.026 | Protein PP4-X |
|
gi|119622366 | 8.0 | 10.91 | −2.23 | −2.08 | 0.018 | 0.00011 | hCG1813095 |
|
gi|119622046 | 16 | 12.02 | −1.58 | −1.68 | 0.0019 | 0.023 | hCG1646228 |
|
gi|119612724 | 30 | 4.88 | −1.79 | −2.02 | 0.014 | 0.0028 | Actin, α, cardiac
muscle, isoform CRA_c |
|
gi|119586559 | 224 | 5.58 | −2.57 | −2.14 | 0.0082 | 0.0052 | Myosin, heavy
polypeptide 6, cardiac muscle, α (cardiomyopathy, hypertrophic 1),
isoform CRA_b |
|
gi|119576987 | 34 | 9.33 | −1.88 | −2.03 | 0.011 | 0.0041 | DNA cross-link
repair 1B |
| Only in RAA
(n=15) |
|
|
|
|
|
|
|
|
gi|94981553 | 19 | 4.92 | −2.72 | – | 0.024 | – | Myosin regulatory
light chain 2, ventricular/cardiac muscle isoform |
|
gi|62897129 | 71 | 5.28 | −1.55 | – | 0.027 | – | Heat shock 70kDa
protein 8 isoform 1 variant |
|
gi|55749932 | 54 | 5.21 | 2.04 | – | 0.0035 | – | Desmin |
|
gi|51103391 | 12 | 8.98 | −1.68 | – | 0.011 | – | Immunoglobulin
variable region VL kappa domain |
|
gi|5031875 | 65 | 6.4 | 1.6 | – | 0.017 | – | Lamin-A/C isoform
2 |
|
gi|46593007 | 53 | 5.94 | 4.82 | – | 0.048 | – | Cytochrome b-c1
complex subunit 1, mitochondrial precursor |
|
gi|4505257e | 68 | 6.08 | 1.62 | – | 0.029 | – | Moesin |
|
gi|4505047 | 39 | 6.16 | −1.63 | – | 0.023 | – | Lumican
precursor |
|
gi|4503057e | 20 | 6.76 | 1.99 | – | 0.0016 | – | α-Crystallin B
chain |
|
gi|31615330 | 68 | 5.66 | 2.09 | – | 0.0091 | – | Chain A, human
serum albumin mutant R218h complexed with thyroxine |
|
gi|223170 | 47 | 5.54 | 1.56 | – | 0.032 | – | Fibrinogen γ |
|
gi|145942333 | 9.6 | 5.5 | 1.79 | – | 0.019 | – | Immunoglobulin
heavy chain variable region |
|
gi|119581148 | 58 | 6.95 | −1.69 | – | 0.002 | – | Keratin 9 |
|
gi|116283748e | 66 | 5.15 | −2.04 | – | 0.0068 | – | Nestin protein |
|
gi|114615454 | 44 | 6.57 | 1.79 | – | 0.015 | – | PREDICTED:
dihydrolipoamide dehydrogenase isoform 1 |
| Only in LAA
(n=14) |
|
|
|
|
|
|
|
|
gi|998467 | 3.1 | 4.41 | – | 1.57 | – | 0.044 | 48 kDa histamine
receptor subunit peptide 4 |
|
gi|62414289 | 54 | 5.06 | – | 2.71 | – | 0.0035 | Vimentin |
|
gi|56204818 | 38 | 5.39 | – | −1.84 | – | 0.0016 | Actin, α 1,
skeletal muscle |
|
gi|4885079 | 33 | 9.31 | – | 2.23 | – | 0.0085 | ATP synthase
subunit γ, mitochondrial isoform H (heart) precursor |
|
gi|48476973 | 137 | 5.29 | – | −1.67 | – | 0.0047 | Rhabdomyosarcoma
antigen MU-RMS-40.7B |
|
gi|4501893 | 104 | 5.31 | – | 2.17 | – | 0.0078 | α-Actinin-2 |
|
gi|34811370 | 13 | 10.15 | – | −2.26 | – | 0.0062 | Chain B, crystal
structure of the 46kDa domain of human cardiac troponin in the
Ca2+ saturated form |
|
gi|34190601e | 35 | 5.18 | – | −1.69 | – | 0.00024 | G protein subunit α
o1 protein |
|
gi|22080672 | 9.6 | 10.96 | – | 1.6 | – | 0.00089 | MHC class I
antigen |
|
gi|15029922e | 50 | 4.83 | – | −1.67 | – | 0.011 | Ribonuclease H1
protein |
|
gi|119618538 | 13 | 10.95 | – | 1.76 | – | 0.0086 | hCG2016250, isoform
CRA_g |
|
gi|119612436 | 36 | 8.27 | – | −1.97 | – | 0.0002 | hCG1779566, isoform
CRA_a |
|
gi|110590599e | 77 | 6.85 | – | 1.58 | – | 0.014 | Chain A, apo-human
serum transferrin |
|
gi|109102505 | 35 | 5.82 | – | −1.6 | – | 0.0024 | PREDICTED: similar
to serine/threonine-protein phosphatase PP1-β catalytic subunit
(PP-1B) isoform 1 |
Comparison of differentially expressed
proteins between RAA and LAA tissues
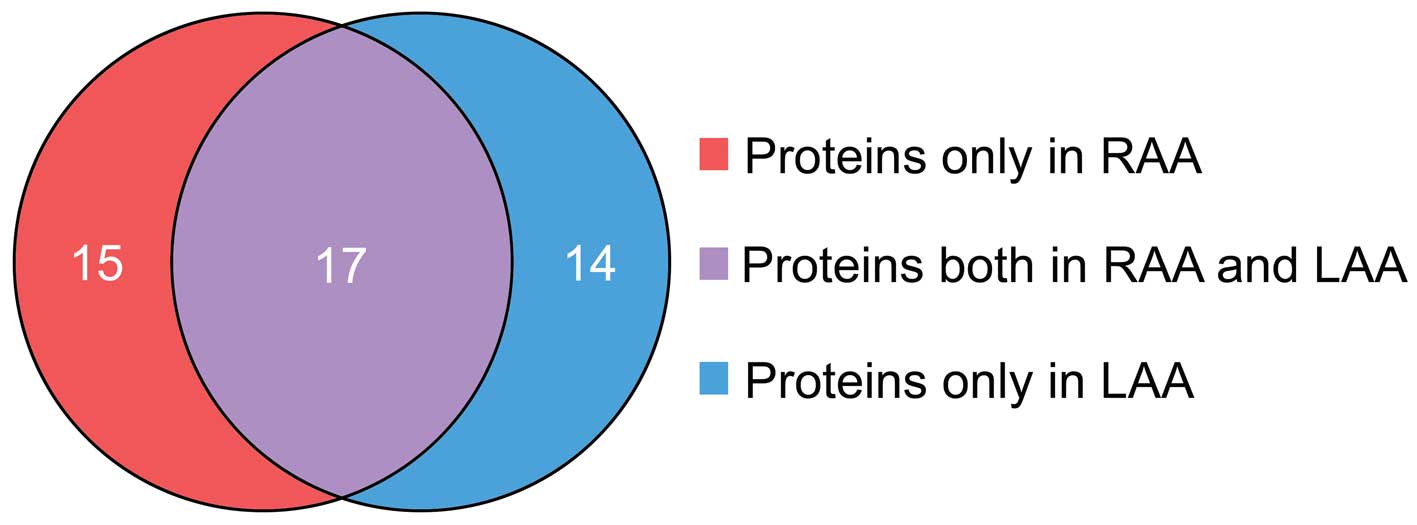
Using 2-D DIGE proteomics methodology, a total of 46
differentially expressed proteins were detected (in RAA or LAA
tissues). Among these, 32 proteins were expressed in RAA tissues,
and 31 proteins in LAA tissues. Furthermore, 17 proteins were
identified in RAA and LAA tissues, whereas 15 proteins were
detected only in RAA tissues and 14 proteins were only in LAA
tissues (Table II; Fig. 2).
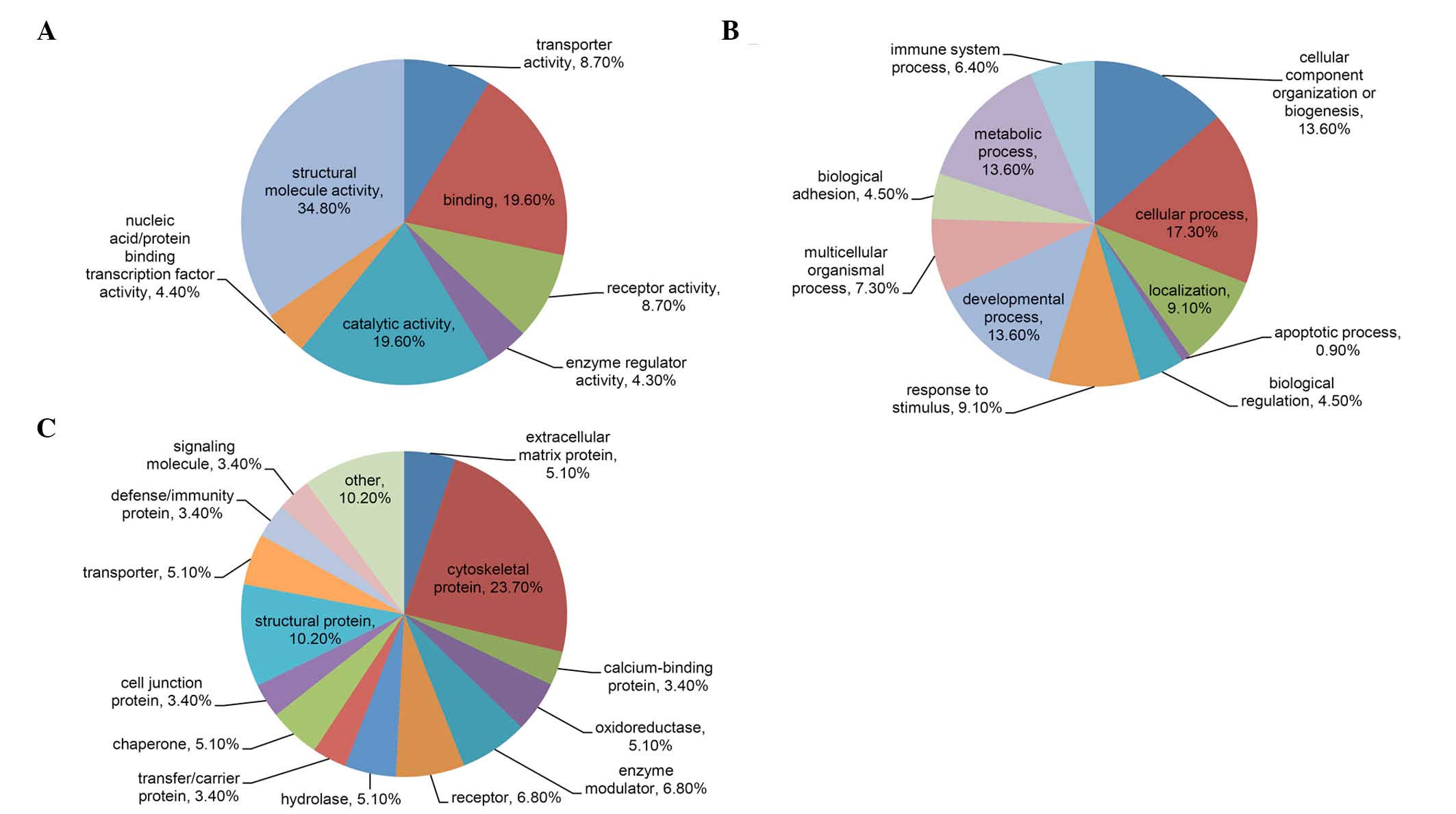
Classification of differentially
expressed proteins and the protein-protein interaction network
According to cellular function and processes, a
total of 46 differentially expressed proteins were distributed into
categories based on molecular function, biological processes and
protein classes using the PANTHER classification system. Results
are presented in Fig. 3. The most
dominant function that the identified proteins were involved in was
structural activity (34.8%), followed by binding (19.6%), catalytic
activity (19.6%), transporter activity (8.7%), receptor activity
(8.7%), nucleic acid/protein binding transcription factor activity
(4.4%) and enzyme regulator activity (4.3%). The majority of the
identified proteins had functions in cellular processes (17.3%),
cellular component organization or biogenesis (13.6%), metabolic
processes (13.6%) and developmental processes (13.6%). Other
biological processes that the proteins were involved in included
localization (9.1%), response to stimuli (9.1%), and multicellular
organismal processes (7.3%). As for protein class, the majority of
the proteins belonged in categories associated with cytoskeletal
proteins (23.7%) and structural proteins (10.2%), while others
belonged to categories associated with enzyme modulators (6.8%),
receptors (6.8%) and calcium-binding proteins (3.4%).
The protein interaction network for the identified
proteins was constructed by STRING (Fig. 4) in order to improve the
understanding of the pathogenic mechanisms in AF with RMVD.
Biological systems can be modeled as complex network systems with
many interactions between components of different pathways. These
interactions provide us with essential information about the
function and behavior of the analyzed proteins (9).
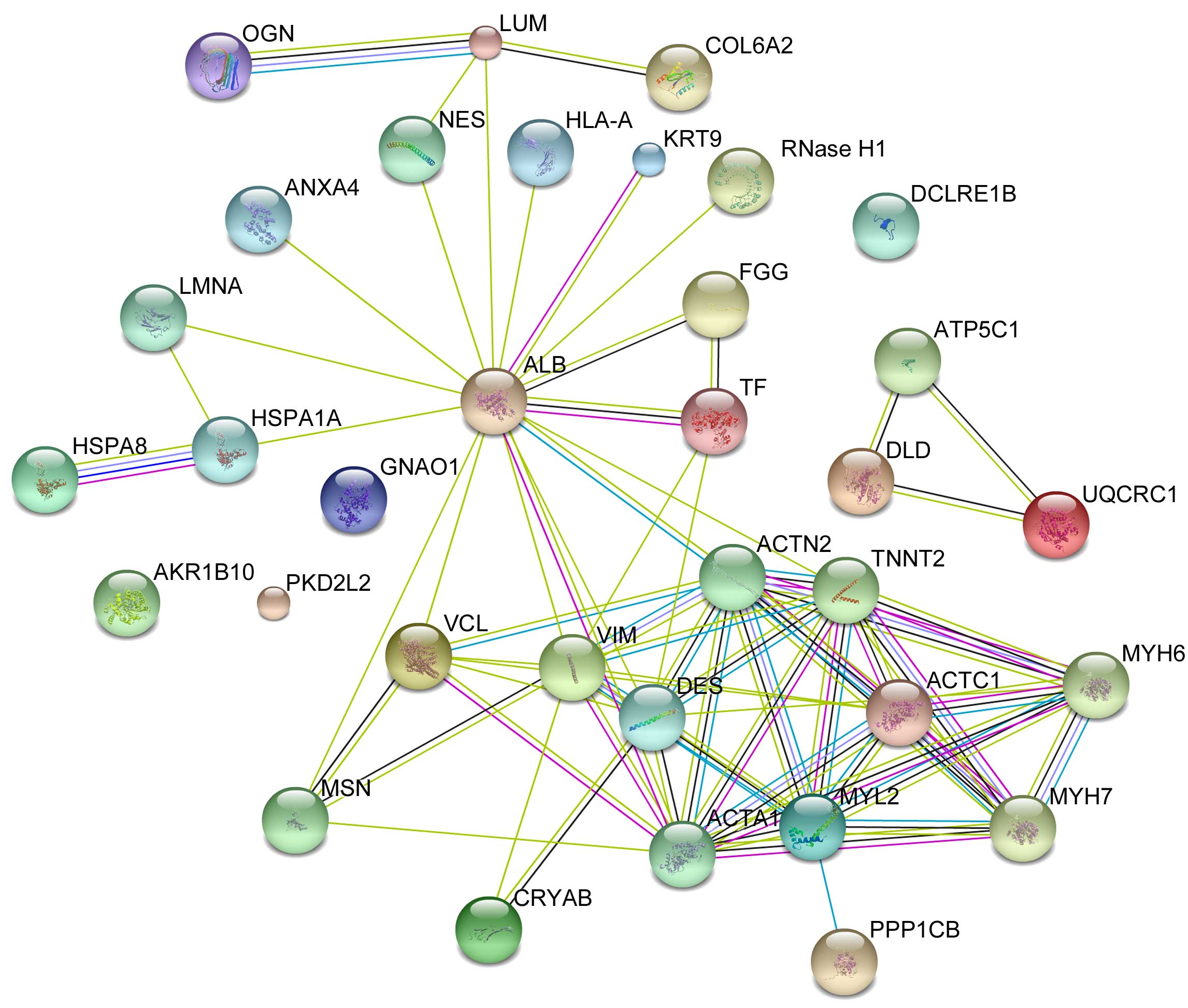 | Figure 4.The protein-protein interaction
network predicted by STRING. OGN, osteoglycin; LUM, lumican;
COL6A2, collagen type VI α 2 chain; ANXA4, annexin A4; NES, nestin;
HLA-A, major histocompatibility complex class I A; KRT9, keratin 9;
RNase H1, ribonuclease H1; DCLRE1B, DNA cross-link repair 1B; LMNA,
lamin A/C; ALB, albumin; FGG, fibrinogen γ chain; HSPA8, heat shock
protein family A (Hsp70) member 8; HSPA1A, heat shock protein
family A (Hsp70) member 1A; TF, transferrin; ATP5C1, ATP synthase
H+ transporting mitochondrial F1 complex γ polypeptide
1; DLD, dihydrolipoamide dehydrogenase; UQCRC1,
ubiquinol-cytochrome c reductase core protein I; AKR1B10, aldo-keto
reductase family 1 member B10; GNAO1, G protein subunit α o1;
PKD2L2, polycystin 2 like 2 transient receptor potential cation
channel; VCL, vinculin; VIM, vimentin; ACTN2, actinin α 2; TNNT2,
troponin T2 cardiac type; DES, desmin; ACTC1, actin α cardiac
muscle 1; MYH6, myosin heavy chain 6; MSN, moesin; ACTA1, actin α 1
skeletal muscle; MYL2, myosin light chain 2; MYH7, myosin heavy
chain 7; CRYAB, crystallin α B; PPP1CB, protein phosphatase 1
catalytic subunit β; STRING, Search Tool for the Retrieval of
Interacting Genes/Proteins. |
Validation of the 2-D DIGE proteomics
data by western blot analysis
To validate the data obtained from the 2-D DIGE
proteomics, western blot analysis was performed on 9 selected
proteins (Table II). According to
the western blot data, AKR1B10 and ANAX4 were AF-associated
proteins in RAA and LAA tissues (Fig.
5), whereas CRYAB and MSN were AF-associated proteins in only
RAA tissues. RNase H1 and transferrin were AF-associated proteins
only in LAA tissues (Fig. 5). The
western blot data validated the results for all of these proteins,
excluding osteoglycin, NES and GNAO1, AF-associated proteins which
were not detected to be differentially expressed in the RAA or LAA
of AF tissues when assessed using western blot analysis (Fig. 5).
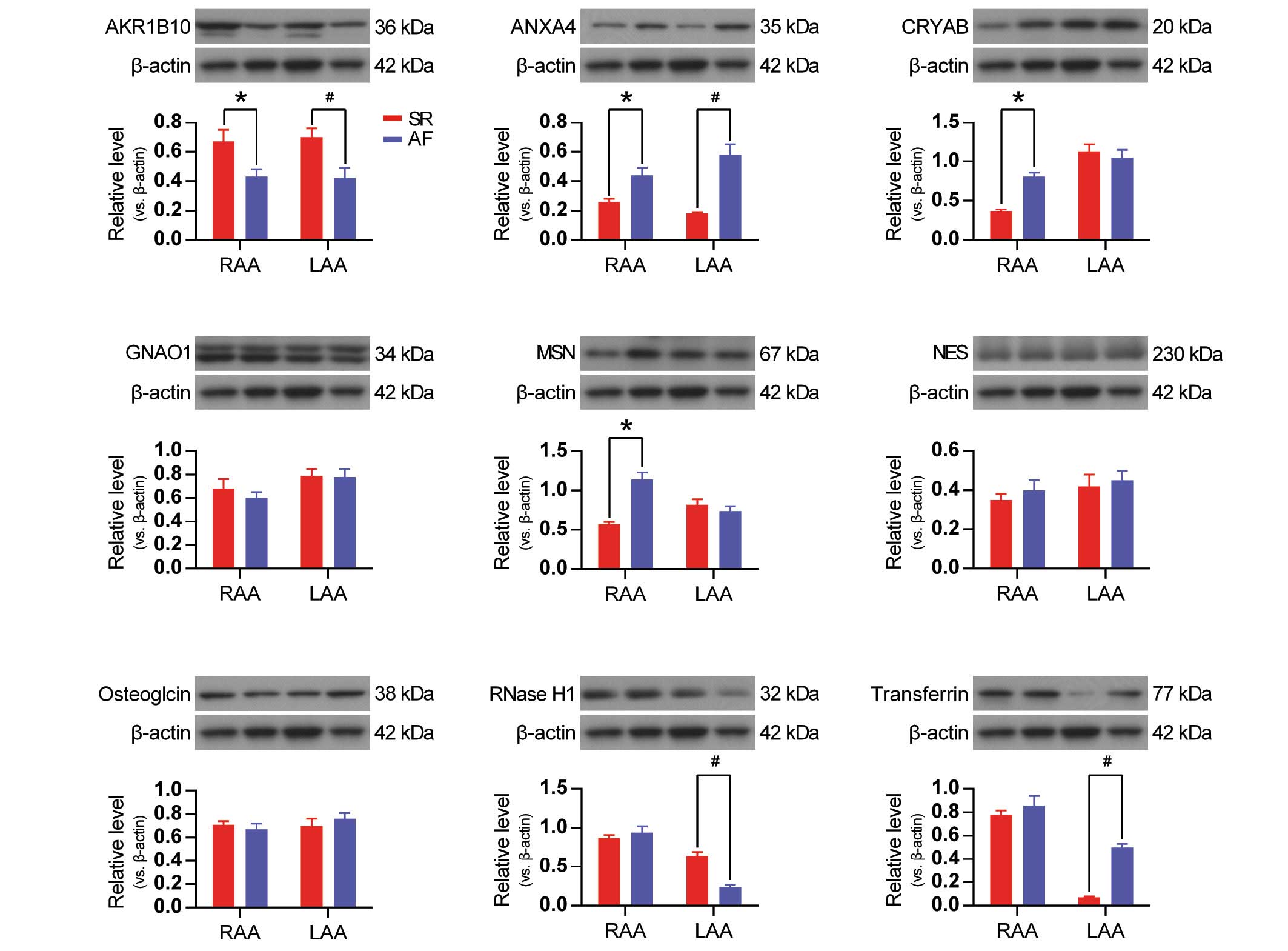 | Figure 5.Validation of the two-dimensional
fluorescence difference gel electrophoresis proteomics data by
western blot analysis of nine selected differentially expressed
proteins. β-actin is used as an internal loading control to
normalize the results. Data are presented as the mean ± standard
deviation of three independent experiments. *P<0.05, AF-RAA vs.
SR-RAA. #P<0.05, AF-LAA vs. SR-LAA. AKR1B10,
aldo-keto reductase family 1 member B10;; SR, sinus rhythm; AF,
atrial fibrillation; RAA, right atrial appendage; LAA, left atrial
appendage; CRYAB, crystallin α B; GNAO1, G protein subunit α o1;
MSN, moesin; NES, nestin; RNase H1, ribonuclease H1. |
Comparative analysis of the expression
of six validated proteins between the RAA and LAA tissues from RMVD
patients with SR
Six differentially expressed proteins were
identified and validated using 2-D DIGE proteomics and western blot
analysis. These proteins had different tissue distributions between
RAA and LAA. Additionally, it was aimed to elucidate whether
expression of these proteins was also different between RAA and LAA
tissues based on SR status. Western blot analysis was used to
compare the expression level of these proteins between the SR-RAA
and SR-LAA groups. Expression levels of AKR1B10, ANAX4 and RNase H1
were not different in the RAA and LAA of patients with SR (Fig. 6). However, CRYAB and MSN expression
was increased in the SR-LAA group compared with SR-RAA (Fig. 6). Transferrin expression was
increased in the SR-RAA group compared with the SR-LAA group
(Fig. 6).
Discussion
Previous studies have identified that both the RA
(29,30) and LA (31,32)
serve an important role in AF. However, intervention strategies
that only target the RA or LA do not ameliorate all of the
complications associated with AF (33–35).
This may reflect the necessity to understand the different
mechanisms involved in AF between the RA and LA. The development of
AF involves complex changes in gene expression and subsequent
changes in protein expression and activity. Thus, it is important
to understand the protein mechanisms of AF to facilitate the
development of potential novel mechanism-based therapeutic
strategies. Proteomic technology provides a comprehensive strategy
to investigate AF-associated proteins. Recently, there have been
many proteomic studies associated with AF (10–16).
However, the majority of proteomics studies on AF primarily focus
on the RA and less on the LA. Although two previous studies
(12,16) used proteomics analysis to identify
AF-associated proteins in RA and LA tissue from patients with
mitral valve disease, potential differences in AF-associated
proteins between the RA and LA were not determined. Therefore, the
potential differential expression of AF-associated proteins between
the RA and LA remain unknown. To the best of our knowledge, the
current study is the first to compare the potential differences of
AF-associated proteins in the RA and LA from RMVD patients.
Profiling protein expression patterns at specific
stages of disease can reflect the status of disease progression. We
hypothesize that if there are different mechanisms involved in AF
between the RA and LA, then changes in protein expression profiles
will also reflect this pattern. The differentially expressed
proteins were compared between RA and LA to reveal different
disease mechanisms underlying AF in the RA and LA. The present
study demonstrated that the development of AF in patients with RMVD
was associated with significant changes in protein expression in
RAA and LAA tissues and that these AF-associated proteins had
different distributions in RAA and LAA. Certain AF-associated
proteins were differentially expressed both in the RA and LA
compared with expression in SR samples, while others were different
only in the RA or LA. The current study further demonstrated that
the expression of certain proteins were not changed between the RAA
and LAA in patients with SR, whereas there is a difference in the
protein expression profiles of SR and AF between the RAA and LAA.
Therefore, it was hypothesize that differential distribution of
these AF-associated proteins may reflect different protein
mechanisms in the RA and LA during AF. Genomic (8,17),
morphological (18,19), and electrophysiological (20–22)
differences have been observed between the RA and LA in mechanisms
involved in AF. Thus, the present study provided proteomic evidence
that different mechanisms regulate the RA and LA in AF.
Protein expression profiles also reflect certain
spatial characteristics (depending on cell, tissue or organ type).
The current study observed that the protein expression levels of
CRYAB, MSN and transferrin were different when comparing RAA and
LAA tissues in patients with RMVD and SR. CRYAB and MSN expression
was increased in the SR-LAA group compared with the SR-RAA group,
however they were identified as AF-associated proteins only in RAA
tissue. By contrast, transferrin expression was increased in the
SR-RAA group compared with the SR-LAA, and was identified as an
AF-associated protein only in LAA tissue. These results suggest
that CRYAB, MSN and transferrin are tissue dependent AF-associated
proteins that may be involved in the progression from SR to AF as
observed by the initial expression studies performed in RAA and LAA
of RMVD patients with SR. The difference in the initial expression
of these proteins between RAA and LAA in patients with RMVD and SR
may due to the progress from healthy to RMVD status. Cooley et
al (36) reported that
microRNA expression profiles caused differential changes in the RA
and LA with the development from healthy to valvular heart diseased
condition. Thus, potentially, the certain proteins become
differentially expressed in the RA and LA during RMVD compared with
healthy settings. In addition, in patients with RMVD, the
association between LA size and AF is well established and LA
dilatation is considered to be a cause and a consequence of AF
(5). The results of the present
study revealed that the LA size of patients with AF was
significantly greater than in patients with SR (Table I), thus, potentially, the
significant structural remodeling occurring in the LA may also
alter the protein expression profiles and cause, at least
partially, differential regulation of AF-associated proteins in the
RA and LA in patients with RMVD.
The current study identified several AF-associated
proteins using 2-D DIGE proteomics analysis. Of these proteins, a
number may participate in the mechanisms associated with AF,
whereas others maybe a result of AF. The exact role of these
differentially expressed proteins in AF requires further
investigation. Stringent bioinformatics analysis will be required
to select for candidate proteins for future functional studies.
Bioinformatics analysis was performed in the current study using
the PANTHER and STRING systems to classify AF-associated proteins
and predict a protein-protein interaction network (Figs. 5 and 6). This analysis may provide
investigators with vital information to direct future research.
A major limitation of this study was the small
number of patients. This was due, in part, to the difficulty in
finding patients with RMVD and SR. In addition, because the study
was performed with human tissues with existing disease, experiments
could not be conducted to modulate the protein levels. Therefore,
exact targets and pathways by which alterations in these proteins
may cause AF in patients with RMVD remain elusive and require
further investigation. Finally, patients in this study were a
specific cohort with preserved systolic left ventricular function
and little comorbidity, undergoing mitral valve replacement
surgery. Thus, changes identified in this population may not be
representative of other cohort populations.
In conclusion, the current study identified
differentially expressed proteins that have potential associations
with AF in the RAA and LAA tissues from patients with RMVD. The
different distribution of these AF-associated proteins may reflect
different mechanisms underlying RAA and LAA involvement in AF.
These findings may be useful for the biological understanding of AF
in patients with RMVD and suggest potential therapeutic targets for
AF.
Acknowledgements
This work is supported by the Pearl River Scholar
Program (grant no. 80000-3210003) and the National Natural Science
Foundation of China (grant nos. 81370215 and 81500260). The authors
wish to acknowledge Dr. Kun-hua Hu (Key Laboratory on Proteomics,
Sun Yat-sen University, Guangzhou, China) for his technical
expertise. We are grateful to Dr. Gang Dai and Dr. Rong Fang (Key
Laboratory on Assisted Circulation, Ministry of Health, Guangzhou,
China) for their technical expertise.
Glossary
Abbreviations
Abbreviations:
|
AF
|
atrial fibrillation
|
|
SR
|
sinus rhythm
|
|
RMVD
|
rheumatic mitral valve disease
|
|
RA
|
right atrium
|
|
LA
|
left atrium
|
|
RAA
|
right atrial appendage
|
|
LAA
|
left atrial appendage
|
|
2-D DIGE
|
two-dimensional fluorescence
difference gel electrophoresis
|
|
IEF
|
isoelectric focusing
|
|
MALDI-TOF/TOF MS
|
matrix-assisted laser
desorption/ionization time-of-flight mass spectrometer/mass
spectrometry
|
|
MS/MS
|
mass spectrometer/mass
spectrometry
|
|
PMF
|
peptide mass fingerprint
|
References
|
1
|
Murphy NF, Simpson CR, Jhund PS, Stewart
S, Kirkpatrick M, Chalmers J, MacIntyre K and McMurray JJ: A
national survey of the prevalence, incidence, primary care burden
and treatment of atrial fibrillation in Scotland. Heart.
93:606–612. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Darby AE and Dimarco JP: Management of
atrial fibrillation in patients with structural heart disease.
Circulation. 125:945–957. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rosiak M, Dziuba M, Chudzik M,
Cygankiewicz I, Bartczak K, Drozdz J and Wranicz JK: Risk factors
for atrial fibrillation: Not always severe heart disease, not
always so ‘lonely’. Cardiol J. 17:437–442. 2010.PubMed/NCBI
|
|
4
|
Benjamin EJ, Levy D, Vaziri SM, D'Agostino
RB, Belanger AJ and Wolf PA: Independent risk factors for atrial
fibrillation in a population-based cohort. The Framingham Heart
Study. JAMA. 271:840–844. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Carabello BA: Modern management of mitral
stenosis. Circulation. 112:432–437. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nattel S: New ideas about atrial
fibrillation 50 years on. Nature. 415:219–226. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lam L, Lind J and Semsarian C: Application
of proteomics in cardiovascular medicine. Int J Cardiol. 108:12–19.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu H, Qin H, Chen GX, Liang MY, Rong J,
Yao JP and Wu ZK: Comparative expression profiles of microRNA in
left and right atrial appendages from patients with rheumatic
mitral valve disease exhibiting sinus rhythm or atrial
fibrillation. J Transl Med. 12:902014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Deng L, Jia HL, Liu CW, Hu KH, Yin GQ, Ye
JW, He CH, Chen JH, Xie YP, Dang R, et al: Analysis of
differentially expressed proteins involved in hand, foot and mouth
disease and normal sera. Clin Microbiol Infect. 18:E188–E196. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lai LP, Lin JL, Lin CS, Yeh HM, Tsay YG,
Lee CF, Lee HH, Chang ZF, Hwang JJ, Su MJ, et al: Functional
genomic study on atrial fibrillation using cDNA microarray and
two-dimensional protein electrophoresis techniques and
identification of the myosin regulatory light chain isoform
reprogramming in atrial fibrillation. J Cardiovasc Electrophysiol.
15:214–223. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
De Souza AI, Cardin S, Wait R, Chung YL,
Vijayakumar M, Maguy A, Camm AJ and Nattel S: Proteomic and
metabolomic analysis of atrial profibrillatory remodelling in
congestive heart failure. J Mol Cell Cardiol. 49:851–863. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Modrego J, Maroto L, Tamargo J, Azcona L,
Mateos-Cáceres P, Segura A, Moreno-Herrero R, Pérez-Castellanos N,
Delpón E, Pérez-Villacastín J, et al: Comparative expression of
proteins in left and right atrial appendages from patients with
mitral valve disease at sinus rhythm and atrial fibrillation. J
Cardiovasc Electrophysiol. 21:859–868. 2010.PubMed/NCBI
|
|
13
|
Mayr M, Yusuf S, Weir G, Chung YL, Mayr U,
Yin X, Ladroue C, Madhu B, Roberts N, De Souza A, et al: Combined
metabolomic and proteomic analysis of human atrial fibrillation. J
Am Coll Cardiol. 51:585–594. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
García A, Eiras S, Parguiña AF, Alonso J,
Rosa I, Salgado-Somoza A, Rico TY, Teijeira-Fernández E and
González-Juanatey JR: High-resolution two-dimensional gel
electrophoresis analysis of atrial tissue proteome reveals
down-regulation of fibulin-1 in atrial fibrillation. Int J Cardiol.
150:283–290. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang WJ, Zhou R, Zeng XR, Tan XQ, Cheng
ZH, Tang MH, Gou LT, Chen LJ, Tong AP, He Y and Yang JL:
Comparative proteomic analysis of atrial appendages from rheumatic
heart disease patients with sinus rhythm and atrial fibrillation.
Mol Med Rep. 4:655–661. 2011.PubMed/NCBI
|
|
16
|
Zhang P, Wang W, Wang X, Wang X, Song Y,
Han Y, Zhang J and Zhao H: Protein analysis of atrial fibrosis via
label-free proteomics in chronic atrial fibrillation patients with
mitral valve disease. PLoS One. 8:e602102013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu H, Zhang W, Zhong M, Zhang G and Zhang
Y: Differential gene expression during atrial structural remodeling
in human left and right atrial appendages in atrial fibrillation.
Acta Biochim Biophys Sin (Shanghai). 43:535–541. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park JH, Lee JS, Ko YG, Lee SH, Lee BS,
Kang SM, Chang BC and Pak HN: Histological and biochemical
comparisons between right atrium and left atrium in patients with
mitral valvular atrial fibrillation. Korean Circ J. 44:233–242.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Embi AA, Scherlag BJ and Ritchey JW:
Glycogen and the propensity for atrial fibrillation: Intrinsic
anatomic differences in glycogen in the left and right atria in the
goat heart. N Am J Med Sci. 6:510–515. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
John B, Stiles MK, Kuklik P, Chandy ST,
Young GD, Mackenzie L, Szumowski L, Joseph G, Jose J, Worthley SG,
et al: Electrical remodelling of the left and right atria due to
rheumatic mitral stenosis. Eur Heart J. 29:2234–2243. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tai CT, Lo LW, Lin YJ and Chen SA:
Arrhythmogenic difference between the left and right atria in a
canine ventricular pacing-induced heart failure model of atrial
fibrillation. Pacing Clin Electrophysiol. 35:188–195. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li D, Zhang L, Kneller J and Nattel S:
Potential ionic mechanism for repolarization differences between
canine right and left atrium. Circ Res. 88:1168–1175. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen J, Mandapati R, Berenfeld O, Skanes
AC, Gray RA and Jalife J: Dynamics of wavelets and their role in
atrial fibrillation in the isolated sheep heart. Cardiovasc Res.
48:220–232. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu W, Zhou XW, Liu S, Hu K, Wang C, He Q
and Li M: Calpain-truncated CRMP-3 and −4 contribute to potassium
deprivation-induced apoptosis of cerebellar granule neurons.
Proteomics. 9:3712–3728. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pang J, Liu WP, Liu XP, Li LY, Fang YQ,
Sun QP, Liu SJ, Li MT, Su ZL and Gao X: Profiling protein markers
associated with lymph node metastasis in prostate cancer by
DIGE-based proteomics analysis. J Proteome Res. 9:216–226. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fu H, Li W, Liu Y, Lao Y, Liu W, Chen C,
Yu H, Lee NT, Chang DC, Li P, et al: Mitochondrial proteomic
analysis and characterization of the intracellular mechanisms of
bis(7)-tacrine in protecting against glutamate-induced
excitotoxicity in primary cultured neurons. J Proteome Res.
6:2435–2446. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jin S, Shen JN, Guo QC, Zhou JG, Wang J,
Huang G, Zou CY, Yin JQ, Liu SJ, Liu W, et al: 2-D DIGE and
MALDI-TOF-MS analysis of the serum proteome in human osteosarcoma.
Proteomics Clin Appl. 1:272–285. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tang ZX, Chen GX, Liang MY, Rong J, Yao
JP, Yang X and Wu ZK: Selective antegrade cerebral perfusion
attenuating the TLR4/NF-kB pathway during deep hypothermia
circulatory arrest in a pig model. Cardiology. 128:243–250.
2014.PubMed/NCBI
|
|
29
|
Zrenner B, Ndrepepa G, Karch MR, Schneider
MA, Schreieck J, Schömig A and Schmitt C: Electrophysiologic
characteristics of paroxysmal and chronic atrial fibrillation in
human right atrium. J Am Coll Cardiol. 38:1143–1149. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang JL, Lin YJ, Lee PC, Chang HY,
Hartono B, Lo LW, Chang SL, Hu YF, Suenari K, Li CH, et al: Right
atrial substrate remodeling and atrioventricular node conduction
properties in patients with paroxysmal atrial fibrillation. Int J
Cardiol. 158:447–449. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Casaclang-Verzosa G, Gersh BJ and Tsang
TS: Structural and functional remodeling of the left atrium:
Clinical and therapeutic implications for atrial fibrillation. J Am
Coll Cardiol. 51:1–11. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Matsuyama TA, Tanaka H, Adachi T, Jiang Y,
Ishibashi-Ueda H and Takamatsu T: Intrinsic left atrial
histoanatomy as the basis for reentrant excitation causing atrial
fibrillation/flutter in rats. Heart Rhythm. 10:1342–1348. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Calò L, Lamberti F, Loricchio ML, De Ruvo
E, Colivicchi F, Bianconi L, Pandozi C and Santini M: Left atrial
ablation versus biatrial ablation for persistent and permanent
atrial fibrillation: A prospective and randomized study. J Am Coll
Cardiol. 47:2504–2512. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim JB, Bang JH, Jung SH, Choo SJ, Chung
CH and Lee JW: Left atrial ablation versus biatrial ablation in the
surgical treatment of atrial fibrillation. Ann Thorac Surg.
92:1397–1404; discussion 1404–1405. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jiang Z, Ma N, Yin H, Ding F, Liu H and
Mei J: Biatrial ablation versus limited right atrial ablation for
atrial fibrillation associated with atrial septal defect in adults.
Surg Today. 45:858–863. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cooley N, Cowley MJ, Lin RC, Marasco S,
Wong C, Kaye DM, Dart AM and Woodcock EA: Influence of atrial
fibrillation on microRNA expression profiles in left and right
atria from patients with valvular heart disease. Physiol Genomics.
44:211–219. 2012. View Article : Google Scholar : PubMed/NCBI
|