Introduction
In recent years, the incidence of skeletal muscle
injury has increased. It is well known that skeletal muscles only
possess limited self-renewal capacity. Skeletal muscles respond to
mechanical load, and this mechanical stimulation has been reported
to promote myogenesis by provoking muscle precursor cell
activation, proliferation and differentiation (1). In the field of regenerative medicine,
appropriate mechanical stimulation has been applied to
muscle-derived stem cells in order to promote skeletal muscle
repair (2). Furthermore, it has
been suggested that external mechanical stimuli are transformed
into intracellular signals through certain mechanisms, in order to
regulate myoblast activity (3,4).
However, the mechanisms underlying the transformation of external
mechanical stimuli into intracellular signals remain unclear.
Nuclear factor (NF)-κB, which belongs to the Rel
family, refers to several transcription factors. The mammalian
NF-κB family comprises seven members, including p65 (RelA), c-Rel,
RelB, p50/p105 (NF-κB1) and p52/p100 (NF-κB2). NF-κB usually exists
in the form of hetero- or homodimers, and the typical NF-κB complex
is composed of a p50 and p65 heterodimer. In non-stimulated cells,
the NF-κB complex is located in the cytoplasm and is inhibited by
the inhibitor of NF-κB (I-κBα). Once stimulated, the degradation of
I-κBα and phosphorylation of NF-κB induce the translocation of
NF-κB into the nucleus where it binds to target DNA (5). The translocation of NF-κB has been
reported to regulate several cellular processes, including immunity
and inflammation, cellular proliferation and differentiation
(6–8). The role of NF-κB is indispensable and
complex due to its presence in both proliferation and
differentiation stages during the myogenic process (9). As for stretch-induced skeletal muscle
alterations, the NF-κB signaling pathway has been shown to be
activated in murine diaphragm muscles subjected to longitudinal
mechanical stretch, resulting in upregulated expression of the
stretch-response gene, Ankyrin repeat domain 2 (10). NF-κB activation by phosphorylation
of p65 NF-κB has also been detected during cyclic stretch-induced
myogenic differentiation (11). A
previous study revealed that NF-κB transcriptional activity is
beneficial to myoblast proliferation under appropriate stretch
induction (12).
MicroRNAs (miRNAs/miRs) are small non-coding RNAs,
some of which are highly evolutionarily conserved. miRNAs regulate
gene expression at the post-transcriptional level by inhibiting
translation of target mRNA or promoting target mRNA degradation
(13). miRNAs have been identified
as novel components in the gene regulatory network of myogenesis
(14). A previous study reported
that miRNAs are essential for the generation and maintenance of
skeletal muscles (15). In
particular, miRNAs specifically expressed in muscles, including
miR-1 and miR-206, are able to promote skeletal myoblast
differentiation (16,17). Conversely, another muscle-specific
miRNA, miR-133, serves an opposite role in myogenesis by
maintaining myoblasts in a proliferative state (16). Furthermore, local injection of
miR-1, miR-133 and miR-206 can upregulate myogenin and myogenic
differentiation 1 (MyoD) expression in injured skeletal muscles,
and thus accelerates muscle regeneration (18). miRNAs that are ubiquitously
expressed can also influence skeletal muscle development. miR-181
inhibits skeletal myogenesis, whereas miR-146a and miR-26a
positively regulate myogenesis (19–21).
However, interactions between the NF-κB signaling pathway and
miRNAs during mechanical stretch-induced myoblast proliferation and
differentiation remain unclear.
The present study subjected C2C12 myoblasts to
cyclic stretch to investigate the interactions between NF-κB
activation and miRNA expression during mechanical stretch-induced
myoblast proliferation. The study focused on the myoblast
proliferation process, since it represents the early stages of
muscle regeneration. The computer-controlled Flexcell system was
used to apply cyclic mechanical strain to C2C12 myoblasts. The
appropriate mechanical stretch conditions that could promote C2C12
myoblast proliferation were initially confirmed, and the miRNA
expression alterations during mechanical stretch-induced myoblast
proliferation were determined using high-throughout sequencing and
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) methods. In addition, the interactions between NF-κB
activation and miRNA expression were explored by treating
stretch-induced myoblasts with or without an NF-κB inhibitor.
Materials and methods
Cell culture
C2C12 mouse myoblast cells were purchased from
American Type Culture Collection (Manassas, VA, USA). The frozen
stock was thawed and maintained as monolayer cultures in Dulbecco's
modified Eagle's medium supplemented with 10% fetal bovine serum
(both Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100
U/ml penicillin and 1,000 U/ml streptomycin at 37°C in a humidified
atmosphere containing 5% CO2. The medium was changed
twice per week.
Mechanical stretch stimulation of
C2C12 cells
The model of in vitro stretching of C2C12
myoblasts was established using the computer-controlled vacuum
stretch system (FX-5000 Tension system; FlexCell International
Corporation, Burlington, NC, USA). Briefly, C2C12 cells were seeded
into type-I collagen-coated flexible-bottom six-well plates
(BioFlex collagen I plates; FlexCell International Corporation) at
a density of 1×105 cells/well in culture medium. The
cells in the flexible-bottom six-well plates were then incubated at
37°C in a humidified 5% CO2 atmosphere overnight.
Subsequently, C2C12 cells underwent cyclic strain (10% deformation)
at 0.125, 0.25 and 0.5 Hz for 2 h per day for 4 consecutive days.
Cells cultured under the same conditions without cyclic strain were
considered the control group.
Cell viability assay and cell cycle
analysis
C2C12 cell viability was assessed 1, 2, 3 and 4 days
after initiation of cyclic strain using the Cell Counting kit-8
(CCK-8) assay. Briefly, C2C12 cells in the control group, and
0.125, 0.25 and 0.5 Hz groups were digested with 0.25% pancreatic
enzyme (trypsin; Gibco; Thermo Fisher Scientific, Inc.) containing
0.02% EDTA. The cells were then reseeded in 96-well plates at a
density of 5×103 cells/well in culture medium (8
duplicate wells/group). The plates were incubated for 24 h at 37°C,
after which the cell medium was replaced with 100 µl culture medium
containing 10 µl CCK-8 reagent (Dojindo Molecular Technologies,
Inc., Kumamoto, Japan). Following a further incubation for 3 h at
37°C, the optical density (OD) values were measured at 450 nm using
a microplate reader (Thermo Fisher Scientific, Inc.). Cell cycle
analysis was performed by flow cytometry (FCM). Briefly, stretched
C2C12 cells (10% deformation; 0.125 Hz) at day 4 and control cells
were collected. Subsequently, the cells were permeabilized with 70%
(v/v) ethanol and incubated overnight at 4°C. Finally, the cells
were stained with 500 µl propidium iodide (100 µg/ml) at 37°C for
30 min and analyzed by FCM. The relative DNA proliferation index
(DPI) was calculated using the following formula: [DPI = (S% +
G2 / M%) / (S% + G2 / M% + G0 /
G1%)], according to our previous report (7).
High-throughput sequencing
High-throughput sequencing was applied to detect
differentially expressed miRNAs in C2C12 cells between the control
group and the 0.125 Hz stretch group at day 4. Total RNA was
extracted from the C2C12 cells using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Small RNAs (18–75 nt) were separated from
total RNAs by polyacrylamide gel electrophoresis and were purified.
The small RNAs were ligated with 5′ and 3′ adapters using T4 RNA
Ligase (Thermo Fisher Scientific, Inc.), and were reverse
transcribed to cDNA using SuperScript II (Life Technologies; Thermo
Fisher Scientific, Inc.). Generation of double-stranded cDNA was
performed prior to PCR amplification. High-throughput sequencing of
the purified DNA samples was performed using the Illumina Genome
Analyzer IIx (Illumina, San Diego, CA, USA). To select
differentially expressed miRNAs between the two groups, the
frequency of miRNAs was normalized to calculate the ratio of 0.125
Hz stretched cells to control cells. A differentially expressed
miRNA was indicated by a stretched/control ratio >2 and a
statistically significant result, as determined by independent
t-test with Bonferroni correction.
NF-κB inhibition
Two selective and irreversible NF-κB inhibitors:
BAY11-7082 (BAY; Beyotime Institute of Biotechnology, Jiangsu,
China) and pyrrolidine dithiocarbamate (PDTC; Beyotime Institute of
Biotechnology) were used in the present study. In order to
determine the effects of NF-κB inhibition on the stretched
myoblasts, C2C12 cells that underwent 0.125 Hz cyclic strain (10%
deformation) for 2 h per day for 4 consecutive days were
simultaneously treated with 2.5 µM BAY, 5 µM BAY or 10 µM PDTC. The
effects of NF-κB inhibition on stretched C2C12 myoblasts were
subsequently detected according to the aforementioned CCK-8 method
and FCM analysis at corresponding time points.
Western blot analysis
Cells were lysed with radioimmunoprecipitation assay
buffer (EMD Millipore, Billerica, MA, USA) and protein
concentration was determined using the bicinchoninic acid assay kit
(Thermo, Fisher Scientific, Inc.). Protein samples (50 µg) were
separated by SDS-PAGE (5% stacking gel, 10% separating gel), and
were then transferred to polyvinylidene fluoride (PVDF) membranes
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). The membranes were
then rinsed with TBS-0.05% Tween-20 (TBST) and blocked with 5%
fat-free dried milk solution at room temperature for 1.5 h. Primary
antibodies: Rabbit anti-NF-κB (ab16502; 1:2,000), rabbit
anti-phosphorylated (P)-NF-κB (ab86299; 1:1,000), mouse anti-I-κBα
(ab211340; 1:1,000) and mouse anti-GAPDH (ab8245; 1:2,000) (all
Abcam, Cambridge, MA, USA) were added, and the membranes were
incubated overnight at 4°C. Subsequently, the PVDF membranes were
rinsed 3 times with TBST (5 min/wash) and were incubated with
horseradish peroxidase-conjugated goat anti-mouse IgG (ab97023;
1:3,000 and 1:5,000 for anti-I-κBα and anti-GAPDH primary
antibodies, respectively) and goat anti-rabbit IgG (ab6721; 1:5,000
and 1:3,000 for anti-NF-κB and anti-P-NF-κB primary antibodies,
respectively) secondary antibodies from Abcam, prior to
visualization by enhanced chemiluminescence. Experiments were
repeated three times. Relative expression levels were determined
using ImageJ software, version 2.1.4.7 (National Institutes of
Health, Bethesda, MD, USA) and were normalized to loading
controls.
RT-qPCR
Total RNA was extracted from C2C12 cells in the
control group, 0.125 Hz stretch group and NF-κB inhibitor + 0.125
Hz stretch group at day 4 using TRIzol® according to the
manufacturer's protocol. RT was performed using an ABI 9700 PCR
amplification machine (Applied Biosystems; Thermo Fisher
Scientific, Inc.) and a miR RT kit (Takara Bio, Inc., Otsu, Japan).
Briefly, aliquots (2 µg) of total RNA were transcribed into cDNA in
a total volume of 20 µl with 5X PrimeScript buffer, 1 mmol/l
nucleotide mixture, 50 pmol/l primer, 100 U PrimeScript Reverse
Transcriptase, and 20 U RNase inhibitor. The RT reaction was
conducted at 37°C for 15 min, 85°C for 5 sec and 4°C prior to qPCR
analysis. qPCR was performed in triplicate using a PCR detection
system (Stratagene Mx3000P qPCR system; Agilent Technologies, Santa
Clara, CA, USA). miRNA specific primer sequences are listed in
Table I. All primers were
synthesized by Guangzhou Vipotion Biotechnology Co., Ltd.
(Guangzhou, China). qPCR was conducted using the Taq PCR kit
(Takara Bio, Inc.) in a 20 µl reaction volume containing 2 µl miRNA
RT product, 0.2 µl 1 U/µl Taq DNA polymerase, 10 µl 5X PCR
Buffer, 0.6 µl 5 µM primer and deionized water. Reaction conditions
were as follows: Initial denaturation at 95°C for 3 min, 40 cycles
of 95°C for 12 sec and 62°C for 60 sec, followed by an extension
step at 72°C for 1 min. The relative miRNA expression levels were
calculated using the comparative threshold cycle 2−ΔΔCq
method (22).
 | Table I.miR specific primer sequences. |
Table I.
miR specific primer sequences.
| Primer name | Sequence
(5′-3′) |
|---|
| mmu-mir-500 |
AAUGCACCUGGGCAAGGGUUCA |
| mmu-mir-1934 |
AGGAUGACGGUGGGGCUGGUGA |
| mmu-mir-31 |
AGGCAAGAUGCUGGCAUAGCUG |
| mmu-mir-378a |
ACUGGACUUGGAGUCAGAAGG |
| mmu-mir-331 |
GCCCCUGGGCCUAUCCUAGAA |
| mmu-mir-5097 |
GUUCAUGUCCCUGUUCAGGCGCCA |
| mmu-mir-1941 |
AGGGAGAUGCUGGUACAGAGGCUU |
Statistical analysis
Data were analyzed by one-way analysis of variance
and the post-hoc Bonferroni test was used for pair-wise
comparisons. Statistical analyses were performed using SPSS 19.0
(IBM SPSS, Armonk, NY, USA) and the data were presented as the mean
± standard deviation, error bars were representative of three
independent experiments. P<0.05 was considered to indicate a
statistically significant difference.
Results
Alterations to the miRNA expression
profile during mechanical stretch-induced myoblast
proliferation
To investigate the effects of various stretch
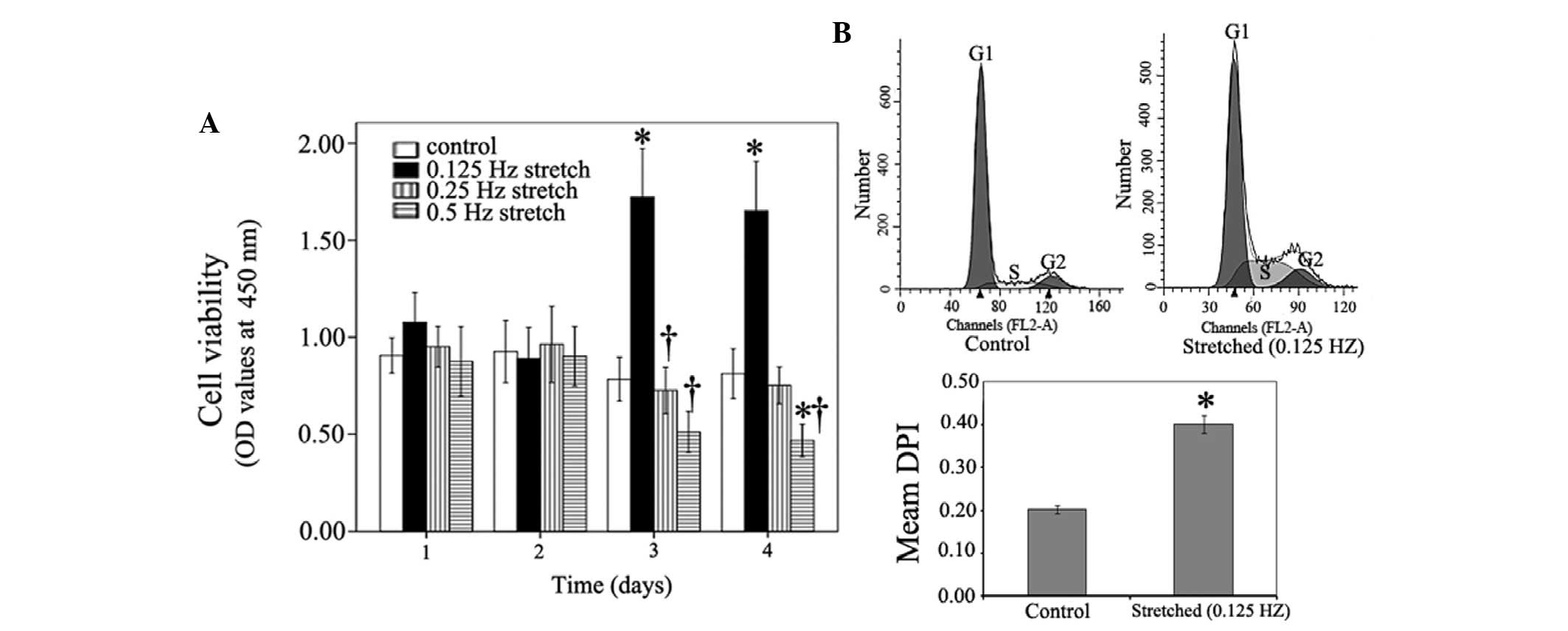
frequencies on myoblast viability, the CCK-8 assay was performed 1,
2, 3 and 4 days after the cells underwent cyclic stretching. At day
3, the mean OD value of the 0.125 Hz stretch group (10%
deformation) was significantly higher compared with in the control
group; 1.65 vs. 0.71 respectively (P<0.05; Fig. 1A). At day 4, the mean OD value of
the 0.125 Hz stretched cells remained higher compared with the
control cells; 1.63 vs. 0.81 (P<0.05). Furthermore, cell cycle
progression differed between the control cells and the 0.125 Hz
stretched myoblasts (10% deformation) at day 4. As shown in
Fig. 1B, the number of cells in
the proliferative (S or G2) phase was markedly higher in
the 0.125 Hz stretch group compared with in the control group. At
day 4, the mean DPI in the 0.125 Hz stretch group was significantly
increased compared with in the control cells (Fig. 1B). These results suggest that 10%
deformation cyclic strain at 0.125 Hz can enhance myoblast
viability and stimulate myoblast entry into the proliferative
phase, thus promoting C2C12 cell proliferation. Conversely, 10%
deformation cyclic strain at 0.5 Hz markedly inhibited C2C12 cell
viability at day 4 compared with the control (Fig. 1A). Therefore, 10% deformation
cyclic mechanical stretch at 0.125 Hz were considered the
appropriate stretch conditions.
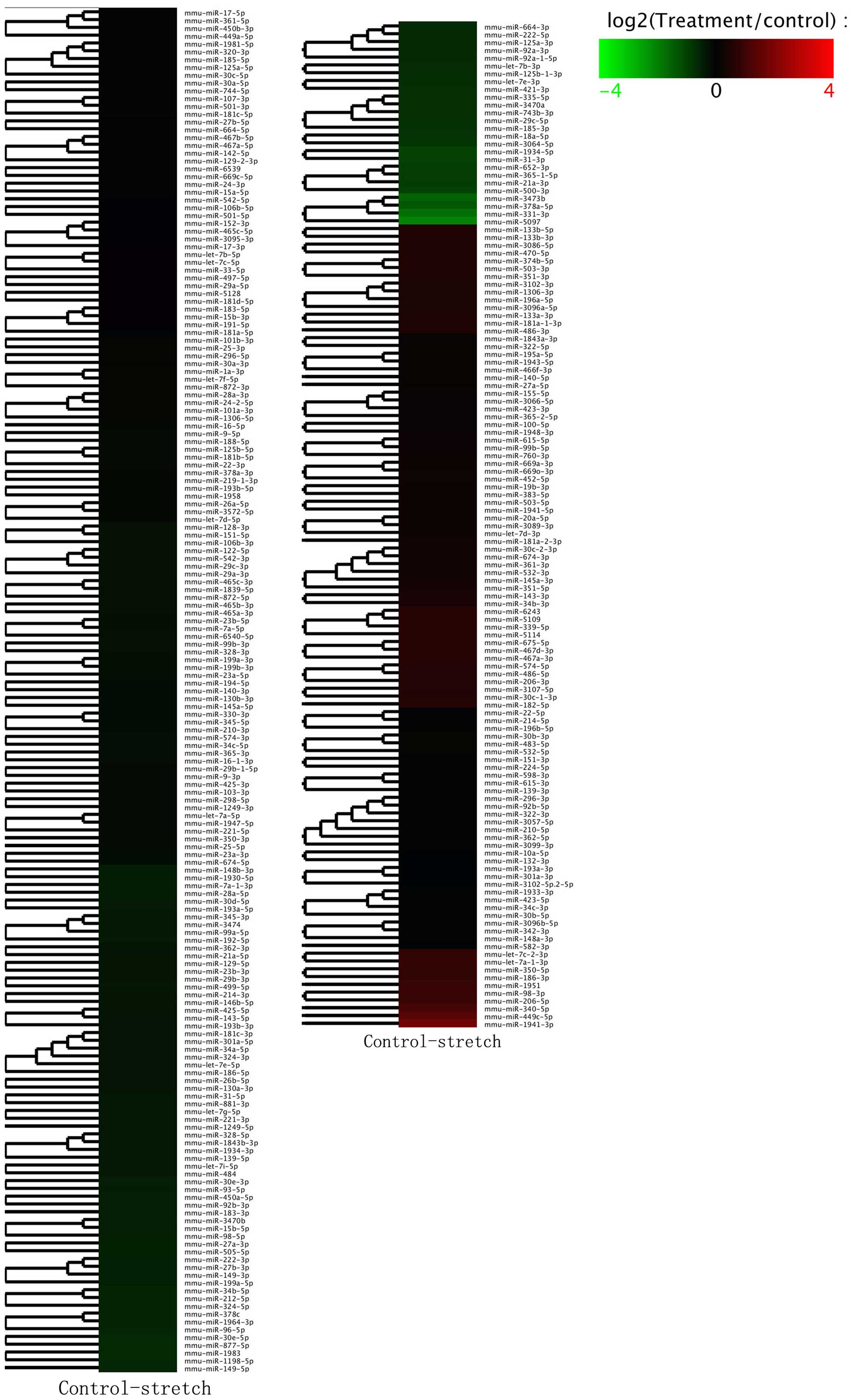
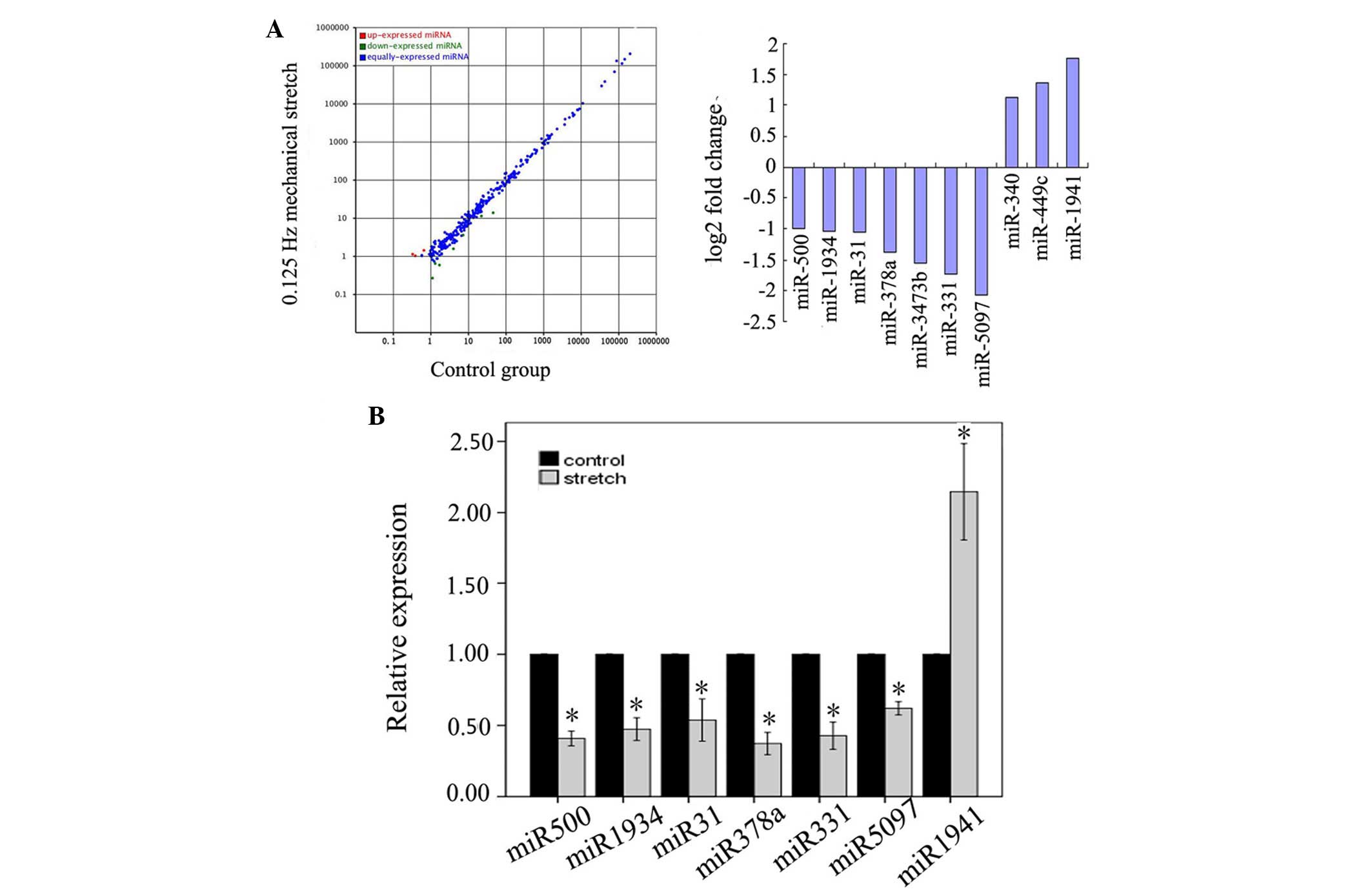
Alterations to the miRNA profile between the control
group and the 0.125 Hz mechanical stretch group were determined by
high-throughput sequencing. The hierarchical clustering analysis of
differentially expressed miRNAs is presented in Fig. 2. Within the miRNA expression
profiles, 10 miRNAs exhibited >2-fold variation in the 0.125 Hz
mechanical stretch group compared with in the control group. Seven
(miR-500, −1934, −31, −378a, −3473b, −331 and −5097) of which were
downregulated, whereas three (miR-340, −449c and −1941) were
upregulated in the mechanical stretch group (Fig. 3A). Among them, miR-500, miR-1934,
miR-31, miR-378a, miR-331, miR-5097 and miR-1941, were validated
through the RT-qPCR method (Fig.
3B).
Differentially expressed miRNAs
induced by cyclic mechanical stretch are reversed by NF-κB
inhibition in C2C12 myoblasts
The present study aimed to explore the association
between NF-κB activation and miRNA expression during
stretch-induced myoblast proliferation. As shown in Fig. 4A, 0.125 Hz mechanical stretch
increased the expression levels of NF-κB p65 and P-NF-κB p65,
whereas the expression levels of the inhibitory factor I-κBα were
decreased compared with the control group (P<0.05). However,
treatment with 2.5 or 5 µM NF-κB inhibitor BAY inhibited NF-κB p65
and P-NF-κB p65 expression, and increased the expression levels of
I-κBα compared with the stretch group. Treatment with 5 µM BAY
resulted in a more significant inhibition of NF-κB activation
(P<0.05). These results suggest that NF-κB is activated in the
process of stretch-mediated myoblast proliferation, and the NF-κB
inhibitor BAY can effectively inhibit activation of NF-κB p65 in
stretched C2C12 cells.
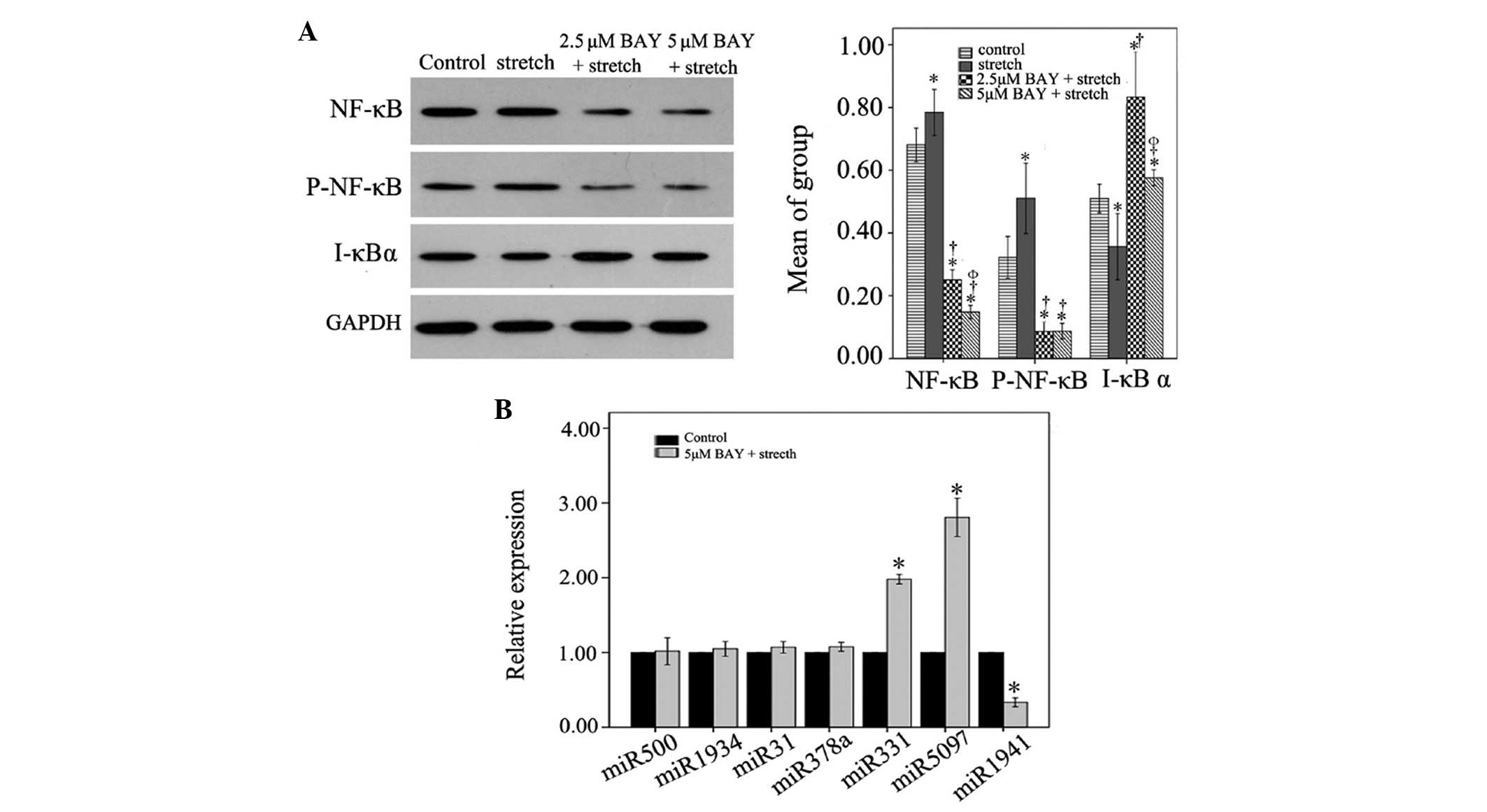 | Figure 4.(A) Effects of NF-κB inhibition on
the expression levels of NF-κB p65, P-NF-κB p65, and I-κBα proteins
in 0.125 Hz stretched C2C12 cells. *P<0.05 vs. the control
group. †P<0.05 vs. the stretch group.
ΦP<0.05 vs. the 2.5 µM BAY group. n=3. (B) Relative
expression levels of miR-500, miR-1934, miR-31, miR-378, miR-331,
miR-5097 and miR-1941 in control and 5 µM BAY + mechanical stretch
groups. Expression levels in the 5 µM Bay + 0.125 Hz mechanical
stretch group were normalized to the control group. *P<0.05 vs.
the control group. n=3. miR, microRNA; NF-κB, nuclear factor-κB;
P-, phosphorylated; I-κBα, inhibitor of NF-κB; BAY, BAY11-7082. |
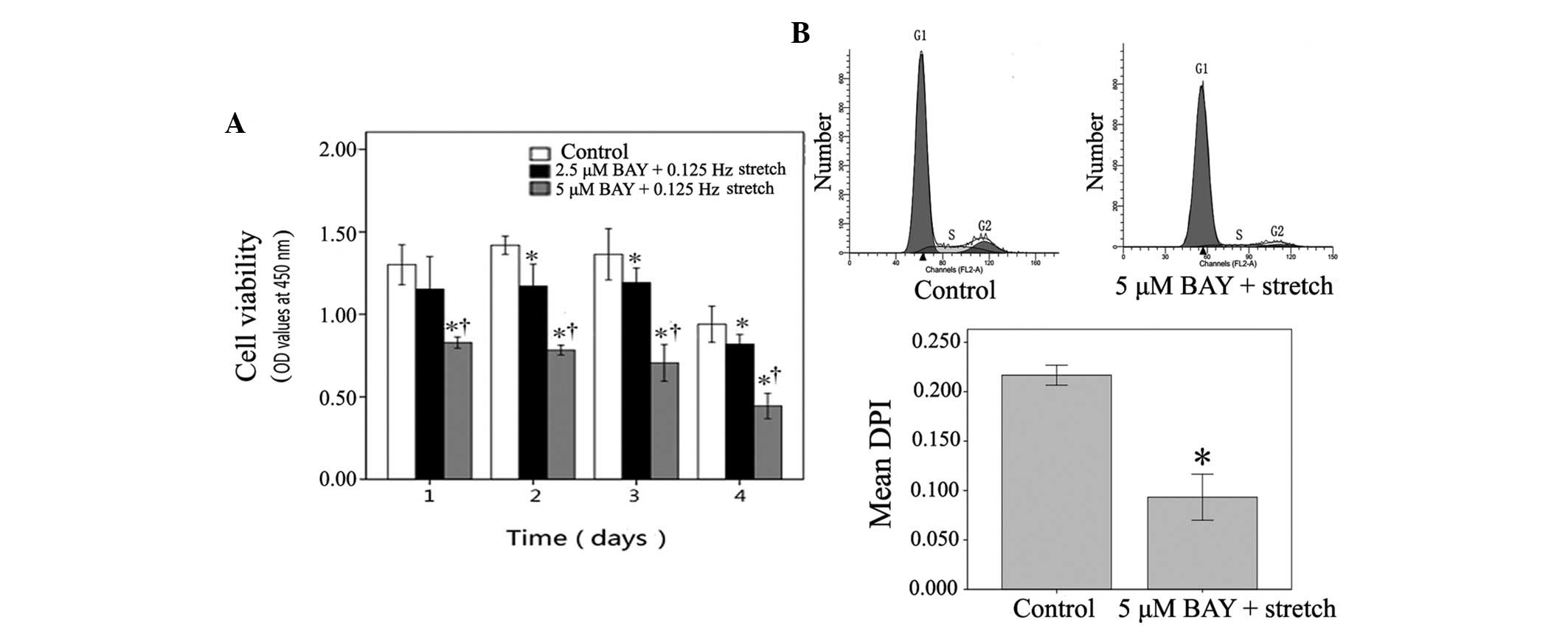
Compared with 2.5 µM BAY, 5 µM BAY displayed an
increased inhibitory effect on NF-κB activation. Therefore, 5 µM
BAY was selected as the dose for NF-κB inhibition in subsequent
experiments regarding miRNA expression. As shown in Fig. 4B, 0.125 Hz stretch-induced
downregulation of miR-500, miR-1934, miR-31 and miR-378a in C2C12
cells was reversed by 5 µM BAY. Furthermore, 0.125 Hz
stretch-induced reduced expression of miR-331 and miR-5097 was
significantly reversed by 5 µM BAY (P<0.05). Conversely, the
0.125 Hz mechanical stretch-induced expression of miR-1941 was
significantly reduced following treatment with 5 µM BAY
(P<0.05). These data indicate that 5 µM Bay can reverse
mechanical stretch-induced miRNA expression alterations.
The effects of the NF-κB inhibitor BAY were also
determined on mechanical stretch-induced myoblast proliferation, as
assessed by CCK-8 assay and FCM. The 0.125 Hz mechanical
stretch-induced increased OD value was significantly decreased in
C2C12 cells following treatment with 2.5 µM BAY on days 2, 3 and 4.
Treatment with 5 µM BAY was able to inhibit stretched myoblast
viability on the first day, and sustain the inhibitory effect
between days 2 and 4 (P<0.05; Fig.
5A). FCM revealed that stretched C2C12 cells treated with 5 µM
BAY were arrested in G1 phase at day 4, and the mean DPI
in stretched C2C12 cells treated with 5 µM BAY was significantly
decreased compared with the control cells (P<0.05; Fig. 5B). These results suggest that the
NF-κB inhibitor BAY may inhibit stretch-induced myoblast
proliferation.
The present study used another NF-κB inhibitor,
PDTC, to confirm the effects of NF-κB inhibition on stretched
myoblasts. The results, as shown in Fig. 6, revealed that 10 µM PDTC elicited
similar effects to BAY. Treatment of C2C12 cells with 10 µM PDTC
inhibited activation of NF-κB, reversed alterations in
stretch-induced miRNA expression and inhibited stretch-induced
myoblast proliferation.
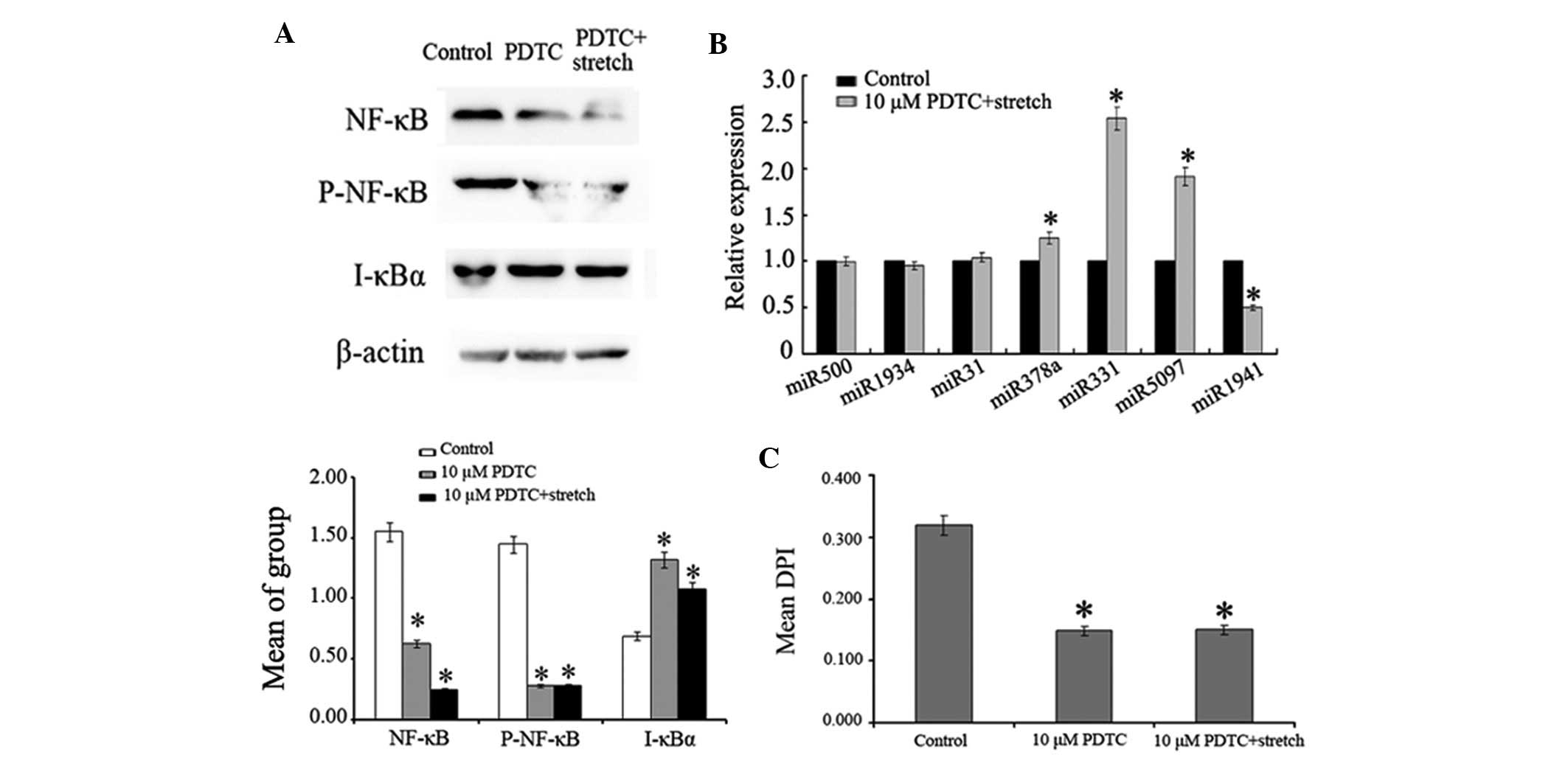 | Figure 6.(A) Effects of 10 µM PDTC on the
expression levels of NF-κB p65, P-NF-κB p65 and I-κBα proteins in
0.125 Hz stretched C2C12 cells, as detected by western blotting.
(B) Relative expression levels of miR-500, miR-1934, miR-31,
miR-378, miR-331, miR-5097 and miR-1941 in control and 10 µM PDTC +
mechanical stretch groups. (C) Effects of 10 µM PDTC on 0.125 Hz
stretch-induced myoblast proliferation. *P<0.05 vs. the control
group. n=3. miR, microRNA; NF-κB, nuclear factor-κB; P-,
phosphorylated; I-κBα, inhibitor of NF-κB; PDTC, pyrrolidine
dithiocarbamate; DPI, DNA proliferation index. |
Discussion
Variations in the duration, magnitude and frequency
of mechanical stretch have discrepant influences on myoblasts.
Appropriate external mechanical stimulation can promote the
proliferation of myoblasts cultured in vitro (23,24).
In addition, it has previously been reported that mechanical
stretch (15% strain) may induce conspicuous injury to muscle cells
(25), whereas cyclic tensile
stretch (10% strain) at 1 Hz for 1 h followed by 23 h of relaxation
could enhance myoblast proliferation (24). The results of the present study
revealed that only 0.125 Hz mechanical stretch could promote
myoblast proliferation when C2C12 myoblasts were exposed to 10%
tensile cyclic strain at various frequencies (0.125, 0.25 and 0.5
Hz). Similar findings have been reported in human bladder smooth
muscle cells cultured in vitro; 5% equibiaxial stretch at
0.1 Hz promoted smooth muscle cell proliferation and decreased
apoptotic rate (25).
In the present study, seven mechanical
stretch-associated miRNAs were detected in C2C12 cells: MiR-500,
miR-1934, miR-31, miR-378a, miR-331, miR-5097 and miR-1941, which
were identified using high-throughput sequencing and RT-qPCR
analysis. Among them, miR-500 has previously been detected in the
dorsal root ganglia of rats and is thought to be involved in nerve
reconstruction (26). Furthermore,
miR-31 is able to promote the expression of myogenic factor 5 and
myoblast proliferation. Targeted inhibition of miR-31 has been
reported to be an effective therapeutic strategy for dystrophin
rescue in human Duchenne muscular dystrophy (27,28).
In addition, miR-378 may suppress proliferation of myoblasts by
modulating the mitogen-activated protein kinases (MAPK) pathway and
bone morphogenetic proteins (29,30).
The overexpression of miR-378 promotes myoblast differentiation by
strengthening MyoD activity (30).
Increased expression of miR-331 has also been detected during
myoblast differentiation (31).
These findings are similar to the results of the present study; the
expression levels of miR-31, miR-378 and miR-331 were reduced
during the process of 10% strain, 0.125 Hz mechanical
stretch-induced myoblast proliferation. In addition, to the best of
our knowledge, miR-1941, miR-1934, miR-5097 and miR-500 may be
considered novel candidates for stretch-induced myogenesis, which
have not been reported previously.
The present study further explored the effects of
NF-κB, which participates in the enhanced proliferative process
elicited by mechanical stretch. There are several signaling
pathways associated with mechanical stretch in skeletal muscle
cells, including protein kinase C (32), MAPK (33), phosphatidylinositol-3 kinase/Akt
(34), NF-κB (12), calcineurin (35) and nitric oxide synthase (36). However, findings published over the
past few years have suggested that these signaling pathways have a
close connection and serve crosstalk roles in inflammatory and
immune responses (37–39). Notably, MAPK appears to be an
upstream regulator of the NF-κB signaling pathway, which can
successfully induce NF-κB-dependent myogenesis (8–11).
In the present study, the NF-κB signaling pathway was activated by
mechanical stretch, as determined by the markedly increased protein
expression levels of NF-κB p65 and P-NF-κB p65. Following treatment
with NF-κB inhibitors, translocation of NF-κB in stretch-loaded
myoblasts was inhibited, which was accompanied by the reversal of
miRNA expression and the suppression of proliferation in C2C12
myoblasts. Therefore, these results indicated that the NF-κB
signaling pathway is activated and exerts direct effects in
stretch-induced myoblasts. These results are concordant with the
findings reported by Kumar et al, which indicated that NF-κB
is indispensable for stretch-induced proliferation of C2C12 cells
(12).
The results of the present study identified a link
between NF-κB and miRNA expression. The expression levels of
miR-500, miR-1934, miR-31, miR-378a, miR-331 and miR-5097 were
downregulated in myoblasts following mechanical stretch, whereas
NF-κB inhibition increased the expression of these miRNAs.
Conversely, the upregulation of miR-1941 was markedly reduced by
NF-κB inhibition in mechanically stretched C2C12 cells. Similarly,
bioinformatics analysis in a previous study suggested the presence
of an NF-κB binding element in the promoter region of miR-26a;
inhibition of NF-κB in cardiac fibroblasts increased miR-26a
expression, whereas overexpression of miR-26a inhibited NF-κB
activity, thus indicating a feedback loop in cardiac fibroblasts
(40). The results of the present
study are the first, to the best of our knowledge, to reveal that
miRNA expression is NF-κB-dependent during stretch-induced myoblast
proliferation, thus suggesting a possible mechanism regarding the
transformation of external mechanical stimuli into intracellular
signals.
The present study used two structurally different
NF-κB inhibitors in order to confirm the effects of the NF-κB
signaling pathway on stretched myoblasts. BAY is a specific
inhibitor of NF-κB, due to its inhibitory effect on the activation
of NF-κB and the phosphorylation of I-κBα (41). PDTC is able to inhibit activation
of NF-κB specifically by suppressing the release of the inhibitory
subunit I-κB from the latent cytoplasmic form of NF-κB (42–44).
The results of the present study revealed that the two NF-κB
inhibitors were able to inhibit activation of NF-κB, reverse
stretch-induced miRNA profile alterations in C2C12 myoblasts, and
suppress stretch-induced myoblast proliferation. Accordingly, these
data confirmed the interaction between miRNAs and the NF-κB
signaling pathway during stretch-induced myogenesis.
In conclusion, cyclic stretch (10% deformation;
0.125 Hz) was able to activate the NF-κB signaling pathway, reduce
the expression levels of miR-500, −1934, −31, −378, −331 and −5097,
increase the expression of miR-1941, and promote proliferation of
C2C12 cells. However, treatment with NF-κB inhibitors inhibited
NF-κB activation in mechanically stretched C2C12 cells, which was
accompanied by reversed expression of miRNAs and suppressed cell
proliferation. These results demonstrated that cyclic
stretch-mediated cell proliferation and miRNA profile alterations
in C2C12 myoblasts occur via activation of the NF-κB signaling
pathway, thus providing a possible mechanism during stretch-induced
myogenesis.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 31100700) and the
Guangdong Medical Research Foundation (grant no. A2015412).
References
|
1
|
Tidball JG: Mechanical signal transduction
in skeletal muscle growth and adaptation. J Appl Physiol (1985).
98:1900–1908. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Beckman SA, Chen WC, Tang Y, Proto JD,
Mlakar L, Wang B and Huard J: Beneficial effect of mechanical
stimulation on the regenerative potential of muscle-derived stem
cells is lost by inhibiting vascular endothelial growth factor.
Arterioscler Thromb Vasc Biol. 33:2004–2012. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Klumpp D, Horch RE, Kneser U and Beier JP:
Engineering skeletal muscle tissue-new perspectives in vitro and in
vivo. J Cell Mol Med. 14:2622–2629. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ross TD, Coon BG, Yun S, Baeyens N, Tanaka
K, Ouyang M and Schwartz MA: Integrins in mechanotransduction. Curr
Opin Cell Biol. 25:613–618. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kurihara Y and Furue M: Interferon-gamma
enhances phorbol myristate acetate-induced cell attachment and
tumor necrosis factor production via the NF-kB pathway in THP-1
human monocytic cells. Mol Med Rep. 7:1739–1744. 2013.PubMed/NCBI
|
|
6
|
Karin M and Ben-Neriah Y: Phosphorylation
meets ubiquitination: The control of NF-[kappa]B activity. Annu Rev
Immunol. 18:621–663. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang LY, Wang HY, Ouyang J, Yu L, Chen B,
Qin JQ and Qiu XZ: Low concentration of lipopolysaccharide acts on
MC3T3-E1 osteoblasts and induces proliferation via the
COX-2-independent NFkappaB pathway. Cell Biochem Funct. 27:238–242.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baeza-Raja B and Muñoz-Cánoves P: p38
MAPK-induced nuclear factor-kappaB activity is required for
skeletal muscle differentiation: Role of interleukin-6. Mol Biol
Cell. 15:2013–2026. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bakkar N and Guttridge DC: NF-kappaB
signaling: A tale of two pathways in skeletal myogenesis. Physiol
Rev. 90:495–511. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mohamed JS, Lopez MA, Cox GA and Boriek
AM: Anisotropic regulation of Ankrd2 gene expression in skeletal
muscle by mechanical stretch. FASEB J. 24:3330–3340. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ji G, Liu D, Liu J, Gao H, Yuan X and Shen
G: p38 mitogen-activated protein kinase up-regulates NF-kappaB
transcriptional activation through RelA phosphorylation during
stretch-induced myogenesis. Biochem Biophys Res Commun.
391:547–551. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kumar A, Murphy R, Robinson P, Wei L and
Boriek AM: Cyclic mechanical strain inhibits skeletal myogenesis
through activation of focal adhesion kinase, Rac-1 GTPase, and
NF-kappaB transcription factor. FASEB J. 18:1524–1535. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang K, Xu Z, Wang N, Tian Y, Sun X and Ma
Y: Analysis of microRNA and gene networks in human chronic
myelogenous leukemia. Mol Med Rep. 13:453–460. 2016.PubMed/NCBI
|
|
14
|
Wang H, Sun H and Guttridge DC: microRNAs:
Novel components in a muscle gene regulatory network. Cell Cycle.
8:1833–1837. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
O'Rourke JR, Georges SA, Seay HR, Tapscott
SJ, McManus MT, Goldhamer DJ, Swanson MS and Harfe BD: Essential
role for Dicer during skeletal muscle development. Dev Biol.
311:359–368. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen JF, Mandel EM, Thomson JM, Wu Q,
Callis TE, Hammond SM, Conlon FL and Wang DZ: The role of
microRNA-1 and microRNA-133 in skeletal muscle proliferation and
differentiation. Nat Genet. 38:228–233. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lu J, McKinsey TA, Zhang CL and Olson EN:
Regulation of skeletal myogenesis by association of the MEF2
transcription factor with class II histone deacetylases. Mol Cell.
6:233–244. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nakasa T, Ishikawa M, Shi M, Shibuya H,
Adachi N and Ochi M: Acceleration of muscle regeneration by local
injection of muscle-specific microRNAs in rat skeletal muscle
injury model. J Cell Mol Med. 14:2495–2505. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yamamoto M and Kuroiwa A: Hoxa-11 and
Hoxa-13 are involved in repression of MyoD during limb muscle
development. Dev Growth Differ. 45:485–498. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kuang W, Tan J, Duan Y, Duan J, Wang W,
Jin F, Jin Z, Yuan X and Liu Y: Cyclic stretch induced miR-146a
upregulation delays C2C12 myogenic differentiation through
inhibition of Numb. Biochem Biophys Res Commun. 378:259–263. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wong CF and Tellam RL: MicroRNA-26a
targets the histone methyltransferase Enhancer of Zeste homolog 2
during myogenesis. J Biol Chem. 283:9836–9843. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Otis JS, Burkholder TJ and Pavlath GK:
Stretch-induced myoblast proliferation is dependent on the COX2
pathway. Exp Cell Res. 310:417–425. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kook SH, Lee HJ, Chung WT, Hwang IH, Lee
SA, Kim BS and Lee JC: Cyclic mechanical stretch stimulates the
proliferation of C2C12 myoblasts and inhibits their differentiation
via prolonged activation of p38 MAPK. Mol Cells. 25:479–486.
2008.PubMed/NCBI
|
|
25
|
Wazir R, Luo DY, Dai Y, Yue X, Tian Y and
Wang KJ: Expression and proliferation profiles of PKC, JNK and
p38MAPK in physiologically stretched human bladder smooth muscle
cells. Biochem Biophys Res Commun. 438:479–482. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu B, Zhou S, Qian T, Wang Y, Ding F and
Gu X: Altered microRNA expression following sciatic nerve resection
in dorsal root ganglia of rats. Acta Biochim Biophys Sin
(Shanghai). 43:909–915. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Crist CG, Montarras D and Buckingham M:
Muscle satellite cells are primed for myogenesis but maintain
quiescence with sequestration of Myf5 mRNA targeted by microRNA-31
in mRNP granules. Cell Stem Cell. 11:118–126. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cacchiarelli D, Incitti T, Martone J,
Cesana M, Cazzella V, Santini T, Sthandier O and Bozzoni I: miR-31
modulates dystrophin expression: New implications for Duchenne
muscular dystrophy therapy. EMBO Rep. 12:136–141. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hou X, Tang Z, Liu H, Wang N, Ju H and Li
K: Discovery of MicroRNAs associated with myogenesis by deep
sequencing of serial developmental skeletal muscles in pigs. PLoS
One. 7:e521232012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jeon TI, Park JW, Ahn J, Jung CH and Ha
TY: Fisetin protects against hepatosteatosis in mice by inhibiting
miR-378. Mol Nutr Food Res. 57:1931–1937. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dmitriev P, Barat A, Polesskaya A,
O'Connell MJ, Robert T, Dessen P, Walsh TA, Lazar V, Turki A,
Carnac G, et al: Simultaneous miRNA and mRNA transcriptome
profiling of human myoblasts reveals a novel set of myogenic
differentiation-associated miRNAs and their target genes. BMC
Genomics. 14:2652013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nobe K and Paul RJ: Distinct pathways of
Ca(2+) sensitization in porcine coronary artery: Effects of
Rho-related kinase and protein kinase C inhibition on force and
intracellular Ca(2+). Circ Res. 88:1283–1290. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Martineau LC and Gardiner PF: Insight into
skeletal muscle mechanotransduction: MAPK activation is
quantitatively related to tension. J Appl Physiol (1985).
91:693–702. 2001.PubMed/NCBI
|
|
34
|
Sakamoto K, Aschenbach WG, Hirshman MF and
Goodyear LJ: Akt signaling in skeletal muscle: Regulation by
exercise and passive stretch. Am J Physiol Endocrinol Metab.
285:E1081–E1088. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bassel-Duby R and Olson EN: Role of
calcineurin in striated muscle: Development, adaptation, and
disease. Biochem Biophys Res Commun. 311:1133–1141. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tidball JG, Spencer MJ, Wehling M and
Lavergne E: Nitric-oxide synthase is a mechanical signal transducer
that modulates talin and vinculin expression. J Biol Chem.
274:33155–33160. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Parola C, Salogni L, Vaira X, Scutera S,
Somma P, Salvi V, Musso T, Tabbia G, Bardessono M, Pasquali C, et
al: Selective activation of human dendritic cells by OM-85 through
a NF-kB and MAPK dependent pathway. PLoS One. 8:e828672013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hu H, Li Z, Zhu X, Lin R and Chen L:
Salidroside reduces cell mobility via NF-k B and MAPK signaling in
LPS-Induced BV2 microglial cells. Evid Based Complement Alternat
Med. 2014:3838212014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hong GE, Kim JA, Nagappan A, Yumnam S, Lee
HJ, Kim EH, Lee WS, Shin SC, Park HS and Kim GS: Flavonoids
Identified from Korean Scutellaria baicalensis Georgi inhibit
inflammatory signaling by suppressing activation of NF-kB and MAPK
in RAW 264.7 cells. Evid Based Complement Alternat Med.
2013:9120312013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wei C, Kim IK, Kumar S, Jayasinghe S, Hong
N, Castoldi G, Catalucci D, Jones WK and Gupta S: NF-kB mediated
miR-26a regulation in cardiac fibrosis. J Cell Physiol.
228:1433–1442. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Moore LB, Sawyer AJ, Charokopos A, Skokos
EA and Kyriakides TR: Loss of monocyte chemoattractant protein-1
alters macrophage polarization and reduces NFkB activation in the
foreign body response. Acta Biomater. 11:37–47. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Guo RM, Xu WM, Lin JC, Mo LQ, Hua XX, Chen
PX, Wu K, Zheng DD and Feng JQ: Activation of the p38 MAPK/NF-kB
pathway contributes to doxorubicin-induced inflammation and
cytotoxicity in H9c2 cardiac cells. Mol Med Rep. 8:603–608.
2013.PubMed/NCBI
|
|
43
|
Huang T, Gao D, Jiang X, Hu S, Zhang L and
Fei Z: Resveratrol inhibits oxygen-glucose deprivation-induced
MMP-3 expression and cell apoptosis in primary cortical cells via
the NF-kB pathway. Mol Med Rep. 10:1065–1071. 2014.PubMed/NCBI
|
|
44
|
Li T, Zhang Q, Zhang J, Yang G, Shao Z,
Luo J, Fan M, Ni C, Wu Z and Hu X: Fenofibrate induces apoptosis of
triple-negative breast cancer cells via activation of NF-kB
pathway. BMC Cancer. 14:962014. View Article : Google Scholar : PubMed/NCBI
|