Introduction
Hypertrophic scar (HS) is a fibroproliferative
disorder of the skin characterized by aberrant wound healing and
pathological, excessive dermal fibrosis (1). Skin damage, as occurs with burns,
activates fibroblasts and causes them to produce extracellular
matrix proteins, including collagen I and III, and growth factors,
including transforming growth factor-β and insulin-like growth
factor I. Excessive extracellular matrix formation combined with
inadequate remodeling of scar tissue results in an aesthetically
and functionally unsatisfactory, painful, pruritic scar that may
impair skin functions. Current advances in molecular biology and
genetics have led to novel non-surgical preventive and therapeutic
options, however none of these treatments are efficient (2). Thus, it is important to further
elucidate the pathophysiology of HS and to identify potential
targets for the development of an effective strategy for HS
prevention and treatment.
MicroRNAs (miRNAs) are small noncoding RNAs of 19–25
nucleotides. miRNAs regulate gene expression at the
post-transcriptional level by degradation or translational
repression of a target mRNA via binding 3′untranslated regions
(3′-UTR) of target genes. It has been estimated that up to 90% of
human genes are regulated by miRNAs and >3,700 miRNAs have been
identified in human cells (3,4).
miRNAs are involved in the regulation of the majority, if not all,
molecular and cellular processes, and aberrant expression of miRNAs
is associated with the initiation and progression of numerous
pathological disorders, including cancer, cardiovascular disease
and multiple fibrotic diseases (5,6).
However, the full miRNA expression signature of HS has not been
previously identified and the functions of miRNAs in the pathology
of HS remain poorly understood. An in-depth understanding of the
specific miRNAs that are involved in the pathogenesis of HS may
provide important novel targets for the prevention and treatment of
HS.
The present study aimed to identify potential
therapeutic targets for the clinical treatment of HS. Specifically,
the current study determined miRNA expression associated with HS by
miRNA microarray analysis. Of the miRNAs differentially expressed
between HS and control tissues, miRNA-21 (miR-21) was significantly
upregulated in vitro and in vivo. Therefore, the
present study focused on investigating the function of miR-21 in HS
and its potential as a target for the prevention and treatment of
HS.
Materials and methods
Patient specimens and primary cell
culture
Hypertrophic scar fibroblasts (HSFBs) were
established as primary cell lines from HS tissues obtained from
patients recovering from severe burns. A total 22 patients (13 male
and 9 female; age range, 17–42) were recruited in the Department of
Plastic and Aesthetic Surgery, Southwest Hospital (Chongqing,
China) between 2014 and 2015. The scars were all in a proliferative
phase and there was no local infection or ulceration present. None
of the cases had been previously treated with glucocorticosteroids
or radiotherapy. NS specimens were obtained from donor sites during
scar resection. The experimental protocols were approved by the
ethics committee of the Third Military Medical University
(Chongqing, China) and written informed consent was obtained from
each patient and donor. NS and HS tissues were cut into a size of
~1×1×0.5 cm and incubated in a solution of collagenase type I (0.5
mg/ml) and trypsin (0.2 mg/ml) at 37°C for 6 h. Primary fibroblasts
were cultured in Dulbecco's modified Eagle medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA). HSFBs from each donor
were used at passages 3–6 in the experiments. When required, the
chemically synthesized miR-21 antagomir
(5′-UAGCUUAUCAGACUGAUGUUGA-3′) and antagomir control
5′-UCACAACCUCCUAGAAAGAGUAGA-3′; Guangzhou RiboBio Co., Ltd.,
(Guangzhou, China) were diluted to final concentrations of 20 nM in
cell culture media to disturb miR-21 expression. Cells were
incubated with miRNA antagomir for 6 h at 37°C prior to changing
the medium. Following a further 48 h incubation, the cells were
used for further experiments.
Microarray analysis
miRNAs were isolated from HS and corresponding
normal skin (NS) tissues using the mirVana miRNA Isolation kit
(Ambion; Thermo Fisher Scientific, Inc.). Total RNA was quantified
using the NanoDrop 2000 (Thermo Fisher Scientific, Inc.) and RNA
integrity was assessed using the Agilent Bioanalyzer 2100 (Agilent
Technologies, Inc., Santa Clara, CA, USA). Each group consisted of
three samples and miRNA expression profiling was performed by miRNA
microarray analysis using a chip containing 2158 mature miRNAs
(Human miRNA Microarray Slide, Design ID: 046064; Agilent
Technologies, Inc.). Proprietary ‘spike-in’ controls were used at
each step of the process. Feature Extraction software (version no.
10.7.1.1; Agilent Technologies, Inc.) was used to analyze array
images to obtain raw data. GeneSpring GX software (version no.
12.5; Agilent Technologies, Inc.) was employed to perform basic
analysis of the raw data. Using this software, the raw data was
normalized with the quantile algorithm. Hierarchical clustering was
performed to highlight distinguishable miRNA expression patterns
within samples.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) for collagen type I
α1chain (COL1A1), collagen type I α 2 chain (COL1A2), fibronectin
(Fn) and miR-21 analysis. RT-qPCR was performed to detect miRNA
expression according to the manufacturer's protocol using the
TaqMan® microRNA assay kit (Applied Biosystems; Foster
City, CA). Reactions were performed using Stratagene Mx3000P
instrument (Agilent Technologies, Inc.) in triplicate. A total of
ten pairs of HS/NS samples obtained from the patients, and all the
samples obtained from the hypertrophic scar model of rabbit ears
were used in the qPCR. Relative miRNA expression was calculated
using the 2-ΔΔCq method (7) and levels were normalized to the
expression of U6 small nuclear RNAs. The sequences of the primers
were as follows: Forward, 5′-TCTAGACATGTTCAGCTTTGTGGAC-3′, and
reverse, 5′-TCTGTACGCAGGTGATTGGTG-3′ for human COL1A1; forward,
5′-GAGGGCAACAGCAGGTTCACTTA-3′ and reverse,
5′-TCAGCACCACCGATGTCCA-3′ for human COL1A2; forward,
5′-ACCCAGTCCCGAAGGCACTA-3′ and reverse,
5′-CCTGTACATCTAAAGGCATGAAGCA-3′ for human fibronectin; forward,
5′-GCCCGCTAGCTTATCAGACTGATG-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′
for miR-21; and forward, 5′-GCGCGTCGTGAAGCGTTC-3′ and reverse,
5′-GTGCAGGGTCCGAGGT-3′ for U6.
In situ hybridization (ISH)
ISH was performed as previously described (8). To determine the location of miR-21,
an antisense locked nucleic acid (LNA)-modified oligonucleotide
probe was used (Exiqon A/S, Vedbaek, Denmark). LNA/DNA oligos
contained locked nucleic acids at eight consecutive centrally
located bases and had the following sequence: LNA-miR-21,
5′-TCAACATCAGTCTGATAAGCTA-3′.
Cell proliferation and apoptosis
assay
Fibroblasts were transfected with the miR-21
antagomir or antagomir control (Guangzhou RiboBio Co., Ltd.)
according to the manufacturer's protocols. Fibroblasts were
initially seeded at 50–60% confluence and treated for 6 h at 37°C
with miR-21 antagomir or antagomir control at a final concentration
of 20 nM. The transfected cells were then prepared as a single-cell
suspension. A total of 1×104 cells were seeded into 96-well plates
and cultured for 5 days. At designated time points, 20 µl 0.5% MTT
was added to each well and incubated for 4 h prior to measuring the
absorbance of the plates at 570 nm using a microplate reader (Model
680; Bio-Rad Laboratories, Inc., Hercules, CA, USA). The Annexin
V-FITC Apoptosis Detection kit (eBioscience, Inc., San Diego, CA,
USA) was used to analyze apoptosis. Apoptotic cells were then
analyzed by a FACSAria flow cytometer (BD Biosciences, Franklin
Lakes, NJ, USA) using CellQuest software version 5.1 (BD
Biosciences).
Collagen synthesis analysis by
H3-proline incorporation assay
Cells at the logarithmic phase (5×104/ml) were
incubated in 96-well plates for 48 h at 37°C, with 200 µl of medium
in each well., A 100 µl mixture of H3-proline (5 µCi/ml; Yahui Co.,
Beijing, China), vitamin C (25 µg/ml) and β-aminopropionitrile (50
µg/ml; Yahui Co.) was added and cells were then cultured for
another 10 h. The supernatants were then discarded, and cells were
digested with trypsin for 2 min, washed and collected onto a glass
fiber filter (0.5×104/well). Following precipitation and fixation
with 10% trichloroacetic acid for 20 min at −4°C and absolute
ethanol, the filter was dried at 70°Cin an oven and placed in a
scintillation bottle containing 5 ml of scintillation fluid. A
Beckman LS6500 scintillation counter (Beckman Coulter Inc., Brea,
CA, USA) was used to count γ-rays. The results were expressed as
counts per min.
Hypertrophic scar model of rabbit ear
and drug administration
Adult male New Zealand white rabbits (n=8) weighing
between 2.2 and 2.5 kg (age, 5–7 months) were used in this study.
All animals were obtained from the Laboratory Animal Center of the
Third Military Medical University (Chongqing, China), and housed
alone at 15–25°C, with a 12-h light/dark cycle and access to food
and water provided ad libitum. The HS model was produced following
the standardized protocol as previously described (9). The animals were anaesthetized with
intramuscular ketamine (40 mg/kg) and xylazine (4 mg/kg). Cleaning
of the ears was conducted with iodine and no local anesthesia was
used. Four wounds, reaching down to bare cartilage on the ventral
surface of each ear, were created by means of a 7 mm punch biopsy.
Experiments were conducted in accordance with the Guidelines for
the Care and Use of Laboratory Animals of Third Military Medicine
University, and the experimental protocols used in the present
study were approved by the Animal Care Committee of TMMU.
The rabbits were randomly assigned to either the
intervention group or the control group (n=4 in each group).
Chemically synthesized antagomir or antagomir control (RiboBio Co.,
Ltd.) were used to inhibit miR-21 expression. Experimental wounds
were injected, under the new epithelium, with 100 µl solution
containing 10 nM miR-21 antagomir or antagomir control. The
injections were carried out on days 15 and 25 after wounding. On
day 65 post-wounding, rabbits in each group were anaesthetized with
intramuscular ketamine (40 mg/kg) and xylazine (4 mg/kg) and then
scars were harvested. One half of each scar was fixed in 4%
paraformaldehyde for 24 h, embedded in paraffin, cut in 4-mm
sections, and stained with hematoxylin and eosin. The thickness of
the epidermis in uninjured skin and of the epidermis of the scar
was measured at ×400 magnification under optical microscope
(Olympus Corporation, Tokyo, Japan). The other half was frozen for
RT-qPCR analysis, which was performed according to the
aforementioned description. The degree of dermal hypertrophy of
each scar was expressed as the scar elevation index (SEI). This
index is the ratio of the area of newly formed dermis of the scar
to the area of surrounding normal dermis. An SEI value greater than
1 was considered to indicate a hypertrophic dermis.
Statistical analysis
All values are presented as the mean ± standard
deviation of three independent experiments. Statistical analysis
was performed using analysis of variance, Chi-squared test or
paired Student's t-test using SPSS17.0 (SPSS, Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
miRNA expression profiling of HS
To determine whether miRNAs are involved in the
development of HS, miRNA expression profiling was performed on HS
and matched NS tissues (n=3). A total of 2158 miRNAs were screened
using an Agilent Human miRNA Bioarray (Human miRNA Microarray
Slide, Design ID 046064; Agilent Technologies, Inc.). The
expression of certain miRNA was significantly different between the
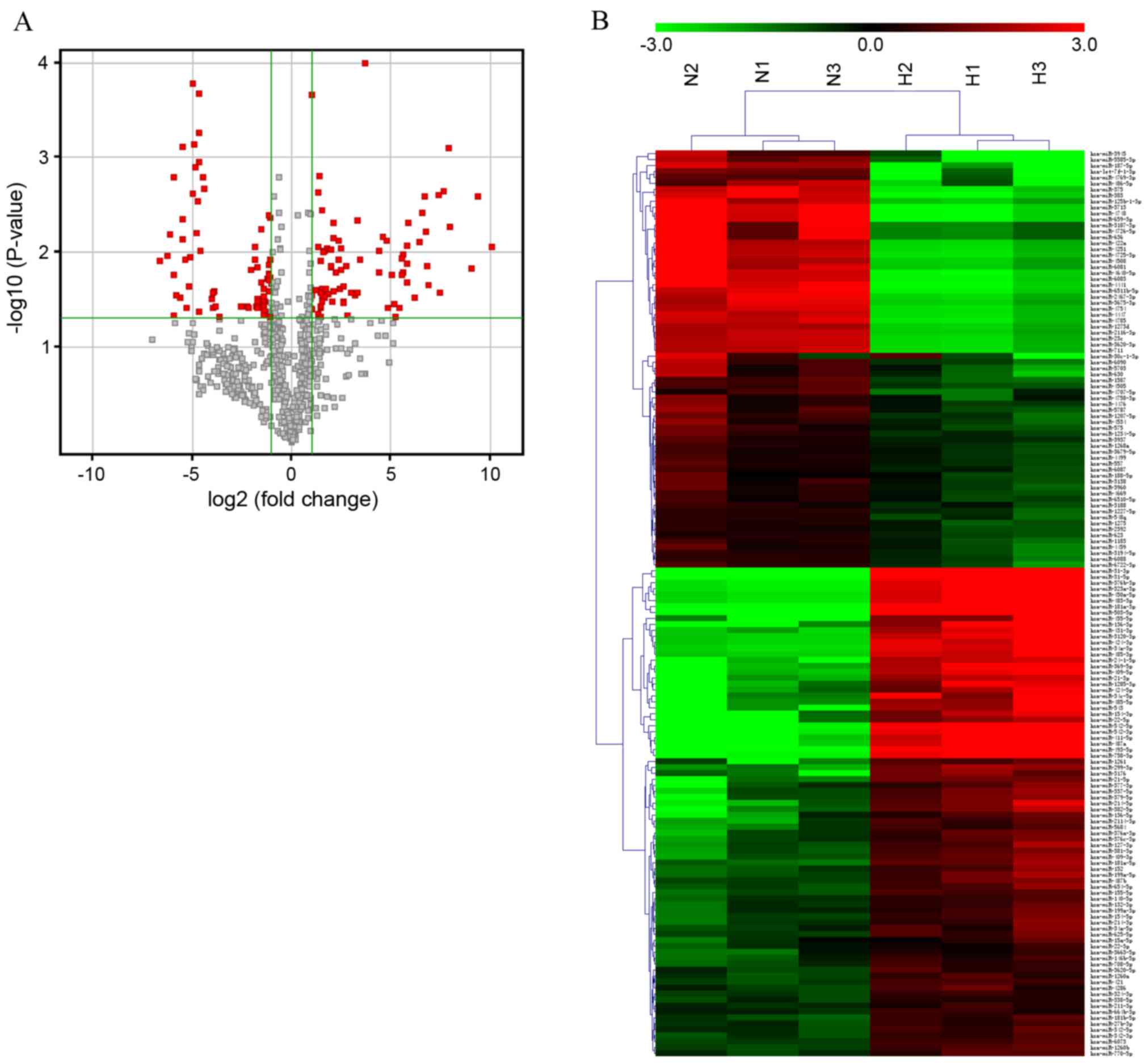
two groups (Fig. 1A). The raw data
was normalized with the quantile algorithm. In any conditions, the
probes flagged as ‘Detected’ in at least 100% of samples were
selected for further data analysis. Differentially expressed miRNAs
were identified through fold change (FC) and the calculation of
P-values was performed using Student's t-test. The threshold set
for up and downregulated genes was an FC ≥2 and P<0.05. Target
genes of the miRNAs identified as differentially expressed were
predicted using the following databases: Targetscan (www.targetscan.org), microRNAorg (http://www.microrna.org) and the PITA algorithm
(https://genie.weizmann.ac.il/pubs/mir07/mir07_prediction.html).
Analysis identified a total of 152 aberrantly
expressed miRNAs in HS, including 82 miRNAs that were upregulated
and 70 miRNAs that were downregulated. Hierarchical clustering
involves unsupervised sample clustering of candidate genes,
obtained by appropriate analysis and filtering, to identify novel
clusters that changed in a similar direction in the development of
HS. Differentially expressed miRNAs were used to generate a heat
map, and HS and NS clusters were clearly visible (Fig. 1B). Table I lists the 40 miRNAs with the
greatest differences in expression between the HS and NS groups. Of
the dysregulated miRNAs identified, miR-31-5p was the most
upregulated and miR-3713 was the most downregulated miRNA in HS
compared with NS tissue, with FC values of 1090.30 and −98.72,
respectively. Dysregulated miRNAs were grouped based on their
absolute FC values as follows: 12 miRNAs with an FC >100, 57
miRNAs between 10 and 100, 14 miRNAs between 5 and 10, and 69
miRNAs between 2 and 5. Given that miRNAs are important in numerous
pathophysiological processes, the data suggest that these specific
miRNAs may be implicated in the development of HS.
 | Table I.Top 40 up and downregulated miRs
identified by microarray analysis. |
Table I.
Top 40 up and downregulated miRs
identified by microarray analysis.
| Upregulated miRs |
|---|
|
|---|
| miR name | FC (abs) | P-value |
|---|
| Upregulated |
|
|
|
hsa-miR-31-5p | 1090.30 | 0.009 |
|
hsa-miR-503-5p | 654.82 | 0.003 |
|
hsa-miR-31-3p | 532.56 | 0.015 |
|
hsa-miR-542-5p | 248.59 | 0.005 |
|
hsa-miR-181a-3p | 244.34 | 0.001 |
|
hsa-miR-542-3p | 202.18 | 0.002 |
|
hsa-miR-483-3p | 177.35 | 0.026 |
|
hsa-miR-493-5p | 165.60 | 0.003 |
|
hsa-miR-450a-5p | 118.70 | 0.022 |
|
hsa-miR-323a-3p | 112.27 | 0.014 |
|
hsa-miR-487a | 108.29 | 0.006 |
|
hsa-miR-758-3p | 104.53 | 0.003 |
|
hsa-miR-411-5p | 96.54 | 0.004 |
|
hsa-miR-376b-3p | 87.80 | 0.008 |
|
hsa-miR-154-3p | 72.48 | 0.030 |
|
hsa-miR-369-5p | 57.18 | 0.005 |
|
hsa-miR-409-5p | 56.06 | 0.009 |
|
hsa-miR-34a-3p | 55.91 | 0.013 |
|
hsa-miR-3120-3p | 55.88 | 0.021 |
|
hsa-miR-24-1-5p | 48.98 | 0.016 |
|
hsa-miR-136-3p | 48.82 | 0.011 |
|
hsa-miR-485-3p | 46.73 | 0.018 |
|
hsa-miR-424-3p | 45.83 | 0.012 |
|
hsa-miR-22-5p | 42.09 | 0.038 |
|
hsa-miR-455-5p | 37.82 | 0.049 |
|
hsa-miR-543 | 35.33 | 0.036 |
|
hsa-miR-431-3p | 32.87 | 0.017 |
|
hsa-miR-34c-5p | 29.35 | 0.039 |
|
hsa-miR-1285-3p | 27.86 | 0.008 |
|
hsa-miR-21-3p | 24.41 | 0.007 |
|
hsa-miR-485-5p | 21.43 | 0.016 |
|
hsa-miR-424-5p | 21.37 | 0.010 |
|
hsa-miR-214-5p | 12.94 | 0.000 |
|
hsa-miR-382-5p | 10.87 | 0.012 |
|
hsa-miR-299-3p | 10.15 | 0.005 |
|
hsa-miR-3176 | 9.80 | 0.027 |
|
hsa-miR-21-5p | 9.31 | 0.029 |
|
hsa-miR-379-5p | 8.03 | 0.027 |
|
hsa-miR-1261 | 7.14 | 0.047 |
|
hsa-miR-337-5p | 6.57 | 0.014 |
| Downregulated |
|
|
|
hsa-miR-3713 | 98.72 | 0.012 |
|
hsa-miR-659-3p | 77.69 | 0.011 |
|
hsa-miR-4748 | 69.06 | 0.007 |
|
hsa-miR-4441 | 61.18 | 0.002 |
|
hsa-miR-5585-3p | 60.28 | 0.047 |
|
hsa-miR-383 | 60.08 | 0.017 |
|
hsa-miR-375 | 55.28 | 0.029 |
|
hsa-miR-187-5p | 49.09 | 0.030 |
|
hsa-miR-6511b-5p | 45.33 | 0.001 |
|
hsa-miR-6083 | 45.16 | 0.007 |
|
hsa-miR-4640-5p | 44.61 | 0.005 |
|
hsa-miR-4725-3p | 40.22 | 0.012 |
|
hsa-miR-125b-1-3p | 40.07 | 0.038 |
|
hsa-miR-422a | 35.22 | 0.023 |
|
hsa-miR-4251 | 34.83 | 0.011 |
|
hsa-miR-4785 | 32.05 | 0.000 |
|
hsa-miR-2467-3p | 31.71 | 0.002 |
|
hsa-miR-4754 | 29.63 | 0.001 |
|
hsa-miR-4447 | 28.50 | 0.001 |
|
hsa-miR-6081 | 27.06 | 0.006 |
|
hsa-miR-3675-3p | 26.78 | 0.003 |
|
hsa-miR-711 | 25.55 | 0.000 |
|
hsa-miR-23c | 25.25 | 0.001 |
|
hsa-miR-3620-3p | 25.08 | 0.001 |
|
hsa-miR-3945 | 24.74 | 0.043 |
|
hsa-miR-4508 | 24.33 | 0.010 |
|
hsa-miR-2116-3p | 21.99 | 0.002 |
|
hsa-miR-1273d | 20.75 | 0.002 |
|
hsa-miR-636 | 16.16 | 0.031 |
|
hsa-let-7f-1-3p | 15.26 | 0.038 |
|
hsa-miR-4726-5p | 15.17 | 0.027 |
|
hsa-miR-3187-3p | 15.14 | 0.026 |
|
hsa-miR-486-5p | 14.49 | 0.037 |
|
hsa-miR-4769-3p | 12.68 | 0.048 |
|
hsa-miR-630 | 5.74 | 0.038 |
|
hsa-miR-5703 | 4.69 | 0.038 |
|
hsa-miR-6090 | 4.47 | 0.039 |
|
hsa-miR-1587 | 4.13 | 0.015 |
|
hsa-miR-30c-1-3p | 3.67 | 0.009 |
|
hsa-miR-4707-5p | 3.54 | 0.012 |
miR-21 expression is upregulated in HS
tissues and HSFBs
Numerous differentially expressed miRNAs, which may
be associated with HS development, were identified by the
preliminary expression profiling study (Table I). Of those identified, miR-21,
also termed miR-21-5p, was upregulated in HS tissue compared with
NS tissue and was selected for further analysis, because a previous
study had investigated the function of phosphatase and tensin
homolog (PTEN) in hypertrophic scar (10) and miR-21 was demonstrated to
directly target PTEN (11).
Additionally, existing evidence suggested that miR-21 promotes
fibrosis in other diseases (12,13).
An miR-21 antagomir may have the potential to be used
therapeutically in HS, which, to the best of our knowledge, has not
been previously investigated. Therefore, the present study selected
miR-21 for investigation as a therapeutic target in HS.
In order to further validate the functions of
miR-21in the pathogenesis and progression of HS, RT-qPCR analysis
of miR-21 expression in HS and NS tissue samples was performed in
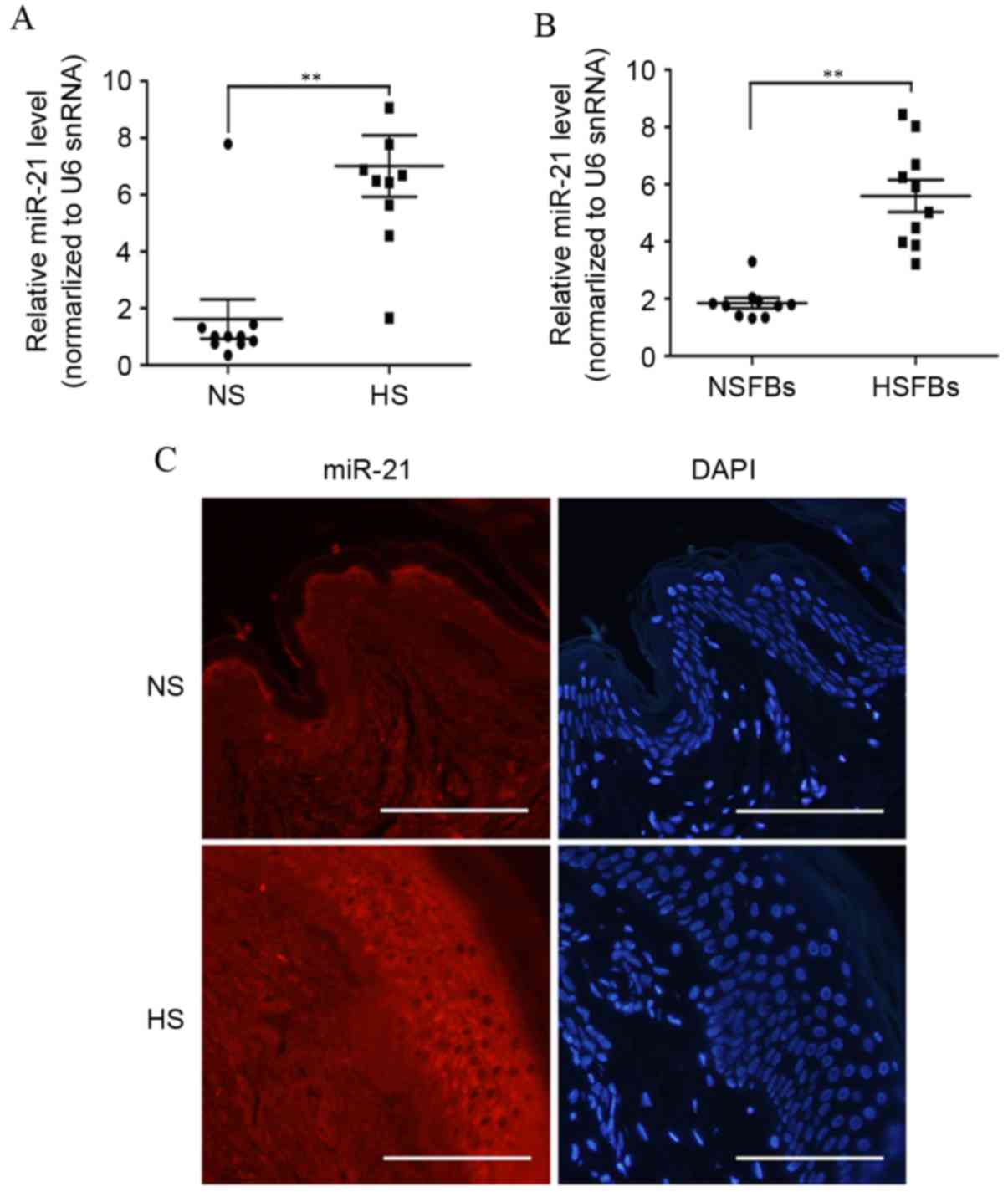
more patients. As demonstrated in Fig.
2A, the level of miR-21 expression was significantly higher in
10 cases of HS than in NS tissues (P=0.0019). As fibroblasts and
myofibroblasts are important in HS development (1), the present study further investigated
the expression of miR-21 in 10 samples of NSFBs (normal skin
fibroblasts) and HSFBs. As demonstrated in Fig. 2B, miR-21was significantly
overexpressed in HSFBs compared with NSFBs (P<0.0001). These
data support the suggestion that miR-21 may act as a pro-fibrotic
factor in HS.
ISH with an miR-21specific probe was then performed
to examine the expression of miR-21 in HS and NS tissues (Fig. 2C). ISH of skin tissues highlighted
extensive cytoplasmic staining for miR-21, which was localized to
epithelial cells and subdermal fibroblasts. The expression of
miR-21 in HS tissues was higher compared with NS tissues. Together,
these data demonstrated that miR-21 was overexpressed in HS,
suggesting that miR-21 may participate in the pathogenesis of
hypertrophic scar development.
miR-21 antagomir inhibits the
proliferation of HSFBs
HSFBs are important for the production and
deposition of collagen and are the primary effectors in HS
formation. To further investigate the potential for miR-21 to be a
therapeutic target for HS treatment, HSFBs were transfected with
miR-21 antagomir for 48 h andmiR-21 expression levels were
detected. An miRNA antagomir control was used as a control. Levels
of miR-21 in anti-miR-21-transfected HSFBs were significantly
decreased compared with the antagomir control group (Fig. 3A, P<0.0001). The present study
then assessed growth and apoptosis in HSFBs and NSFBs following
transfection of the miR-21 antagomir. Initially, the MTT assay was
used to evaluate the effect of miR-21 on cell growth. The results
demonstrated that the miR-21antagomir significantly decreased the
proliferation of HSFBs to a nearly normal level (P<0.01;
Fig. 3B), suggesting that
suppression of miR-21 with an antagomir inhibited HSFB growth. The
differences were most significant at day 4, compared with the HSFBs
transfected with the antagomir control. Flow cytometric analysis of
annexin V and propidium iodide stained cells was performed to
assess whether decreased viability was a result of induction of
cell apoptosis. Annexin V-positive early apoptotic cells were
increased in HSFBs transfected with miR-21antagomir compared with
the antagomir control group (Fig. 3C
and D). These results indicate that miR-21 affects cell
proliferation and apoptosis, and suggests that miR-21 may function
as an anti-apoptotic factor in the development of HS.
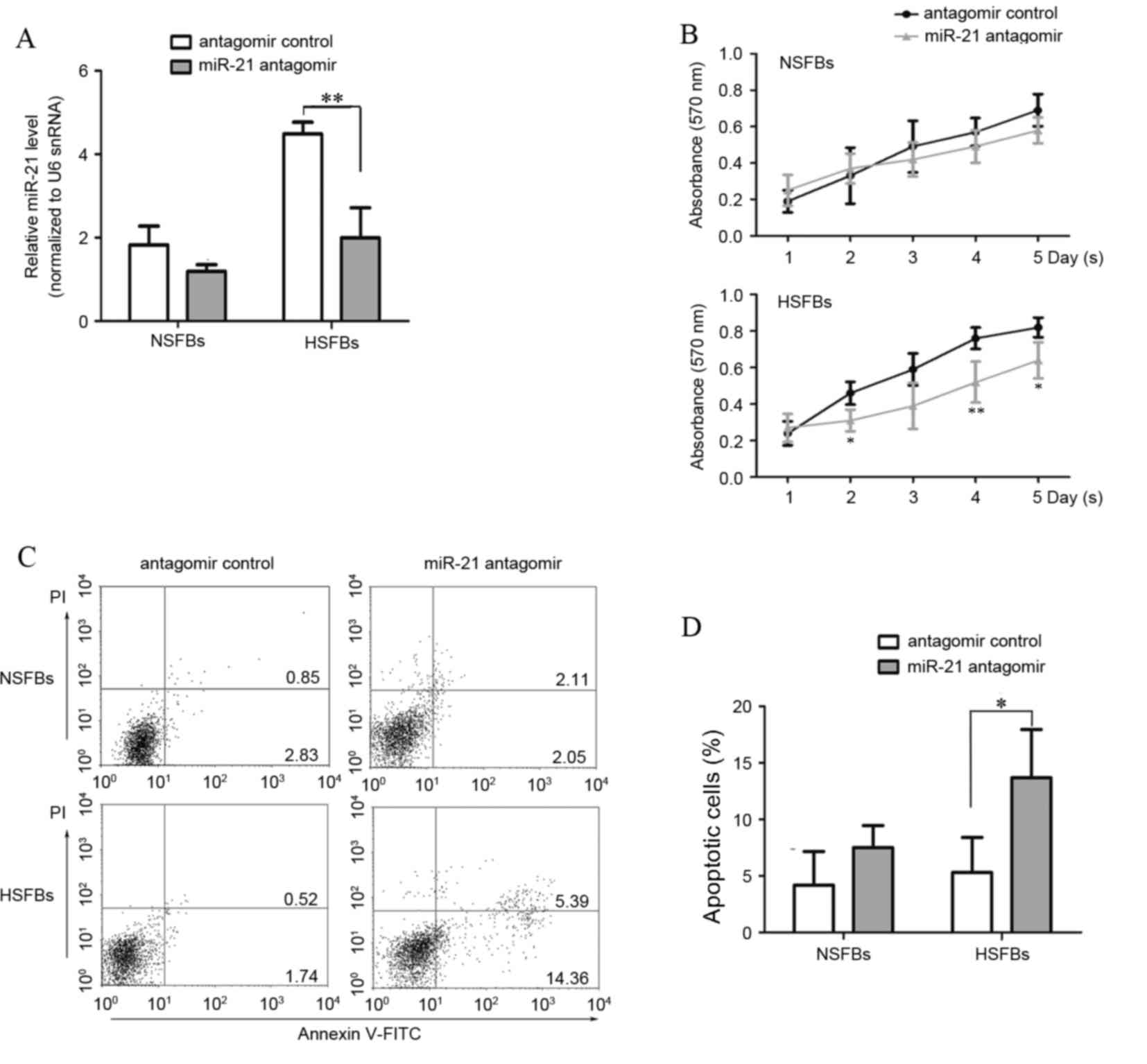 | Figure 3.miR-21 antagomir inhibits
proliferation and induces apoptosis in HSFBs. HSFBs and NSFBs were
treated with miR-21 antagomir or miR-21 antagomir control for 48 h.
(A) At 48 h after transfection, miR-21 expression levels were
evaluated by reverse transcription-quantitative polymerase chain
reaction analysis. (B) MTT assays were performed at 24 h intervals
for five days. *P<0.05, **P<0.01 vs. the antagomir control.
(C) Apoptosis was measured by flow cytometric analysis using
Annexin V and PI staining. Right lower quadrant, early apoptotic
cells; right upper quadrant, late apoptotic cells. (D) The rate of
apoptosis in different groups was calculated and compared. miR,
microRNA; HSFBs, hypertrophic scar fibroblasts; NSFBs, normal skin
fibroblasts; PI, propidium iodide; snRNA, small nuclear RNA.
*P<0.05, **P<0.01. |
miR-21 antagomir decreases the
expression of fibrosis-associated genes and collagen synthesis in
HSFBs
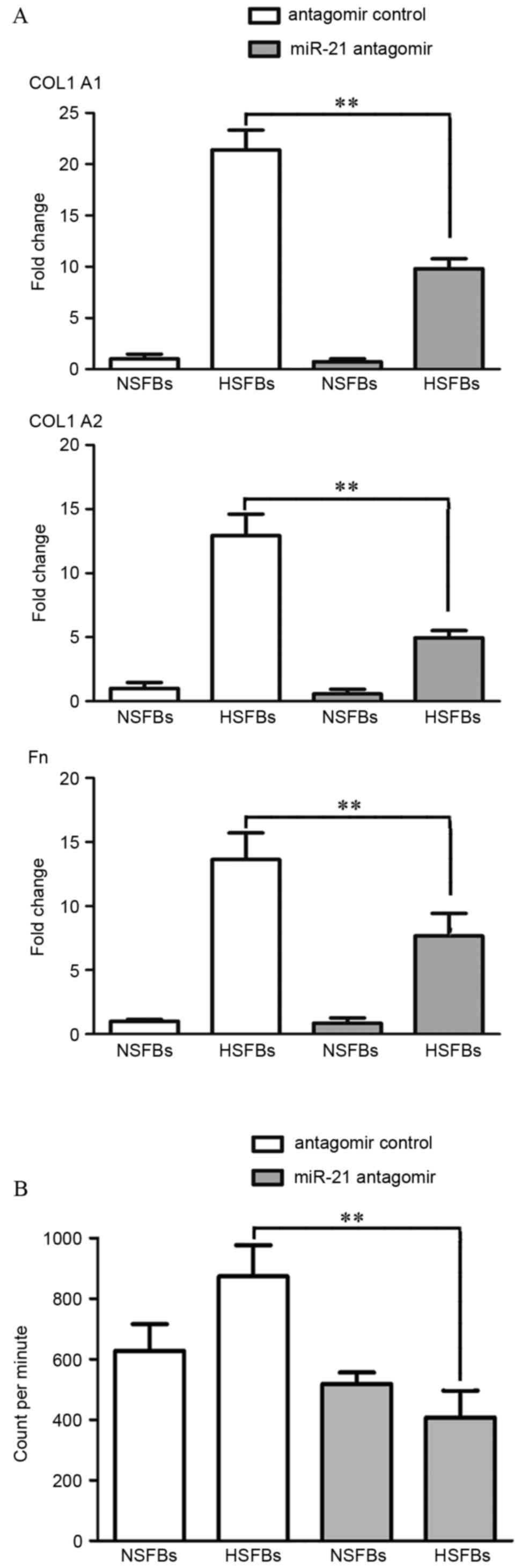
HS is characterized by excessive collagen synthesis.
Thus, the present study investigated the effects of altered miR-21
expression on fibrogenesis. The mRNA expression of
fibrosis-associated genes, including COL1A1, COL1A2 and Fn,
following treatment of HSFBs and NSFBs with miR-21 antagomir or
antagomir control was investigated. Compared with NSFBs, mRNA
levels of COL1A1, COL1A2 and Fn were elevated in HSFBs, and were
significantly decreased by miR-21 antagomir transfection in HSFBs
(P<0.01; Fig. 4A).
Additionally, the H3-proline incorporation assay demonstrated that
HSFBs exhibited increased collagen synthesis compared with NSFBs in
the antagomir control groups. However, after 48 h of treatment with
miR-21 antagomir, collagen synthesis by HSFBs was significantly
decreased compared with HSFBs treated with the antagomir control
(P<0.0001), whereas collagen synthesis by NSFBs changed only
marginally (Fig. 4B). These
results suggest that miR-21 has a pathological role in skin
fibrosis and inflammation by promoting the synthesis of collagen,
and targeting miR-21 may be a potential therapeutic strategy to
block the progression of abnormal scar formation.
Inhibition of miR-21 in vivo
diminishes the HS formation in rabbit ears
To investigate whether inhibition of miR-21 has
therapeutic potential in the treatment of HS, miR-21 antagomir or
antagomir control were administered by subcutaneous injection
following complete healing of wounds (day 15) in a HS model in
rabbit ears. miR-21 continued to increase over 9 days following
wound epithelization. However, the increase in miR-21 was
significantly suppressed by miR-21 antagomir (Fig. 5A). The extent of skin fibrosis was
subsequently analyzed. Macroscopic images of scar samples were
taken at each time point by digital camera (Fig. 5B). At 10 days after complete
healing (day 25), the scars in the control group were severely
congested and appeared purple and swollen. By contrast, the
perfusion of scars in the intervention group was dramatically
reduced and these scars appeared pink and less swollen following
injection of miR-21 antagomir. At 20 days after healing (day 35),
the intra-scar microvascular perfusion in both groups had decreased
and the scars had become less red. However, scars in the control
group remained thick, and had become hardened, while scars in the
intervention group were contracted and had softened. The diameter
of scars in both groups was measured with a slide gauge. Between
days 35 and 65, the diameter of scars in the miR-21 antagomir
treated group was reduced compared with the control group
(P<0.05; Fig. 5C). The
thickness of scars, measured as SEI, in both groups decreased over
time. However, the decrease was faster in the intervention group
compared with the control group and significant statistical
differences in the SEI were observed between the control and
intervention group on days 35 and 65 (P<0.05; Fig. 5D). Compared with the control group,
administration of miR-21 antagomir led to an ~80% reduction of the
SEI in rabbit ear HS model on day 65 (P<0.0001).
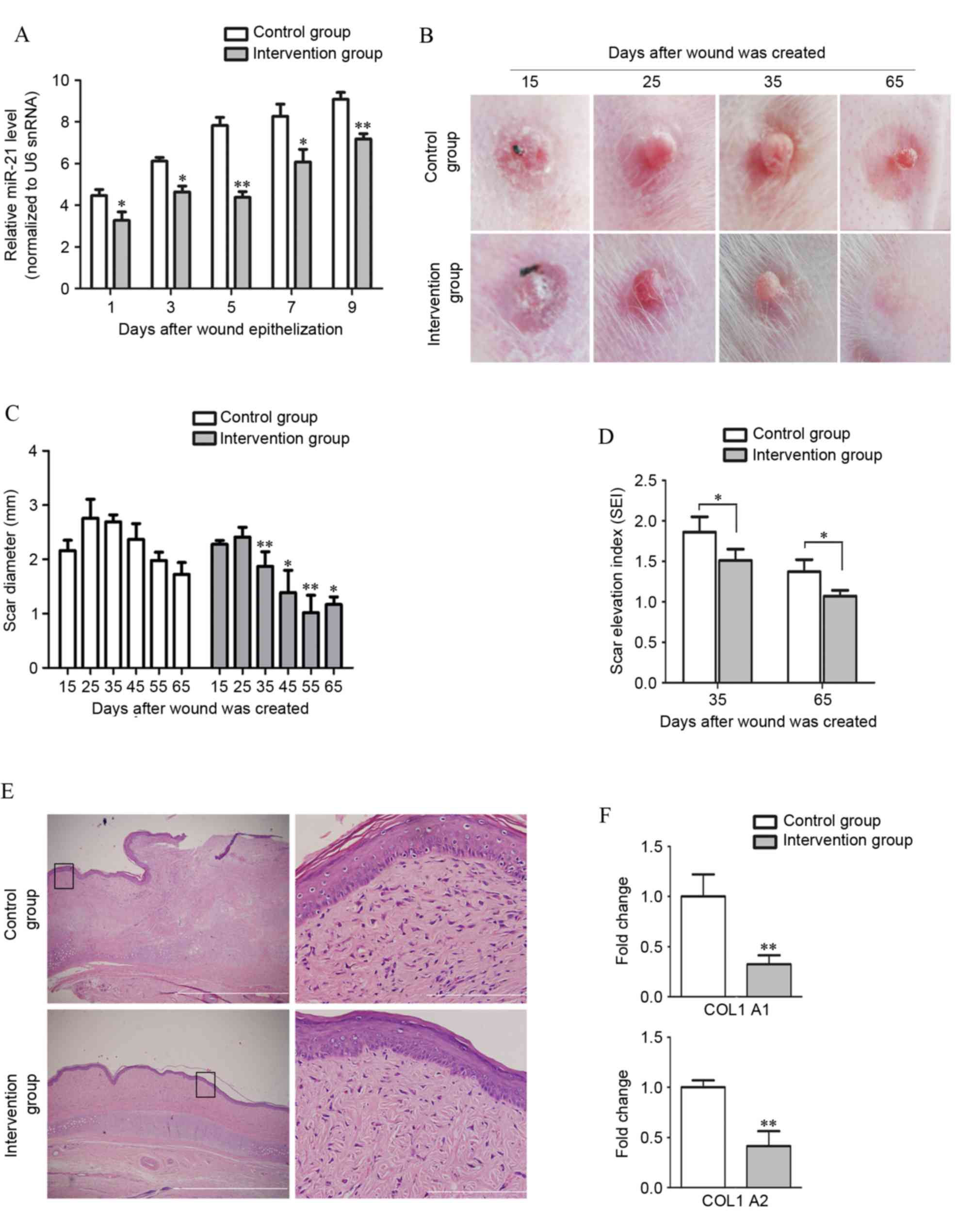 | Figure 5.Local treatment with miR-21 antagomir
significantly decreases scar formation. (A) miR-21 antagomir
administration in the intervention group significantly suppressed
the upregulation of miR-21 in scar tissue (n=3) following wound
epithelialization, as compared with the miRNA antagomir control
administration group. (B) Representative images of scar formation
in the rabbit ear model. Scars in the group withanti-miR-21
intervention exhibited a delay in formation process compared with
the control group. (C) Quantification of formed HS. The diameter of
scars in both groups was measured with a slide gauge. Between day
25 and day 65, the diameter of scars in the miR-21 antagomir
intervention group was significantly smaller compared with the
control group. (D) Comparison of SEI on days 35 and 65. The degree
of dermal hypertrophy of each scar was expressed as the SEI. This
index is the ratio of the area of newly formed dermis of the scar
to the area of surrounding normal dermis. (E) Pathological analyses
by hematoxylin and eosin staining confirmed that scar formation was
inhibited in the miR-21 antagomir-treated group when comparing the
collagen distribution on day 65. The distribution of collagen
fibers in the control group were dense, enlarged and disordered,
with more cells in the epidermis and dermis of HS, while collagen
fibers were more mature and well arranged with few cells in the
intervention group. Boxes in the left column indicate the areas
that are enlarged in the corresponding panels in the right column.
Scale bars: 1 µm (right) and 50 µm (left). (F) Expression of COL1A1
and COL1A2 in different groups of scars was measured and compared
by reverse transcription-quantitative polymerase chain reaction
analysis. miR, microRNA; miRNA, microRNA; HS, hypertrophic scar;
SEI, scar elevation index; snRNA, small nuclear RNA; COL1A1,
collagen type I α 1 chain; COL1A2, collagen type I α 2 chain.
*P<0.05, **P<0.01 vs. the control group. |
On day 35 post-wounding, scar samples were harvested
and collagen distribution was observed by hematoxylin and eosin
staining (Fig. 5E). Collagen
fibers in the control group were dense, enlarged and disordered,
with more cells in the epidermis and dermis of HS, while collagen
fibers were more mature and well arranged with few cells in the
intervention group, which suggested downregulation of proliferative
activity in HS by miR-21 antagomir administration. Finally, mRNA
expression of COLA1 and COLA2 in scars was measured by RT-qPCR and
the expression levels of COLA1 (P<0.0001) and COLA2
(P<0.0001) were significantly repressed by miR-21 antagomir
treatment (Fig. 5F). These
differences between the two groups suggest a therapeutic effect of
miR-21 antagomir in HS.
Discussion
HS is one of the most common skin disorders and is
characterized by pathological, excessive dermal fibrosis. Patients
with HS experience a severe reduction in quality of life due to
physical, psychological and social sequelae (1,14).
There is currently no efficient treatment for HS, partially due to
the incomplete understanding of the molecular pathogenesis of this
disease. Recently, it has emerged that miRNAs have critical roles
in various pathophysiological processes and have been reported to
participate in tissue development, differentiation, repair and
fibrosis (3–6). Strategies that interfere with miRNA
function are considered to offer novel opportunities for clinical
treatment. However, little is known about the expression and
functions of miRNAs in HS formation and how miRNAs regulate
fibrosis in HS remains elusive.
In the present study, miRNA expression profiling was
performed and a unique miRNA signature associated with HS
development was identified. miRNA microarray profiling identified
at least 152 miRNAs with altered expression in HS, including 82
upregulated and 70 downregulated miRNAs. The majority of these
miRNAs have been reported to function as oncogenes or tumor
suppressors (5), which supports
the assumptions regarding the similarity of excess proliferation in
HS and cancer cells. Certain other miRNAs were reported to be
closely associated with the major events of organ fibrosis. For
example, miR-31-5p was reported to be upregulated in keloid
(15) and systemic sclerosis
(16), miR-503 inhibits
epithelial-to-mesenchymal transition in gastric cancer cells
(17) and miR-34a has a role in
the progression of cardiac tissue fibrosis by directly targeting
SMAD family member 4 (18), which
suggests that miR-34a may be a novel marker for cardiac fibrosis
progression. Microarray analysis in the present study indicated
that certain miRNAs are involved in the pathological process of HS
formation, and subgroup analysis of miRNAs should be performed to
further investigate thisassociation.
miRNA expression analysis in other fibrotic
processes has also been reported. Zarjou et al (19) identified a unique miRNA signature
associated with renal fibrosis, including upregulated miR-21,
miR-142-3p, miR-142-5p, miR-214 and miR-223, and significantly
downregulated miR-101a, miR-193 and miR-218. It was reported that
miRNA alteration drove the acute and late stages of
radiation-induced fibrosis in a murine skin model (20). In a keloid study, 293
differentially expressed miRNAs were identified (21). Several differentially expressed
miRNAs identified in the present study were also reported in these
studies, including miR-21, miR-214 and miR-34, suggesting that
these miRNAs may have an important role in the regulation of
fibrosis formation. However, several differentially expressed
miRNAs identified in the present study were distinct from the study
on keloid (21). The reason for
the discrepancy may depend on the miRNAs that were screened in the
experiments, and also on the differences between keloid and HS.
Although both involve persistent fibrosis of the skin, HS and
keloid have pathophysiological and clinical differences (1). HS are typically raised, red or pink,
and sometimes pruritic, however they do not exceed the margins of
the original wound, whereas keloids infiltrate surrounding normal
tissue and rarely regress. Additionally, HS usually subsides over
time, while keloid continues to evolve without a quiescent or
regressive phase. Accordingly, in the present study, the expression
of miR-30c-1-3p and miR-23c was downregulated in HS, whereas in
keloid, miR-30a-5p and miR-23b-3p were significantly upregulated in
a comparative study of miRNA profiling performed by Li et al
(15). It is assumed that the
differences in the expression of these miRNAs may partially explain
why keloid and HS have different clinical presentations and
prognosis. Further investigation is required to verify this
assumption.
In addition tithe increased expression of miR-21 in
HS, as identified in the microarray analysis performed by the
present study, evidence suggests that miR-21 may participate in
numerous fibrotic processes, including renal fibrosis (22), systemic sclerosis (23) and idiopathic pulmonary fibrosis
(24). The results of these
studies indicate that miR-21 has an important role in fibrogenesis.
Hsa-miR-21 is located on human chromosome 17q23.2 and is
transcribed to produce primary-miR-21. The miR-21 strand of the
duplex (miR-21-5p) is incorporated into the RNA-induced silencing
complex (RISC), while miR-21-3p (previously termed miR-21*)
separates from the duplex and is rapidly degraded. miR-21 then
facilitates the binding of RISC to the 3-UTR of the target mRNA,
thus, inhibiting protein translation or promoting degradation
(25). In the present study,
RT-qPCR confirmed that miR-21 was upregulated in HS tissue and
HSFBs. ISH also highlighted extensive expression of miR-21 in
epithelial cells and dermal fibroblasts. Similarly, Wang et
al (26) reported a similar
miR-21 expression pattern in epithelial cells of the epidermis and
mesenchymal cells of the dermis in the wound. These results
indicated that miR-21 may have multiple roles during the course of
fibrosis.
It is commonly accepted that HS is caused by high
cell proliferation rates and excessive collagen deposition, and
abnormal HSFBs are involved in this process. The in vitro
experiments performed in this study demonstrated that inhibition of
miR-21 significantly inhibited the growth of HSFBs by inducing
apoptosis (27). In addition,
knockdown of miR-21 reduced the expression of fibrosis-associated
genes COL1A1, COL1A2 and Fn. These data suggest that an miR-21
antagomir may be used as a strategy to inhibit HS development
clinically. To test this hypothesis, an in vivo rabbit ear
scar model of hypertrophic scarring was applied (28). In the present study, miR-21
antagomir was injected directly into HS and, subsequently, miR-21
antagomir significantly decreased the thickness of scars.
Additionally, scars in the intervention group were flat and
softened compared with the control group. Collagen deposition was
also reduced following miR-21 antagomir administration and the
distribution of fibers was less disordered. These in vivo
and in vitro results indicated that miR-21-induced cell
proliferation and collagen deposition are important in the
development of HS. The experiments performed in the present study
suggest that targeting miR-21 may be a potential therapeutic
strategy in HS prevention and treatment, and miR-21 antagomir may
be an effective agent to decrease miR-21 expression and reduce
excessive fibrosis (29).
It is important to note that miRNAs modulate
biological functions via multiple target mRNAs. It is likely that
miR-21 participates in fibrotic events in kidney, lung, heart or
other organs by regulating a unique array of targets. Liu et
al (30) reported that miR-21
promoted collagen deposition by targeting SMAD family member 7,
whilst Hatley et al (31)
reported that miR-21 drove tumorigenesis through inhibition of
negative regulators of the Ras/MEK/extracellular signal-regulated
kinase pathway. A previous study reported that loss of PTEN, a
well-established miR-21 target, in dermal fibroblasts caused skin
fibrosis (32). Our previous study
also observed that PTEN expression was significantly decreased in
HS, and that PTEN inhibited the proliferation and function of HSFBs
through the Akt serine/threonine kinase (AKT) pathway (10). Furthermore, Zhu et al
(33) recently reported that
miR-21 regulated human telomerase reverse transcriptase expression
via the PTEN/phosphatidylinositol-3 kinase/AKT signaling pathway by
directly targeting PTEN, thus, controlling growth of HSFBs.
Therefore, it is possible that the anti-fibrotic effect of the
miR-21 antagomir may be achieved by stabilizing PTEN in HSFBs.
In conclusion, the present study revealed a unique
miRNA expression signature associated with the development of HS by
microarray analysis. Furthermore, experiments demonstrated that an
miR-21 antagomir reversed the phenotype of fibrotic fibroblasts
in vitro and attenuated HS formation in an in vivo
rabbit ear HS model. The results of the present studydemonstrated
that miR-21 has an important role in HS formation and suggest that
the miR-21 antagomir is a potential agent for the treatment of HS
and other fibrotic skin conditions, including keloid.
Acknowledgements
This research was supported by grants from the
National Natural Science Foundation of China (grant nos. 81301638
and 81301639), the State Key Laboratory of Trauma, Burn and
Combined Injury of China development program (grant no.
SKLZZ201122), and the Grant of Wuhan Young and Middle-aged Medical
Personnel Training Project. This manuscript has been edited and
proofread by Medjaden Bioscience, Ltd.
References
|
1
|
Wolfram D, Tzankov A, Pülzl P and
Piza-Katzer H: Hypertrophic scars and keloids-a review of their
pathophysiology, risk factors, and therapeutic management. Dermatol
Surg. 35:171–181. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Karagoz H, Bayram Y, Sever C, Sahin C,
Kulahci Y and Ulkur E: A review of the prevention and treatment of
hypertrophic scars: Part II experimental studies. Arch Clin Exp
Surg. 2:49–58. 2013. View Article : Google Scholar
|
|
3
|
Bentwich I, Avniel A, Karov Y, Aharonov R,
Gilad S, Barad O, Barzilai A, Einat P, Einav U, Meiri E, et al:
Identification of hundreds of conserved and nonconserved human
microRNAs. Nat Genet. 37:766–770. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Backes C and Keller A: Reanalysis of 3,707
novel human microRNA candidates. Proc Natl AcadSci USA.
112:E2849–E2850. 2015. View Article : Google Scholar
|
|
5
|
Bouyssou JM, Manier S, Huynh D, Issa S,
Roccaro AM and Ghobrial IM: Regulation of microRNAs in cancer
metastasis. Biochim Biophys Acta. 1845:255–265. 2014.PubMed/NCBI
|
|
6
|
Lei W, Li G, Zheng J, Shui X, Huang S and
Chen C: Roles of microRNA in vascular diseases in cardiac and
pulmonary systems. Pharmazie. 69:643–647. 2014.PubMed/NCBI
|
|
7
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Obernosterer G, Martinez J and Alenius M:
Locked nucleic acid-based in situ detection of microRNAs in
mouse tissue sections. Nat Protoc. 2:1508–1514. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tandara AA and Mustoe TA: The role of the
epidermis in the control of scarring: Evidence for mechanism of
action for silicone gel. J Plast ReconstrAesthet Surg.
61:1219–1225. 2008.
|
|
10
|
Guo L, Chen L, Bi S, Chai L, Wang Z, Cao
C, Tao L and Li S: PTEN inhibits proliferation and functions of
hypertrophic scar fibroblasts. Mol Cell Biochem. 361:161–168. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ou H, Li Y and Kang M: Activation of
miR-21 by STAT3 induces proliferation and suppresses apoptosis in
nasopharyngeal carcinoma by targeting PTEN gene. PLoS One.
9:e1099292014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Duffield JS, Grafals M and Portilla D:
MicroRNAs are potential therapeutic targets in fibrosing kidney
disease: Lessons from animal models. Drug Discov Today Dis Models.
10:e127–e135. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dong S, Ma W, Hao B, Hu F, Yan L, Yan X,
Wang Y, Chen Z and Wang Z: microRNA-21 promotes cardiac fibrosis
and development of heart failure with preserved left ventricular
ejection fraction by up-regulating Bcl-2. Int J Clin Exp Pathol.
7:565–574. 2014.PubMed/NCBI
|
|
14
|
Gabriel V: Hypertrophic scar. Phys Med
Rehabil Clin N Am. 22:301–310. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li C, Bai Y, Liu H, Zuo X, Yao H, Xu Y and
Cao M: Comparative study of microRNA profiling in keloid fibroblast
and annotation of differential expressed microRNAs. Acta Biochim
Biophys Sin (Shanghai). 45:692–699. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu H, Li Y, Qu S, Luo H, Zhou Y, Wang Y,
Zhao H, You Y, Xiao X and Zuo X: MicroRNA expression abnormalities
in limited cutaneous scleroderma and diffuse cutaneous scleroderma.
J Clin Immunol. 32:514–522. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Peng Y, Liu YM, Li LC, Wang LL and Wu XL:
microRNA-503 inhibits gastric cancer cell growth and
epithelial-to-mesenchymal transition. Oncol Lett. 7:1233–1238.
2014.PubMed/NCBI
|
|
18
|
Huang Y, Qi Y, Du JQ and Zhang DF:
MicroRNA-34a regulates cardiac fibrosis after myocardial infarction
by targeting Smad4. Expert Opin Ther Targets. 18:1355–1365.
2014.PubMed/NCBI
|
|
19
|
Zarjou A, Yang S, Abraham E, Agarwal A and
Liu G: Identification of a microRNA signature in renal fibrosis:
Role of miR-21. Am J Physiol Renal Physiol. 301:F793–F801. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Simone BA, Ly D, Savage JE, Hewitt SM, Dan
TD, Ylaya K, Shankavaram U, Lim M, Jin L and Camphausen K: microRNA
alterations driving acute and late stages of radiation-induced
fibrosis in a murine skin model. Int J Radiat Oncol Biol Phys.
90:44–52. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu ZY, Lu L, Liang J, Guo XR, Zhang PH and
Luo SJ: Keloid microRNA expression analysis and the influence of
miR-199a-5p on the proliferation of keloid fibroblasts. Genet Mol
Res. 13:2727–2738. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Glowacki F, Savary G, Gnemmi V, Buob D,
Van der Hauwaert C, Lo-Guidice JM, Bouyé S, Hazzan M, Pottier N,
Perrais M, et al: Increased circulating miR-21 levels are
associated with kidney fibrosis. PLoS One. 8:e580142013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu H, Luo H, Li Y, Zhou Y, Jiang Y, Chai
J, Xiao X, You Y and Zuo X: MicroRNA-21 in scleroderma fibrosis and
its function in TGF-β-regulated fibrosis-related genes expression.
J Clin Immunol. 33:1100–1109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li P, Li J, Chen T, Wang H, Chu H, Chang
J, Zang W, Wang Y, Ma Y, Du Y, et al: Expression analysis of serum
microRNAs in idiopathic pulmonary fibrosis. Int J Mol Med.
33:1554–1562. 2014.PubMed/NCBI
|
|
25
|
da Costa Martins PA and De Windt LJ:
miR-21: A miRaculous Socratic paradox. Cardiovasc Res. 87:397–400.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang T, Feng Y, Sun H, Zhang L, Hao L, Shi
C, Wang J, Li R, Ran X, Su Y and Zou Z: miR-21 regulates skin wound
healing by targeting multiple aspects of the healing process. Am J
Pathol. 181:1911–1920. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Buscaglia LE and Li Y: Apoptosis and the
target genes of microRNA-21. Chin J Cancer. 30:371–380. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Morris DE, Wu L, Zhao LL, Bolton L, Roth
SI, Ladin DA and Mustoe TA: Acute and chronic animal models for
excessive dermal scarring: Quantitative studies. Plast Reconstr
Surg. 100:674–681. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mattes J, Yang M and Foster PS: Regulation
of microRNA by antagomirs: A new class of pharmacological
antagonists for the specific regulation of gene function? Am J
Respir Cell Mol Biol. 36:8–12. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu G, Friggeri A, Yang Y, Milosevic J,
Ding Q, Thannickal VJ, Kaminski N and Abraham E: miR-21 mediates
fibrogenic activation of pulmonary fibroblasts and lung fibrosis. J
Exp Med. 207:1589–1597. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hatley ME, Patrick DM, Garcia MR,
Richardson JA, Bassel-Duby R, van Rooij E and Olson EN: Modulation
of K-Ras-dependent lung tumorigenesis by MicroRNA-21. Cancer Cell.
18:282–293. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Parapuram SK, Shi-wen X, Elliott C, Welch
ID, Jones H, Baron M, Denton CP, Abraham DJ and Leask A: Loss of
PTEN expression by dermal fibroblasts causes skin fibrosis. J
Invest Dermatol. 131:1996–2003. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhu HY, Li C, Bai WD, Su LL, Liu JQ, Li Y,
Shi JH, Cai WX, Bai XZ, Jia YH, et al: MicroRNA-21 regulates hTERT
via PTEN in hypertrophic scar fibroblasts. PLoS One. 9:e971142014.
View Article : Google Scholar : PubMed/NCBI
|