Introduction
S100 calcium binding protein A6 (S100A6), which is
also known as calcyclin, is a low-molecular-weight calcium-binding
protein of the S100 family. S100A6 was originally purified from
Ehrlich ascite tumor cells (1) and
was subsequently detected in a wide range of cell types, including
epithelial cells, neurons and fibroblasts (2). S100A6 interacts with various proteins
to regulate cell proliferation and differentiation, the dynamics of
cytoskeletal constituents and calcium homeostasis (3). Increased expression of S100A6 has
been reported in numerous malignancies, including colorectal
carcinoma, osteosarcoma, pancreatic cancer and urothelial carcinoma
(4–8); however, its highest expression has
been observed in epithelial cells and fibroblasts (2). S100A6 has previously been
demonstrated to be present in cancer tissues and is associated with
tumor behavior and patient prognosis (9); however, the association between
S100A6 and endometriosis remains to be fully elucidated.
Endometriosis is defined as the presence of
endometrial glands and stroma within extrauterine sites, and may
result in pelvic pain and infertility. It affects ~10% of women of
reproductive age (10).
Endometriosis is not considered a malignant disorder; however, it
does exhibit various characteristics similar to tumor cells,
including the ability to metastasize and invade distant regions.
However, the details of its etiology, pathogenesis and
pathophysiology remain to be fully elucidated. It has previously
been demonstrated that the Wnt/β-catenin signaling pathway, which
is involved in development, tissue self-renewal and various
diseases (11,12), is aberrantly activated in the
endometrium of patients with endometriosis (13).
Our previous study demonstrated that S100A6 was
highly expressed in eutopic endometrial stromal cells (14), and upregulated levels of S100A6
increased the expression of β-catenin; one of the most important
signaling molecules in the Wnt/β-catenin signaling pathway
(15). Furthermore, inhibition of
S100A6 has been reported to decrease the proliferation and
invasiveness of pancreatic cancer cells in vitro (16). However, the function of S100A6 in
endometriosis has yet to be fully elucidated. The present study
therefore hypothesized that S100A6 may be important in the
development of endometriosis via regulating the expression of
β-catenin.
Materials and methods
Ethics statement
The research protocol was approved by the
Consultative Committee for the Protection of People in Biomedical
Research of Jiangxi Maternal and Child Health Hospital (Nanchang,
China). Written informed consent was obtained from each patient
prior to participation in the investigation.
Cell culture and recombinant
lentiviral constructs
Eutopic endometrial stromal cells were derived from
the endometrial tissue of 20 female patients (8 proliferative phase
and 12 secretory phase cases) with ovarian endometriosis (mean age,
43.6±3.2 years) that presented at the Jiangxi Province Key
Laboratory of Molecular Medicine (Nanchang, China). The patients
had not received any prior hormone therapy or chemotherapy. A
section of each endometrial specimen was submitted for histological
diagnosis and the remainder was frozen in liquid nitrogen and later
prepared for use in subsequent experiments.
Cells were cultivated in Dulbecco's modified Eagle's
medium (DMEM)/F12 medium (Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 10% fetal bovine serum (FBS; Thermo
Fisher Scientific, Inc.) and incubated at 37°C in a humidified
atmosphere containing 5% CO2. Experimental recombinant
lentivirus [small interfering RNA (si)S100A6-enhanced green
fluorescent protein (EGFP)-specific] and control recombinant
lentivirus (without siS100A6) vectors were purchased from Shanghai
GenePharma Co., Ltd. (Shanghai, China). EGFP was included as a
reporter gene. The interference fragment sequence used for S100A6
siRNA was 5′-AAGCTGCAGGATGCTGAAATT-3′, based on previous research
findings (17).
siRNA transfection with
lentiviruses
Eutopic endometrial stromal cells were seeded in
6-well plates and grown to 60–70% confluence. Cells were then
transfected with the control recombinant lentivirus vector
(Lv-control), siS100A6 lentivirus vector (Lv-siS100A6), or with no
vector (untreated control) using Polybrene® reagent
(Sigma-Aldrich; Merck Millipore, Darmstadt, Germany) in accordance
with the manufacturer's protocol. All transfections were performed
in triplicate. Transfected cells were cultured at 37°C in an
atmosphere containing 5% CO2 for 4 days, and were
subsequently used for a range of analytical experiments.
Western blot analysis
A total of 4 days post-transfection, cells were
lysed in ice-cold radioimmunoprecipitation assay buffer containing
1% protease inhibitors (Roche Diagnostics, Basel, Switzerland).
Proteins were harvested and the protein concentration was
determined using the Bicinchoninic Acid Protein Assay kit (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Proteins (20 µg) were
separated by 8% SDS-PAGE and transferred to a nitrocellulose
membrane (EMD Millipore, Billerica, MA, USA). Membranes were
blocked for 2 h with 5% non-fat dried milk in Tris-buffered saline
containing 0.1% Tween-20 (TBST) at room temperature, and were then
incubated at 4°C overnight with rabbit anti-human β-catenin (1:800;
cat. no. ab6302), S100A6 (1:1,000; cat. no. 181974), or β-actin
(1:1,000; cat. no. ab6276) primary antibodies (Abcam, Cambridge,
UK). Membranes were subsequently washed three times in TBST and
incubated with horseradish peroxidase-conjugated goat anti-rabbit
immunoglobin G (1:10,000; Beijing Zhongshan Jinqiao Biotechnology
Co., Ltd., Beijing, China; cat. no. ZDR-5306) for 2 h at room
temperature. Following three washes in TBST, specific bands were
detected using the enhanced chemiluminescence system (EMD
Millipore). The bands were analyzed by using Gel-Pro Analyzer 4.0
software (Media Cybernetics, Inc., Rockville, MD, USA). The
monoclonal anti-β-actin antibody served as an internal control for
loading.
Cell proliferation assay
A total of 4 days post-transfection, cells were
cultivated in 96-well plates at a density of 3×103 cells/well in a
total volume of 100 µl/well. A total of 10 µl Cell Counting Kit-8
(CCK-8; Dojindo Molecular Technologies, Inc., Kumamoto, Japan) was
added to each well when the cells reached 40–50% confluence. The
absorbance of each well was measured at a wavelength of 450 nm
using a microtiter plate reader (Molecular Devices, LLC, Sunnyvale,
CA, USA). All experiments were performed in triplicate and results
are expressed as the mean ± standard deviation.
Cell migration assay
Migration assays were conducted in modified Boyden
chambers (BD Biosciences, Franklin Lakes, San Jose, CA, USA) with
8-µm pore filter inserts in 24-well plates. A total of 4 days
post-transfection, 2×104 cells suspended in serum-free DMEM/F12
were added to the upper chamber. DMEM/F12 supplemented with 20% FBS
was added to the lower chamber as a chemoattractant. Following a 48
h cultivation at 37°C, the membrane containing migrated cells was
fixed using methanol and stained with crystal violet. Migration was
detected by light microscopy following removal of the non-filtered
cells on the upper side of the membrane with cotton swabs. Three
independent experiments were performed.
Cell apoptosis assay
A total of 4 days post-transfection, cells were
harvested, diluted to a concentration of 3×105 cells/ml, and washed
twice with ice-cold phosphate-buffered saline. Subsequently, the
cells were incubated in the dark, at room temperature for 15 min
with phycoerythrin (PE) Annexin V and 7-aminoactinomycin D (7-AAD)
and 380 µl 1X binding buffer from the Annexin V-PE/7-AAD apoptosis
kit (Multisciences, Beijing, China) according to the manufacturer's
protocol, and were analyzed by flow cytometry. Cells undergoing
early apoptosis only bound to PE Annexin V, whereas cells that
bound to PE Annexin V and 7-AAD were in the late stages of, or had
already undergone, apoptosis. The experiment was repeated three
times.
RNA extraction and reverse
transcription (RT)-semi-quantitative polymerase chain reaction
(PCR)
A total of 4 days post-transfection, total RNA was
isolated from cells grown to 70–80% confluence using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Total RNA (2 µg)
was used for the synthesis of first-strand cDNA using First Strand
cDNA Synthesis kit (Promega Corporation, Madison, WI, USA). PCR
primers were designed using Primer Premier 5.0 software (Premier
Biosoft International, Palo Alto, CA, USA) from reported sequences
(Table I). Target gene expression
was normalized to β-actin levels.
 | Table I.Primer sequences used for polymerase
chain reaction. |
Table I.
Primer sequences used for polymerase
chain reaction.
| Gene | Primer sequence | Product size
(bp) |
|---|
| S100A6 | Sense:
5′-ACACCCTGAGCAAGAAGGAG-3′ | 198 |
|
| Antisense:
5′-CCCTTGAGGGCTTCATTGTA-3′ |
|
| β-catenin | Sense:
5′-GATTTGATGGAGTTGGACATGG-3′ | 149 |
|
| Antisense:
5′-TCTTCCTCAGGATTGCCTT-3′ |
|
| β-actin | Sense:
5′-ATCATGTTTGAGACCTTCAACA-3′ | 318 |
|
| Antisense:
5′-CATCTCTTGCTCGAAGTCCA-3′ |
|
PCR reactions were performed in a 20-µl volume
containing 0.5 µl cDNA, 20 pmol of each primer, 0.25 mM of each
dNTP, 1 unit Taq DNA polymerase (Promega Corporation), and the
buffer supplied by the manufacturer. β-catenin amplification used
the following thermal cycling conditions: Initial denaturation at
94°C for 3 min followed by 35 cycles of denaturation at 94°C for 30
sec, primer annealing at 53°C for 30 sec and primer extension at
72°C for 25 sec, with a final extension at 72°C for 5 min. S100A6
and β-actin amplification was similar, with the exception of an
annealing temperature of 57°C. PCR products were separated by 1.5%
agarose gel electrophoresis and were subjected to densitometric
scanning using an Epson photo scanner (Epson America Inc., Long
Beach, CA, USA). Bands were semi-quantified using Gel-Pro Analyzer
4.0 software (Media Cybernetics, Inc., Rockville, MD, USA). All
experiments were conducted in triplicate.
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical analysis was conducted using SPSS software version 19.0
(IBM SPSS, Armonk, NY, USA). Statistical analyses were performed
using a one-way analysis of variance and Student's t-test for
comparisons between two different groups, or χ2 test. P<0.05 was
considered to indicate a statistically significant difference.
Results
EGFP expression in eutopic endometrial
stromal cells
Eutopic endometrial stromal cells were transfected
with Lv-control or Lv-siS100A6. The efficiency of transfection was
assessed 4 days post-transfection by analyzing EGFP expression
using fluorescence microscopy, it was indicated that transfection
was efficient.
Analysis of post-transfection
intracellular S100A6 expression and its interrelation with
β-catenin
Western blotting and RT-PCR were performed to
measure intracellular S100A6 expression 4 days following the
transfection of eutopic endometrial stromal cells. S100A6 protein
expression was decreased in cells transfected with Lv-siS100A6
compared with untreated and Lv-control-transfected cells,
(0.239±0.013, 0.689±0.010 and 0.706±0.008 for Lv-siS100A6 group,
untreated group and Lv-control group, respectively; P<0.05),
following normalization to β-actin levels (Fig. 1A and B). There was no significant
difference in the expression of S100A6 between untreated and
Lv-control cells (P>0.05). The mRNA expression levels of S100A6
were also lower in cells transfected with Lv-siS100A6 compared with
the untreated and Lv-control-transfected cells (P<0.05; Fig. 1C).
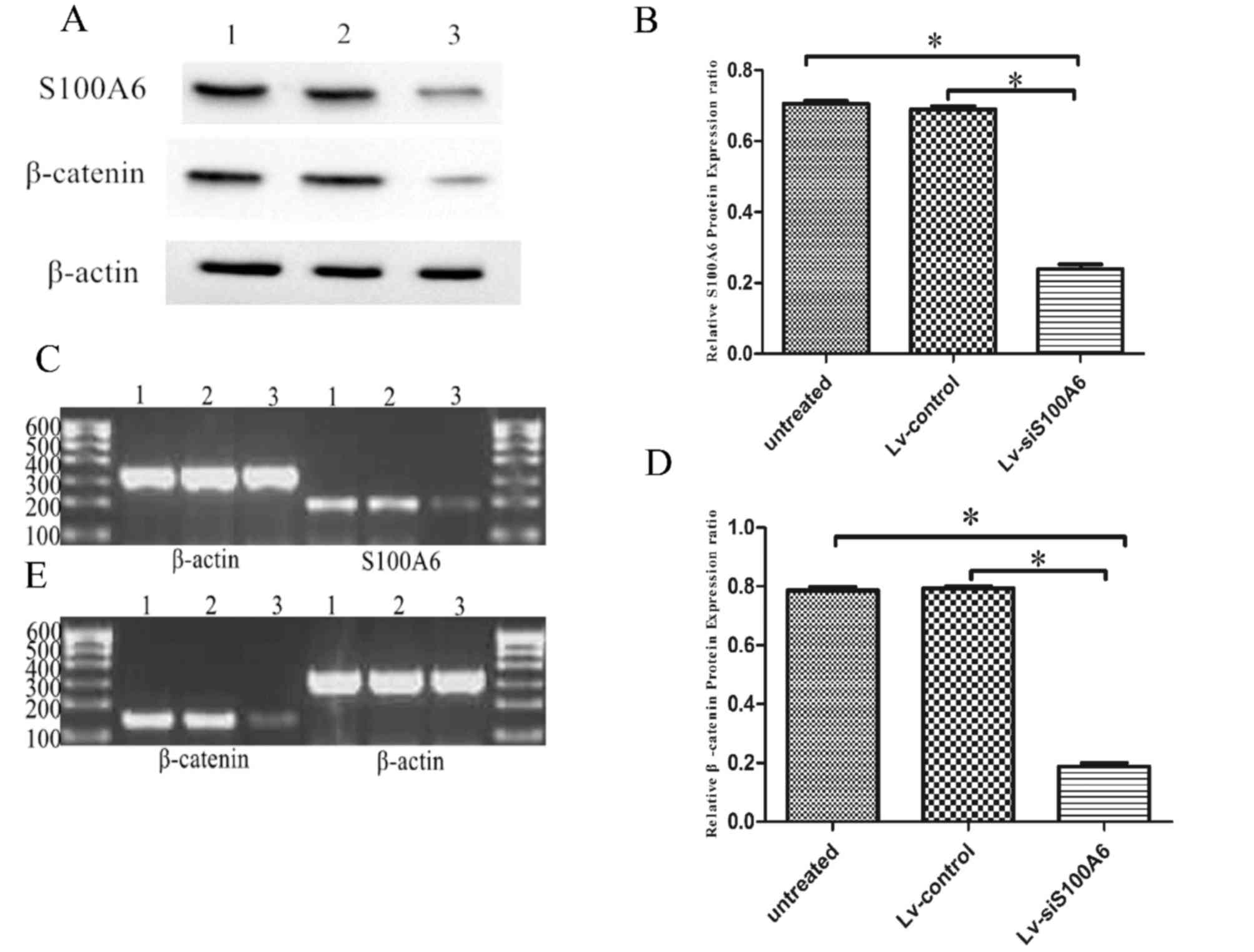 | Figure 1.Silencing of S100A6 downregulated the
mRNA and protein expression levels of β-catenin in eutopic
endometrial stromal cells, as determined by RT-PCR and western blot
analysis. (A) Western blot analysis indicated decreased S100A6 and
β-catenin protein levels in eutopic endometrial stromal cells
following treatment with Lv-siS100A6 compared with Lv-control. Lane
1, untreated group; lane 2, Lv-control group; lane 3, Lv-siS100A6
group. The effect of siS100A6 on S100A6 (B) protein and (C) mRNA
expression, and β-catenin (D) protein and (E) mRNA expression was
evaluated 4 days post-transfection by western blotting and RT-PCR,
respectively. Data are presented as the mean ± standard deviation.
*P<0.05 vs. untreated and Lv-control groups. S100A6, S100
calcium binding protein A6; si, small interfering RNA; Lv-siS100A6,
siS100A6 recombinant lentivirus vector; Lv-control, control
recombinant lentivirus vector; RT-PCR, reverse
transcription-polymerase chain reaction. |
To further examine the interaction between S100A6
and β-catenin, the expression levels of β-catenin were detected
using western blotting and RT-PCR, in cells post-transfection with
Lv-siS100A6 or Lv-control constructs. As presented in Fig. 1A and D, β-catenin expression was
significantly lower in Lv-siS100A6-transfected cells (0.188±0.012)
compared with Lv-control (0.794±0.006) and untreated cells
(0.787±0.011) following normalization to β-actin levels. In
addition, β-catenin mRNA levels were observed to be markedly
decreased in Lv-siS100A6-transfected cells, whereas no notable
difference was detected in β-catenin levels between Lv-control and
untreated cells (Fig. 1E).
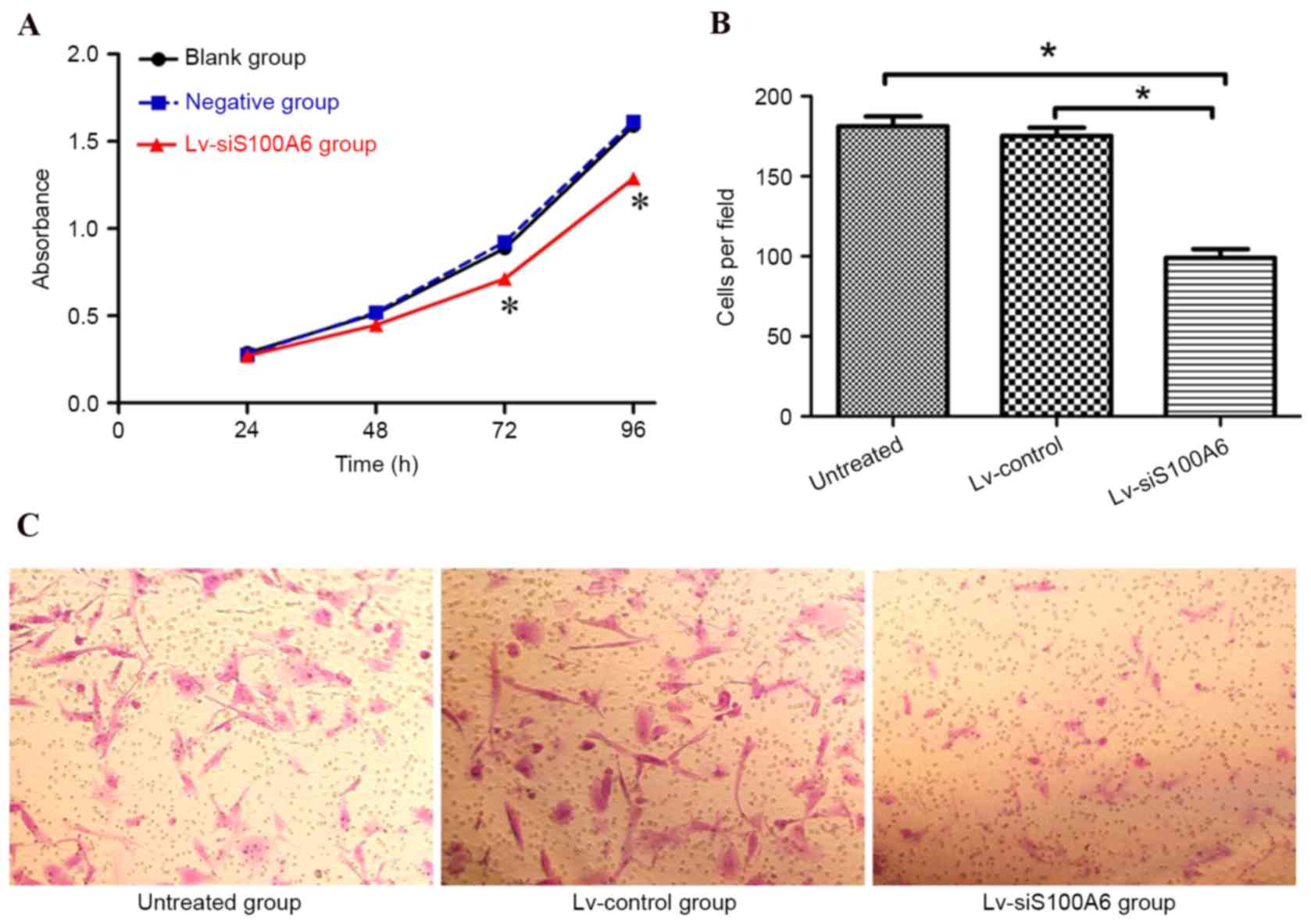
siS100A6 suppresses the proliferation
and migration of eutopic endometrial stromal cells in vitro
To further investigate the functional roles of
S100A6 in endometriosis, the present study examined the effects of
S100A6 silencing on eutopic endometrial stromal cells. As presented
in Fig. 2A, the CCK-8
proliferation assay revealed that the growth rate was significantly
reduced in cells transfected with the Lv-siS100A6 construct
compared with untreated cells or those transfected with the
Lv-control construct (P<0.05).
The effects of siS100A6 on the migratory capacity of
eutopic endometrial stromal cells were also investigated. As
presented in Fig. 2B and C, the
number of migrated cells was significantly reduced in the
Lv-siS100A6 group compared with the untreated group or those
transfected with Lv-control, (182.00±13.21/per field,
170.00±20.32/per field and 96.00±21.34/per field in untreated,
Lv-control-, and Lv-siS100A6-transfected cells, respectively).
siS100A6 promotes apoptosis in eutopic
endometrial stromal cells in vitro
To determine whether the siS100A6-induced inhibition
of proliferation and migration may be a consequence of the
induction of cell death, the present study examined the number of
early apoptotic eutopic endometrial stromal cells 4 days
post-transfection. Flow cytometric analysis revealed that few
Lv-control-transfected (13.73%) or untreated cells (12.25%)
underwent early apoptosis, whereas transfection with the
Lv-siS100A6 construct significantly increased the percentage of
early apoptotic cells to 35.44% (Fig.
3A and B; P<0.05).
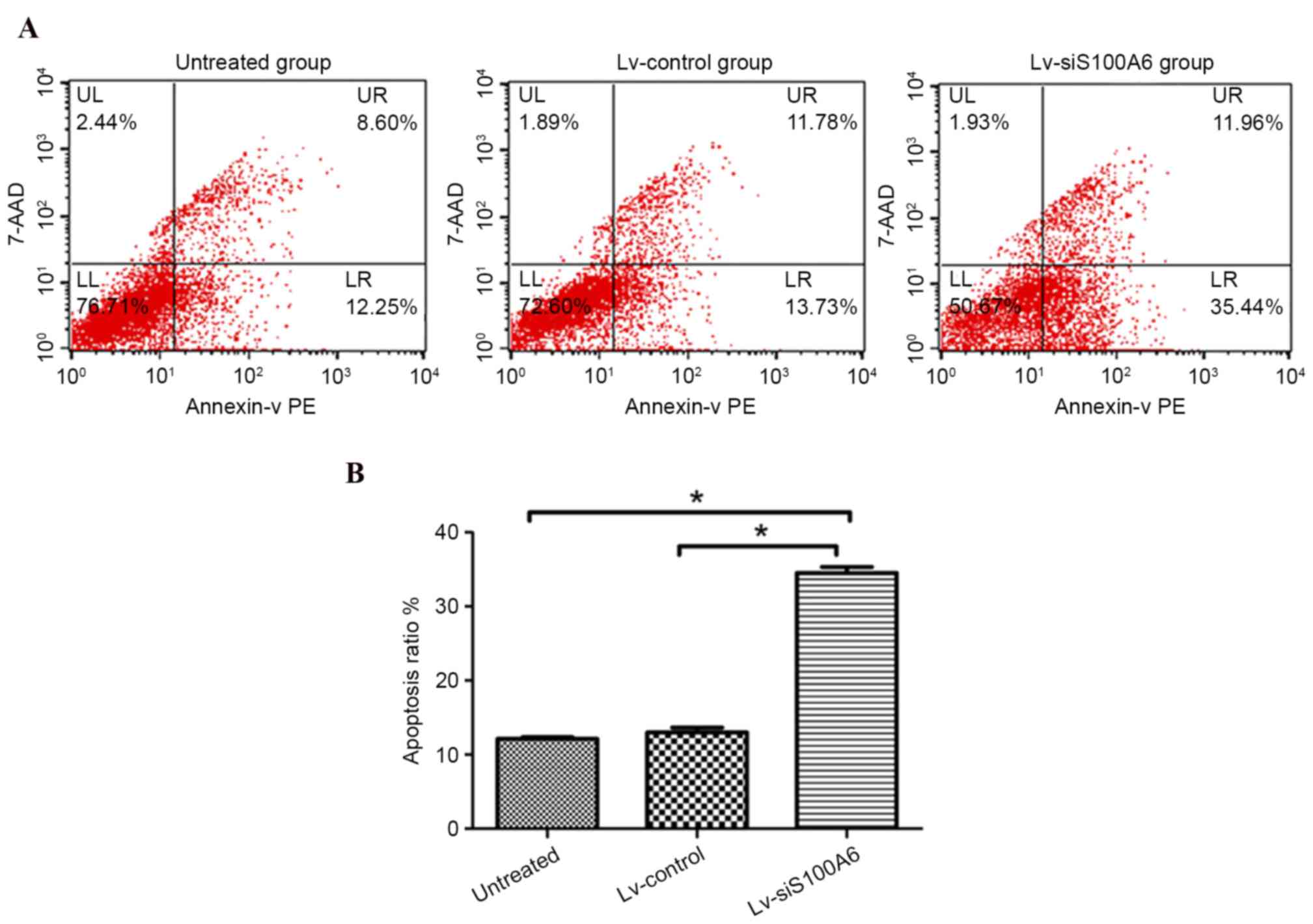 | Figure 3.Eutopic endometrial stromal cell
apoptotic rates were detected by flow cytometry following
transfection with Lv-siS100A6. (A) Analysis of apoptosis was
conducted 4 days post-transfection. Cells undergoing early
apoptosis only bound to PE Annexin V, whereas cells that bound to
PE Annexin V and 7-AAD were in the late stages, or had already
undergone apoptosis. (B) Knockdown of endogenous S100A6 induced
eutopic endometrial stromal cell apoptosis. The LR quadrants were
used to determine apoptotic ratio. The results represent one of
three experiments performed. Data are presented as the mean ±
standard deviation. *P<0.05 vs. untreated and Lv-control groups.
S100A6, S100 calcium binding protein A6; si, small interfering RNA;
Lv-siS100A6, siS100A6 recombinant lentivirus vector; Lv-control,
control recombinant lentivirus vector; PE, phycoerythrin; 7-AAD,
7-aminoactinomycin D; UL, upper left; UR, upper right; LL, lower
left; LR, lower right. |
Discussion
S100 proteins are the largest subgroup of the
EF-hand Ca2+ binding protein family, with cell- and
tissue-specific expression patterns. They are multi-functional
signaling proteins, which serve roles in cell growth, cell cycle
progression, differentiation, transcription and secretion (18). A particular characteristic of these
proteins is that individual members are localized in discrete
cellular compartments, from which a few are able to relocate, upon
Ca2+ activation (19,20).
These proteins may temporally and spatially transduce the
Ca2+ signal by interacting with various specific target
proteins. Notably, the majority of S100 genes are located within a
gene cluster on chromosome lq21, which is a site associated with
frequent chromosomal abnormalities (21).
Numerous S100 protein members have been revealed to
be associated with tumor development and metastasis (22,23).
Du et al (24) reported
that S100P induced the dissociation of non-muscle myosin IIA
filaments, leading to a weakening of focal adhesion sites and an
enhancement of cell migration. Zhang et al (25) demonstrated that S100A4 regulated
migration and invasion in HepG2 cells via matrix metallopeptidase
(MMP) 9 signaling. MMPs are important in cell invasion and
migration, and MMP2 expression was revealed to significantly
increase in a p53-dependent manner following S100A14 overexpression
(26). Similarly, Li et al
(27) reported that MMP9
expression was upregulated following S100A6 treatment, which
promoted cell proliferation, migration and tumorigenesis of
HepG2.2.15 cells.
The authors of the present study previously
demonstrated that S100A6 was highly expressed in eutopic
endometrial stromal cells (14),
and revealed that S100A6 overexpression increased the proliferation
and migration of these cells, and inhibited their apoptosis
(15). Endometriotic and malignant
cells undergo blood vessel development and a decrease in the number
of apoptotic cells (28).
Spuijbroek et al (29)
confirmed in vitro that the ectopic endometrium and cancer
cells share an invasive ability.
In the present study, S100A6 silencing by RNA
interference (RNAi) significantly inhibited the proliferation and
migration of eutopic endometrial stromal cells, while promoting
their apoptosis. Endometriosis was previously suggested to occur
following aberrant activation of the Wnt/β-catenin signaling
pathway (12). Intervention of the
pathway in vivo resulted in alterations in eutopic
endometrial invasion and adhesion, which affected the development
of endometriosis (30).
Furthermore, the pathway was observed to promote eutopic
endometrial cell proliferation and to improve the efficiency of
eutopic endometrial implantation, invasion, metastasis and
angiogenesis (30).
β-catenin is a central molecule of the Wnt/β-catenin
signaling pathway and the present study observed that its
expression was decreased following S100A6 inhibition. Kilanczyk
et al (31) observed that
the expression of S100A6 was regulated by β-catenin in colorectal
cancer cells, suggesting that the S100A6 gene may be a
transcriptional target of β-catenin. In addition, reduced β-catenin
levels have been reported in HepG2.2.15 cells, which result in
decreased S100A6 levels (32).
β-catenin binds to E-cadherin to form adherence junctions (33). Upon loss of binding with
E-cadherin, unphosphorylated β-catenin accumulates in the cytoplasm
and partially translocates into the nucleus, resulting in the
activation of β-catenin/Tcf-mediated transcription and downstream
target genes (34). Expression of
these genes, including c-myc avian myelocytomatosis viral
oncogene homolog, cyclin D and MMPs, may be responsible for
cellular proliferation and migration. Furthermore, Shaco-Levy et
al (35) reported the
expression of MMP-2, MMP-9, E-cadherin and β-catenin in
endometriosis.
In conclusion, the present study demonstrated that
suppression of S100A6 expression with RNAi significantly decreased
the expression levels of β-catenin, inhibited the growth and
migration of eutopic endometrial stromal cells, and promoted their
apoptosis. These results suggested that the multiple effects of
S100A6 may be dependent on regulation of β-catenin expression via
activation of the Wnt/β-catenin signaling pathway. Furthermore, the
results indicated that S100A6 is important in endometrial
metastasis and may increase understanding regarding the precise
functions of S100A6 in the promotion and progression of
endometriosis. Inhibition of S100A6 may be considered a novel
approach for the targeted therapy of endometriosis.
Acknowledgements
The present study was supported by the Jiangxi
Provincial Natural Science Foundation of China (grant no.
20123BCB22010).
References
|
1
|
Kuźnicki J and Filipek A: Purification and
properties of a novel Ca2+-binding protein (10.5 KDa) from
Ehrlich-ascites-tumour cells. Biochem J. 247:663–667. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kuźnicki J, Kordowska J, Puzianowska M and
Wozniweicz BM: Calcyclin as a marker of human epithelial cells and
fibroblasts. Exp Cell Res. 200:425–430. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Leśniak W, Słomnicki ŁP and Filipek A:
S100A6-new facts and features. Biochem Biophys Res Commun.
390:1087–1092. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Duan L, Wu R, Zou Z, Wang H, Ye L, Li H,
Yuan S, Li X, Zha H, Sun H, et al: S100A6 stimulates proliferation
and migration of colorectal carcinoma cells through activation of
the MAPK pathways. Int J Oncol. 44:781–790. 2014.PubMed/NCBI
|
|
5
|
Li Y, Wagner ER, Yan Z, Wang Z, Luther G,
Jiang W, Ye J, Wei Q, Wang J, Zhao L, et al: The calcium-binding
protein S100A6 accelerates human osteosarcoma growth by promoting
cell proliferation and inhibiting osteogenic differentiation. Cell
Physiol Biochem. 37:2375–2392. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luu HH, Zhou L, Haydon RC, Deyrup AT,
Montag AG, Huo D, Heck R, Heizmann CW, Peabody TD, Simon MA and He
TC: Increased expression of S100A6 is associated with decreased
metastasis and inhibition of cell migration and anchorage
independent growth in human osteosarcoma. Cancer Lett. 229:135–148.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zihao G, Jie Z, Yan L, Jing Z, Jing C, Xue
L, Jing Z, Heng LW, Ru G and Jianyu H: Analyzing S100A6 expression
in endoscopic ultrasonography-guided fine-needle aspiration
specimens: A promising diagnostic method of pancreatic cancer. J
Clin Gastroenterol. 47:69–75. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nishi M, Matsumoto K, Kobayashi M,
Yanagita K, Matsumoto T, Nagashio R, Ishii D, Fujita T, Sato Y and
Iwamura M: Serum expression of S100A6 is a potential detection
marker in patients with urothelial carcinoma in the urinary
bladder. Biomed Res. 35:351–356. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang J, Zhang K, Jiang X and Zhang J:
S100A6 as a potential serum prognostic biomarker and therapeutic
target in gastric cancer. Dig Dis Sci. 59:2136–2144. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Giudice LC and Kao LC: Endometriosis.
Lancet. 364:1789–1799. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Clever H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wend P, Holland JD, Ziebold U and
Birchmeier W: Wnt signaling in stem and cancer stem cells. Semin
Cell Dev Biol. 21:855–863. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Matsuzaki S, Darcha C, Maleysson E, Canis
M and Mage G: Impaired down-regulation of E-cadherin and
beta-catenin protein expression in endometrial epithelial cells in
the mid-secretory endometrium of infertile patients with
endometriosis. J Clin Endocrinol Metab. 95:3437–3445. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiaoling Z, Lin Q, Xiaohong Y, Huihong Z
and Huai L: Expression of S100A6 in eutopic and ectopic endometrium
of ovarian endometriosis patients. Chinese Journal of Practical
Gynecology and Obstetrics. 26:778–780. 2010.
|
|
15
|
Liu Z, Zhang X, Chen M, Cao Q and Huang D:
Effect of S100A6 over-expression on β-catenin in endometriosis. J
Obstet Gynaecol Res. 41:1457–1462. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ohuchida K, Mizumoto K, Ishikawa N and
Tanaka M: The role of S100A6 in pancreatic cancer development and
its clinical implication as a diagnostic marker and therapeutic
target. Clin Cancer Res. 11:7785–7793. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fuli Z, Ying Z, Yaning Z, Miao D and Jian
W: Influence of S100A6 siRNA on biological features of human
ovarian cancer cell. Chinese Journal of Woman and Child Health
Research. 25:48–51. 2014.
|
|
18
|
Yammani RR: S100 proteins in cartilage:
Role in arthritis. Biochim Biophys Acta. 1822:600–606. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Van Dieck J, Brandt T, Teufel DP,
Veprintsev DB, Joerger AC and Fersht AR: Molecular basis of S100
proteins interacting with the p53 homologs p63 and p73. Oncogene.
29:2024–2035. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shimamoto S, Kubota Y, Tokumitsu H and
Kobayashi R: S100 proteins regulate the interaction of Hsp90 with
Cyclophilin 40 and FKBP52 through their tetratricopeptide repeats.
FEBS Lett. 584:1119–1125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schäfer BW, Wicki R, Engelkamp D, Mattei
MG and Heizmann CW: Isolation of a YAC clone covering a cluster of
nine S100 genes on human chromosome lq21: Rationale for a new
nomenclature of the S100 protein family. Cenomics. 25:638–643.
1995.
|
|
22
|
Schäfer BW and Heizmann CW: The S100
family of EF-hand calcium-binding proteins: Functions and
pathology. Trends Biochem Sci. 21:134–140. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Saleem M, Kweon MH, Johnson JJ, Adhami VM,
Elcheva I, Khan N, Bin Hafeez B, Bhat KM, Sarfaraz S, Reagan-Shaw
S, et al: S100A4 accelerates tumorigenesis and invasion of human
prostate cancer through the transcriptional regulation of matrix
metalloproteinase 9. Proc Natl Acad Sci USA. 103:14825–14830. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Du M, Wang G, Ismail TM, Gross S, Fernig
DG, Barraclough R and Rudland PS: S100P dissociates myosin IIA
filaments and focal adhesion sites to reduce cell adhesion and
enhance cell migration. J BiolChem. 287:15330–15344. 2012.
|
|
25
|
Zhang J, Zhang DL, Jiao XL and Dong Q:
S100A4 regulates migration and invasion in hepatocellular carcinoma
HepG2 cells via NF-kB-dependent MMP-9 signal. Eur Rev Med Pharmacol
Sci. 17:2372–2382. 2013.PubMed/NCBI
|
|
26
|
Chen H, Yuan Y, Zhang C, Luo A, Ding F, Ma
J, Yang S, Tian Y, Tong T, Zhan Q and Liu Z: Involvement of S100A14
protein in cell invasion by affecting expression and function of
matrix metalloproteinase (MMP)-2 via p53-dependent transcriptional
regulation. J Biol Chem. 287:17109–17119. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li Z, Tang M, Ling B, Liu S, Zheng Y, Nie
C, Yuan Z, Zhou L, Guo G, Tong A and Wei Y: Increased expression of
S100A6 promotes cell proliferation and migration in human
hepatocellular carcinoma. J Mol Med(Berl). 92:291–303. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Swiersz LM: Role of endometriosis in
cancer and tumor development. Ann N Y Acad Sci. 955:281–295,
396–406. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Spuijbroek MD, Dunselman GA, Menheere PP
and Evers JL: Early endometriosis invades the extracellular matrix.
Fertil Steril. 58:929–933. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liang JY, Li CD and Zhang WY: Effects of
activating and inhibiting Wnt/β-catenin signaling pathway on murine
model of eutopic endometrium and endometriosis. Zhonghua Yi Xue Za
Zhi. 92:1352–1356. 2012.(In Chinese). PubMed/NCBI
|
|
31
|
Kilańczyk E, Graczyk A, Ostrowska H,
Kasacka I, Leśniak W and Filipek A: S100A6 is transcriptionally
regulated by β-catenin and interacts with a novel target, lamin
A/C, in colorectal cancer cell. Cell Calcium. 51:470–477. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee YT, Dimitrova YN, Schneider G,
Ridenour WB, Bhattacharya S, Soss SE, Caprioli RM, Filipek A and
Chazin WJ: Structure of the S100A6 complex with a fragment from the
c-terminal domain of Siah-1 interacting protein: A novel mode for
S100 protein target recognition. Biochemistry. 47:10921–10932.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fuchs SY, Ougolkow AV, Spiegelman VS and
Minamoto T: Oncegenice beta-catenin signaling networks in
colorectal cancer. Cell Cycle. 4:1522–1539. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Valenta T, Hausmann G and Basler K: The
many faces and functions of β-catenin. EMBO J. 31:2714–2736. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shaco-Levy R, Sharabi S, Benharroch D,
Piura B and Sion-Vardy N: Matrix metalloproteinases 2 and 9,
E-cadherin, and beta-catenin expression in endometriosis, loe-grade
endometrial carcinoma and non-neoplastic eutopic endometrium. Eur J
Obstet Gynecol Reprod Biol. 139:226–232. 2008. View Article : Google Scholar : PubMed/NCBI
|