Introduction
Diabetic nephropathy (DN) is the most severe and
most common long-term microvascular complication in diabetes
mellitus (DM), and 20–40% of diabetic patients are at risk of
developing DN (1). In addition
with changing lifestyle and dietetic habits, the incidence of
diabetes has increased annually worldwide (2). Chronic renal failure resulting from
DN is the leading cause for dialysis and renal transplantation in
Europe, America and developed areas of China (3). However, the pathogenesis of DN
remains to be fully elucidated. Renal tubular lesions appear in the
early stage of DN and tubulointerstitial fibrosis, which is closely
associated with renal failure, has a substantial effect on renal
function in DN (4,5).
The activity of the transforming growth factor
(TGF)-β1 signaling pathway can be negatively regulated by
Ski-related novel protein N (SnoN), which is a member of the Ski
family of protooncoproteins, which are involved in binding to small
mothers against decapentaplegic (Smad) complexes (6). SnoN has been found to be expressed at
high levels and localized exclusively to the nucleus in normal
renal tissues (7), which strictly
limits the activity of the TGF-β1 signaling pathway. In this
respect, the elevated expression of SnoN is considered a critical
gatekeeper and may be responsible for the restriction of Smad
signaling in normal conditions. However, the expression level of
SnoN is progressively reduced in a time-dependent manner in the
fibrotic kidney (8), which renders
the restriction mechanism ineffective and can lead to renal
fibrosis resulting from the enhanced TGF-β1 signaling pathway. The
TGF-β1/Smad signaling pathway has been recognized as a pathway
closely associated with renal hypertrophy and tubulointerstitial
fibrosis (9). SnoN is an important
negative regulator of TGF-β1/Smad signaling, and its expression
level has a marked effect on the TGF-β1/Smad pathway (9). Several E3 ubiquitin ligases have been
shown to control the TGF-β1/Smad signaling pathway with
dual-directional regulation, including Smad ubiquitination
regulatory factor 2 (smurf2). Upon TGF-β1/Smad stimulation, the
phosphorylated (p-) Smad2 interacts with smurf2 and the degradation
of SnoN degradation by ubiquitination is activated by binding to
smurf, which enhances the activity of TGF-β1/Smad. Smurf2 also
functions as a mediator to promote the degradation of TGF-β1
receptor and Smad2, leading to the weakened or depressed biological
function of TGF-β1 (10–12). These studies demonstrated that
smurf2 is important in balancing regulation of the TGF-β1/Smad
signaling pathway.
In our previous study, SnoN depression was found in
DN. To clarify the underlying mechanism, DM rats were treated as
in vivo models. The results indicated that the protein level
of smurf2 was upregulated and that of SnoN was downregulated in the
DM rat renal tissues. The expression level of smurf2 was negatively
correlated with SnoN, however, the mRNA level of SnoN showed no
significant difference. Based on these results, it was hypothesized
that the depressed expression of SnoN was due to increased
degradation, which was induced by smurf2. To further examine the
regulatory mechanism, NRK-52E rat primary renal tubular epithelial
cells and DM rats were utilized in the present study to determine
the effect of smurf2 on the expression of SnoN, and to elucidate
the molecular mechanism underlying SnoN degradation by TGF-β1
stimulation and inhibition assays.
Materials and methods
Reagents
Streptozotocin (STZ) was purchased from
Sigma-Aldrich; Merck Millipore (Darmstadt, Germany). DMEM (low
glucose), DMEM (high glucose) and fetal bovine serum (FBS) were
purchased from HyClone; GE Healthcare Life Sciences (Logan, UT,
USA). Primary antibodies against smurf2 (cat. no. 12024), Smad2
(cat. no. 3103) and p-Smad2 (cat. no. 3108) were purchased from
Cell Signaling Technology, Inc. (Danvers, MA, USA), whereas primary
antibody against SnoN (cat. no. NBP1-77306PEP) was purchased from
Novus Biologicals (Littleton, CO, USA).
Cell culture
The NRK-52E rat primary renal tubular epithelial
cells and 293 human embryonic kidney cell line were purchased from
Stem Cell Bank (Shanghai, China). The NRK-52E cells were cultured
in DMEM high medium (HyClone; GE Healthcare Life Sciences)
containing 5% FBS. The 293 cells were cultured in DMEM high medium
containing 10% FBS. All cells were grown in a humidified 5%
CO2 incubator at 37°C.
Cell treatment
Effects of glucose on cell culture
To examine the reversibility of the effect of
glucose, 2×105 cells were cultured in medium with 24.5
mM glucose at 37°C for 24 h and then switched to medium with 5.5 mM
glucose for 0–4 h. The cells were harvested at different time
points (0, 15 and 30 min for RT-qPCR and 0, 2 and 4 h for western
blot) and the expression levels of smurf2, SnoN, TGF-β1 and Smad2
were detected using reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) and western blot analyses.
Effects of TGF-β1 in cell culture
To investigate the effect of TGF-β1,
2×105 cells were treated with 2 ng/ml TGF-β1 (HumanZyme,
Inc., Chicago, IL, USA) at 37°C for 0–4 h. To inhibit the TGF-β1
signaling pathway, 5 µM TGF-β1 inhibitor (SB431542; Selleck
Chemicals, Houston, TX, USA) was added 2 h prior to the switch to
high glucose. DMSO solvent was used as a control for the TGF-β1
inhibitor. The cells were harvested at different time points (0, 15
and 30 min for RT-qPCR and 0, 2 and 4 h for western blot), and the
expression levels of smurf2, SnoN, TGF-β1 and Smad2 were detected
using RT-qPCR and western blot analyses.
Infection with adenovirus
To induce the overexpression of smurf2,
2.5×105 NRK-52E cells were infected with the AD-smurf2
and AD-negative control (NC) adenovirus which were purchased from
GeneChem Co., Ltd. (Shanghai, China) according to the
manufacturer's protocol, and the medium containing the adenovirus
was replaced with fresh medium 12 h following infection. The cells
were harvested 48 h following infection, and the expression levels
of smurf2, SnoN and the TGF-β1 signaling pathway (smad and p-smad)
were detected using RT-qPCR and western blot analyses. Similarly,
for smurf2 knockdown, the NRK-52E cells were infected with the
AD-short hairpin (sh)smurf2 and AD-shNC adenovirus, respectively,
and the expression levels of smurf2, SnoN and the TGF-β1 signaling
pathway (smad and p-smad) were detected.
Animal model
Male SD rats (n=50; 8 weeks old; 180–200 g; Beijing
Vital River Laboratory Animal Technology Co., Ltd., Beijing, China)
were intraperitoneally injected with 60 mg/kg STZ to establish the
DN rat model. The rats were fasted 12 h prior to model
establishment without water deprivation. The 1% STZ was prepared
with citrate buffer (pH 4.5; 0.1 M) on ice. Prior to injection, the
urine glucose and caudal vein glucose levels of the rats were
confirmed to be normal. Caudal vein glucose was detected 48 and 72
h following injection and rats with glucose concentrations
maintained ≥6.7 mmol/l were recognized as diabetic (DM) rats and
the standard remained unchanged during the experiment.
Overexpression and knockdown models were established through left
renal vein (LRV) injection, prior to which the left renal artery
and vein of the DM rats required closure prior to injection,
according to surgical methods described by Guadalupe et al
(13). The experimental rats were
randomly divided into six groups: i) smurf2 knockdown group, in
which 5×109 pfu/ml AD-shsmurf2 was injected into the
LRV; ii) AD-shNC group, in which 4×109 pfu/ml AD-shNC
was injected into the LRV; iii) smurf2 overexpression group, in
which 5×10 9 pfu/ml AD-smurf2 was injected into the LRV;
iv) AD-NC group, in which 4×109 pfu/ml AD-NC was
injected into the LRV; v) sham group, in which normal saline was
injected into the LRV; vi) control group of untreated DM rats. The
rats were sacrificed by cervical dislocation and the protein
expression levels of smurf2 and SnoN were detected in the renal
tissues of the different groups.
All animal experiments were performed in strict
accordance with the recommendations in the guidelines for the
Animal Care and Use Committee of Beijing Friendship Hospital
(Beijing, China). The rats were housed at 18–26°C on a 12-h
light/dark cycle with free access to water and standard rat chow.
The animals were allowed to acclimatize for a minimum of 1 week.
The environment was maintained at a relative humidity of 30–70%.
All surgery was performed under sodium pentobarbital anesthesia,
and all efforts were made to minimize suffering.
Detection of expression levels of
smurf2, SnoN, TGF-β1 and Smad2 using RT-qPCR analysis
RNA isolation and RT
The total RNA was isolated using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The isolated RNA was reverse transcribed
into cDNA using a ReverTra Ace qPCR RT kit (Toboyo Co., Ltd.,
Osaka, Japan). Briefly, a total of 1,000 ng RNA was used for the
initial RT reaction.
qPCR
The qPCR analysis to measure the expression of mRNA
was performed using a KAPA SYBR FAST qPCR kit (Kapa Biosystems,
Inc., Wilmington, MA, USA) according to the manufacturer's
protocol. cDNA was diluted 1:19 with ddH2O and 5 µl was
added into each well. The qPCR data collection was performed on an
ABI 7500 apparatus (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The reaction was performed for 30 sec at 95°C, 40 cycles at
95°C for 30 sec, and 60°C for 30 sec. RNA 18S used as an endogenous
control in each sample. The relative quantification of mRNA
expression was calculated using the 2−ΔΔCq method
(12) relative to the level of
18S. All reactions were replicated three times.
The qPCR primers for mRNA and 18S were as follows:
Smurf2, forward 5′-AACAAGACCGGCGTCAGAAT-3′ and reverse
5′-TGTTCATAGCCTTCGGGTAG-3′; SnoN, forward
5′-GACAAATTTCTCCTTGGTTCCG-3′ and reverse
5′-CTCTCCATCCGTTTCCATCAGT-3′; Smad2, forward
5′-GCCGCCCGAAGGGTAGAT-3′ and reverse 5′-TTCTGTTCTCCACCACCTGC-3′;
18S, forward 5′-CAGCCACCCGAGATTGAGCA-3′ and reverse
5′-TAGTAGCGACGGGCGGTGTG-3′.
Statistical analysis
Statistical comparisons between two different groups
were determined using Student's t-test with GraphPad Prism 5.0
(GraphPad Software, Inc., La Jolla, CA, USA). The results are
presented as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Western blot analysis
Following the various treatments for the indicated
intervals, the cells and tissues were lysed in lysis buffer. The
protein concentrations were determined using an Enhanced BCA
Protein Assay kit (Beyotime Institute of Biotechnology, Haimen,
China) according to the manufacturer's protocol. The proteins (50
µg) were separated on a 10% SDS-polyacrylamide gel and transferred
electrophoretically onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). The membranes were blocked with 5%
nonfat milk in Tris-buffered saline/0.1% Tween 20 for 1 h at room
temperature, and subsequently incubated with primary antibodies
with the dilution of 1:1,000 at 4°C overnight. The membranes were
then incubated with horseradish peroxidase-conjugated secondary
antibody (goat anti-rabbit immunoglobulin G; cat. no. BS13278; and
goat anti-mouse immunoglobulin G; cat. no. BS12478, respectively;
Bioworld Technology, Inc., St Louis Park, MN, USA) with the
dilution of 1:5,000 at room temperature for 1 h. The protein bands
were visualized using an enhanced chemiluminescence detection
system (GE Healthcare Life Sciences, Chalfont, UK).
Histology and immunohistochemistry (IHC)
Alterations in renal morphology were examined in
methyl Carnoy-fixed, paraffin-embedded tissue sections (4 µm)
stained with hematoxylin and eosin or periodic acid-Schiff stain.
Immunostaining was performed with the paraffinized sections using a
microwave-based antigen retrieval technique. The antibodies used
included smurf2, SnoN, Smad2 and pSmad2 and were all diluted at
1:50. The primary antibodies were incubated at 4°C overnight. An
isotype-matched rabbit IgG was used as a negative control
throughout the experiment. All slides were counterstained with
hematoxylin for visualization of the nuclei.
Results
Protein levels of SnoN are negatively
correlated with smurf2 in low glucose
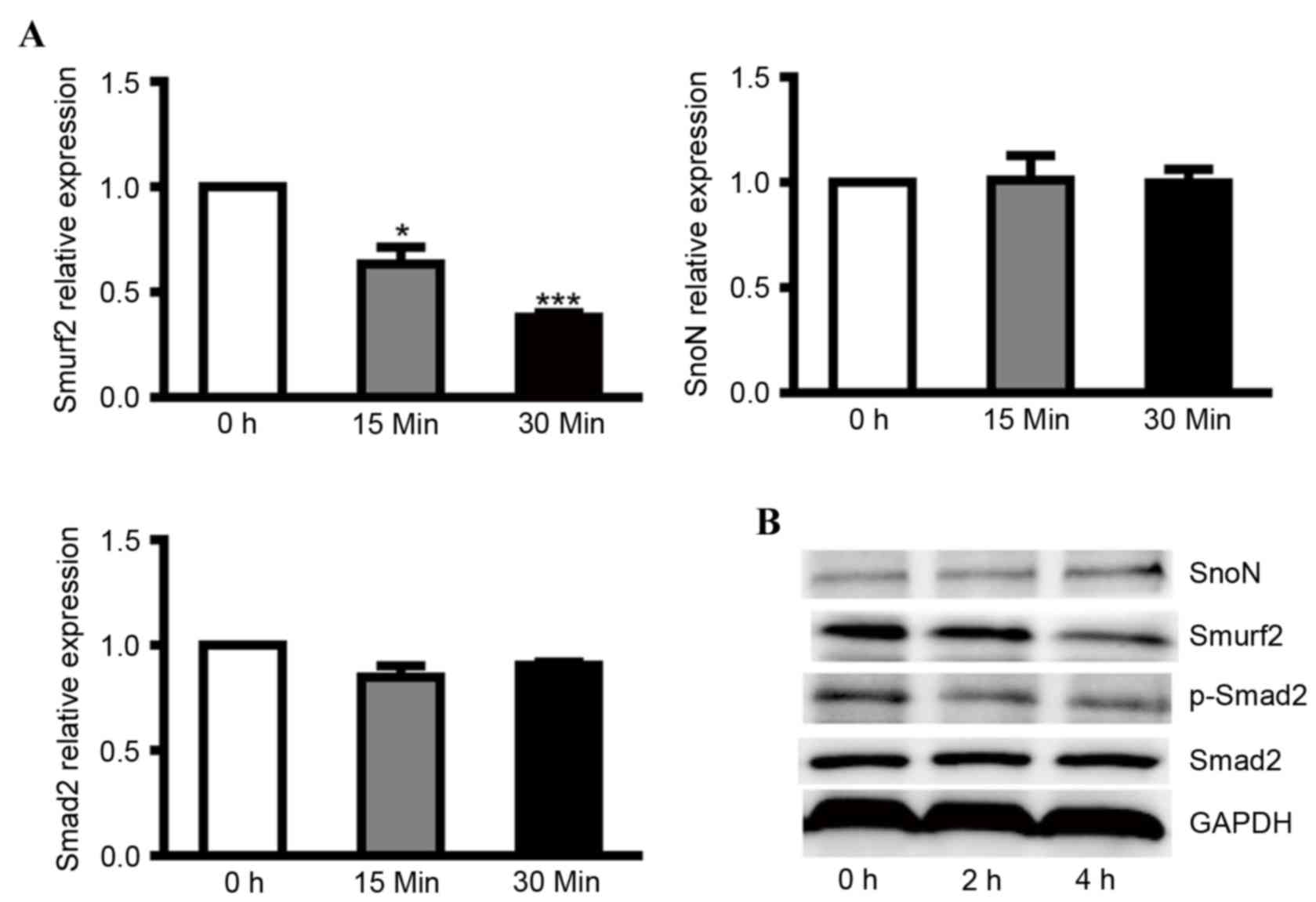
To clarify the regulatory effect of smurf2 on SnoN
in DN, NRK-52E cells were cultured overnight with 24.5 mM glucose
(normal) and then treated with medium containing 5.5 mM glucose
(low). After 30 min, low glucose treatment induced a lower mRNA
expression level of smurf2, however, no significant differences
were found in the mRNA levels of SnoN and Smad2 (Fig. 1A). At the protein level, low
glucose caused higher expression of SnoN and lower expression of
smurf2 and p-Smad2 in the NRK-52E cells (Fig. 1B). These results revealed that low
glucose inactivated the TGF-β1/Smad signaling pathway and that
depressed expression of smurf2 in NRK-52E cells caused the
degradation of SnoN by ubiquitination.
SnoN degradation through
ubiquitination is mediated by smurf2 in vitro
To evaluate whether the degradation of SnoN through
ubiquitination was mediated by smurf2, the NRK-52E cells were
transfected with smurf2 recombinant adenovirus. An
immunofluorescent assay was used to detect the expression of
smurf2. Compared with the control, the NRK-52E cells transfected
with AD-smurf2 showed enhanced red fluorescence (Fig. 2A). No significant difference in the
ectopic overexpression of smurf2 was found in SnoN and Smad2 at the
mRNA level (Fig. 2B). The protein
level of SnoN was reduced, but no effect on the expression of
p-Smad2 was observed (Fig.
2B).
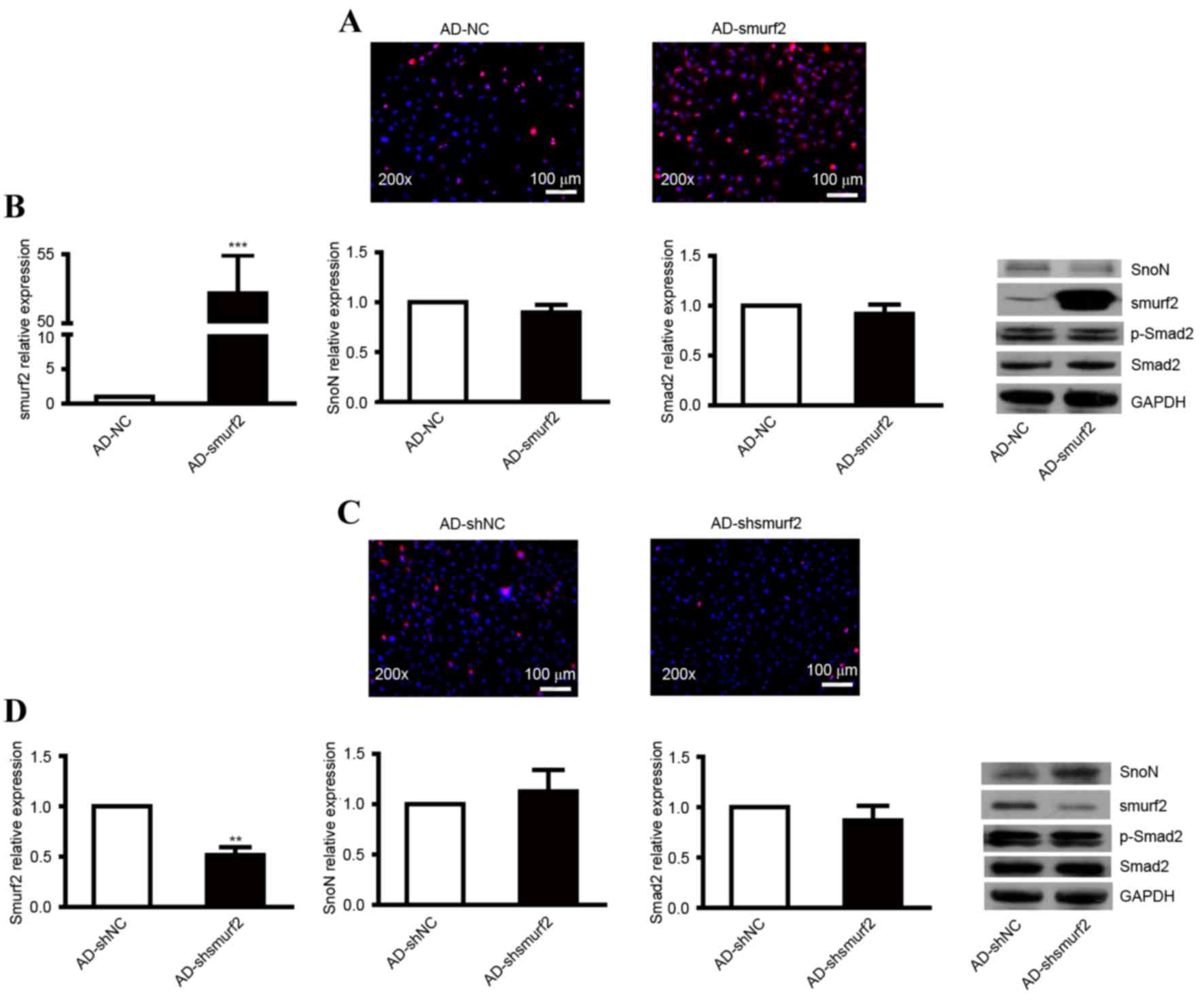 | Figure 2.Expression levels of SnoN and p-Smad2
in NRK-52E cells transfected with recombinant adenovirus. (A)
Immunofluorescence analysis of the expression of smurf2 in NRK-52E
cells infected with AD-smurf2. (B) RT-qPCR analysis was performed
to detect the mRNA levels of smurf2, SnoN and Smad2, and western
blot analysis was performed to detect the expression levels of
smurf2, SnoN and p-Smad2 in NRK-52E cells infected with AD-smurf2.
(C) Immunofluorescence analysis of the expression of smurf2 in
NRK-52E cells treated with AD-shsmurf2. (D) RT-qPCR analysis was
performed to detect the mRNA levels of smurf2, SnoN and Smad2, and
western blot analysis was performed to detect the expression levels
of smurf2, SnoN and p-Smad2 in NRK-52E cells infected with
AD-shsmurf2. ***P<0.001, vs. control. SnoN, ski-related novel
protein N; p-Smad2, phosphorylated small mothers against
decapentaplegic; smurf2, Smad ubiquitination regulatory factor 2;
AD, adenovirus; NC, negative control; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction. |
The present study also infected NRK-52E cells with
recombinant adenovirus AD-shsmurf2 to knockdown the expression of
smurf2. An immunofluorescent assay was used to detect the
expression of smurf2. Compared with the control, NRK-52E cells
infected with AD-shsmurf2 showed weakened red fluorescence
(Fig. 2C). RT-qPCR analysis was
performed to detect the expression levels of smurf2, SnoN and Smad2
in the NRK-52E cells infected with AD-shsmurf2. The results
indicated no significant differences in the expression of SnoN or
Smad2; however, the expression of smurf2 was decreased, compared
with that in NRK-52E cells infected with AD-shNC (Fig. 2D). By performing a western blot
assay, it was found that the downregulated expression of smurf2
increased the protein level of SnoN, however, there was no obvious
alteration in the expression of p-Smad2 (Fig. 2D). Taken together, these results
indicated that smurf2 led to the degradation of SnoN by
ubiquitination when the TGF-β1/Smad signaling pathway was activated
continuously.
Smurf2 induces SnoN degradation in
vivo
To further validate the effect of smurf2 on the
degradation of SnoN in vivo, smurf2 overexpression and
knockdown DM rat models were established. DM rats were injected
with adenovirus AD-smurf2 and AD-shsmurf2, respectively. The smurf2
expression profile was upregulated in tissues collected from the
AD-smurf2 group, compared with those from the control, however, no
significant difference in the expression levels of SnoN or Smad2
were observed at the mRNA level (Fig.
3A). At protein level, the overexpression of smurf2 decreased
the expression of SnoN, but had no effect on the expression of
p-Smad2 (Fig. 3A). This difference
was further confirmed using immunohistochemistry (IHC), which
revealed the expression of smurf2 and SnoN in renal tubules. Smurf2
was localized to the nucleus and plasma membrane; however, SnoN was
predominantly expressed in the plasma membrane. The results of the
IHC staining showed that the overexpression of smurf2 decreased the
expression of SnoN (Fig. 3B),
which was consistent with the results of the western blot
analysis.
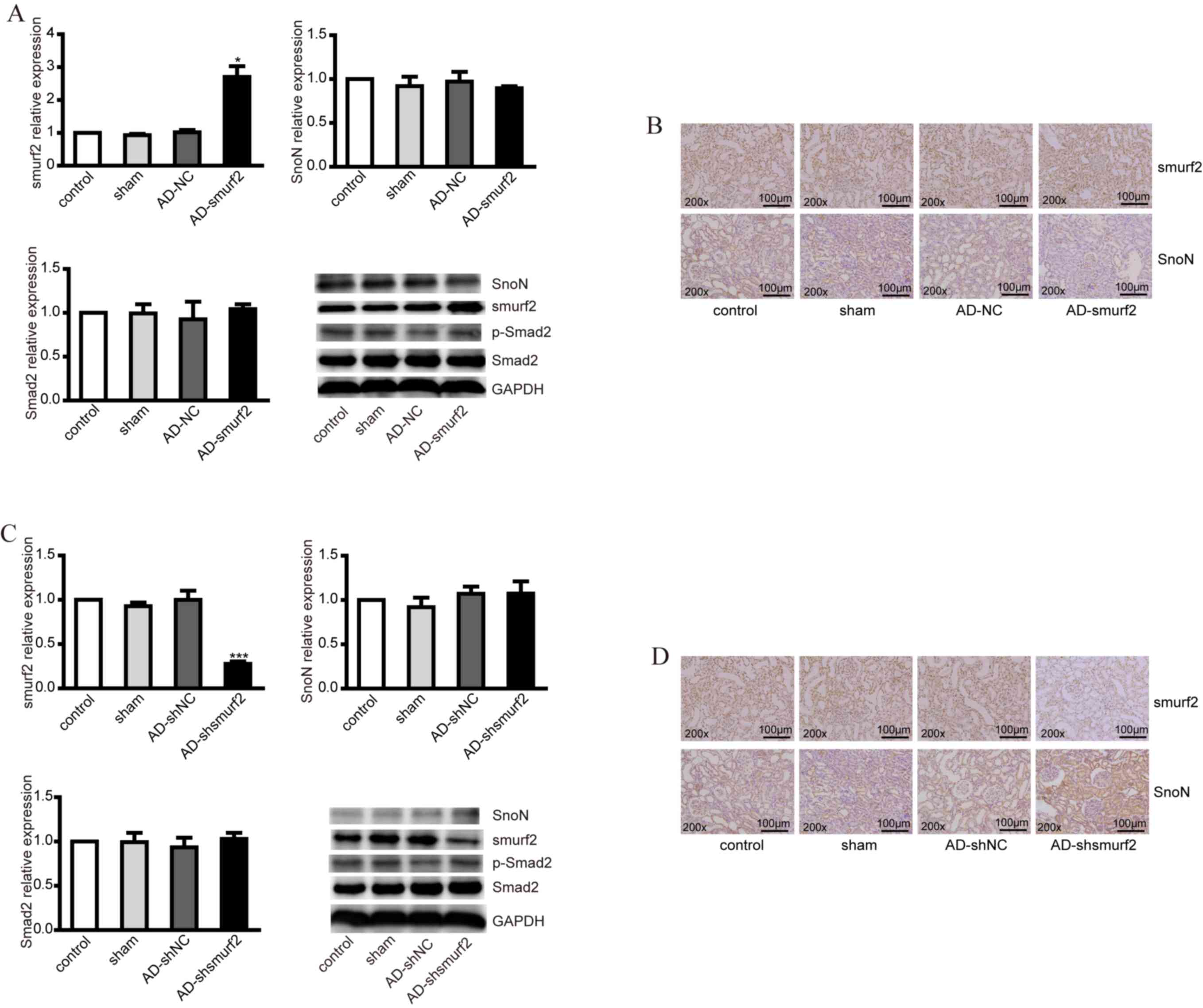 | Figure 3.Expression levels of smurf2, SnoN and
p-Smad2 in DM rats injected with recombinant adenovirus. (A)
RT-qPCR analysis was performed to detect the mRNA levels of smurf2,
SnoN and Smad2, and western blot analysis was performed to detect
the expression levels of smurf2, SnoN and p-Smad2 in animal models
established via injection of recombinant adenovirus AD-smurf2 into
DM rats. (B) Immunohistochemical staining showed the expression of
smurf2 and SnoN in renal tubules of DM rats in different groups.
(C) RT-qPCR analysis was performed to detect the mRNA levels of
smurf2, SnoN and Smad2 and western blot analysis was performed to
detect the expression levels of smurf2, SnoN and p-Smad2 in the DM
rats injected with recombinant adenovirus AD-shsmurf. (D)
Immunohistochemical staining showed the expression of smurf2 and
SnoN in renal tubules of the DM rats in different groups.
*P<0.05 and ***P<0.001, vs. control. DM, diabetes mellitus;
SnoN, ski-related novel protein N; p-Smad2, phosphorylated small
mothers against decapentaplegic; smurf2, Smad ubiquitination
regulatory factor 2; AD, adenovirus; NC, negative control; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction. |
In addition, the DM rats were injected with
adenovirus AD-shsmurf2 to knockdown the expression of smurf2 in
vivo. However, no significant differences were observed in SnoN
or Smad2 at mRNA level following RT-qPCR analysis (Fig. 3C). From the western blot assays, it
was found that the decreased expression of smurf2 increased the
protein level of SnoN; however, no alterations were observed in the
expression of p-Smad2 (Fig. 3C).
The IHC staining showed a higher expression level of SnoN, compared
with control group (Fig. 3D).
TGF-β1/Smad signaling is required for
the degradation of SnoN
Although it was confirmed that the degradation of
SnoN is important in the development of DN and that smurf2 was the
key factor in SnoN degradation by ubiquitination, the mechanism of
SnoN degradation remains to be fully elucidated. The TGF-β1/Smad
signaling pathway is considered to be pathway associated with renal
hypertrophy and tubulointerstitial fibrosis, whereas SnoN is the
key negative regulator of TGF-β1/Smad and it has been widely
recognized that SnoN has the ability to inhibit the signaling
pathway (7–9). To verify the role of the TGF-β1/Smad
signaling pathway in the degradation of SnoN, the present study
treated NRK-52E with 2 ng/ml TGF-β1. The results indicated that
TGF-β1 did not affect the mRNA levels of smurf2, SnoN or Smad2
(Fig. 4A), however, the ectopic
expression of TGF-β1 resulted in decreased expression of SnoN and
increased expression of p-Smad2 following treatment with TGF-β1 for
4 h (Fig. 4A).
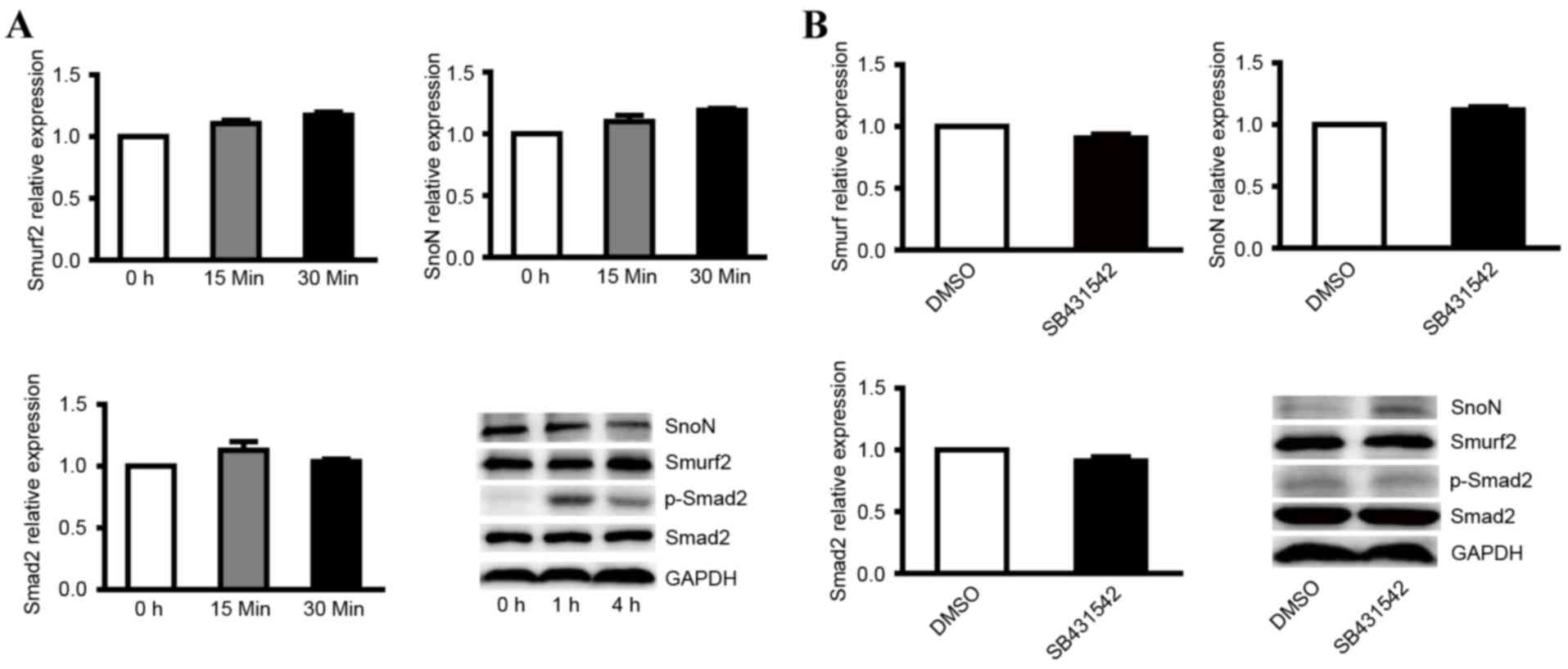 | Figure 4.Expression levels of smurf2, SnoN and
p-Smad2 in NRK-52E cells treated with TGF-β1 and its inhibitor. (A)
RT-qPCR analysis was performed to detect the mRNA levels of smurf2,
SnoN and Smad2, and western blot analysis was performed to detect
the expression levels of smurf2, SnoN and p-Smad2 in NRK-52E cells
treated with TGF-β1 at different time points. (B) RT-qPCR analysis
was performed to detect the mRNA levels of smurf2, SnoN and Smad2,
and western blot analysis was performed to detect the expression
levels of smurf2, SnoN and p-Smad2 in NRK-52E cells treated with
TGF-β1 inhibitor. SnoN, ski-related novel protein N; p-Smad2,
phosphorylated small mothers against decapentaplegic; smurf2, Smad
ubiquitination regulatory factor 2; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; TGF-β1,
transforming growth factor-β1. |
In order to verify the expression level of active
genes following inhibition of the TGF-β1/Smad signaling pathway,
NRK-52E cells were also treated with the TGF-β1 kinase inhibitor,
SB431542, which specifically targets the TGF-β1 receptor. The
results indicated that the TGF-β1 kinase inhibitor did not affect
the mRNA levels of smurf2, SnoN or Smad2 in cells treated with 5 µM
SB431542 (Fig. 4B). Western blot
analysis showed that inhibiting the TGF-β1/Smad signaling pathway
enhanced the expression of SnoN and decreased the expression of
p-Smad2 (Fig. 4B). These results
suggested that the TGF-β1/Smad signaling is indispensable in the
degradation of SnoN by ubiquitination mediated by smurf2.
Discussion
DN is one of the most important microvascular
complication of diabetes and is the leading cause of end-stage
kidney failure worldwide, contributing to morbidity and mortality
rates (14). Due to the complexity
of metabolic disorders, the treatment of DN is more difficult,
compared with other kidney diseases. As the pathogenesis of DN
remains to be fully elucidated and there is no effective drugs, it
is important to investigate the pathogenesis for the prevention and
further treatment of DN. Fukasawa et al (15) suggested that the reduction in the
protein expression of SnoN is critical in the progression of
obstructive nephropathy and results in dysregulation of the
TGF-β1/Smad signaling pathway, which is important in the
pathogenesis of renal fibrosis. However, ureteral obstruction is
the initiating factor of obstructive nephropathy accompanied with
rapid onset and apparent inflammatory reaction. Hyperglycemia is
the initiating factor of DN, in addition to glucose metabolic
disturbance, slow onset and marginal inflammatory reaction, and DN
has a more complex pathogenesis and higher morbidity rates,
compared with obstructive nephropathy. There have been no reports
on whether the progression of DN is associated with the protein
expression of SnoN. In a DM rat model, it was found that the
protein expression of SnoN was significantly downregulated and
showed a negative correlation with smurf2 in the rat renal tissue
(16,17). However, there was no apparent
difference in the mRNA expression of SnoN between DM rat and normal
rat, indicating that downregulation of the protein expression of
SnoN was possibly caused by degradation mediated by smurf2. To
verify this, the present study infected NRK-52E or rat renal
tissues with adenovirus to knock down or overexpress smurf2, and
then detected the protein expression of SnoN, the level of which
was decreased following infection.
In general, the activated TGF-β1 signaling pathway
causes degradation of SnoN protein and leads to the activation of
TGF-β1 downstream targets. The protein expression of SnoN is then
increased and affects the transcriptional activity of the Smad
complex (18). It has been
reported that the activation of TGF-β1 reduces the half-life of
SnoN from 4 h to 45 min through degradation by ubiquitination, in
which E3 ubiquitin ligase was involved. There are different
ubiquitination pathways between cells and diseases (9). TGF-β1 is important in the process of
DN. SnoN effectively inhibits TGF-β1/Smad signaling activity
through several mechanisms (19,20).
Several reports have indicated that SnoN directly interacts with
intranuclear Smad and inhibits the gene transcriptional activity
induced by Smad; secondly, SnoN recruits other nuclear
transcription co-repressors, including nuclear hormone receptor
co-repressor; finally, SnoN prevents Smad from binding to the
uclear transcription co-activator, p300/CBP. As a significant
negative regulator of the TGF-β1/Smad signaling pathway, SnoN
inhibits the target gene of TGF-β1 pre-transcriptionally and
decreases the potency and duration of TGF-β1/Smad signaling.
Although it is generally recognized that SnoN specifically inhibits
the TGF-β1/Smad signaling pathway and decreased SnoN protein is
important in the process of DN, the mechanism remains to be fully
elucidated. The present study treated NRK-52E with TGF-β1 or TGF-β1
inhibitor and detected the gene expression of the TGF-β1/Smad
signaling pathway. The results indicated that the TGF-β1/Smad
negatively regulated the protein expression of SnoN, demonstrating
that the degradation of SnoN by smurf2 was mediated by the
TGF-β1/Smad signaling pathway during the process of DN.
In the present study, NRK-52E cells and DM rat renal
tissues were also infected with adenovirus to knock down or
overexpress smurf2, to demonstrate that the degradation of SnoN
protein by ubiquitination was mediated by smurf2. The NRK-52E cells
were also infected with TGF-β1 or its inhibitor, and it was
confirmed that the reduction in nuclear transcription of the
co-repressor SnoN protein was regulated by the TGF-β1/Smad
signaling pathway during the process of DN. These results provide a
theoretical basis for the treatment of DN.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. 81300607).
References
|
1
|
Standards of medical care in
diabetes-2012. Diabetes Care. 35:(Suppl 1). S11–S63. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
InterAct Consortium: Dietary fibre and
incidence of type 2 diabetes in eight European countries: The
EPIC-InterAct Study and a meta-analysis of prospective studies.
Diabetologia. 58:1394–1408. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wan J, Li P, Liu DW, Chen Y, Mo HZ, Liu
BG, Chen WJ, Lu XQ, Guo J, Zhang Q, et al: GSK-3β inhibitor
attenuates urinary albumin excretion in type 2 diabetic db/db mice,
and delays epithelial-to-mesenchymal transition in mouse kidneys
and podocytes. Mol Med Rep. 14:1771–1784. 2016.PubMed/NCBI
|
|
4
|
Vallon V and Thomson SC: Renal function in
diabetic disease models: The tubular system in the pathophysiology
of the diabetic kidney. Annu Rev Physiol. 74:351–375. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tang SC, Leung JC and Lai KN: Diabetic
tubulopathy: An emerging entity. Contrib Nephrol. 170:124–134.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bottinger EP and Bitzer M: TGF-beta
signaling in renal disease. J Am Soc Nephrol. 13:2600–2610. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Krakowski AR, Laboureau J, Mauviel A,
Bissell MJ and Luo K: Cytoplasmic SnoN in normal tissues and
nonmalignant cells antagonizes TGF-beta signaling by sequestration
of the Smad proteins. Proc Natl Acad Sci USA. 102:12437–12442.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang J, Zhang X, Li Y and Liu Y:
Downregulation of Smad transcriptional corepressors SnoN and Ski in
the fibrotic kidney: An amplification mechanism for TGF-beta1
signaling. J Am Soc Nephrol. 14:3167–3177. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Deheuninck J and Luo K: Ski and SnoN,
potent negative regulators of TGF-beta signaling. Cell Res.
19:47–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cai Y, Zhou CH, Fu D and Shen XZ:
Overexpression of Smad ubiquitin regulatory factor 2 suppresses
transforming growth factor-β mediated liver fibrosis. J Dig Dis.
13:327–334. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Inoue Y and Imamura T: Regulation of
TGF-beta family signaling by E3 ubiquitin ligases. Cancer Sci.
99:2107–2112. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Meng XM, Chung AC and Lan HY: Role of the
TGF-β/BMP-7/Smad pathways in renal diseases. Clin Sci (Lond).
124:243–254. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ortiz-Muñoz G, Mallavia B, Lopez-Franco O,
Hernandez-Vargas P, Egido J and Gomez-Guerrero C: Renal delivery of
adenovirus and antisense oligonucleotides in rats by retrograde
renal vein injection. Methods Mol Biol. 886:321–329. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Marchant V, Droguett A, Valderrama G,
Burgos ME, Carpio D, Kerr B, Ruiz-Ortega M, Egido J and Mezzano S:
Tubular overexpression of Gremlin in transgenic mice aggravates
renal damage in diabetic nephropathy. Am J Physiol Renal Physiol.
309:F559–F568. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fukasawa H, Yamamoto T, Togawa A, Ohashi
N, Fujigaki Y, Oda T, Uchida C, Kitagawa K, Hattori T, Suzuki S, et
al: Ubiquitin-dependent degradation of SnoN and Ski is increased in
renal fibrosis induced by obstructive injury. Kidney Int.
69:1733–1740. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rui-Xia L, Bing G, Ying X, Ming-jun S and
Yuan-yuan W: Expression of Smurf 2 in kidney of diabetic rat and
its relationship with downregulation of SnoN. Chinese Journal of
Pathophysiology. 26:1743–1748. 2010.
|
|
17
|
Liu R, Wang Y, Xiao Y, Shi M, Zhang G and
Guo B: SnoN as a key regulator of the high glucose-induced
epithelial-mesenchymal transition in cells of the proximal tubule.
Kidney Blood Press Res. 35:517–528. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tecalco-Cruz AC, Sosa-Garrocho M,
Vázquez-Victorio G, Ortiz-García L, Domínguez-Hüttinger E and
Macías-Silva M: Transforming growth factor-β/SMAD Target gene SKIL
is negatively regulated by the transcriptional cofactor complex
SNON-SMAD4. J Biol Chem. 287:26764–26776. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang J, Dai C and Liu Y: A novel mechanism
by which hepatocyte growth factor blocks tubular epithelial to
mesenchymal transition. J Am Soc Nephrol. 16:68–78. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Javelaud D, van Kempen L, Alexaki VI, Le
Scolan E, Luo K and Mauviel A: Efficient TGF-β/SMAD signaling in
human melanoma cells associated with high c-SKI/SnoN expression.
Mol Cancer. 10:22011. View Article : Google Scholar : PubMed/NCBI
|