Introduction
Bladder cancer (BC) is the fifth most common type of
malignant tumor worldwide (1) and
one of the most frequently occurring in the urinary system. It is
predominantly present in Europe, North America and Australia, with
approximately 420,000 newly diagnosed cases each year and a leading
cause of cancer-associated mortality (2,3). Of
all urinary malignancies, BC is the primary cause of mortality in
China (4,5).
A number of studies have revealed that tobacco smoke
(TS), the environment and diet are primary risk factors for BC, in
addition to drinking water contaminants, including chlorinated
byproducts and arsenic, the use of pioglitazone, obesity,
hypertension and diabetes (2,6,7). A
previous study reported that 23% of female and 50% of male BC cases
have been attributed to TS (8). It
has been reported that current tobacco smokers have a fourfold
greater risk of developing BC, compared with non-smokers (9). Progress in the understanding of the
molecular mechanisms underlying the initiation and progression of
BC has been made; however, the molecular pathogenesis remains to be
fully elucidated.
Epithelial-mesenchymal transition (EMT) is an
important mechanism in embryonic progression and cancer development
(10). Cells progressively lose
their epithelial characteristics and acquire mesenchymal features
during the process of EMT (11).
In addition to facilitating tumor invasion and metastasis, EMT is
also involved in the initiation of tumorigenesis by promoting cell
malignant transformation. TS has been previously demonstrated to
promote the initial progression of EMT (12–14).
TS-triggered EMT has been revealed to regulate early events in
carcinogenesis, including downregulation of E-cadherin, loss of
cell-cell adhesion and increased mobility of cells. However, the
underlying molecular mechanisms by which TS induces EMT remain to
be fully elucidated.
Extracellular signal-regulated kinase (ERK) 5 is the
least studied member of the mitogen-activated protein kinase (MAPK)
family, and is implicated in important cellular processes,
including gene expression, proliferation, apoptosis, angiogenesis,
cell motility and differentiation (15–18).
A number of studies have investigated the functional role of ERK5
in cancer oncogenesis; however, its role in EMT regulation requires
further investigation. It has been reported that ERK5 promotes EMT
(19–21) and triggers a motility and invasive
phenotype of cells (22–24). Additional studies have suggested a
differential regulatory role of ERK5 in EMT (25,26).
The present study, to the best of our knowledge, is
the first to investigate the role of ERK5 in TS-induced urocystic
EMT in vivo. The study was designed to investigate ERK5
regulation of TS-induced EMT in bladder tissue of mice, using in
vivo TS exposure models. The findings of the present study
suggest that ERK5 may regulate TS-mediated urocystic EMT and
provide insight into the underlying molecular mechanisms of
TS-associated bladder tumorigenesis.
Materials and methods
Chemicals and reagents
XMD8-92 was purchased from Tocris Bioscience
(Bristol, UK). Primary antibodies, phosphorylated (p)-ERK5 (cat.
no. 3371S), p-c-Jun (cat. no. 9164S), p-c-Fos (cat. no. 5348),
E-cadherin (cat. no. 3195S), N-cadherin (cat. no. 4061) and
Vimentin (cat. no. 3932S) were all obtained from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Antibodies for zonula
occludens (ZO)-1 (cat. no. sc-8146) and GAPDH (cat. no. sc-20357)
were from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). Primers
for E-cadherin, ZO-1, N-cadherin, Vimentin and GAPDH were
synthesized by Invitrogen; Thermo Fisher Scientific, Inc. (Waltham,
MA, USA).
Mice and TS exposure
Male BALB/c mice (weight, 18–22 g; age, 8 weeks;
n=12) were purchased from the Animal Research Center of Nanjing
Medical University (Nanjing, China). Mice were group-housed in
polypropylene cages, maintained on a 12 h light/dark cycle, at
22±0.5°C room temperature, at 40–60% humidity with free access to
water and the AIN-76A diet. Animals were handled in accordance with
the recommendations in the guidelines of the Animal Care and
Welfare Committee of Nanjing Medical University. The present study
protocol was approved by the Committee of the Ethics of Animal
Experiments of Nanjing Medical University.
The control group (n=6) was exposed to filtered air
(FA), and the TS-exposure group (n=6) was exposed to TS in a
smoking apparatus designed by the authors. One commercial cigarette
(Hongtashan, one of the most consumed cigarettes in China, which
contains 12 mg tar and 1.1 mg nicotine per cigarette) was combusted
to generate TS by a smoke machine at a constant rate (each
cigarette took ~5 min to burn out). The smoke was delivered to
whole-body exposure chambers with a target concentration of total
particulate matter (TPM) of 80 mg/m3. Mice were exposed
for 6 h daily for 12 weeks. Exposures were monitored and
characterized as follows: For the control group, carbon monoxide
was at 13.98±2.65 mg/m3 and TPM was at 0
mg/m3; and for the TS-exposure group, carbon monoxide
was at 168.77±19.36 mg/m3 and TPM was at 81.05±3.82
mg/m3. Following the final TS exposure, mice were
sacrificed by exposure to 20% CO2 and the bladder
tissues were isolated, frozen and stored at −80°C until
analysis.
Delivery of ERK5 inhibitor in
mice
A total of 24 mice were divided into four groups
(n=6/group) as follows: i) FA-exposure group, in which mice were
exposed to FA; ii) TS-exposure group, in which mice were exposed to
TS; iii) TS + dimethyl sulfoxide (DMSO) group, in which mice were
injected with 15 µl DMSO and exposed to TS; and iv) TS + XMD8-92,
in which mice were injected with XMD8-92 and exposed to TS. XMD8-92
was reconstituted in DMSO and injected intraperitoneally (2 mg/kg
body weight) every other day. Mice were weighed weekly. Following
the completion of exposure, mice were sacrificed and bladder
tissues were collected for analysis.
Western blot analysis
Proteins were extracted from bladder tissues (~200
mg) using a lysate buffer (5 mmol/l EDTA, 50 mmol/l Tris, 1% SDS,
pH 7.5, 10 µg/ml aprotinin, 1% sodium deoxycholate, 1% NP-40, 1 mM
PMSF, 1% Triton-X 100, and 10 µg/ml leupeptin). Protein
concentrations were quantified using a BCA Protein Assay kit
(Pierce; Thermo Fisher Scientific, Inc.). Protein (60 µg/lane) was
loaded on 10% SDS-PAGE and then transferred to polyvinylidene
membranes (EMD Millipore, Billerica, MA, USA). The membranes were
blocked by 5% (w/v) non-fat milk for 1 h at room temperature and
subsequently probed with primary antibodies p-ERK5 (1:500), p-c-Jun
(1:500), p-c-Fos (1:500), E-cadherin (1:1,000), N-cadherin (1:500),
Vimentin (1:1,000) and ZO-1 (1:500), GAPDH (1:5,000) overnight at
4°C, and then incubated with a horseradish peroxidase-conjugated
goat anti-mouse (1:5,000) and goat anti-rabbit (1:10,000) secondary
antibodies. Immunoreactive bands were detected by enhanced
chemiluminescence (GE Healthcare Life Sciences, Chalfont, UK).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA was isolated from frozen bladder tissues (~100
mg) using the RNAiso Plus kit according to the manufacturer's
protocol (Takara Bio, Inc., Otsu, Japan). RNA (2 µg) was reverse
transcribed into cDNA using AMV Reverse Transcriptase (Promega
Corporation, Madison, WI, USA). cDNA was analyzed by PCR using the
Power SYBR-Green Master Mix (Takara Bio, Inc.). The thermal cycling
profile for PCR was 94°C for 5 min, followed by 36 cycles of 30 sec
at 94°C, with 30 sec annealing intervals at their correct
temperatures (56–60°C) and 30 sec at 72°C. The primers used were as
follows: E-cadherin, forward 5′-TCGACACCCGATTCAAAGTGG-3′, reverse
5′-TTCCAGAAACGGAGGCCTGAT-3′; ZO-1, forward
5′-GCAGCCACAACCAATTCATAG-3′, reverse 5′-GCAGACGATGTTCATAGTTTC-3′;
Vimentin, forward 5′-CCTTGACATTGAGATTGCCA-3′, reverse
5′-GTATCAACCAGAGGGAGTGA-3′; N-cadherin, forward
5′-ATCAAGTGCCATTAGCCAAG-3′, reverse 5′-CTGAGCAGTGAATGTTGTCA-3′; and
GAPDH, forward 5′-GCTGCCCAACGCACCGAATA-3′, reverse
5′-GAGTCAACGGATTTGGTCGT-3′; GAPDH served as a control. Fold
alterations in gene expression were calculated by a comparative
threshold cycle (Cq) method using the formula 2−ΔΔCq
(27,28).
Statistical analysis
Data are expressed as the mean ± standard deviation.
Statistical analyses were performed with SPSS software, version
16.0 (SPSS, Inc., Chicago, IL, USA). One-way analysis of variance
was used for comparison of statistical differences of multiple
groups, followed by the least significant difference test. An
unpaired Student's t-test was used for the comparison between two
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
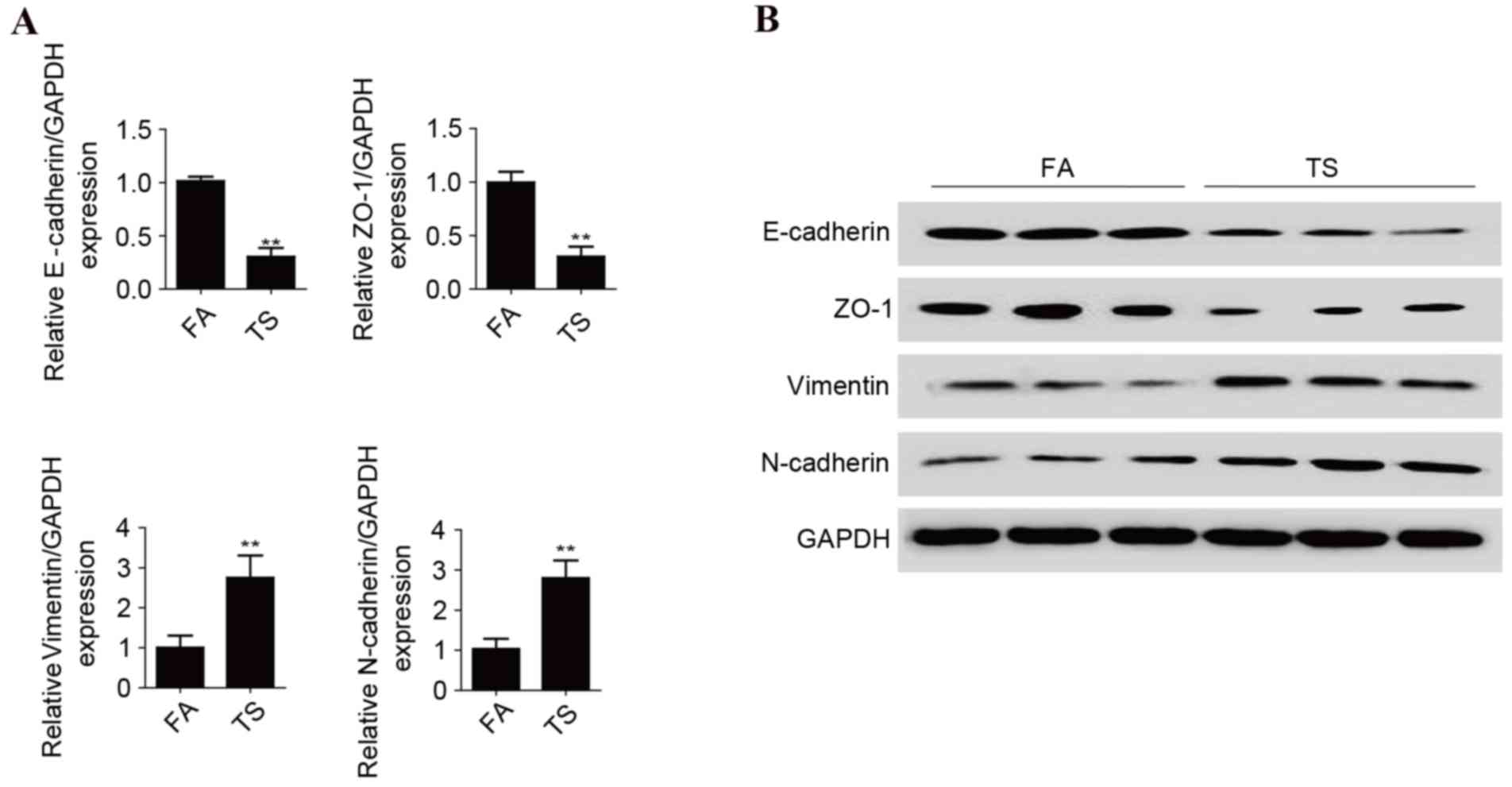
TS exposure alters the expression of
EMT markers in bladder tissues of mice
TS is one of the key risk factors for BC, and
TS-induced EMT is important in TS-associated malignant
transformation. The present study investigated whether TS exposure
induces EMT in bladder tissues. BALB/c mice were exposed to TS for
12 weeks, and the expression of the epithelial and mesenchymal
markers in the bladders of mice were examined. RT-qPCR results
revealed that TS exposure decreased the mRNA expression levels of
E-cadherin and ZO-1. Conversely, the mRNA expression levels of
Vimentin and N-cadherin were increased (P<0.01 vs. FA control;
Fig. 1A). TS exposure reduced
E-cadherin and ZO-1 protein expression levels, and elevated
Vimentin and N-cadherin protein levels, as demonstrated by western
blot analysis (Fig. 1B).
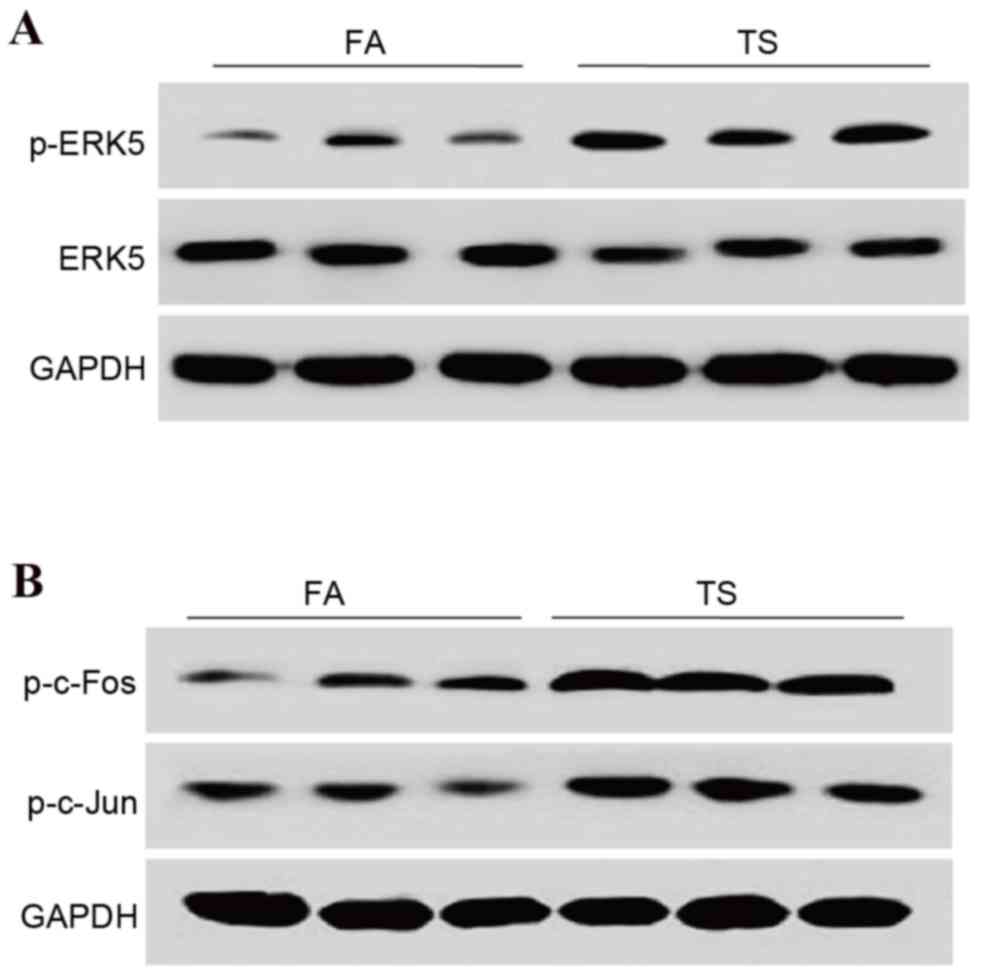
TS increases ERK5/activator protein
(AP)-1 activation in bladder tissue
To determine if TS-induced bladder EMT alterations
are associated with ERK5 activation, the expression levels of
p-ERK5 were measured. It was demonstrated that TS exposure
activated the urocystic ERK5 pathway (Fig. 2A). TS exposure also increased AP-1
protein expression in the bladder of mice, as indicated by elevated
levels of p-c-Jun and p-c-Fos (Fig.
2B). The data therefore suggested that ERK5 activity may be
important in TS-elicited EMT in bladder tissue.
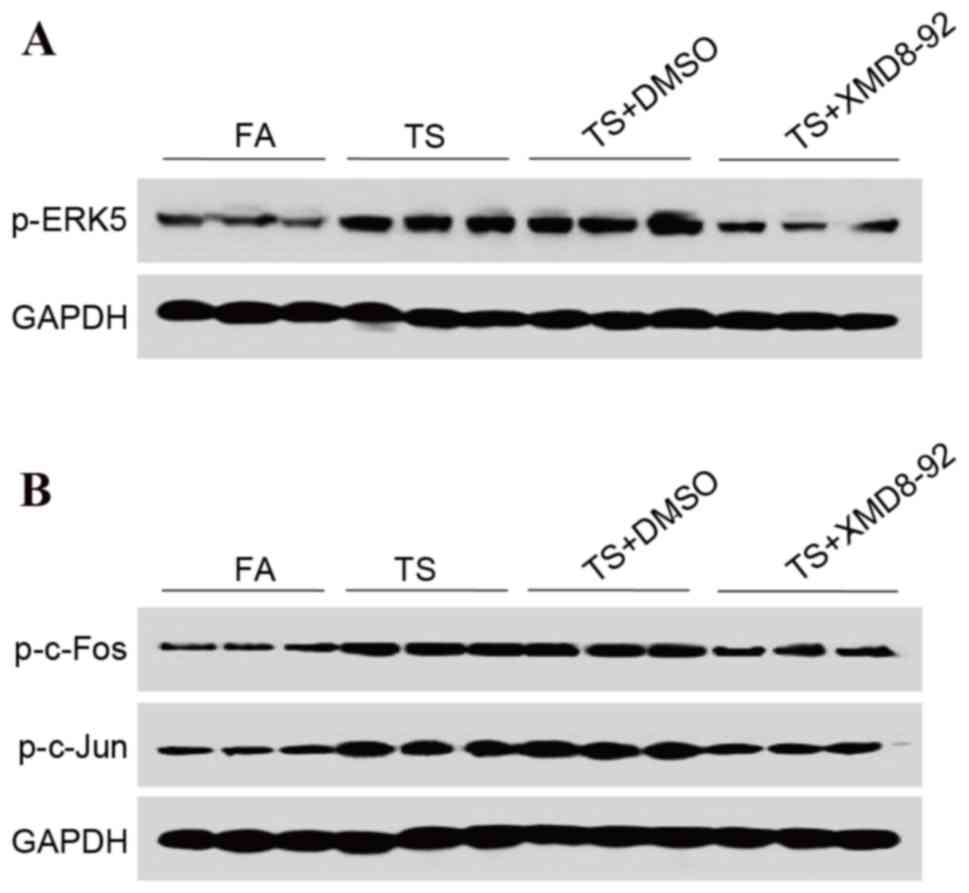
XMD8-92 attenuates TS-induced ERK5
activation in bladder tissue
The aforementioned results revealed that TS-induced
urocystic EMT was associated with ERK5 activation; therefore, the
present study aimed to further determine the role of ERK5 in
urocystic EMT regulation. Mice were treated with XMD8-92 (2 mg/kg
body weight), a highly specific ERK5 inhibitor that suppresses ERK5
activation. Western blot analyses revealed that XMD8-92
downregulated p-ERK5 expression levels (Fig. 3A). In addition, treatment with
XMD8-92 markedly decreased TS-induced AP-1 activation (Fig. 3B).
ERK5 suppression reverses TS-triggered
urocystic EMT
To further determine the role of ERK5 in
TS-triggered EMT in the bladder tissue of mice, the expression of
the EMT markers were examined. ERK5 suppression reversed TS-induced
alterations in the mRNA levels of E-cadherin, ZO-1, Vimentin
(P<0.01 vs. FA control; P<0.001 vs. TS) and N-cadherin
(P<0.01 vs. FA control; P<0.05 vs. TS; Fig. 4A). Western blot analyses
demonstrated that XMD8-92 treatment attenuated both the TS-induced
decrease of E-cadherin and ZO-1 levels, and the increase of
Vimentin and N-cadherin in the bladders of the mice (Fig. 4B). These data demonstrated the
regulatory role of ERK5 in TS-induced EMT in the bladder
tissue.
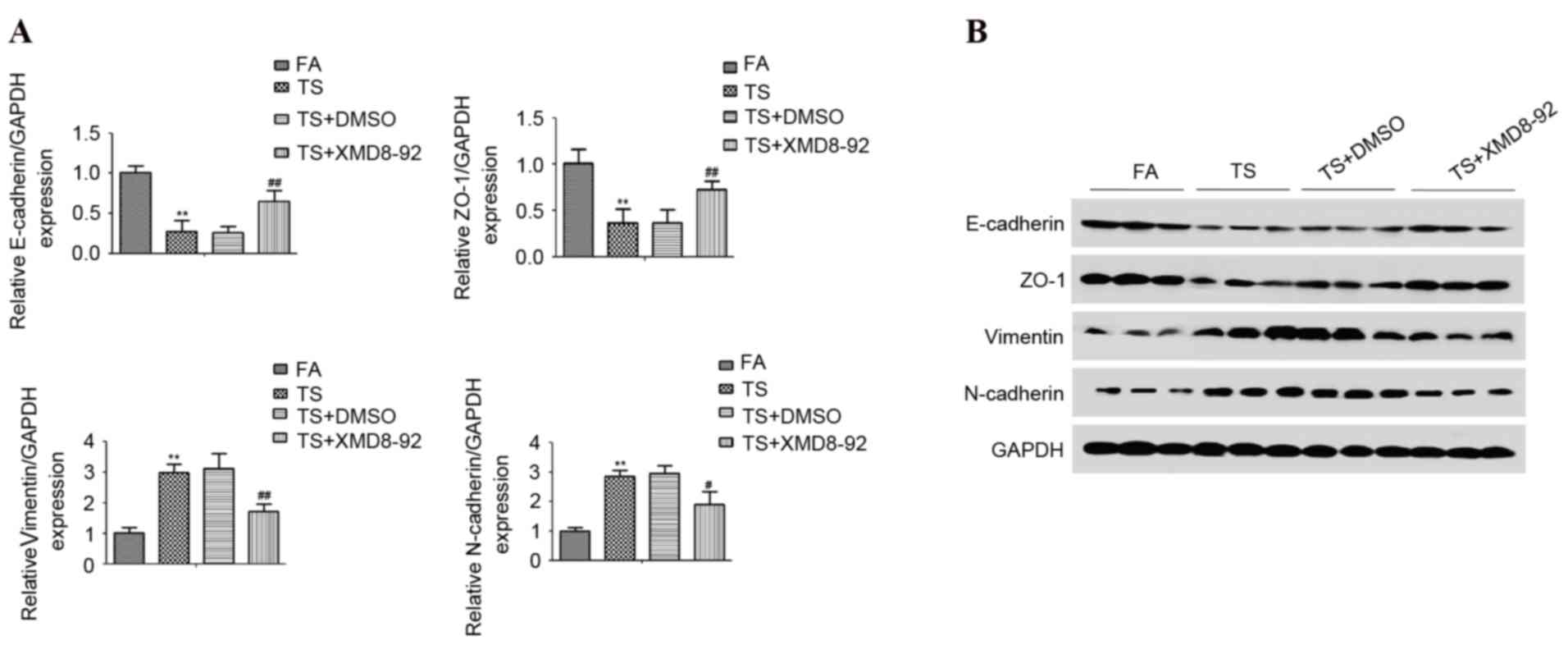 | Figure 4.ERK5 suppression reverses TS-induced
urocystic epithelial-mesenchymal transition alterations in mice.
(A) Reverse transcription-quantitative polymerase chain reaction
analyses of E-cadherin, ZO-1, Vimentin and N-cadherin mRNA. (B)
Western blot analyses of E-cadherin, ZO-1, Vimentin and N-cadherin
proteins. Data are expressed as the mean ± standard deviation.
**P<0.01 vs. FA control; #P<0.05 vs. TS;
##P<0.001 vs. TS. DMSO, dimethyl sulfoxide; ERK,
extracellular signal-regulated kinase; FA, filtered air; TS,
tobacco smoke; XMD8-92, a highly specific ERK5 inhibitor; ZO-1,
zonula occludens-1. |
Discussion
BC is one of the primary causes of cancer-associated
mortality, and an association between the occurrence of BC and TS
has previously been established (27,29).
However, the underlying molecular mechanisms by which TS results in
BC development remain to be fully elucidated. The present study
demonstrated that TS exposure induced EMT in the bladder of mice.
To the best of our knowledge, the present study is the first to
demonstrate that ERK5 regulates TS-mediated bladder EMT in
vivo. The findings suggested that exposure to TS elevated ERK5
activation, and TS-associated bladder EMT alteration was reversed
by ERK5 inhibition. These results explain the important role of
ERK5 in TS-triggered urocystic EMT and furthers our current
understanding of the underlying molecular mechanisms of
TS-associated BC.
It has previously been demonstrated that exposure of
cells to carcinogens induces EMT during tumor transformation and
formation (12–14,30,31),
suggesting the involvement of EMT in the initiation of
tumorigenesis by promoting cell malignant transformation. The
present study revealed that exposure to TS induced EMT in the
bladder of mice. TS exposure altered the expression of EMT markers:
Decreasing the expression levels of epithelial markers E-cadherin
and ZO-1, and increasing the expression of mesenchymal markers
Vimentin and N-cadherin. These results revealed that exposure to TS
triggered urocystic EMT in the bladder of mice.
The underlying mechanisms of EMT induction by TS
exposure remain to be fully elucidated. ERK5 is a lesser-studied
member of the MAPK family. Various reports have suggested a
functional role for ERK5 in cancer oncogenesis, and ERK5 has been
demonstrated to promote EMT (19–21);
however, its specific role in EMT regulation has not yet been
verified. Previous studies have indicated that ERK5 triggers the
motility and invasive phenotype of cells (22–24).
Additional studies have suggested a differential regulatory role of
ERK5 in EMT (25,26). These reports have suggested that
ERK5 regulation of EMT may be sensitive to cell type and/or the
cellular microenvironment.
The function of ERK5 in TS-induced urocystic EMT
in vivo has not yet been investigated. The present study
demonstrated that TS-induced urocystic EMT was associated with an
upregulation of ERK5 activation in the bladders of mice. To
determine the role of ERK5 in urocystic EMT regulation, mice were
treated with XMD8-92 (2 mg/kg body weight), a highly specific ERK5
inhibitor. Inhibition of ERK5 attenuated TS-induced alterations in
EMT markers, including the decreased expression levels of the
epithelial markers E-cadherin and ZO-1, and the increased
expression of the mesenchymal markers Vimentin and N-cadherin.
These data indicated that ERK5 positively regulated TS-induced
urocystic EMT.
In conclusion, the present study demonstrated that
ERK5 positively regulates TS-induced urocystic EMT in vivo.
These findings indicate the important role of ERK5 in TS-associated
carcinogenesis and provide a potential strategy for the search of a
novel interventional target in TS-associated bladder
tumorigenesis.
Acknowledgements
The present study was supported by The Anhui Public
Welfare Research Linkage Plan (grant no. 1501ld04045) and The
National Natural Science Foundation of China (grant no.
81072330).
References
|
1
|
Ding M, Li Y, Wang H, Lv Y, Liang J, Wang
J and Li C: Diagnostic value of urinary microRNAs as non-invasive
biomarkers for bladder cancer: A meta-analysis. Int J Clin Exp Med.
8:15432–15440. 2015.PubMed/NCBI
|
|
2
|
Montella M, Di Maso M, Crispo A, Grimaldi
M, Bosetti C, Turati F, Giudice A, Libra M, Serraino D, La Vecchia
C, Tambaro R, Cavalcanti E, et al: Metabolic syndrome and the risk
of urothelial carcinoma of the bladder: A case-control study. BMC
Cancer. 15:7202015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Babjuk M, Burger M, Zigeuner R, Shariat
SF, Van Rhijn BW, Compérat E, Sylvester RJ, Kaasinen E, Böhle A,
Redorta J Palou, et al: EAU guidelines on non-muscle-invasive
urothelial carcinoma of the bladder: Update 2013. Eur Urol.
64:639–653. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fang H, Yao B, Yan Y, Xu H, Liu Y, Tang H,
Zhou J, Cao L, Wang W, Zhang J and Zhao Y: Diabetes mellitus
increases the risk of bladder cancer: An updated meta-analysis of
observational studies. Diabetes Technol Ther. 15:914–922. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang YY, Wang XL and Yu ZJ: Vitamin C and
E intake and risk of bladder cancer: A meta-analysis of
observational studies. Int J Clin Exp Med. 7:4154–4164.
2014.PubMed/NCBI
|
|
8
|
Letasˇiová S, Medve'ová A, Šovčíková A,
Dušinská M, Volkovová K, Mosoiu C and Bartonová A: Bladder cancer,
a review of the environmental risk factors. Environ Health. 11
Suppl 1:S112012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen LM, Nergard JC, Ni L, Rosser CJ and
Chai KX: Long-term exposure to cigarette smoke extract induces
hypomethylation at the RUNX3 and IGF2-H19 loci in immortalized
human urothelial cells. PLoS One. 8:e655132013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu J, Hu G, Chen D, Gong AY, Soori GS,
Dobleman TJ and Chen XM: Suppression of SCARA5 by Snail1 is
essential for EMT-associated cell migration of A549 cells.
Oncogenesis. 2:e732013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang GX, Cao LP, Kang PC, Zhong XY, Lin
TY and Cui YF: Interleukin-6 induces epithelial-mesenchymal
transition in human intrahepatic biliary epithelial cells. Mol Med
Rep. 13:1563–1569. 2016.PubMed/NCBI
|
|
12
|
Shin VY, Jin HC, Ng EK, Sung JJ, Chu KM
and Cho CH: Activation of 5-lipoxygenase is required for nicotine
mediated epithelial-mesenchymal transition and tumor cell growth.
Cancer Lett. 292:237–245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang L, Gallup M, Zlock L, Basbaum C,
Finkbeiner WE and McNamara NA: Cigarette smoke disrupts the
integrity of airway adherens junctions through the aberrant
interaction of p120-catenin with the cytoplasmic tail of MUC1. J
Pathol. 229:74–86. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang H, Liu H, Borok Z, Davies KJ, Ursini
F and Forman HJ: Cigarette smoke extract stimulates
epithelial-mesenchymal transition through Src activation. Free
Radic Biol Med. 52:1437–1442. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Drew BA, Burow ME and Beckman BS:
MEK5/ERK5 pathway: The first fifteen years. Biochim Biophys Acta.
1825:37–48. 2012.PubMed/NCBI
|
|
16
|
Nishimoto S and Nishida E: MAPK
signalling: ERK5 versus ERK1/2. EMBO Rep. 7:782–786. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hayashi M, Fearns C, Eliceiri B, Yang Y
and Lee JD: Big mitogen-activated protein kinase 1/extracellular
signal-regulated kinase 5 signaling pathway is essential for
tumor-associated angiogenesis. Cancer Res. 65:7699–7706.
2005.PubMed/NCBI
|
|
18
|
Wang X and Tournier C: Regulation of
cellular functions by the ERK5 signalling pathway. Cell Signal.
18:753–760. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou C, Nitschke AM, Xiong W, Zhang Q,
Tang Y, Bloch M, Elliott S, Zhu Y, Bazzone L, Yu D, et al:
Proteomic analysis of tumor necrosis factor-alpha resistant human
breast cancer cells reveals a MEK5/Erk5-mediated
epithelial-mesenchymal transition phenotype. Breast Cancer Res.
10:R1052008. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Antoon JW, Martin EC, Lai R, Salvo VA,
Tang Y, Nitzchke AM, Elliott S, Nam SY, Xiong W, Rhodes LV, et al:
MEK5/ERK5 signaling suppresses estrogen receptor expression and
promotes hormone-independent tumorigenesis. PLoS One. 8:e692912013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Madak-Erdogan Z, Ventrella R, Petry L and
Katzenellenbogen BS: Novel roles for ERK5 and cofilin as critical
mediators linking ERα-driven transcription, actin reorganization
and invasiveness in breast cancer. Mol Cancer Res. 12:714–727.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ramsay AK, McCracken SR, Soofi M, Fleming
J, Yu AX, Ahmad I, Morland R, Machesky L, Nixon C, Edwards DR, et
al: ERK5 signalling in prostate cancer promotes an invasive
phenotype. Br J Cancer. 104:664–672. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim SM, Lee H, Park YS, Lee Y and Seo SW:
ERK5 regulates invasiveness of osteosarcoma by inducing MMP-9. J
Orthop Res. 30:1040–1044. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shukla A, Miller JM, Cason C, Sayan M,
MacPherson MB, Beuschel SL, Hillegass J, Vacek PM, Pass HI and
Mossman BT: Extracellular signal-regulated kinase 5: A potential
therapeutic target for malignant mesotheliomas. Clin Cancer Res.
19:2071–2083. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zuo Y, Wu Y, Wehrli B, Chakrabarti S and
Chakraborty C: Modulation of ERK5 is a novel mechanism by which
Cdc42 regulates migration of breast cancer cells. J Cell Biochem.
116:124–132. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Woo CH, Massett MP, Shishido T, Itoh S,
Ding B, McClain C, Che W, Vulapalli SR, Yan C and Abe J: ERK5
activation inhibits inflammatory responses via peroxisome
proliferator-activated receptor delta (PPARdelta) stimulation. J
Biol Chem. 281:32164–32174. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liang Z, Xie W, Wu R, Geng H, Zhao L, Xie
C, Li X, Zhu M, Zhu W, Zhu J, et al: Inhibition of tobacco
smoke-induced bladder MAPK activation and epithelial-mesenchymal
transition in mice by curcumin. Int J Clin Exp Pathol. 8:4503–4513.
2015.PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Geng H, Zhao L, Liang Z, Zhang Z, Xie D,
Bi L, Wang Y, Zhang T, Cheng L, Yu D and Zhong C: ERK5 positively
regulates cigarette smoke-induced urocystic epithelial-mesenchymal
transition in SV-40 immortalized human urothelial cells. Oncol Rep.
34:1581–1588. 2015.PubMed/NCBI
|
|
30
|
Sun JL, Chen DL, Hu ZQ, Xu YZ, Fang HS,
Wang XY, Kan L and Wang SY: Arsenite promotes intestinal tumor cell
proliferation and invasion by stimulating epithelial-to-mesenchymal
transition. Cancer Biol Ther. 15:1312–1319. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tellez CS, Juri DE, Do K, Bernauer AM,
Thomas CL, Damiani LA, Tessema M, Leng S and Belinsky SA: EMT and
stem cell-like properties associated with miR-205 and miR-200
epigenetic silencing are early manifestations during
carcinogen-induced transformation of human lung epithelial cells.
Cancer Res. 71:3087–3097. 2011. View Article : Google Scholar : PubMed/NCBI
|