Introduction
Reactive oxygen species (ROS) that are generated
in vivo are a major cause of human aging and disease
(1). ROS expression leads to
disease through toxic effects on cells and tissues. ROS include
free radical species, such as superoxide anion
(O2−), hydroxyl radical and singlet oxygen,
as well as non-radical species such as hydrogen peroxide
(H2O2). ROS are neutralized by antioxidant
enzymes, including superoxide dismutase (SOD), catalase (CAT),
glutathione peroxidase (GPx) and glutathione reductase, or by small
molecules, such as ascorbic acid, α-tocopherol and glutathione
(GSH), during normal physiological metabolic processes. Oxidizing
agents and oxidation inhibitors function to maintain
reduction-oxidation (redox) homeostasis in the body. However,
perturbations in this balance, either caused by excess production
of ROS or decreased or malfunctioning antioxidant defenses, may
result in oxidative stress, potentially leading to pathological
effects (1–4). ROS may be detrimental to cells by
inducing oxidative damage to lipids, proteins and DNA (5). Therefore, it is important to balance
ROS levels with endogenous antioxidant enzymes and, when required,
antioxidant compounds.
Oxidative stress has been associated with a number
of diseases, including inflammatory diseases, acquired
immunodeficiency syndrome, gastric ulcers, hypertension,
neurological disorder, alcoholism and smoking-related diseases
(4). Over 90% of ROS are produced
in the mitochondria during cellular respiration. During this
process, most oxygen is converted into water (H2O), and
a one-electron reduction of molecular oxygen (O2)
produces O2−, a primary ROS, which is
subsequently transformed into hydrogen peroxide
(H2O2) by SOD. H2O2 is
an oxygen metabolite of central importance that is produced in the
mitochondria, as well as in other sites in the cell.
H2O2 itself has weak biological reactivity;
however, if present at a cellular concentration of at least 10 µM,
it can directly damage cellular components, including inactivating
enzymes by oxidizing active site thiol groups (2,3).
ROS levels are tightly controlled in the body by
antioxidant enzymes, including SOD, CAT, GPx and thiol-containing
small molecules such as GSH. SODs catalyze the dismutation of
O2− to H2O2 and
O2, and are located in the cytoplasm [copper (Cu)- and
zinc (Zn)-dependent SOD] and in the mitochondria
[manganese-dependent (Mn)-SOD] (6). CAT is a tetrameric iron-porphyrin
protein in peroxisomes that converts H2O2 to
H2O and O2 (7). CAT and Cu/Zn-SOD are expressed
constitutively, whereas Mn-SOD expression within the mitochondria
is induced by oxidative stress. GSH is a sulfhydryl peptide that
may directly react with O2− or
N2-containing free radicals, or is able to donate
electrons in the enzymatic dismutation of
H2O2 to H2O and O2 by
GPx (8).
ROS removal involves the cooperation of 29
associated genes and 15 corresponding gene expression products
(3). The regulation of
intracellular antioxidant activity is regulated by a complex
network of these genes. The expression of antioxidant enzymes is
altered by oxidative stress. The antioxidant defense system within
cells includes ~50 antioxidant-related genes that are divided into
four categories (3–9): i) CAT/SOD family members, which
include the classic antioxidant enzymes CAT and SODs; ii) proteins
involved in GSH metabolism, including GPx, glutaredoxins,
glutathione reductase and glucose-6-phosphate dehydrogenase, which
promote the antioxidant activity of thiol-containing small
molecules, such as GSH, and GSH recycling; iii) proteins involved
in redox balance, including enzymes of thioredoxin (TRX)
metabolism, TRX, TRX reductase (TRR) and peroxiredoxin (PRDX); and
iv) pentose phosphate cycle proteins, including glucose 6-phosphate
dehydrogenase. The present study investigated the antioxidant
effects of a purified protein on the mRNA expression the
antioxidant enzymes SOD, CAT, GPx, and TRR.
Cyclophilin (Cyp) is a multifunctional protein
family, which includes cyclophilin-type peptidylprolyl
cis-trans isomerase (PPI), a protein which may be inhibited
by cyclosporin A (CsA) (10). CsA
generates ROS and lipid peroxidation in cells, which appears to be
directly associated with its pathological effects. Perez et
al (11) used
2′,7′-dichlorofluorescein diacetate (DCF-DA) and demonstrated that
1–10 µM CsA generated an oxidized DCF signal, interpreted as being
H2O2-derived. CsA was demonstrated to
increase lipid peroxidation in the rat kidney and liver in
vivo (12,13). Cyp is a ubiquitous protein that is
present in all subcellular compartments, and therefore may
potentially be involved in a variety of processes, including
protein trafficking and maturation, receptor complex stabilization,
apoptosis, receptor signaling, RNA processing and spliceosome
assembly (14–20). It has been proposed that
CsA-induced oxidative stress may activate or deactivate
transcription factors and thus affect gene transcription.
Similarly, ROS generated by CsA may activate or deactivate various
signaling molecules and influence their downstream transduction
systems. The ROS induced by CsA has been reported to affect
mitogen-activated protein kinase and transforming growth factor-β
signaling (21). In particular,
there is increased expression of TRX 1 via the ERK/MAPK-2
signaling, to overcome the oxidative stress resulting from ROS. In
the present study, PPI, known to exhibit antioxidant activity, was
cloned from Pyropia yezoensis. In addition, hydrogen
peroxide was used to experimentally induce oxidative stress that
results from CsA.
In the sea, the exposure of seaweeds to sunlight and
oxygen leads to formation of ROS; however, no oxidative damage is
evident in their structural and functional components, which
suggests that they may have an efficient antioxidant defense
system. Hence, several seaweed extracts are attracting scientific
interest to identify new and effective antioxidant compounds
(22–27). A recent report described
antioxidant agents isolated from seaweed (22). Ismail and Tan (23) compared the antioxidant capacity of
commercial seaweeds and confirmed high antioxidant activities of
Porphyra sp., Laminaria sp., Undaria sp. and
Hijikia sp. Other studies have described high antioxidative
properties of many compounds isolated from seaweed, including
phlorotannin, fucoxanthin, carotenoids and tocopherols (24–27).
Previous studies on the antioxidant properties of algae focused
primarily on crude extracts (28–30).
In the present study, the gene expression and purification of the
antioxidant protein PPI from P. yezoensis is described. In
addition, the antioxidant activity of recombinant PPI, that is, its
protection against ROS, was investigated in a human hepatocellular
carcinoma cell line.
Materials and methods
Seaweeds and cDNA synthesis
P. yezoensis was collected directly from
Myeongji (Busan, Korea) and stored at −70°C. Frozen samples were
lyophilized using a freezing dryer (−80°C, 24 h) and homogenized
using a blender into powder, prior to mRNA extraction. mRNA (5 µg)
was extracted from 20 mg P. yezoensis with the GeneJET Plant
RNA Purification Mini kit (Thermo Scientific, Inc., Waltham, MA,
USA), according to the manufacturer's protocol. Quantification of
RNA was performed using the NanoDrop 2000 Spectrophotometer (Thermo
Fisher Scientific, Inc., Waltham, MA, USA). The ratio of absorbance
at 260 nm and 280 nm was used to assess the purity of RNA. A ratio
of <2.0 was accepted as pure for RNA. The mRNA was used for
first strand cDNA synthesis and double strand cDNA synthesis using
a PrimeScript Double strand cDNA synthesis kit (Takara Bio, Inc.,
Otsu, Japan) following the manufacturer's protocol.
Preparation and identification of
recombinant PPI protein
To prepare PPI, a forward primer
(5′-GGCCCATATGGGGAACCCGCAGGTGTTCT-3′) containing the Nde1
site (underlined) and initiation codon (italic) and reverse primer
(5′-GGCCCTCGAGGAGCTCGCCGCAGTCCGC-3′) containing Xho1 site
(underlined), were constructed. Polymerase chain reaction (PCR)
amplification was performed using the cDNA of P. yezoensis
(one cycle at 95°C for 5 min; 30 cycles at 95°C for 30 sec, 55°C
for 30 sec, and 72°C for 30 sec; followed by one cycle of 72°C for
7 min) using EXTaq DNA polymerase (Takara Bio Inc., Otsu, Japan).
The PCR products (151 bp) and expression vector pET22b+ were
digested with two restriction enzymes (NdeI and
XhoI). The PCR fragment was subcloned into the digested
pET22b+ vector with a DNA ligation kit (Takara Bio Inc.). The
resulting plasmid was named pETppi. The pETppi was introduced into
Escherichia coli DH5α competent cells, used as the cloning
host for propagation of the expression vector, and finally
retransformed into expression strain E. coli BL21 (DE3),
according to a previously described method (31). The transformation bacteria were
selected on LB agar medium containing 100 µg-ml-1 of ampicillin.
Cultures of the transformed E. coli BL21 overexpressed a
recombinant PPI of the expected molecular mass (~18 kDa), which was
purified by affinity chromatography in Ni-NTA purification system
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. Bacterial cultures were incubated using LB medium at 37°C
until reaching the OD600 of 0.8. For PPI expression,
isopropyl-beta-D-thiogalactopyranoside (IPTG) was added to a final
concentration of 1 mM. Expression was continued for 4 h.
EnterokinaseMax™ enzyme (Thermo Fisher Scientific, Inc.) was used
to separate the 6x His-tag from recombinant PPI protein, according
to the manufacturer's protocol. Analysis of protein expression and
purification were carried out using sodium dodecyl sulfate -poly
acrylamide gel electrophoresis (SDS-PAGE). The purified recombinant
PPI protein was confirmed using SDS-PAGE and electrospray
ionization quadrupole time-of-flight mass spectrometry/mass
spectrometry (ESI-Q-TOF MS/MS), according to a previously described
method (32). The proteins were
identified via an NCBI search using the MASCOT program (http://www.matrixscience.com, Matrixscience, London,
UK).
Cell culture and viability
HepG2 human hepatocellular carcinoma cells were
obtained from the American Type Culture Collection (ATCC, Manassas,
VA, USA). Cells were cultured at 37°C in a humidified 5%
CO2, 95% air equilibrated incubator in minimum essential
medium (MEM) supplemented with heat-inactivated 10% fetal bovine
serum (HyClone, Logan, UT, USA), penicillin (100 U-ml-1) and
streptomycin (100 µg-ml-1). Adherent cells at 70–80% confluence
were detached by trypsin-EDTA solution and re-plated.
Cell viability was estimated with the CytoX Cell
Viability Assay kit (LPS Solution, Daejeon, Korea). Briefly, cells
(1.0×103 cells/well) were seeded in a 96-well plate.
Various concentrations (0.001, 0.01, 0.1 and 1 µg-ml−1)
of recombinant PPI protein were added to the cells along with 1 mM
H2O2 for 1 h at 37°C. Following incubation,
Cyto solution was added to each well and incubated for an
additional 1 h at 37°C. Absorbance was measured with a microplate
reader (BioTek Instruments, Inc., Winooski, VT, USA) at a
wavelength of 450 nm. The viability of purified recombinant PPI
protein-treated cells was expressed as a percentage of that in
negative control cells (−, without H2O2).
Assessment of ROS production
The effects of purified recombinant PPI protein on
ROS production was evaluated using the cell-permeable probe DCF-DA.
DCF-DA (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was
dissolved in 10 mg-ml-1 sterile dimethylsulfoxide and was used at a
concentration of 50 µg-ml-1. HepG2 cells were seeded onto 96-well
plates at 1.0×103 cells/well in MEM, grown until confluence, and
pre-incubated with DCF-DA for 20 min at 37°C in the dark. After
washing twice with phosphate buffered saline (PBS) to remove the
unattached probe, cells were treated with PBS or various
concentrations (between 0.001 and 1 µg-ml-1) of purified
recombinant PPI protein in the presence or absence of 1 mM
H2O2 for 1 h. Fluorescence was measured at
485/20 nm excitation and 535/20 nm emission using a FilterMax F5
Multi-Mode Microplate Reader (Molecular Devices, LLC, Sunnyvale,
CA, USA).
Assessment of antioxidant enzyme
activities
HepG2 cells were grown to 70–80% confluence at 37°C.
Following 4 h starvation, the cells were treated with PBS or
various concentrations (0.001–1 µg-ml-1) of purified recombinant
PPI protein in the presence or absence of 1 mM
H2O2 for 1 h and harvested in lysis buffer
containing 50 mM Tris-HCl (pH 7.5), 150 mM sodium chloride, 0.5%
sodium deoxycholate, 0.1% SDS, 1% triton X-100 and 2 mM
ethylenediaminetetra-acetic acid with inhibitors (1 mM sodium
orthovanadate and 1 mM phenylmethylsulfonyl fluoride). Cell debris
was removed by centrifugation at 13,000 × g at 4°C for 10 min and
the supernatant was used for further measurements. Protein
concentration was determined using the BCA Protein assay (Pierce;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's protocol. Each supernatant contained an equal amount
of protein (5 µg) and was used for the subsequent enzyme activity
assays. The activities of antioxidant enzymes, including CAT, SOD,
GPx and TRR were measured using a Catalase Assay kit
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), SOD Assay Kit-WST
(Sigma-Aldrich; Merck KGaA), Glutathione Peroxidase Cellular
Activity Assay kit (Sigma-Aldrich; Merck KGaA) and Thioredoxin
Reductase Assay kit (Sigma-Aldrich; Merck KGaA) according to the
manufacturer's protocols. The absorbance was measured using a
microplate reader (BioTek Instruments, Inc., Winooski, VT,
USA).
Reverse transcription (RT)-PCR
The effects of recombinant PPI on antioxidant enzyme
mRNA expression in H2O2 treated HepG2 cells
was evaluated by RT-PCR. Total RNA was extracted from HepG2 cells
using TRIzol Reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocols. Quantification of RNA
was performed as described above. The purity of the RNA was
determined at absorbance ratios of 260 and 280 nm (260/280 nm
<2.0). cDNA was synthesized using a First-Strand cDNA Synthesis
kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. In this experiment, primer sets of SOD2,
GPx and CAT were used as reported in Schmidt et al (33) and primer sets of GAPDH and TRR were
designed as described by Aguilar-Melero et al (34). PCR amplification was performed
using the template cDNA (1 ng) and PCR Amplification Kit (Takara
Bio Inc.) Initial denaturation at 95°C for 5 min, followed by 25
cycles of 95°C for 30 sec, 55°C for 30 sec and 72°C for 30 sec,
then a final extension of 72°C for 5 min. Amplified products were
analyzed by 1% agarose gel electrophoresis and stained with
ethidium bromide for detection. The software GeneTools, version
4.03 (SYNGENE, Cambridge, UK) was used for densitometry.
Statistical analysis
Data are expressed as the mean ± standard deviation
and were evaluated by one-way analysis of variance using the
statistical package for social sciences version 10.0 (SPSS, Ins.,
Chicago, IL, USA). Values were compared with controls using
analysis of variance followed by Duncan's multiple range test.
P<0.05 was considered to represent a statistically significant
difference.
Results
Expression and purification of
recombinant PPI protein
The expression vector pETppi of P. yezoensis was
transformed into E. coli BL21 (DE3) and then induced by 1 mM IPTG.
SDS-PAGE analysis of the harvested cells from the pETppi
transformed E. coli (BL21) exhibited high amounts of a
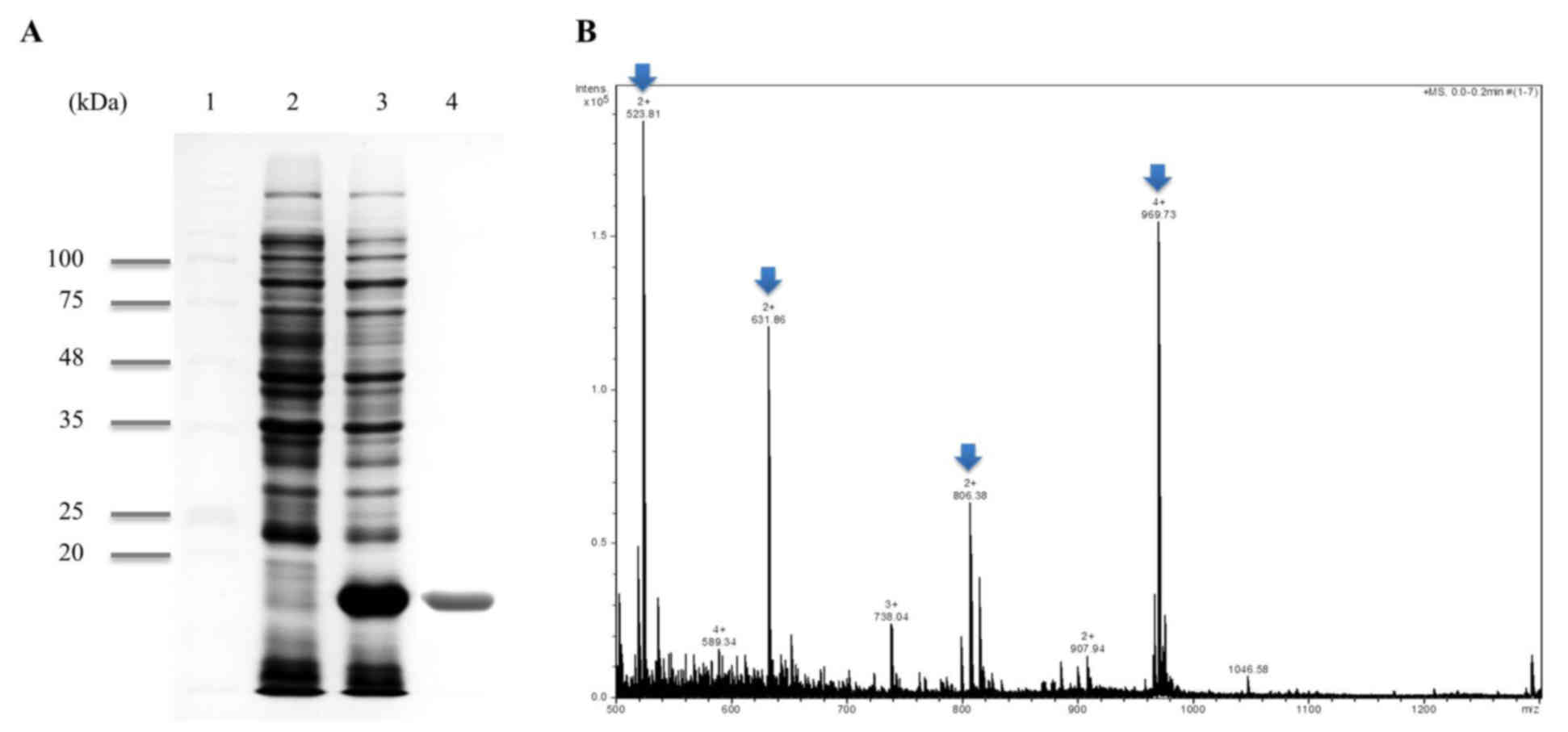
polypeptide with the expected molecular mass of ~18 kDa (Fig. 1A, lane 2) from SDS-PAGE analysis.
Recombinant PPI was purified ~18 kDa protein by the Ni-NTA system
(Fig. 1A, lane 4). It was highly
purified from crude extracts as a His-tagged protein. Recombinant
PPI was identified via MALDI-TOF MS/MS. A total of four peaks from
MALDI-TOF MS were analyzed by MS/MS and the following sequences
were obtained (Fig. 1B, Arrow): i)
VFFDMTIGGAPAGR; ii) VITDF MCQGGDFTR; iii) ADENFTLTHTGPGVLSMANAGK;
and iv) NGSQFFLTTVK. This was 100% homologous to PPI (accession
number KJ728870.1) using the MASCOT program and NCBI database.
Cell viability
Cell viability was determined with the CytoX assay,
which relies on the mitochondrial metabolic capacity of viable
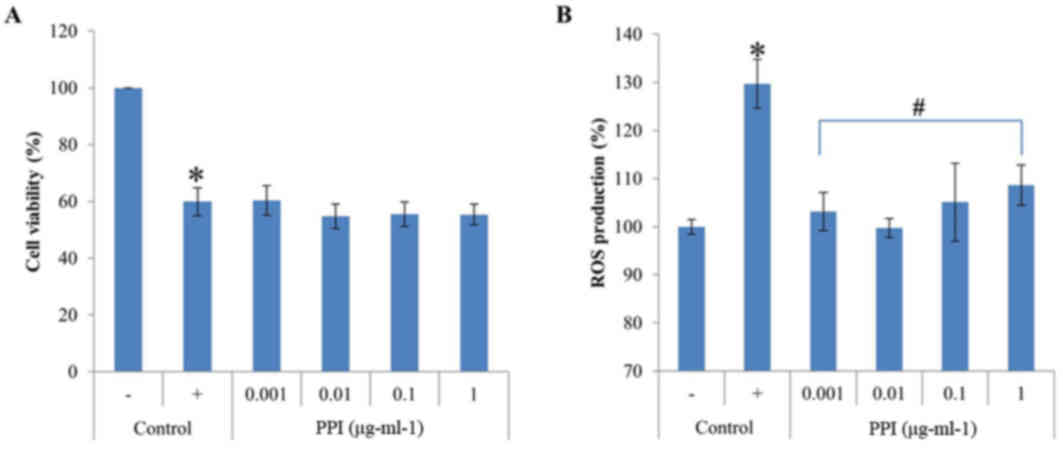
cells. The results revealed that recombinant PPI exposure was not
cytotoxic to HepG2 cells at any of the concentrations examined,
between 0.001 and 1 µg-ml-1 (Fig.
2A). Treatment with H2O2 for 1 h
decreased cell viability to 59.9% that of control cells;
co-treatment with recombinant PPI did not alter the
H2O2-induced decrease in cell viability,
which remained constant at 60.4% (at 0.001 µg-ml-1 recombinant
PPI), 54.7% (at 0.01 µg-ml-1 recombinant PPI), 55.5% (at 0.1
µg-ml-1 recombinant PPI) and 55.4% (at 1.0 µg-ml-1 recombinant
PPI). These results demonstrate that recombinant PPI treatment did
not prevent apoptosis from H2O2-induced
oxidative stress However, the recombinant PPI was not toxic to
HepG2 cells. Therefore, recombinant PPI was then tested further for
its antioxidant.
Inhibition of ROS production
DCF-DA staining was used to examine whether
recombinant PPI exposure inhibited ROS production in HepG2 cells
co-treated with H2O2. HepG2 cells were
challenged with H2O2, and the resulting ROS
levels were 29.7% greater compared with the unchallenged control.
Pretreatment of cells with the various concentrations of
recombinant PPI (0.001–1 µg-ml-1) reduced the
H2O2-mediated increase in ROS formation
(Fig. 2B). Recombinant PPI
treatment showed antioxidant effects in the presence of
H2O2, implying that PPI directly scavenges
ROS or other free radicals.
Induction of antioxidant enzyme
activities in HepG2 cells
To investigate whether the antioxidant properties of
recombinant PPI are related to antioxidant enzyme induction, HepG2
cells were treated with recombinant PPI and the activity of the
antioxidant enzymes CAT, GPx, SOD and TRR were measured. As shown
in Fig. 3A, CAT activity was
dramatically downregulated by H2O2 treatment,
whereas the addition of recombinant PPI (0.001–1 µg-ml-1) was able
to restore CAT activity. GPx activity was reduced by 14% when cells
were exposed to H2O2, but levels recovered in
the presence of recombinant PPI at 1 µg-ml-1 (Fig. 3B). The activity of SOD was not
altered by H2O2 however, increased with the
treatment of recombinant PPI (Fig.
3C). PPI treatment at concentrations between 0.01 and 1 µg-ml-1
was able to increase the H2O2-induced
decrease in TRR activity (Fig.
3D). Recombinant PPI treatments increased the activities of
CAT, GPx and TRR that were reduced by exposure to
H2O2. The results also indicated that
recombinant PPI was also able to significantly increase SOD
activity. Therefore, recombinant PPI appears to exert its
antioxidant effects by modulating the activities of endogenous
antioxidant enzymes.
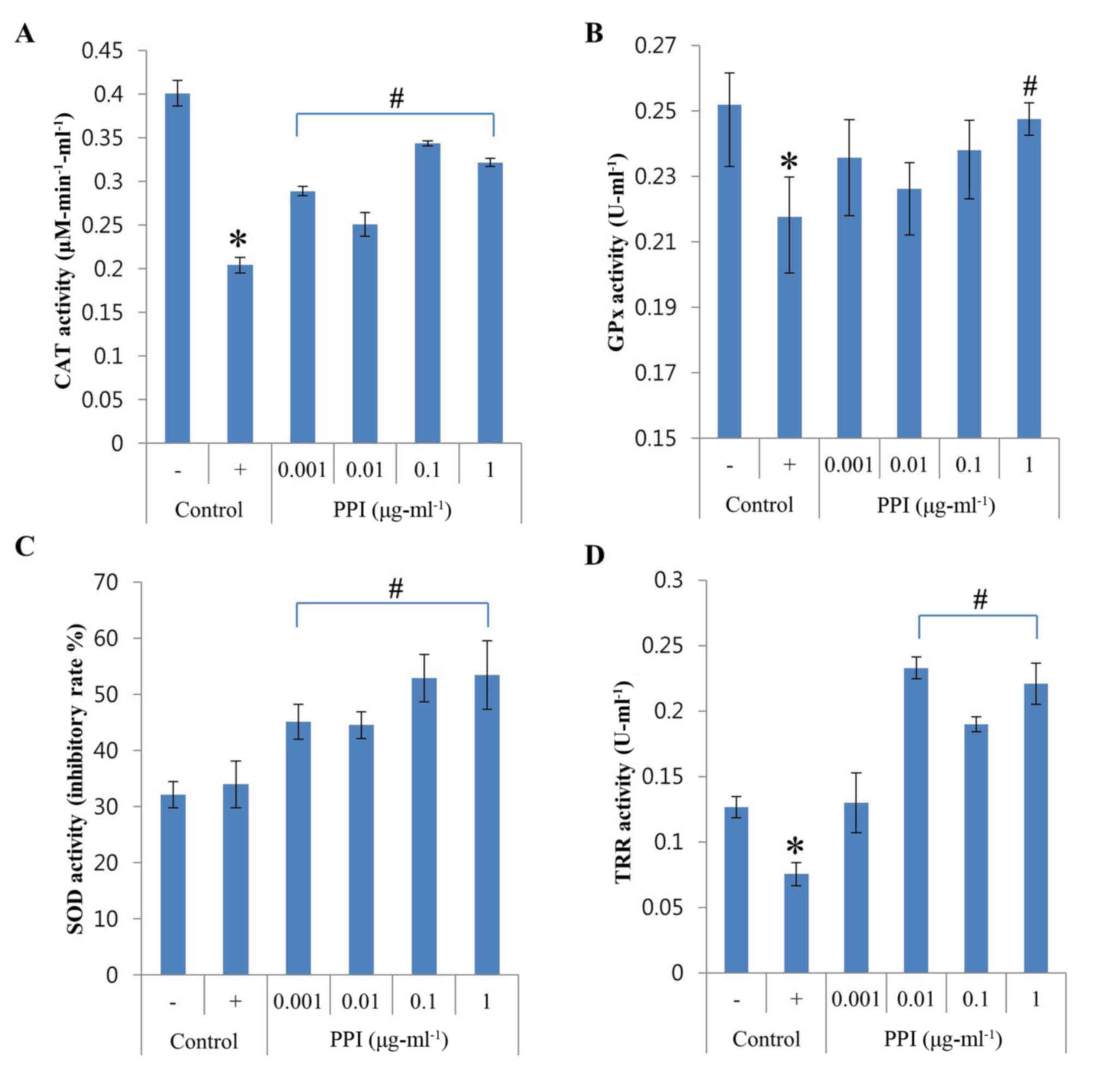 | Figure 3.Effects of PPI on SOD, GPx, CAT and
TRR activities in H2O2-treated HepG2 cells.
(A) CAT activity; (B) GPx activity; (C) SOD activity; and (D) TRR
activity. Values represent the mean ± standard deviation (n=3).
*P<0.05 vs. untreated control; #P<0.05 vs.
H2O2-treated control. -, control cells
without H2O2; +, control cells treated with 1
mM H2O2; CAT, catalase; GPx, glutathione
peroxidase; H2O2, hydrogen peroxide; PPI,
peptidylprolyl cis-trans isomerase; ROS, reactive oxygen
species; SOD, superoxide dismutase; TRR, thioredoxin reductase. |
Induction of antioxidant enzyme
expression in HepG2 cells
In addition to its effects on antioxidant enzyme
activities, the effects of recombinant PPI on mRNA expression were
evaluated in HepG2 cells. In cells treated with
H2O2, CAT mRNA expression levels of were
significantly diminished compared with the
H2O2-treated -treated controls and GAPDH mRNA
expression (Fig. 4A and B).
H2O2 exposure significantly decreased CAT
mRNA expression level, which was then reversed five-fold by
addition of 0.01 µg-ml-1 recombinant PPI. The level of GPx mRNA
expression was highest (ratio 0.52) when cells were pretreated with
recombinant PPI at a concentration of 0.1 µg-ml-1 (Fig. 4A,C). The results demonstrated that
treatment with H2O2 led to decreased CAT and
GPx mRNA expression; however, SOD and TRR mRNA expression levels
were not affected by H2O2 treatment (Fig. 4A,D and E). SOD mRNA expression was
significantly increased when treated with 0.01 and 0.1 µg-ml-1
recombinant PPI, as compared with the positive control (Fig 4A and D). TRR mRNA levels were
highest (ratio 0.87) at a concentration of 0.001 µg-ml-1
recombinant PPI (Fig. 4A and E).
Recombinant PPI treatment in HepG2 cells increased the level of
mRNA expression of reduced antioxidant enzymes when cells were
treated with H2O2. Therefore, PPI increased
the expression of the majority of antioxidant enzymes, however this
differed depending on the concentration of PPI.
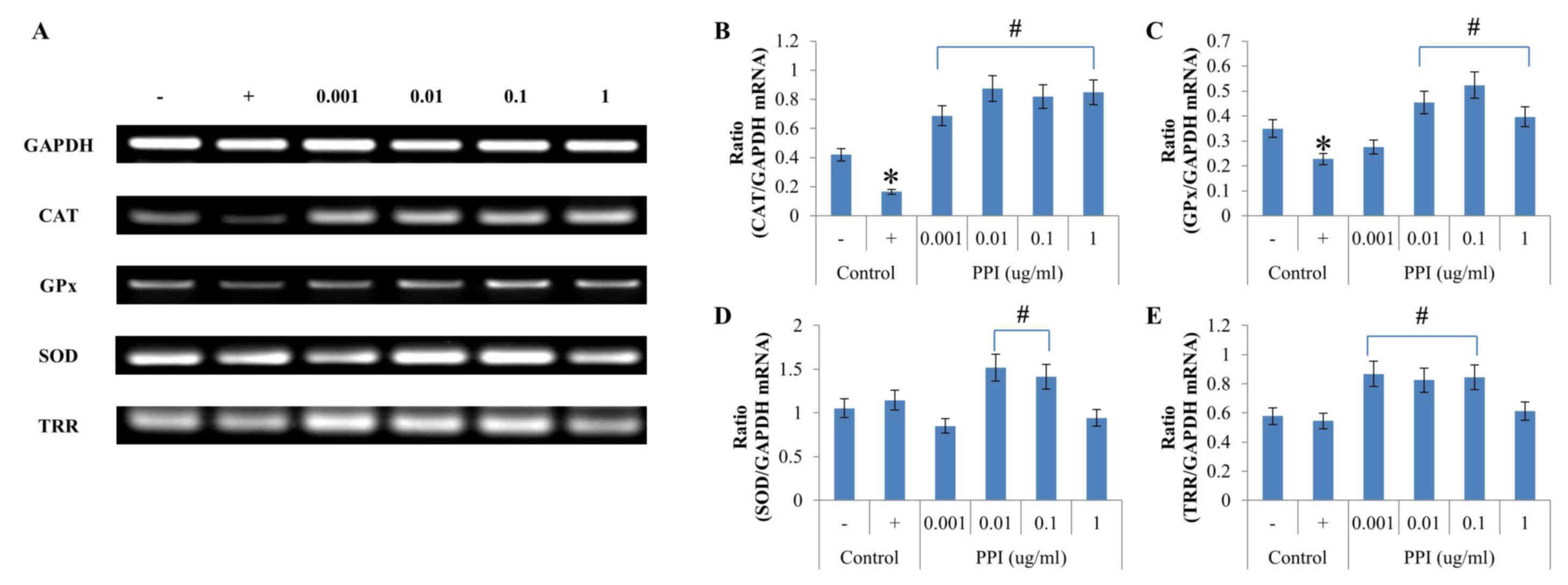 | Figure 4.Effects of PPI on antioxidant enzymes
mRNA expressions in H2O2 treated HepG2 cells.
(A) Reverse transcription-polymerase chain reaction analysis of
SOD, GPx, CAT and TRR. (B-E) Data quantification of antioxidant
enzyme/GAPDH mRNA expression for (B) CAT, (C) GPx, (D) SOD, and (E)
TRR. Values represent the mean ± standard deviation (n=3).
*P<0.05 vs. untreated control; #P<0.05 vs.
H2O2-treated control. -, control cells
without H2O2; +, control cells treated with 1
mM H2O2; CAT, catalase; GPx, glutathione
peroxidase; H2O2, hydrogen peroxide; PPI,
peptidylprolyl cis-trans isomerase; ROS, reactive oxygen
species; SOD, superoxide dismutase; TRR, thioredoxin reductase. |
Discussion
The findings of the present study supported the
hypothesis that recombinant PPI has multiple antioxidant properties
that enable it to protect cells. First, recombinant PPI treatment
made cells more resistant to H2O2-induced
oxidative stresses. Second, recombinant PPI treatment increased
antioxidant enzyme activities, providing cells with a higher
capacity for scavenging ROS. Accordingly, recombinant PPI increased
the H2O2-induced decreases in antioxidant
enzyme activity and their mRNA expression. These data suggest that
recombinant PPI cloned from the red algae P. yezoensis may
be able protect cells as an inducer of antioxidant enzymes. PPI
treatment increased the activities of antioxidant enzymes, such as
CAT, GPx, SOD and TRR.
The mechanism of cell repair against oxidative
stress is directly or indirectly regulated by antioxidants.
Direct-regulation of antioxidants is possible at low
concentrations, however the period of activation time is short.
Conversely, the indirect-regulation of antioxidants may occur via
the Kelch ECH associating protein 1/nuclear factor erythroid
2-related factor 2/antioxidant response element (Keap1/Nrf2/ARE)
pathway, which is relatively long-lasting (35). When oxidative stress was induced by
1 mM H2O2, the activities of CAT and GPx,
which primarily act to remove H2O2, were
decreased. In addition, the activity of TRR, which helps PRDX
directly remove H2O2, decreased. This
suggested that the antioxidant enzymes in the cell disappeared
following activation to remove the increased
H2O2. However, the activity of SOD which is
not directly involved in the removal of H2O2
was not different compared with the negative control.
The treatment of 0.001 µg-ml−1
recombinant PPI directly increased the activities of CAT, GPx and
TRR to remove H2O2, thereby reducing the
intracellular ROS. In addition, as a result of primary antioxidant
activity, SOD activity was increased to remove newly generated ROS.
The recombinant PPI treatment of 0.001 µg-ml−1 increased
the mRNA expressions of CAT, GPx and TRR and did not increase the
mRNA expression of SOD. This suggests that the recombinant PPI
concentration of 0.001 µg-ml−1 does not affect the
expression of SOD, and that the increase in SOD activity may be
self-regulated in cells to remove newly generated ROS. In addition,
an increase in TRR mRNA expression aided PRDX, which is directly
associated with H2O2.
When treated with 1 µg-ml−1 recombinant
PPI, the SOD activity was 53.4%, which was increased compared with
the positive control. However, expression of mRNA was decreased
compared with the positive control. These results suggested that
the SOD antioxidant activity was not regulated by its associated
signaling pathway, however by the antioxidant activity of the
recombinant PPI. It has been reported that the recombinant PPI of
the sweet potato roots gene has its own antioxidant activity
(36). The recombinant PPI in the
present study exhibited 20% antioxidant activity in vitro at
1 µg-ml−1 (data not shown). Therefore, the results of
SOD activity and mRNA expression are different due to the fact that
the antioxidant activity of SOD is regulated by that of PPI.
The recombinant PPI treatment of 0.01–0.1
µg-ml−1 increased the activities of CAT, GPx, SOD and
TRR, and mRNA expression was high. It is predicted that recombinant
PPI regulates the Keap1/Nrf2/ARE signal pathway for antioxidant
enzyme expression. Therefore, the regulation of antioxidation by
recombinant PPI directly promoted the activity of antioxidant
enzymes, however was also indirectly controlled. Recently, Lee
et al (37) reported that
Cyp including PPI, binds to TRR and enhances its antioxidant
activity. When the recombinant PPI is treated at a concentration of
0.01 or more, the activity of TRR may be rapidly increased due to
the combination of PPI and TRR.
However, the expression of mRNA does not entirely
determine the activity of the enzyme (38). Therefore, the present study
primarily confirmed the activity and mRNA expression alterations of
antioxidant enzymes that regulate induced oxidative stress. Gene
regulation and enzyme activity combinations were additionally
predicted. In the majority of cases, the expression of mRNA was
directly associated with the regulation of the antioxidant enzyme
activity. However, when treated with 0.001 µg/ml of PPI, the
expression of TRR mRNA rapidly increased, whereas the activity of
TRR was decreased compared with other experimental groups. Further
experimentation is therefore required on TRR, PRDX and their
associated regulatory mechanisms.
The present study revealed for the first time, to
the best of our knowledge, the antioxidant properties of P.
yezoensis recombinant PPI. The results of the present study
indicated that recombinant PPI treatment controlled the expression
of antioxidant enzymes and maintained the antioxidant capacity of
the cell. Additional studies into the interaction between
recombinant PPI and antioxidant enzymes are necessary to elucidate
the antioxidant mechanisms including the Keap1/Nrf2/ARE signal
pathway of recombinant PPI in the cell.
The present study also reported for the first time,
and to the best of our knowledge, the isolation of PPI mRNA from
P. yezoensis. The biochemical and physiological effects of
recombinant PPI protein during oxidative stress requires further
study. However, results from the present study suggest that
recombinant PPI is an excellent candidate for the development of
therapeutically useful antioxidant agents.
Acknowledgements
The present study was supported by The Basic Science
Research Program through the National Research Foundation of Korea,
funded by the Ministry of Education (grant no.
2012R1A6A1028677).
References
|
1
|
Harman D: Role of free radicals in aging
and disease. Ann N Y Acad Sci. 673:126–141. 1992. View Article : Google Scholar
|
|
2
|
Bhattacharyya A, Chattopadhyay R, Mitra S
and Crowe SE: Oxidative stress: An essential factor in the
pathogenesis of gastrointestinal mucosal diseases. Physiol Rev.
94:329–354. 2014. View Article : Google Scholar :
|
|
3
|
Bentley AR, Emrani P and Cassano PA:
Genetic variation and gene expression in antioxidant related
enzymes and risk of COPD: A systematic review. Thorax. 63:956–961.
2008. View Article : Google Scholar :
|
|
4
|
Lobo V, Patil A, Phatak A and Chandra N:
Free radicals, antioxidants and functional foods: Impact on human
health. Pharmacogn Rev. 4:118–126. 2010. View Article : Google Scholar :
|
|
5
|
Young IS and Woodside JV: Antioxidants in
health and disease. J Clin Pathol. 54:176–186. 2001. View Article : Google Scholar :
|
|
6
|
Apel K and Hirt H: Reactive oxygen
species: Metabolism, oxidative stress, and signal transduction.
Annu Rev Plant Biol. 55:373–399. 2004. View Article : Google Scholar
|
|
7
|
Zelko IN, Mariani TJ and Folz RJ:
Superoxide dismutase multigene family: A comparison of the CuZn-SOD
(SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures,
evolution, and expression. Free Radic Biol Med. 33:337–349. 2002.
View Article : Google Scholar
|
|
8
|
Flora SJ: Structural, chemical and
biological aspects of antioxidants for strategies against metal and
metalloid exposure. Oxid Med Cell Longev. 2:191–206. 2009.
View Article : Google Scholar :
|
|
9
|
de M, Bandeira S, da Fonseca LJ, da S
Guedes G, Rabelo LA, Goulart MO and Vasconcelos SM: Oxidative
stress as an underlying contributor in the development of chronic
complications in diabetes mellitus. Int J Mol Sci. 14:3265–3284.
2013. View Article : Google Scholar :
|
|
10
|
Takahashi N, Hayano T and Suzuki M:
Peptidyl-prolyl cis-trans isomerase is the cyclosporin A-binding
protein cyclophilin. Nature. 337:473–475. 1989. View Article : Google Scholar
|
|
11
|
Pérez de Lema G, Arribas-Gómez I,
Ruiz-Ginés JA, de Arriba G, Prieto A, Rodriguez-Puyol D and
Rodriguez-Puyol M: Reactive oxygen species mediate the effects of
cyclosporine A on human cultured mesangial cells. Transplant Proc.
29:1241–1243. 1997. View Article : Google Scholar
|
|
12
|
McGrath LT, Treacy R, McClean E and Brown
JH: Oxidative stress in cyclosporin and azathioprine treated renal
transplant patients. Clin Chim Acta. 264:1–12. 1997. View Article : Google Scholar
|
|
13
|
Zhong Z, Arteel GE, Connor HD, Yin M,
Frankenberg MV, Stachlewitz RF, Raleigh JA, Mason RP and Thurman
RG: Cyclosporin A increases hypoxia and free radical production in
rat kidneys: Prevention by dietary glycine. Am J Physiol.
275:F595–F604. 1998.
|
|
14
|
Brazin KN, Mallis RJ, Fulton DB and
Andreotti AH: Regulation of the tyrosine kinase Itk by the
peptidyl-prolyl isomerase cyclophilin A. Proc Natl Acad Sci USA.
99:pp. 1899–1904. 2002; View Article : Google Scholar :
|
|
15
|
Ferreira PA, Nakayama TA, Pak WL and
Travis GH: Cyclophilin-related protein RanBP2 acts as chaperone for
red/green opsin. Nature. 383:637–640. 1996. View Article : Google Scholar
|
|
16
|
Galat A: Variations of sequences and amino
acid compositions of proteins that sustain their biological
functions: An analysis of the cyclophilin family of proteins. Arch
Biochem Biophys. 371:149–162. 1999. View Article : Google Scholar
|
|
17
|
Horowitz DS, Lee EJ, Mabon SA and Misteli
T: A cyclophilin functions in pre-mRNA splicing. EMBO J.
21:470–480. 2002. View Article : Google Scholar :
|
|
18
|
Krzywicka A, Beisson J, Keller AM, Cohen
J, Jerka-Dziadosz M and Klotz C: KIN241: A gene involved in cell
morphogenesis in Paramecium tetraurelia reveals a novel protein
family of cyclophilin-RNA interacting proteins (CRIPs) conserved
from fission yeast to man. Mol Microbiol. 42:257–267. 2001.
View Article : Google Scholar
|
|
19
|
Leverson JD and Ness SA: Point mutations
in v-Myb disrupt a cyclophilin-catalyzed negative regulatory
mechanism. Mol Cell. 1:203–211. 1998. View Article : Google Scholar
|
|
20
|
Lin DT and Lechleiter JD: Mitochondrial
targeted cyclophilin D protects cells from cell death by peptidyl
prolyl isomerization. J Biol Chem. 277:31134–31141. 2002.
View Article : Google Scholar
|
|
21
|
O'Connell S, Tuite N, Slattery C, Ryan MP
and McMorrow T: Cyclosporine A-induced oxidative stress in human
renal mesangial cells: A role for ERK 1/2 MAPK signaling. Toxicol
Sci. 126:101–113. 2012. View Article : Google Scholar
|
|
22
|
de Oliveira e Silva AM, Vidal-Novoa A,
Batista-González AE, Pinto JR, Mancini DA Portari, Reina-Urquijo W
and Mancini-Filho J: In vivo and in vitro antioxidant activity and
hepatoprotective properties of polyphenols from Halimeda opuntia
(Linnaeus) Lamouroux. Redox Rep. 17:47–53. 2012. View Article : Google Scholar
|
|
23
|
Ismail A Jr and Tan S: Antioxidant
activity of selected commercial seaweeds. Malays J Nutr. 8:167–177.
2002.
|
|
24
|
Chandini SK, Ganesan P and Bhaskar N: In
vitro antioxidant activities of three selected brown seaweeds of
India. Food Chem. 107:707–713. 2008. View Article : Google Scholar
|
|
25
|
Kumar SR, Hosokawa M and Miyashita K:
Fucoxanthin: A marine carotenoid exerting anti-cancer effects by
affecting multiple mechanisms. Mar Drugs. 11:5130–5147. 2013.
View Article : Google Scholar :
|
|
26
|
Sathya R, Kanaga N, Sankar P and Jeeva S:
Antioxidant properties of phlorotannins from brown seaweed
Cystoseira trinodis (Forsskål) C. Agardh. Arab J Chem. 2013.
View Article : Google Scholar
|
|
27
|
Yuan YV, Bone DE and Carrington MF:
Antioxidant activity of dulse (Palmaria palmata) extract evaluated
in vitro. Food Chem. 91:485–494. 2005. View Article : Google Scholar
|
|
28
|
Balboa EM, Conde E, Moure A, Falqué E and
Domínguez H: In vitro antioxidant properties of crude extracts and
compounds from brown algae. Food Chem. 138:1764–1785. 2013.
View Article : Google Scholar
|
|
29
|
Ganesan P, Kumar CS and Bhaskar N:
Antioxidant properties of methanol extract and its solvent
fractions obtained from selected Indian red seaweeds. Bioresour
Technol. 99:2717–2723. 2008. View Article : Google Scholar
|
|
30
|
Hwang ES and Thi ND: Effects of extraction
and processing methods on antioxidant compound contents and radical
scavenging activities of laver (Porphyra tenera). Prev Nutr Food
Sci. 19:40–48. 2014. View Article : Google Scholar :
|
|
31
|
Fathi-Roudsari M, Akhavian-Tehrani A and
Maghsoudi N: Comparison of three Escherichia coli strains in
recombinant production of reteplase. Avicenna J Med Biotechnol.
8:16–22. 2016.
|
|
32
|
Abu-Reidah IM, Arráez-Román D,
Quirantes-Piné R, Fernández-Arroyo S, Segura-Carretero A and
Fernández-Gutiérrez A: HPLC-ESI-Q-TOF-MS for a comprehensive
characterization of bioactive phenolic compounds in cucumber whole
fruit extract. Food Res Int. 46:108–117. 2012. View Article : Google Scholar
|
|
33
|
Schmidt AJ, Heiser P, Hemmeter UM, Krieg
JC and Vedder H: Effect of antidepressants on mRNA levels of
antioxidant enzymes in human monocytic U-937 cells. Prog
Neuro-Psychoph. 32:1567–1573. 2008. View Article : Google Scholar
|
|
34
|
Aguilar-Melero P, Prieto-Álamo MJ, Jurado
J, Holmgren A and Pueyo C: Proteomics in HepG2 hepatocarcinoma
cells with stably silenced expression of PRDX1. J Proteomics.
79:161–171. 2013. View Article : Google Scholar
|
|
35
|
Dinkova-Kostova AT and Talalay P: Direct
and indirect antioxidant properties of inducers of cytoprotective
proteins. Mol Nutr Food Res. 52 Suppl 1:S128–S138. 2008.
|
|
36
|
Liao JC, Chiu CS, Chen HJ, Huang SS, Hou
WC, Lin WC, Lin YH and Huang GJ: Characterization of a novel
Cyclophilin-type peptidylprolyl isomerase protein from sweet potato
storage roots. Botanical Studies. 53:315–324. 2012.
|
|
37
|
Lee SP, Hwang YS, Kim YJ, Kwon KS, Kim HJ,
Kim K and Chae HZ: Cyclophilin A binds to peroxiredoxins and
activates its peroxidase activity. J Biol Chem. 276:29826–29832.
2001. View Article : Google Scholar
|
|
38
|
Glanemann C, Loos A, Gorret N, Willis LB,
O'Brien XM, Lessard PA and Sinskey AJ: Disparity between changes in
mRNA abundance and enzyme activity in Corynebacterium glutamicum:
Implications for DNA microarray analysis. Appl Microbiol
Biotechnol. 61:61–68. 2003. View Article : Google Scholar
|