Introduction
Diabetes mellitus (DM) is one of the most severe
metabolic diseases, which affects >170 million people worldwide
(1). A major symptom of DM is an
increased blood sugar level, which results in a number of
additional complications, such as skin ulcers, which affect ~15% of
DM patients (1). In addition, DM
affects angiogenesis, which leads to the delay of wound healing
(2); however, the underlying
mechanisms remain unclear.
Wound healing is a complex process that requires the
cooperation of keratinocytes, fibroblasts, endothelial cells,
macrophages and platelets (3).
Fibroblast cell proliferation and migration is known to serve an
important role in the wound repair process, as well as in collagen
deposition and remodeling, wound contraction, angiogenesis and the
formation of extracellular matrix (ECM) (3). It is possible that poor DM-associated
wound healing may be due to altered protein and lipid metabolism
and the abnormal formation of granulation tissue (4). Increased blood sugar levels alter
glycosylation enzymes and induce covalent binding of aldose sugars
to a protein or lipid. Binding of aldose sugar to a protein or
lipid produces a stable product, termed advanced glycation
end-product, which accumulate on the surface of the cell membrane
and ECM proteins leading to delayed turnover rates (4). Reactive oxygen species (ROS)
accumulate in high quantities in the skin of diabetic rats
(5). In addition, nitric oxide
(NO) is a known signaling molecule that stimulates fibroblast cell
proliferation, maturation, differentiation and collagen production
during the wound healing process (6). In fibroblasts from diabetic ulcers,
the cells often possess a dilated endoplasmic reticulum, an
increased number of vesicular bodies and defected micro tubular
structures (7). These subcellular
alterations may be induced by activated protein synthesis and
turnover, as well as defects in microtubule structure thus
inhibiting protein secretion via autonomous trafficking through
vesicles and induction of microtubules to release their products
into the extracellular space in diabetic ulcer fibroblasts
(7,8). Diabetic ulcers exhibit defective
fibroblast proliferation, which may be due to reduced ECM protein
production, and delayed wound contraction and healing (8). However, a limited number of
biochemical studies have been performed to date, that have
investigated the mechanisms underlying DM-mediated delay of wound
healing (5–8).
Fibroblast cell migration is an important step
during the wound closure process, and the phosphoinositide 3-kinase
(PI3K)-Ras-related C3 botulinum toxin substrate 1-c-Jun N-terminal
kinase (JNK) signaling pathway is important in the promotion of
cell migration (9). In a recent
study, RNA-Seq analysis was performed using high-glucose stimulated
human foreskin fibroblasts, in order to examine the response of a
large population of transcripts to high glucose-induced stress
(9). The results demonstrated that
the inflammatory response and Wnt signaling pathways generated
negative and positive responses to high glucose-induced stress,
respectively (10). Xuan et
al (10) revealed that nuclear
factor-κB functions independently of PI3K signaling to activate
JNK, resulting in accelerated fibroblast cell migration.
The present study investigated DM-induced
alterations in a large number of proteins in diabetic rat skin
using two-dimensional (2-D) gel electrophoresis. Proteomic analysis
and liquid chromatography/mass spectrometry (LC/MS) were performed
to identify the differentially expressed proteins. In addition, the
effect of annexin A2 on human foreskin fibroblast cell migration
was analyzed, as well as the potential association between annexin
A2 and JNK, a key regulator of cell migration. The present study
provides useful proteomic information that may enhance what is
currently known regarding the mechanisms underlying skin wound
closure.
Materials and methods
Generation of skin wounds in diabetic
rats
A rat model of type 1 diabetes was generated using a
previously described method (11).
A total of 12 male Sprague-Dawley rats (age, 8 weeks; weight, 150
to 200 g) were housed in cages (12 h light/12 h dark cycle; free
access to water and food) at 25°C and 30% humidity, divided into 2
groups (n=6/group) and were injected intraperitoneally with
streptozotocin (STZ; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany; 50 mg/kg) which was dissolved in sodium citrate buffer (pH
4.5) (12). STZ-injected rats with
blood glucose levels >16.7 mmol/l were considered to have
developed DM. To generate the control group, the same volume of
sodium citrate without STZ was injected into the male rats. To
generate skin wounds, the rats were anesthetized with pentobarbital
(Sigma-Aldrich; Merck KGaA; 45 mg/ml), and two circular wounds
(~200 mm2) that were 1 cm deep were created on the lower
back of each rat. Ethical approval was obtained from the ethics
committee of Wenzhou Medical University (Wenzhou, China).
Analysis of wound healing
Following 8 weeks of DM onset, the wound area was
evaluated every 4 days for 16 days. The skin wounds from 3 diabetic
and 3 control rats were photographed with rulers, and the area of
the unhealed regions was calculated using these images together
with TINA 2.0 software (DesignSoft, Inc., Budapest, Hungary;
http://www.designsoftware.com/)
(11).
Protein preparation and 2-D SDS-PAGE
analysis
Following anesthesia with pentobarbital (45 mg/ml;
Sigma-Aldrich; Merck KGaA), the dorsal area of diabetic rats was
totally depilated with Na2S (8.0%, w/v; Sigma-Aldrich; Merck KGaA)
and two full-thickness circular wounds (~300 mm2 each)
were created on the lower back of each rat using a pair of sharp
scissors and a scalpel. Skin tissues (2 g) that were 1-cm deep were
collected from the diabetic and control rats for protein extraction
2 weeks following the establishment of DM. The following reagents
were used for protein extraction: PBS, 9.5 M urea, 0.1% (w/v) DTT,
2% (w/v) CHAPS and 0.8% (w/v) pharmalyte (Sigma-Aldrich; Merck
KGaA). The extracts were dissolved in immobilized pH gradient (IPG)
rehydration buffer [8 M urea, 2% (w/v) 3-[(3-cholamidopropyl)
dimethylammonio]-1-propanesulfonate hydrate, 2% (w/v) IPG buffer,
0.04 M dithiothreitol, 1X nuclease solution and 0.1% (w/v) of
bromophenol blue] prior to isoelectric focusing electrophoresis.
The following reagents were used for 2-D gel analysis: 1% DTT, 50
mM Tris-HCl, pH 8.8, 6 M urea, 30% (v/v) glycerol, 2% (w/v) SDS,
0.1% (w/v) bromophenol blue, equilibration buffer (2.5% (w/v)
iodoacetamide (Sigma-Aldrich; Merck KGaA). The MS/MS spectra were
processed using Proteome Discoverer software (version 1.3; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) and the database search
was performed using the Mascot search engine (Matrix Science Mascot
version 2.3; http://www.matrixscience.com/) against a concatenated
forward-decoy approach (13).
In-gel digestion
Protein spots were visualized using ImageMaster 2D
Platinum version 6.0 (GE Healthcare Bio-Sciences, Pittsburgh, PA,
USA). A total of 77 protein spots of interested were excised and
digested using a 1% protease cocktail blue (Sigma-Aldrich; Merck
KGaA), which were further analysed using MS. In-gel digestion was
performed in 3 steps according to the procedure described by
Russell et al (14). The
following reagents were used for analysis: 50% (v/v) acetonitrile
(ACN), 10 mM DTT containing 100 mM ammonium bicarbonate and 1%
trypsin solution (Sigma-Aldrich; Merck KGaA).
LC-MS/MS analysis and bioinformatics
analysis
The proteins extracted from the gel sections were
digested using 1% trypsin solution (Sigma-Aldrich; Merck KGaA) and
4 µl of samples were submitted to online nanoflow liquid
chromatography using the easy-nano LC system (Proxeon Biosystems;
Thermo Fisher Scientific, Inc.) with 10 cm capillary columns of an
internal diameter of 75-µm, filled with 3-µm Reprosil-Pur C18-A2
resin (Dr. Maisch GmbH, Ammerbuch-Entringen, Germany) (14). The gradient consisted of 10–30%
(v/v) ACN in 0.1% (v/v) formic acid at a flow rate of 200 nl/min
for 45 min, 30–100% (v/v) ACN in 0.1% (v/v) formic acid at a flow
rate of 200 nl/min for 1 min and 100% CAN in 0.1% formic acid at a
flow rate of 200 nl/min for 10 min. The elution was electrosprayed
through a Proxeon nanoelectrospray ion source by (electrospray
ionization) ESI-MS/MS analysis on a Thermo Fisher LTQ Velos Pro
(Thermo Fisher Scientific, Inc.) using full ion scan mode over the
m/z range 200–1800. Collision-induced dissociation (CID) was
performed in the linear ion trap using a 4.0-Th isolation width and
35% normalized collision energy with helium as the collision gas.
Five independent MS/MS scans were performed on each ion using
dynamic exclusion. The precursor ion that was selected for CID was
dynamically excluded from further MS/MS analysis for 30 sec.
Further corresponding protein identification was performed using
the Swiss-Prot protein sequence database (version 54.5; http://web.expasy.org/docs/swiss-prot_guideline.html).
LC-MS/MS and bioinformatics analyses were performed by LC Bio, Inc.
(Hangzhou, China).
Western blot analysis
Skin tissues (100 mg) were ground using liquid
nitrogen and an ice-cold lysis solution (7 M urea, 2 M thiourea, 2%
CHAPS, 40 mM Tris base, 40 mM dithiothreitol, and 1% protease
inhibitor; Sigma-Aldrich; Merck KGaA) was added to procure whole
cell extracts. Total proteins (20 µg) from skin samples with or
without induced diabetes 2 weeks prior, were separated on a 12%
SDS-PAGE gel and electrotransferred to Immobilon®-P PVDF
Transfer Membranes (EMD Millipore, Billerica, MA, USA). The
membranes were blocked in 1X Tris-buffered saline containing 5%
skim milk and 0.05% Tween-20 for 2 h. All of the reagents for SDS
gel electrophoresis and membrane blocking were purchased the
Sigma-Aldrich; Merck KGaA. To analyse protein levels in the samples
the membranes were incubated with the following primary antibodies
at room temperature (25°C) for 2 h: Anti-eNOS antibody (cat. no.
ab76198; 1:2,000, Abcam, Cambridge, MA, USA), anti-cleaved
(c)-caspase-3 antibody (cat. no. ab2302; 1:2,000, Abcam),
anti-plasminogen activator inhibitor (PAI) −1 antibody (cat. no.
ab125687; 1:2,000; Abcam), anti-cluster of differentiation (CD) 34
antibody (cat. no. ab81289; 1:2,000; Abcam), anti-collagen1
antibody (cat. no. ab34710; 1:2,000; Abcam), anti-annexin A2
antibody (cat. no. ab41803; 1:1,000; Abcam), anti-JNK antibody
(cat. no. ab179461; 1:2,000; Abcam), anti-phosphorylated (p) -JNK
antibody (cat. no. ab4821; 1:2,000; Abcam) and anti-GAPDH antibody
(cat. no. ab8245; 1:2,000; Abcam). The membranes were then
incubated with either the anti-mouse or anti-rabbit horseradish
peroxidase-conjugated secondary antibody (cat. no. 7074; 1:2,000;
Cell Signaling Technology, Inc., Danvers, MA, USA) at room
temperature (25°C) for 2 h. Antigen-antibody complexes were
visualized using an electrochemiluminescence kit (BioTrand, Crystal
Lake, Illinois, USA). Protein levels were normalized against GAPDH
and protein expression was analyzed using Image J2 version 2.0
software (13).
Human foreskin fibroblast cell
culture
Human fibroblast cell culture was performed as
previously described (5). Human
foreskin tissues were collected from 3 patients diagnosed with
redundant prepuce (all three patients were healthy without DM or
other disease diagnoses; ages, 26, 30 and 32 years old) who were
admitted in January 2014 to the Department of Dermatology at the
First Affiliated Hospital of Wenzhou Medical University (Wenzhou,
China). The present study was approved by the ethics committee of
Wenzhou Medical University, and written informed consent was
obtained from all patients involved. The fat was first removed from
the tissue samples, when were subsequently cut into 3×2-mm strips
and incubated overnight at 4°C in 0.05% Dispase I (Sigma-Aldrich;
Merck KGaA). The epidermis was then removed and the dermis was
placed into 25 cm2 flasks pre-treated with foetal bovine
serum (FBS; HyClone; GE Healthcare Life Sciences, Logan, UT, USA).
The flasks were positioned horizontally for 1 h, and then
vertically for 3 h in a culture chamber at 37°C with 5% CO2. The
cells were cultured in Dulbecco's modified Eagle's medium (DMEM;
HyClone; GE Healthcare Life Sciences) containing 5.5 mM glucose
(Sigma-Aldrich; Merck KGaA), 10% FBS and 1% penicillin-streptomycin
(Gibco; Thermo Fisher Scientific, Inc.); the medium was refreshed
every 3 days. When cell confluence had reached 70 to 80% the cells
were digested and passaged using 0.25% trypsin (Gibco; Thermo
Fisher Scientific, Inc.). Cells were cultured for 3 days in 5.5 mM
glucose medium, before they were transferred to media with or
without 25 µM SP600125 (Cell Signaling Technology, Inc.) for up to
1 day prior to the migration assay, or for 1 h prior to western
blot analysis.
Overexpression of annexin A2 in
fibroblast cells
For overexpression of annexin A2, the open reading
frame (ORF) sequences of the annexin A2 gene (ANXA2; GenBank
Accession no. NM_001002858.2; National Center for Biotechnology
Information, https://www.ncbi.nlm.nih.gov/) was amplified for 28
cycles of 95°C for 30 sec, 60°C for 30 sec and 72°C for 90 sec
using the following primers: Forward,
5′-ATCGTAGGATCCATGGGCCGCCAGCTAG-3′, and reverse,
5′-AGCTATTCTAGATCAGTCATCTCCACCACACAGGTA-3′. The amplified
polymerase chain reaction (PCR) fragments were then cloned into the
Bacillus amyloliquefaciens and Xanthomonas badrii
restriction enzyme sites of the pcDNA3 expression vector
(Invitrogen; Thermo Fisher Scientific, Inc.). Primer design and
sequencing were performed by Sangon Biotech Co., Ltd. (Shanghai,
China; http://www.sangon.com/). A total of 2 or
3 µg pcDNA3-ANXA2 plasmid was transformed into human
foreskin primary fibroblasts from the 3 patients using a
Lipofectamine 2000 kit (Invitrogen; Thermo Fisher Scientific, Inc.)
according the manufacturer's instructions. Empty vector pcDNA3
using Lipofectamine 2000 transformed cells was used as control. The
2 and 3 µg of pcDNA3-ANXA2 plasmid transformed cells were
termed OX1 and OX2, respectively. Following 2 days of
transformation, annexin A2 levels in the fibroblast cells were
examined by western blotting, and cell migration tests were
performed.
Cell migration assay
Primary fibroblast cells were seeded onto 6-well
plates at 80–90% confluency and incubated at 37°C overnight in DMEM
containing 0.5% FBS and 5 µg/ml mitomycin-C (Sigma-Aldrich; Merck
KGaA). Linear scratch wounds were subsequently created in the
confluent fibroblast monolayer using a sterile 200 µl pipette tip
(Ningbo MFLab Medical Instruments Co., Ltd., Ningbo, Zhejiang,
China). The medium was immediately replaced with prewarmed (37°C)
fresh DMEM containing 0.5 % FBS and 5 µg/ml mitomycin-C. At 0, 12
and 24 h following wound establishment, images were captured using
a light microscope (IX70; Olympus Corporation, Tokyo, Japan)
equipped with a CCD camera (CoolSNAP HQ; Nippon Roper, Toyko,
Japan), which was controlled by MetaMorph 7.1 software (Molecular
Devices, LLC, Sunnyvale, CA, USA). To quantify cell migration, 10
cells on the border of the wound area were randomly selected from
each well and the migration distance was measured using ImageJ
version 14.8 software (https://imagej.nih.gov/ij/) at the indicated time
points.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software (version 5.0; GraphPad Software, Inc., La Jolla, CA,
USA). Comparisons between 2 groups were performed using a Student's
t-test, while significant differences among 3 groups were analysed
using one-way analysis of variance, followed by Bonferroni's
multiple comparison test. P<0.05 was considered to indicate a
statistically significant difference.
Results
DM impairs skin wound healing
Diabetes-induced skin wound healing was analyzed in
a rat model of diabetes. STZ, an inducer of type 1 diabetes was
used to induce diabetes in rats, and wound healing was assessed by
monitoring and comparing the size of ~200 mm2 circular
wounds created on the waist of each rat at 8 weeks following the
onset of diabetes (8). Wound
repair was then analysed over the course of 16 days, and the
results indicated that the rate of wound healing was significantly
reduced in diabetic rats when compared with non-diabetic normal
rats (Fig. 1).
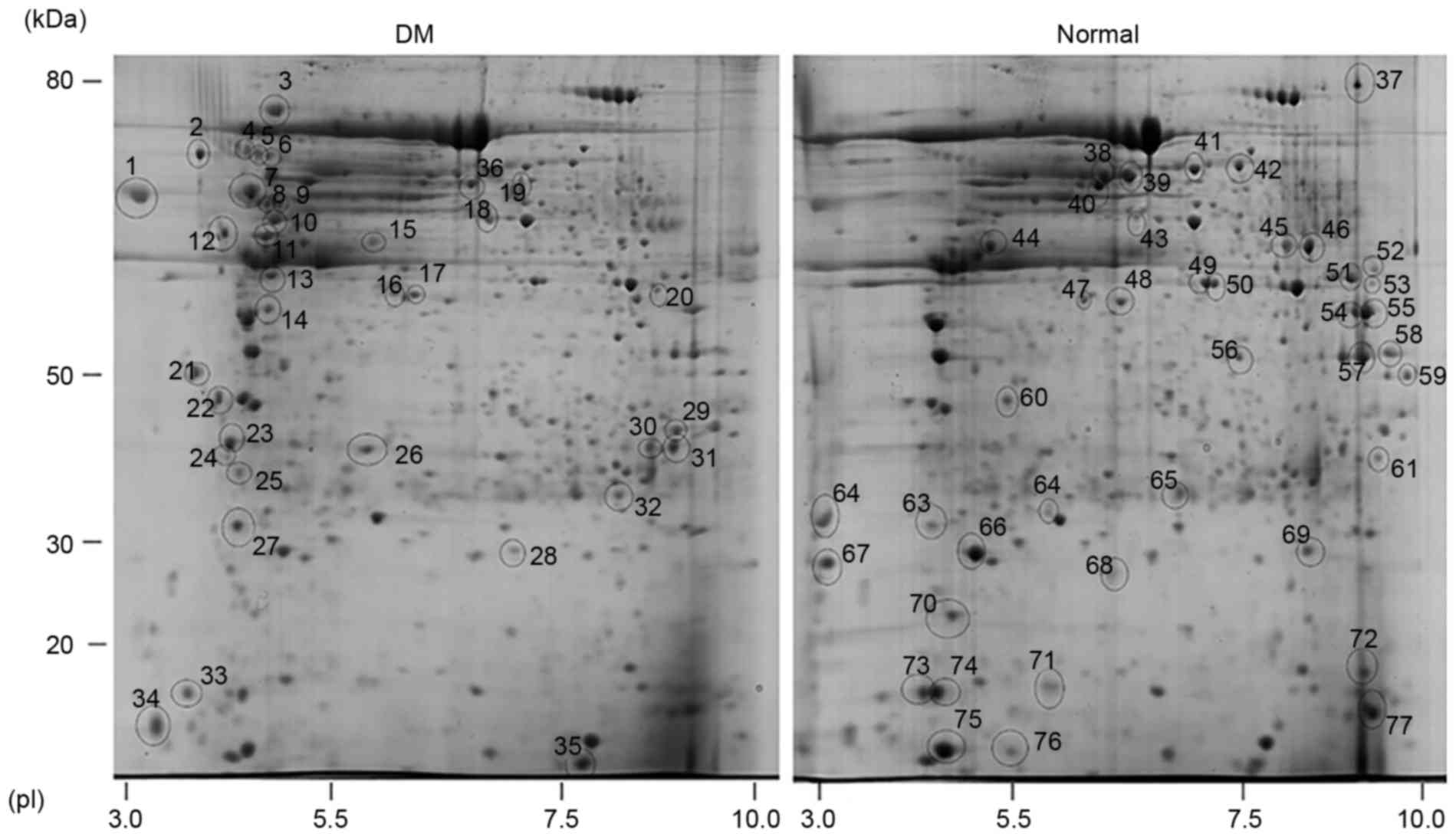
Proteomic analysis of rat skin tissues
under DM stress
In order to gain an improved understanding of the
molecular mechanisms underlying DM-mediated impairment of skin
wound healing, comparative proteomic assays were preformed to
identify differentially expressed proteins in the skin of DM rats
and normal rats using 2-D gel electrophoresis. Total skin protein
extracts were first subjected to 2-D gel analysis followed by
identification by LC/MS. This led to the detection of proteins
altered by DM. Proteins extracted from the normal rat skin were
used as a control (Fig. 2).
Optimization of the 2-D gels and image processing identified 36 and
41 protein spots (representing differentially expressed proteins)
that were induced and repressed by DM, respectively (Fig. 2). The experiments were repeated 3
times and only the reproducible alterations were taken into
account. To identify differentially expressed proteins, the spots
were excised from the 2-D gels and identified by LC/MS and trypsin
digestion. The results presented in Table I demonstrate that 72 proteins were
identified among the 77 spots observed in the 2-D gel (Fig. 2). The major group of proteins
identified, either overexpressed or repressed, were observed to be
involved in regulation of the cytoskeleton (tubulin and actin
formation), carbohydrate metabolism (glycolysis and the
tricarboxylic acid cycle) and protein folding. In the present
study, the calcium binding proteins, calmodulin and calreticulin,
were induced in DM (Table I).
Formation of the ECM is an important step in the wound healing
process (3). DM reduced the levels
of collagen α1 and dermatopontin, two ECM-associated proteins. In
addition to the major group of proteins, RNA metabolism, adenosine
triphosphate (ATP) synthesis and cell migration-associated protein
levels were altered (Table I).
 | Table I.Differentially expressed proteins
identified by liquid chromatography/mass spectrometry analysis. |
Table I.
Differentially expressed proteins
identified by liquid chromatography/mass spectrometry analysis.
| A, Proteins highly
expressed in DM skin |
|---|
|
|---|
| Spot no. | UniProt ID | Protein | Function |
|---|
| 1 | P69897 | Tubulin β-5
chain | Cytoskeleton |
| 8 | P85108 | Tubulin β-2A
chain | Cytoskeleton |
| 9 | Q4QRB4 | Tubulin β | Cytoskeleton |
| 13 | P48675 | Desmin | Cytoskeleton |
| 16 | O35763 | Moesin | Cytoskeleton |
| 17 | O35763 | Moesin | Cytoskeleton |
| 2 | P18418 | Calreticulin | Ca2+
binding |
| 4 | P18418 | Calreticulin | Ca2+
binding |
| 12 | P43742 | Reticulocalbin-3 | Ca2+
binding |
| 34 | P62161 | Calmodulin
(inflammation and apoptosis) | Ca2+
binding |
| 3 | P06761 | 78 kDa
glucose-regulated protein | Protein folding and
oligomerization |
| 7 | P04785 | Protein
disulfide-isomerase | Protein folding |
| 19 | P11598 | Protein
disulfide-isomerase A3 | Protein folding |
| 36 | P11598 | Protein
disulfide-isomerase A3 | Protein folding |
| 24 | P34064 | Proteasome subunit α
type-5 | Protein
degradation |
| 14 | G3V9R8 | Heterogeneous
nuclear ribonucleoprotein C1/C2 | RNA metabolism |
| 6 | P61980 | Heterogeneous
nuclear ribonucleoprotein K | RNA metabolism |
| 22 | B5DEN5 | Elongation factor
1-β | RNA metabolism |
| 33 | P02401 | 60S acidic
ribosomal protein P2 | RNA metabolism |
| 35 | P63324 | 40S ribosomal
protein S12 | RNA metabolism |
| 20 | P62630 | Elongation factor
1-α 1 | RNA metabolism |
| 5 | P33840 |
Antithrombin-III | Plasma protease
inhibitors |
| 10 | P10719 | ATP synthase
subunit β | ATP synthesis |
| 11 | P31000 | Vimentin | Signaling |
| 23 | P20332 | 14-3-3 protein
σ | Signaling |
| 25 | Q79342 | 14-3-3 protein
σ | Signaling |
| 15 | Q9WTT6 | Guanine
deaminase | Purine
metabolism |
| 18 | P04764 | α-enolase | Carbon
metabolism |
| 21 | P02650 | Apolipoprotein
E | Carbon metabolism
(inflammation) |
| 29 | P04797 |
Glyceraldehyde-3-phosphate
dehydrogenase | Carbon
metabolism |
| 30 | P04797 |
Glyceraldehyde-3-phosphate
dehydrogenase | Carbon
metabolism |
| 31 | P04797 |
Glyceraldehyde-3-phosphate
dehydrogenase | Carbon
metabolism |
| 26 | Q07936 | Annexin A2 | Apoptosis and
inflammation |
| 27 | P63029 |
Translationally-controlled tumor
protein | Tumor protein |
| 28 | P23764 | Glutathione
peroxidase 3 | Superoxide |
| 32 | Q04374 | Ig κ chain V–II
region 26–10 | Immune system |
|
| B, Proteins with
reduced expression in DM skin |
|
| 37 | P02454 | Collagen α-1 | Tissue
connection |
| 68 | B2RZ77 | Dermatopontin | ECM matrix |
| 38 | P04785 | Protein
disulfide-isomerase | Protein
folding |
| 40 | P04785 | Protein
disulfide-isomerase | Protein
folding |
| 77 | P73842 |
Peptidyl-prolylcis-trans isomerase A | Protein
refolding |
| 65 | P40112 | Proteasome subunit
β type-3 | Protein
degradation |
| 42 | P39423 | Lamin-A/C | Gene
regulation |
| 47 | P70564 | Serpin B5 | Gene
regulation |
| 61 | Q9WUH4 | Four and a half LIM
domains protein 1 | Signaling |
| 44 | P60711 | Actin, cytoplasmic
1 | Cytoskeleton |
| 48 | Q4FZT0 | Stomatin-like
protein 2 | Cytoskeleton |
| 49 | Q6AYC4 | Macrophage-capping
protein | Cytoskeleton |
| 71 | Q4KLF8 | Actin-related
protein 2/3 complex subunit 5 | Cytoskeleton |
| 66 | P02600 | Myosin light chain
1/3 | Cytoskeleton |
| 67 | P13413 | Troponin I | Cytoskeleton |
| 72 | P45592 | Cofilin-1 | Cytoskeleton |
| 73 | P04466 | Myosin regulatory
light chain 2 | Cytoskeleton |
| 74 | P16409 | Myosin regulatory
light chain | Cytoskeleton |
| 45 | P15429 | β-enolase | Carbon
metabolism |
| 46 | P05065 |
Fructose-bisphosphate aldolase A | Carbon
metabolism |
| 43 | P04764 | α-enolase | Carbon
metabolism |
| 50 | P12007 | Isovaleryl-CoA
dehydrogenase | Carbon
metabolism |
| 51 | P16617 | Phosphoglycerate
kinase 1 | Carbon
metabolism |
| 52 | P41562 | Isocitrate
dehydrogenase | Carbon
metabolism |
| 53 | P17764 | Acetyl-CoA
acetyltransferase | Carbon
metabolism |
| 55 | P05065 |
Fructose-bisphosphate aldolase A | Carbon
metabolism |
| 56 | P07943 | Aldose
reductase | Carbon
metabolism |
| 57 | P04797 |
Glyceraldehyde-3-phosphate
dehydrogenase | Carbon
metabolism |
| 58 | P04636 | Malate
dehydrogenase | Carbon
metabolism |
| 62 | P15429 | β-enolase | Carbon
metabolism |
| 69 | P39069 | Adenylate kinase
isoenzyme 1 | ATP synthesis |
| 59 | A7VJC2 | Heterogeneous
nuclear ribonucleoproteins A2/B1 | RNA metabolism |
| 60 | P55260 | Annexin A4 | Apoptosis and
inflammation |
| 63 | P63029 |
Translationally-controlled tumor
protein | Tumor protein |
| 64 | A48593 | Glutathione
S-transferase P1 | Superoxide |
| 75 | P02625 | Parvalbumin α |
Ca2+-binding |
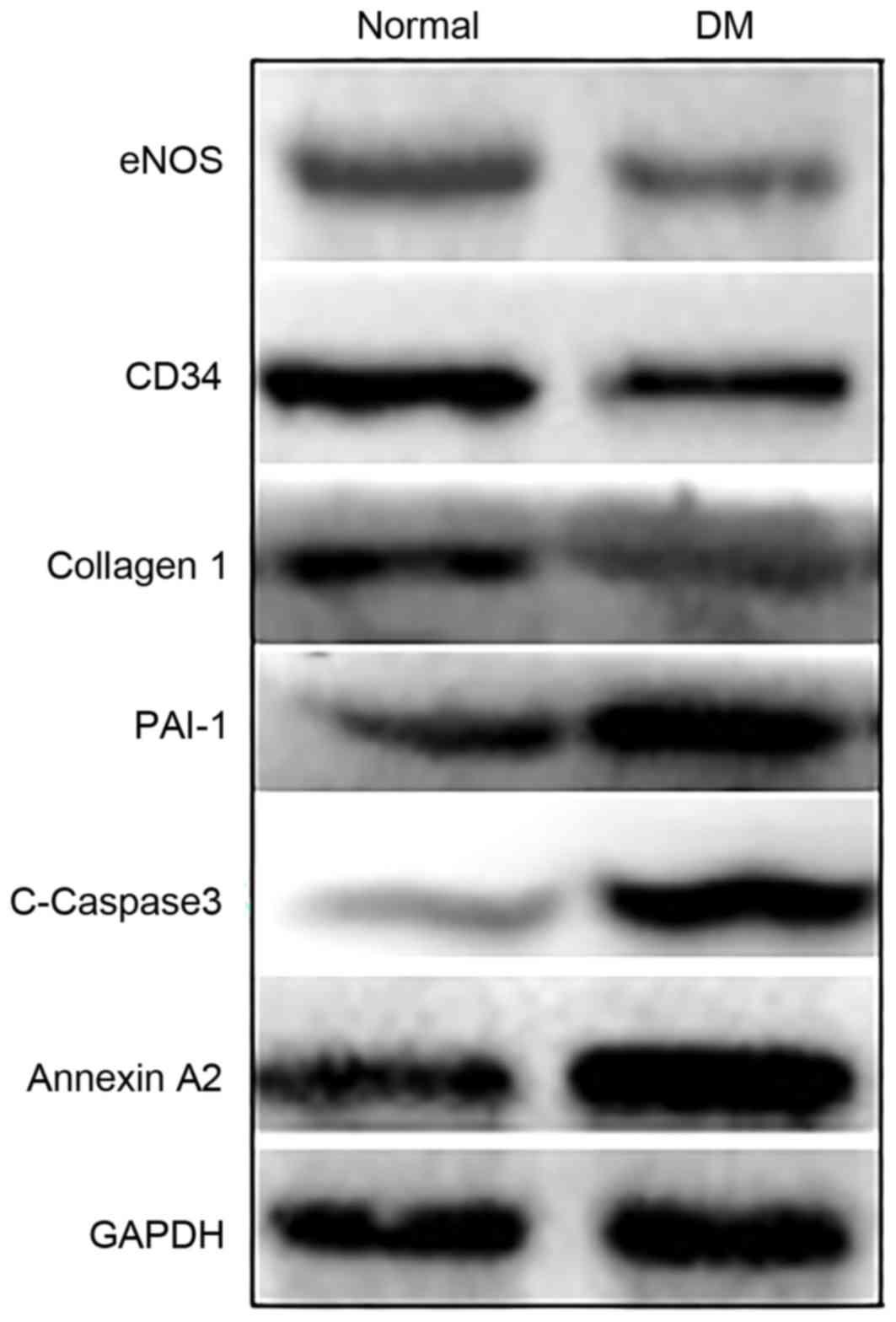
Protein expression was altered in DM
rat skin
To verify the results identified by proteomic assay,
western blot analysis was performed using specific antibodies to
detect select marker proteins in the pathways identified. The
results revealed that the expression levels of CD34, collagen-1 and
endothelial NO synthase were decreased (NO synthesis), whereas
PAI-1 (inflammation), c-caspase3 (apoptosis) and annexin A2 were
increased in DM rat skin when compared with normal rat skin
(Fig. 3). These results were
similar to those identified by the proteomic analysis.
Overexpression of annexin A2 promotes
human foreskin fibroblast cell migration
Annexin A2 was induced in DM rat skin (Fig. 3), and the transcript of its
orthologue gene in humans was previously demonstrated to be induced
by high-glucose treatment of foreskin fibroblast cells (9). Therefore, the function of annexin A2
in fibroblast cells was examined further, by forced overexpression
of annexin A2. Human ANXA2 (GenBank Accession no.
NM_001002858.2) ORF sequences were amplified by PCR, and the PCR
products were then cloned into the pcDNA3 expression vector.
Following sequencing, 2 and 3 µg pcDNA3-ANXA2 plasmid were
transformed into human foreskin primary fibroblast cells using the
Lipofectamine 2000 system to generate two independent
transformants, termed OX1 and OX2, respectively. Compared with the
control cells (untransformed), OX1 and OX2 cells exhibited a
significantly increased cell migration rate following wound
generation (Fig. 4A and B). To
verify the expression levels of annexin A2, western blot analysis
was performed. The results demonstrated that the level of annexin
A2 was increased in OX1 and OX2 when compared with the controls
(Fig. 4C). In addition, annexin A2
overexpression maintained an increased cell migration rate when
compared with the control cells when they were cultured in
high-glucose containing medium (data not shown). However, the level
of p-JNK, a key regulator of cell migration, was not altered by the
overexpression of annexin A2 (Fig.
4C).
Annexin A2 functions downstream of
JNK
As annexin A2 overexpression was not observed to
alter p-JNK protein expression levels, SP600125, an inhibitor of
JNK was used to analyze its effect on annexin A2 expression.
SP600125 significantly inhibited fibroblast cell migration
(Fig. 5A and B), which is
consistent with results presented in a previous study (11). In addition, SP600125 suppressed
p-JNK expression while maintaining normal total JNK expression
levels (Fig. 5C). SP600125
treatment marginally repressed annexin A2 protein expression levels
(Fig. 5C).
Due to the observed inhibitory effects of SP600125
on cell migration and annexin A2 expression levels, its effects
were further examined in OX1 fibroblast cells overexpressing
ANXA2. SP600125+OX1 was associated with a significant
increase in cell migration when compared with SP600125-treated
controls at 12 and 24 h. In addition, SP600125 treatment did not
alter the expression levels of annexin A2 in OX1 transformed cells
(Fig. 6A and B). Western blotting
results demonstrated that SP600125 markedly suppressed p-JNK levels
in OX1 cells (Fig. 6C).
Collectively, these results indicate that annexin A2 may function
downstream of JNK to promote fibroblast cell migration.
Discussion
One of the strategies employed by living organisms
to adapt to environmental alterations, is the rapid reprogramming
of transcriptional and translational regulation via cell signalling
mechanisms. Therefore, the analysis of proteomic alterations
following exposure to specific stressors is a suitable strategy to
understand its regulatory basis (9,10).
In the present study, proteomic analysis was performed on DM rat
skin to identify global alterations in the proteome. The proteomic
results demonstrated that the expression of a number of important
proteins involved in metabolic processes was significantly
altered.
Cell migration, a key process in wound healing, is
multi-step process. During the migration process a protrusion
extends from one cell to attach to a nearby leading edge of another
cell protrusion (3). The cells
then migrate forward and release their adhesions and retractions at
the rear of the cell (15–17). The actin-mediated Wiskott-Aldrich
syndrome protein-family verprolin homologous protein complex is
important in the formation of the lamellipodial protrusions at the
leading edge of the migrating cells (18), and the Rho family guanosine
triphosphatase protein, cell division control protein 42,
reportedly regulates the polarity of migrating cells (19). In the present study,
cytoskeleton-associated proteins were part of a major group of
proteins regulated by DM in rat skin. Actin and the
actin-associated protein complex were repressed by DM, as were
tubulin, myosin, troponin and additional cytoskeleton-associated
proteins. The ECM serves important roles in cell adhesion,
communication between nearby cells, and during differentiation
induced by the deposition of extracellular molecules secreted by
cells (20). In the present study,
collagen α1 and dermatopontin, an ECM-associated protein, were
repressed in DM while moesin, a protein involved in cell-to-cell
recognition, was induced. A previous study identified protein
alterations induced by DM that involved the superoxide
detoxification process (glutathione peroxidase 3 and glutathione
S-transferase P1) and ROS accumulation in rat skin (5), indicating that DM-mediated ROS
accumulation may be induced via inhibition of the superoxide
detoxification pathway. In addition, DM regulation in carbon
metabolism and the ATP synthesis process may affect the supply of
carbon skeletons and energy to the cell, which may in turn inhibit
wound closure (21).
Annexin A2, a member of the annexin protein family,
has been reported to induce cell migration and neoangiogenesis in
breast tumors (22). In addition,
annexin A2 reportedly regulates intestinal epithelial cell
spreading and wound closure (23).
In the present study, annexin A2 was overexpressed while annexin A4
was repressed in DM. Western blot analysis confirmed the increased
levels of annexin A2 in DM rat skin when compared with normal
controls. Similarly, a previous transcriptomics study revealed the
induction of ANXA2 by high-glucose in human foreskin
fibroblast cells (9).
Overexpression of ANXA2 in human foreskin primary fibroblast
cells in the present study resulted in a marked increase in the
rate of cell migration. Furthermore, inhibition of JNK by treating
cells with SP600125, delayed cell migration and reduced annexin A2
levels. However, a significant difference in cell migration was
identified when comparing the SP600126-treated OX1 cells with the
SP600126-treated control cells (Fig.
6), indicating that annexin A2 may function downstream of JNK
during cell migration. A previous study demonstrated that
high-glucose stress increased the nitrosative modification of
annexin A2 in human foreskin fibroblasts, and that regulation
occurred downstream of JNK regulation (5). These results indicated that JNK may
regulate ANXA2 expression and its post-translational
modifications, which may be important for wound repair. In the
present study, the level of annexin A2 was increased in rat DM skin
when compared with normal skin. This may be due to feedback
activation of annexin A2 in an attempt to try to repair the wound
in DM rat skin.
In conclusion, 2-D gel electrophoresis and LC/MS
proteomics were performed to gain insight into the intricate
mechanisms underlying the response to DM-mediated delay of wound
healing in rats. This approach resulted in the identification of a
number of differentially expressed proteins in DM that were
associated with major processes, including carbohydrate metabolism,
protein folding and cell redox homeostasis. The majority of
proteomic studies conducted to date, that have used stress models,
have been performed in rat skin against diabetes. Overexpression of
annexin A2 accelerated the rate of fibroblast migration, and
JNK-annexin A2 was identified as a potential pathway that promotes
cell migration during wound healing. It would be of interest to
analyze skin wound repair in an annexin A2 overexpressed animal
model in future studies.
Acknowledgements
The present study was made possible by an initiative
grant (grant no. QA2576) from Wenzhou Medical University. The
authors would like to thank the undergraduate students from Wenzhou
Medical University for their assistance with the experiments.
References
|
1
|
Yach D, Stuckler D and Brownell KD:
Epidemiologic and economic consequences of the global epidemics of
obesity and diabetes. Nat Med. 12:62–66. 2006. View Article : Google Scholar
|
|
2
|
Braiman-Wiksman L, Solomonik I, Spira R
and Tennenbaum T: Novel insights into wound healing sequence of
events. Toxicol Pathol. 35:767–779. 2007. View Article : Google Scholar
|
|
3
|
Brem H and Tomic-Canic M: Cellular and
molecular basis of wound healing in diabetes. J Clin Invest.
117:1219–1222. 2007. View
Article : Google Scholar :
|
|
4
|
Goldin A, Beckman JA, Schmidt AM and
Creager MA: Advanced glycation end products: Sparking the
development of diabetic vascular injury. Circulation. 114:597–605.
2006. View Article : Google Scholar
|
|
5
|
Xuan YH, Huang BB, Tian HS, Chi LS, Duan
YM, Wang X, Zhu ZX, Cai WH, Zhu YT, Wei TM, et al: High-glucose
inhibits human fibroblast cell migration in wound healing via
repression of bFGF-regulating JNK phosphorylation. PLoS One.
9:e1081822014. View Article : Google Scholar :
|
|
6
|
Obayashi K, Akamatsu H, Okano Y, Matsunaga
K and Masaki H: Exogenous nitric oxide enhances the synthesis of
type I collagen and heat shock protein 47 by normal human dermal
fibroblasts. J Dermatol Sci. 41:121–126. 2006. View Article : Google Scholar
|
|
7
|
Rowe DW, Starman BJ, Fujimoto WY and
Williams RH: Abnormalities in proliferation and protein synthesis
in skin fibroblast cultures from patients with diabetes mellitus.
Diabetes. 26:284–290. 1977. View Article : Google Scholar
|
|
8
|
Loots MA, Lamme EN, Mekkes JR, Bos JD and
Middelkoop E: Cultured fibroblasts from chronic diabetic wounds on
the lower extremity (non-insulin-dependent diabetes mellitus) show
disturbed proliferation. Arch Dermatol Res. 291:93–99. 1999.
View Article : Google Scholar
|
|
9
|
Pang L, Wang Y, Zheng M, Wang Q, Lin H,
Zhang L and Wu L: Transcriptomic study of high-glucose effects on
human skin fibroblast cells. Mol Med Rep. 13:2627–2634. 2016.
|
|
10
|
Xuan Y, Chi L, Tian H, Cai W, Sun C, Wang
T, Zhou X, Shao M, Zhu Y, Niu C, et al: The activation of the
NF-κB-JNK pathway is independent of the PI3K-Rac1-JNK pathway
involved in the bFGF-regulated human fibroblast cell migration. J
Dermatol Sci. 82:28–37. 2016. View Article : Google Scholar
|
|
11
|
Yang Y, Xia T, Zhi W, Wei L, Weng J, Zhang
C and Li X: Promotion of skin regeneration in diabetic rats by
electrospun core-sheath fibers loaded with basic fibroblast growth
factor. Biomaterials. 32:4243–4254. 2011. View Article : Google Scholar
|
|
12
|
Cai L, Wang Y, Zhou G, Chen T, Song Y, Li
X and Kang YJ: Attenuation by metallothionein of early cardiac cell
death via suppression of mitochondrial oxidative stress results in
a prevention of diabetic cardiomyopathy. J Am Coll Cardiol.
48:1688–1697. 2006. View Article : Google Scholar
|
|
13
|
Ni MW, Ye WJ, Cong WT, Hong GY, Zhu ZX,
Duan YM, Zhou X and Jin LT: Fluorescent staining of protein in
sodium dodecyl sulfate polyacrylamide gels by salicylaldehyde
azine. Electrophoresis. 34:3171–3179. 2013. View Article : Google Scholar
|
|
14
|
Russell WK, Park ZY and Russell DH:
Proteolysis in mixed organic-aqueous solvent systems: Applications
for peptide mass mapping using mass spectrometry. Anal Chem.
73:2682–2685. 2001. View Article : Google Scholar
|
|
15
|
Lauffenburger DA and Horwitz AF: Cell
migration: A physically integrated molecular process. Cell.
84:359–369. 1996. View Article : Google Scholar
|
|
16
|
Webb DJ, Parsons JT and Horwitz AF:
Adhesion assembly, disassembly and turnover in migrating cells-over
and over and over again. Nat Cell Biol. 4:E97–E100. 2002.
View Article : Google Scholar
|
|
17
|
Ridley AJ, Schwartz MA, Burridge K, Firtel
RA, Ginsberg MH, Borisy G, Parsons JT and Horwitz AR: Cell
migration: Integrating signals from front to back. Science.
302:1704–1709. 2003. View Article : Google Scholar
|
|
18
|
Smith LG and Li R: Actin polymerization:
Riding the wave. Curr Biol. 14:R109–R111. 2004. View Article : Google Scholar
|
|
19
|
Macara IG: Parsing the polarity code. Nat
Rev Mol Cell Biol. 5:220–231. 2004. View
Article : Google Scholar
|
|
20
|
Abedin M and King N: Diverse evolutionary
paths to cell adhesion. Trends Cell Biol. 20:734–742. 2010.
View Article : Google Scholar :
|
|
21
|
Cong W, Zhao T, Zhu Z, Huang B, Ma W, Wang
Y, Tan Y, Chakrabarti S, Li X, Jin L and Cai L: Metallothionein
prevents cardiac pathological changes in diabetes by modulating
nitration and inactivation of cardiac ATP synthase. J Nutr Biochem.
25:463–474. 2014. View Article : Google Scholar
|
|
22
|
Sharma M, Ownbey RT and Sharma MC: Breast
cancer cell surface annexin II induces cell migration and
neoangiogenesis via tPA dependent plasmin generation. Exp Mol
Pathol. 88:278–286. 2010. View Article : Google Scholar
|
|
23
|
Babbin BA, Parkos CA, Mandell KJ, Winfree
LM, Laur O, Ivanov AI and Nusrat A: Annexin 2 regulates intestinal
epithelial cell spreading and wound closure through Rho-related
signaling. Am J Pathol. 170:951–966. 2007. View Article : Google Scholar :
|