Introduction
Hepatocellular carcinoma (HCC), one of the most
frequently diagnosed malignancies in the world, particularly in
several areas of Asia and Africa, is the third largest cause of
cancer-related death worldwide. Development of HCC is often
associated with chronic liver disease, particularly cirrhosis
caused by hepatitis B virus (HBV) or hepatitis C virus (HCV).
Imaging techniques have detected HCC at an early stage in patients
with chronic HBV or HCV infection (1–4).
Early detection of HCC is rare, whereas very few candidates are
eligible for liver transplantation as a result of the lack of liver
donors (5); thus, recurrence rates
of HCC are very high following medical or surgical treatments
(6,7). Therefore, the development of an
effective targeted gene therapy strategy driven by the
tumor-specific promoter has become an urgent requirement in
treating HCC (8,9).
In recent years, gene therapy has been widely
studied. The aim of gene therapy is to transfect a target gene into
host cells to be specifically expressed, thus killing tumor cells.
Currently, suicide gene therapy has become a promising strategy for
gene therapy, especially the herpes simplex virus thymidine kinase
(HSVtk)/ganciclovir (GCV) system (10). Previous studies have demonstrated
the use of a pro-drug sensitive gene as a suicide gene, to convert
a non-toxic pro-drug into a toxic product, to block the extension
of DNA chains and inhibit the activation of DNA polymerase, finally
leading to cell apoptosis (11–13).
At present, a variety of tumor-specific promoters
have been used for HCC gene therapy, and the alpha-fetoprotein
(AFP) promoter has become an ideal target (14). The regulatory regions and sequence
of the AFP gene not only specifically activate transcription of
exogenous genes but also have significant effects on tumor-specific
transcriptional activity (15).
Furthermore, in clinical application, AFP gene has been used as a
specific marker of HCC.
Therefore, the purpose of the present study was to
construct a plasmid, pcDNA3.1-pAFP-TK, to express thymidine kinase
(TK) driven by the AFP promoter, to study the selective killing
effect on HCC cells.
Materials and methods
Cell lines and culture
HL-7702 human liver cell line and HepG2 human HCC
cell line were purchased from Shanghai Cell Bank of Chinese Academy
of Science (Shanghai, China). HeLa human cervical cancer cells were
purchased from Basic Medical Cell Bank of Chinese Peking Union
Medical University (Beijing, China). HL-7702 cells were maintained
in Iscove's modified Dulbecco's medium (IMDM; Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA) supplemented with 10%
fetal bovine serum (FBS; Thermo Fisher Scientific, Inc., Waltham,
MA, USA), 4 mM L-glutamine, 100 U/ml penicillin and 100 mg/ml
streptomycin. HeLa and HepG2 cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; Hyclone; GE Healthcare Life
Sciences) supplemented with 10% FBS, 4 mM L-glutamine, 100 U/ml
penicillin and 100 mg/ml streptomycin. All cells were maintained at
37°C in a humidified atmosphere containing 5% CO2.
Construction of the plasmid
pcDNA3.1-pAFP-TK and bacterial transformation
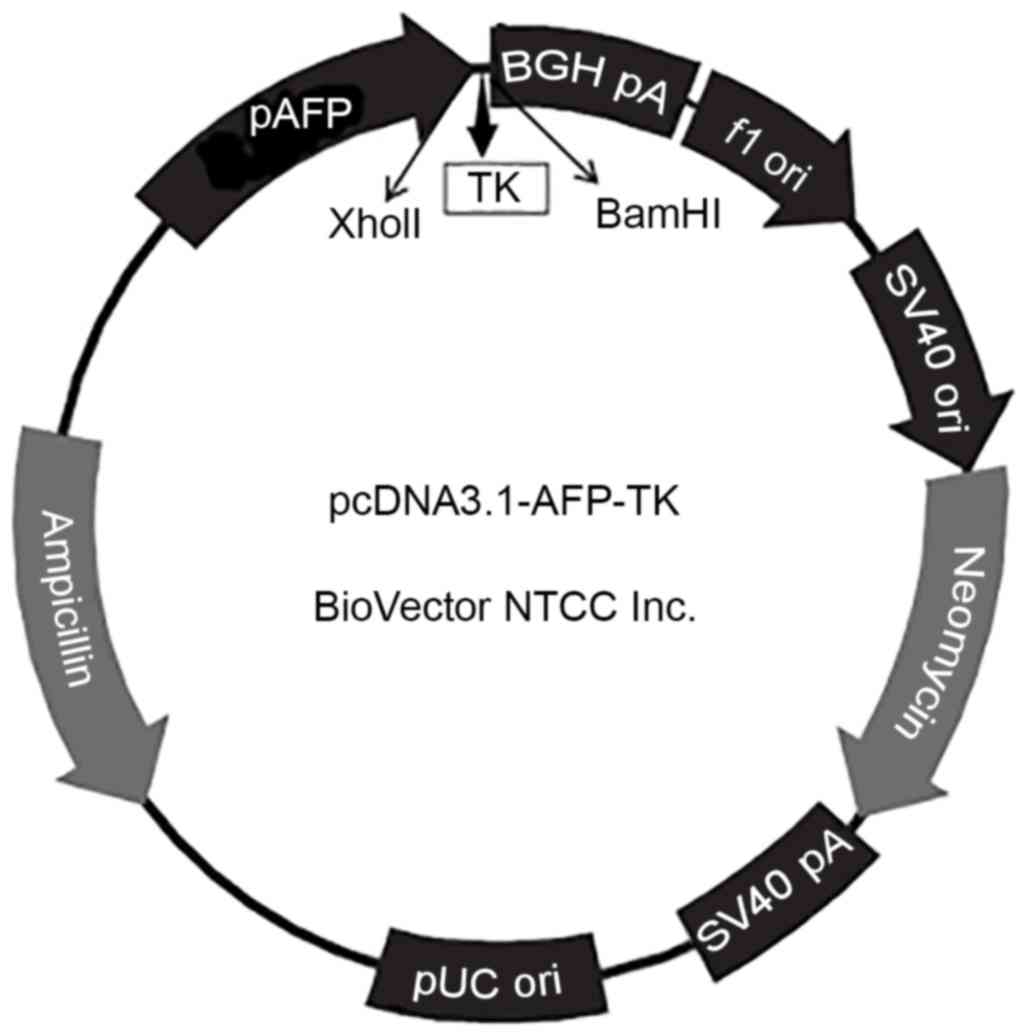
The pcDNA3.1-pAFP-TK plasmid (Fig. 1) was synthesized by BioVector, Inc.
(Beijing, China). Chemically competent Escherichia coli DH5α
cells were purchased from Beijing Solarbio Science & Technology
Co., Ltd. (Beijing, China). Cells (50 µl) were thawed on ice, then
5 ng of plasmid DNA was added, mixed gently, and the mixture
incubated on ice for 30 min. Cells were then transferred to a 42°C
water bath for 90 sec, then placed on ice for a further 2 min.
Sterile lysogeny broth (LB; 400 µl; Beijing Solarbio Science &
Technology Co., Ltd.) without antibiotic was added, and the mixture
was incubated for 3 h at 37°C in a shaker at 200 rpm/min to recover
the cells. The mixture was then centrifuged at 1,006.2 × g for 3
min at 4°C, the supernatant was discarded, and the pellet was
resuspended in 100 µl LB. Cells were plated on solid LB containing
agar and 50 mg/ml ampicillin (Sangon Biotech Co., Ltd., Shanghai,
China) and incubated at 37°C for 12–16 h. Single colonies were then
picked into 100 ml LB and incubated at 37°C for 16 h in a shaker at
260 rpm/min.
Extraction and identification of the
pcDNA3.1-pAFP-TK plasmid
Plasmid extraction kit was purchased from Sangon
Biotech Co., Ltd. and pcDNA3.1-pAFP-TK plasmid DNA was extracted
from E.coli DH5α according to the manufacturer's protocol.
Plasmid DNA was digested with XhoI and BamHI and the
products were visualized under UV transillumination following
separation on a 1% agarose gel stained with fluorescence staining
dye Goldviewna I (Beijing Solarbio Science & Technology Co.,
Ltd.), using the DNA marker DM 2000 Plus (CWBio, Beijing, China).
DNA fragments were subsequently sequenced by BioVector, Inc., and
Basic Local Alignment Search Tool (BLAST; National Center for
Biotechnology Information, National Institutes of Health, Bethesda,
MD, USA) was used to analyze the homology of the HSVtk gene
and plasmid DNA sequences.
Cell transfection
HL-7702, HeLa and HepG2 cells were transfected with
Lipofectamine 2000 Reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocols. For MTT assays,
HepG2 cells were cultured to the exponential phase of growth and
then were seeded in 96 well plates at a density of 104
cells per well. For flow cytometry, HepG2 cells were incubated in 6
well plates at a density of 5×105 cells per well. For
the transfection experiments, cells were divided into the
pcDNA3.1-pAFP-TK group and the control group and were incubated
overnight in 5% CO2 at 37°C. Cells in the control group
received no treatment. The following day, cells in the
pcDNA3.1-pAFP-TK group were washed twice with PBS and fresh
serum-free DMEM was added to each well. Transfection was conducted
with 7 µl Lipofectamine 2000 and 2.5 µg plasmid DNA. Following 6 h
incubation in 5% CO2 at 37°C, the cell medium was
replaced with medium containing 10% FBS and the cells were
incubated for a further 48 h an additional culture of 48 h.
Reverse transcription-polymerase chain
reaction (RT-PCR)
mRNA expression of HSVtk was analyzed by
RT-PCR. HL-7702, HeLa and HepG2 cells were transfected
withcDNA3.1-pAFP-TK, then total RNA was extracted using
TRIzol® reagent (Thermo Fisher Scientific, Inc.). cDNA
was reverse transcribed from the total RNA according to the
manufacturer's protocol, using dNTP Mix, 5X RT buffer, HiFiScript
1st Strand cDNA Synthesis kit and RNase-free water (CWBio). PCR was
then performed, using Goldviewna I (Beijing Solarbio Science &
Technology Co., Ltd.) as the fluorophore, to amplify TK, with
β-actin as an internal control, using the following primer
sequences: TK, forward 5′-CAACAAAAAGCCACGGAAGT-3′ and reverse
5′-ATGCTGCCCATAAGGTATCG-3′; and β-actin, forward
5′-TGACGTGGACATCCGCAAAG-3′ and reverse 5′-CTGGAAGGTGGACAGCGAGG-3′.
The amplification products of TK and β-actin were, respectively,
446 and 205 bp in length. PCR was performed using the following DNA
thermal cycler conditions: 1 cycle of 94°C for 2 min; 30 cycles of
94°C for 30 sec, 55°C for 30 sec, 72°C for 1 min; and a final
elongation step of 72°C for 5 min. The PCR products were visualized
under UV transillumination following separation by 1% agarose gel
electrophoresis stained with fluorescence staining dye Goldviewna I
alongside a DNA marker (Takara Biotechnology Co., Ltd., Dalian,
China).
Western blot
HL-7702, HeLa and HepG2 cells were treated as
described. Total protein was extracted following transfection using
a radioimmunoprecipitation assay lysis buffer (Beyotime Institute
of Biotechnology, Haimen, China) at 4°C for 30 min, and the lysates
were centrifuged at 4°C for 20 min at 4,360.2 × g. Protein
concentration in cell lysates was determined using a bicinchoninic
acid protein assay kit (Boster Systems, Inc., Pleasanton, CA, USA).
Proteins were then separated by 5–12% SDS-PAGE, transferred onto
polyvinylidene fluoride membranes, and blocked by incubation in 5%
skim milk in TBS containing 0.05% Tween-20 at room temperature for
2 h. Membranes were then incubated overnight at 4°C with goat
polyclonal anti-HSVtk (1:800; cat no. sc-28038; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) and anti-β-actin (1:800; cat
no. TA-09; ZSGB-BIO, Beijing, China) primary antibodies, washed in
TBS containing 0.1% Tween-20 (TBST) for 3×10 min, incubated at 4°C
for 1 h with horseradish peroxidase-conjugated secondary antibody
(1:5,000; cat no. A0181; Beyotime Institute of Biotechnology), then
the membranes washed again in TBST for 3×10 min. Finally, the
expression of HSVtk and β-actin was visualized using enhanced
chemiluminescence (Wuhan Boster Biological Technology, Ltd., Wuhan,
China).
MTT assay
HepG2 cells transfected with pcDNA3.1-pAFP-TK (100
µl transfected cells) were seeded in 96-well plates at a density of
104 cells per well and incubated overnight at 37°C in a
humidified atmosphere containing 5% CO2. Cells were then
treated with 0, 1, 5, 10, 20, 40, 60 or 80 µg GCV and incubated in
5% CO2 at 37°C. After 4 days, cells were observed under
the microscope. MTT substrate (20 µl) was added to each well and
the plates were incubated in 5% CO2 at 37°C for a
further 4 h. The medium was then discarded and 150 µl DMSO were
added to each well at room temperature for 10 min. A microplate
reader was used to measure the absorbance at 490 nm
(A490), and the inhibition rate was calculated as:
[1-(A490 of pcDNA3.1-pAFP-TK group/A490 of
the control group)]x100%.
Detection of cell apoptosis by flow
cytometry
Cell apoptosis was detected with an Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis
detection kit (Invitrogen; Thermo Fisher Scientific, Inc.).
pcDNA3.1-pAFP-TK was transfected into HepG2 cells. Cells in the
control group received no transfection and no further treatments.
Following 48 h incubation, the pcDNA3.1-pAFP-TK group was treated
with 150 µg/ml GCV for 2 days. Cells were harvested with 0.25%
trypsin then sedimented by centrifugation at 335.4 × g for 3 min at
room temperature. The supernatant was then discarded, cells were
washed twice with PBS, then 100 µl binding buffer (Nanjing KeyGen
Biotech Co., Ltd., Nanjing, China) was added to resuspend the
cells. Cell suspension (100 µl) was then added to the flow tube,
along with 5 µl Annexin V-FITC at a final concentration of 1 µg/ml
and 10 µl (250 ng) of PI. The cells were mixed and incubated for 15
min at room temperature in the dark. Finally, 400 µl binding buffer
was added and flow cytometry was used to detect cell apoptosis,
using the BD FACSCalibur™ flow cytometer and the BD FACStation™
software (BD Biosciences, Franklin Lakes, NJ, USA).
Detection of mitochondrial membrane
potential by flow cytometry
The mitochondrial membrane potential apoptosis
detection kit was purchased from Beijing ComWin Company (Beijing,
China). HepG2 cells were transfected with pcDNA3.1-pAFP-TK, whereas
the control group received no intervention. Following 48 h
incubation, the pcDNA3.1-pAFP-TK group was treated with 150 µg/ml
GCV for 2 days. The culture media was then discarded, the cells
were washed once with PBS, then 1 ml cell culture medium and 1 ml
JC-1 staining solution was added and the cells were incubated in a
cell culture incubator at 37°C for 20 min. The supernatant was then
removed and the cells were washed twice with 1X JC-1 staining
buffer (Abcam, Cambridge UK). Finally, 2 ml of cell culture medium
was added to the cells and fluorescence was observed under a
fluorescence microscope, to reflect cell apoptosis. Three slides
were observed and >10 fields of view/slide were assessed.
Detection of activated caspase-3 by
caspase-3 staining
The Active Caspase-3 Staining kit was purchased from
BioVision, Inc. (Milpitas, CA, USA). HepG2 cells in 6 well plates
were transfected with pcDNA3.1-pAFP-TK using Lipofectamine2000
(Invitrogen; Thermo Fisher Scientific, Inc.), alongside an
untreated control group. The pcDNA3.1-pAFP-TK group was treated
with 150 µg/ml GCV for 48 h, then all the cells were collected by
centrifugation at 4°C at 335.4 × g for 3 min. Cells were then
resuspended in 300 µl cell culture medium with 1 µl Red-DEVD-FMK
and incubated for 1 h at 37°C with 5% CO2. Cells were
then centrifuged at 335.4 × g for 3 min, the supernatant was
discarded, and cells were resuspended in 100 µl water buffer
provided in the kit. Finally, a drop of the cell suspension was
placed onto a microslide and covered with a coverslip. Red
fluorescence was observed under a fluorescence microscope, to
evaluate the levels of activated caspase-3. Three slides were
observed and >10 fields of view/slide were assessed.
Statistical analysis
Data was expressed as the mean ± standard deviation,
and all assays were performed in triplicate. Statistical
differences were evaluated by Student's t-test using SPSS 16.0
software (SPSS Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Detection of the pcDNA3.1-pAFP-TK
plasmid
Following digestion of pcDNA3.1-pAFP-TK was digested
with XholI and BamHI, a 1131-bp fragment was detected
by agarose gel electrophoresis (Fig.
2), which demonstrated that the HSVtk gene was
successfully inserted into the plasmid pcDNA3.1-pAFP-TK.
Sequence analysis of pcDNA3.1-pAFP-TK
by BLAST
DNA sequencing demonstrated that the construction of
the plasmid pcDNA3.1-pAFP-TK was successful and that the
pcDNA3.1-pAFP-TK vector had a 1131-bp fragment (Fig. 3A). This fragment was confirmed to
be an insertion of the HSVtk gene by BLAST (Fig. 3B).
Detection of HSVtk mRNA
expression
RT-PCR was used to analyze the expression of
HSVtk mRNA in the HL-7702 cells, the HeLa cells and the
HepG2 cells. As demonstrated in Fig.
4, a 446 bp product was observed in HepG2 cells transfected
with the plasmid pcDNA3.1-pAFP-TK, but no product was observed in
HL-7702 cells or HeLa cells. Therefore, the result indicated that
the expression of HSVtk gene mRNA had a high level in the
HepG2 cells.
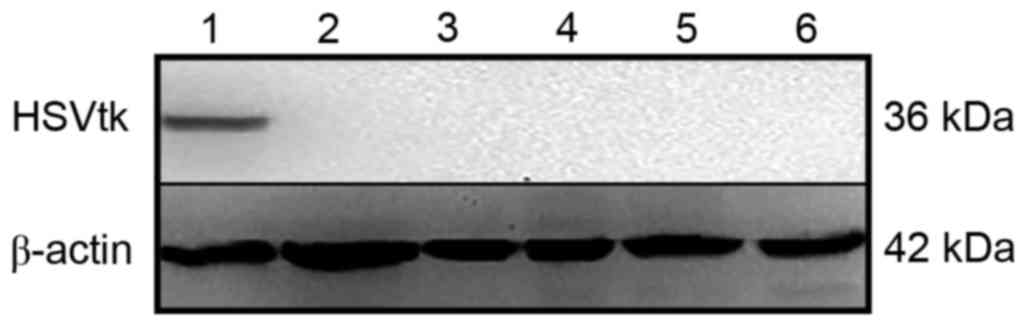
HSVtk suicide gene protein
expression
The expression of HSVtk protein was
demonstrated by western blotting. A 36 kDa protein band was
detected in the HepG2 cells transfected with pcDNA3.1-pAFP-TK, but
no expression was observed in HL-7702 cells or HeLa cells (Fig. 5). The result suggested that
HSVtk was highly expressed in HepG2 cells.
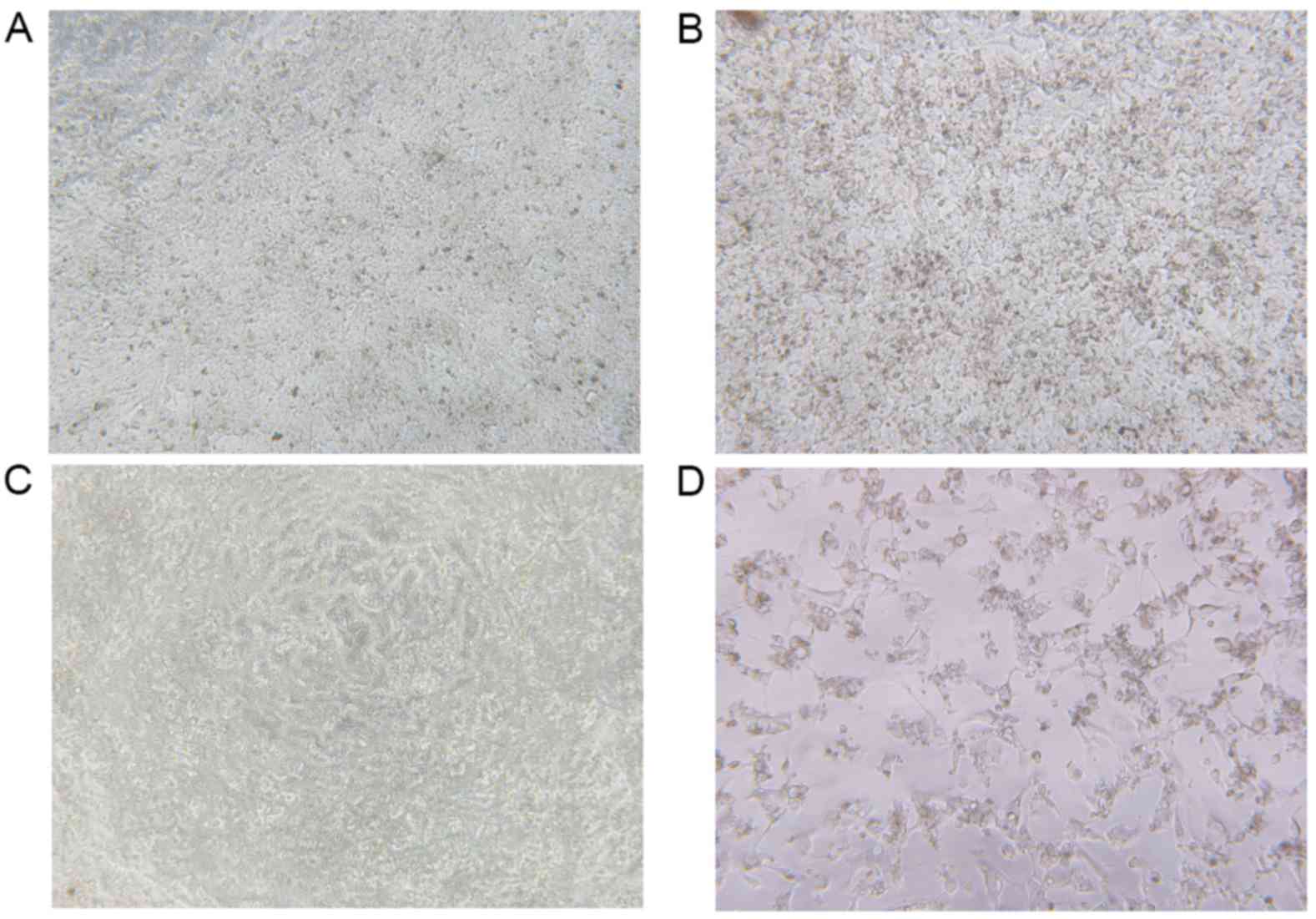
Cell viability assay
MTT assays were performed to investigate cell
viability in the HepG2 cells interfered by the HSVtk/GCV suicide
gene system. With high levels of GCV, transfected cells were killed
and the cell morphology was altered compared with untransfected and
untreated cells (Fig. 6). As
demonstrated by the GCV dose-response curve (Table I and Fig. 7), cell viability in the
pcDNA3.1-pAFP-TK group gradually reduced compared with the control
group as GCV concentration increased.
 | Table I.Cell viability in response to GCV
treatment. |
Table I.
Cell viability in response to GCV
treatment.
|
| GCV (µg) |
|---|
|
|
|
|---|
| Groups | 0 | 1 | 5 | 10 | 20 | 40 | 60 | 80 |
|---|
| Control | 0.542±0.017 | 0.592±0.028 | 0.545±0.038 | 0.571±0.027 | 0.522±0.070 | 0.526±0.110 | 0.508±0.091 | 0.522±0.072 |
|
pcDNA3.1-pAFP-TK | 0.542±0.160 | 0.537±0.110 | 0.357±0.025 | 0.322±0.038 | 0.246±0.045 | 0.198±0.014 | 0.157±0.024 | 0.090±0.075 |
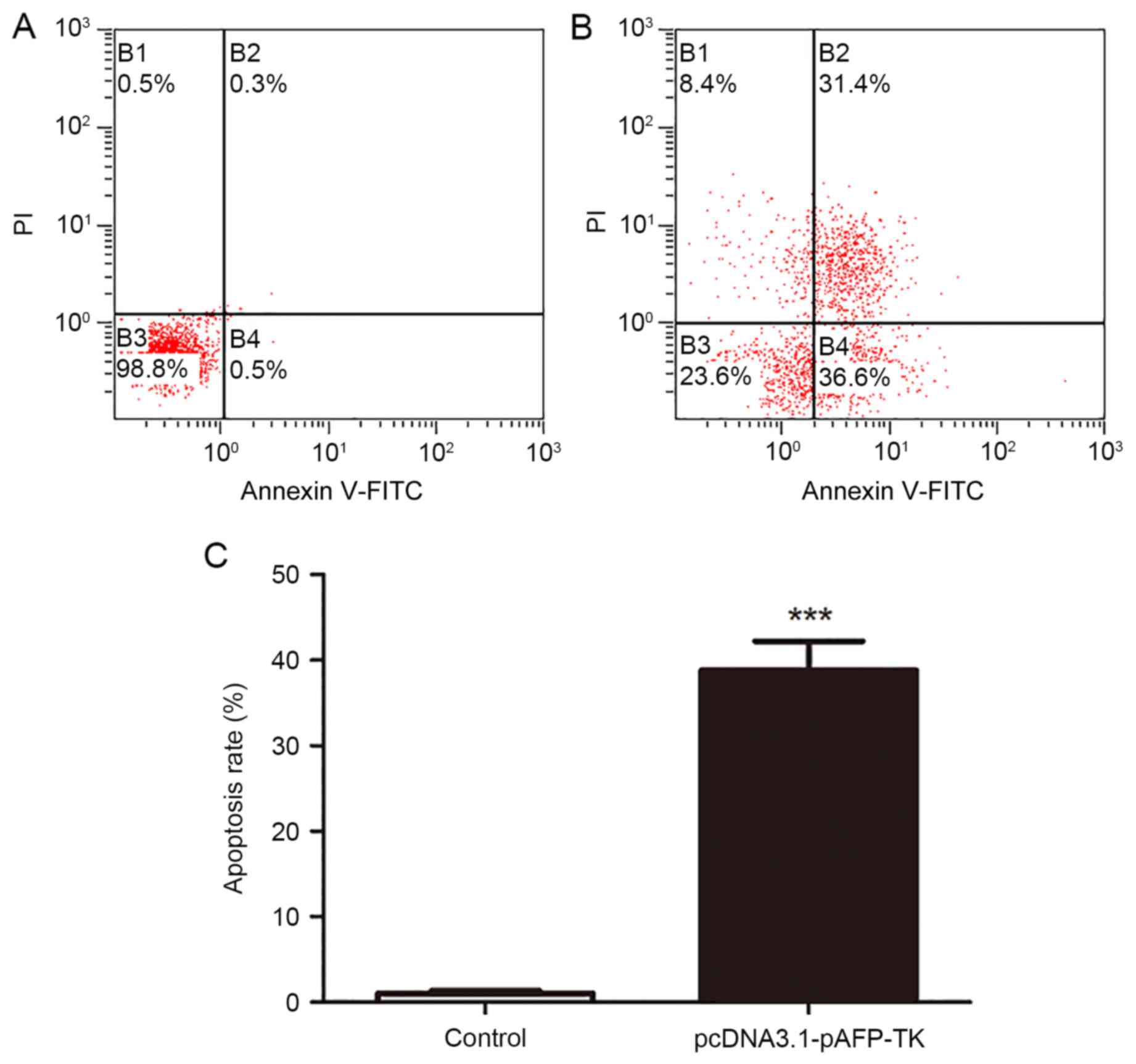
Detection of apoptosis by flow
cytometry
As demonstrated in Fig.
8, the apoptosis rate in the pcDNA3.1-pAFP-TK group
(38.70±6.03%) was significantly higher than the apoptosis rate in
the control group (1.00±0.62%; P<0.001).
Detection of mitochondrial membrane
potential by flow cytometry
As demonstrated in Fig.
9, the mitochondrial membrane potential reduction rate in the
pcDNA3.1-pAFP-TK group (22.84±5.79%) was significantly higher than
the rate in the control group (0.57±0.11; P<0.01).
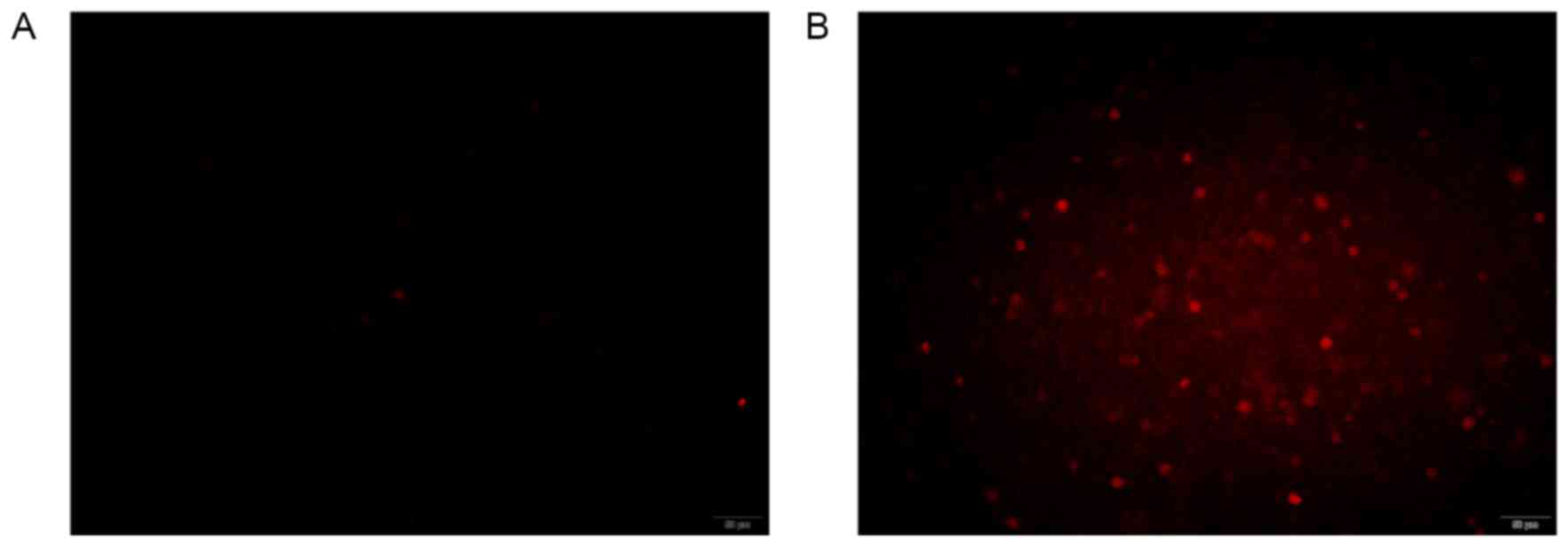
Detection of activated caspase-3 by
fluorescence microscopy
Caspase-3 staining was used to detect activated
caspase-3 in the HepG2 cells. The results demonstrated that
compared with control cells (Fig.
10A), HepG2 cells interfered with HSVtk/GCV suicide gene system
emitted a brighter red signal (Fig.
10B), indicating increased levels of activated caspase-3.
Activated caspase-3 played an important role in the early stage
apoptotic cells, therefore, HSVtk/GCV suicide gene system had a
significant killing effect on the HepG2 cells.
Discussion
The worldwide incidence and mortality rates of HCC
appear to be increasing year by year, the incidence of which ranked
the fifth among all cancer cases (16). HCC represents the third leading
cause of cancer-associated mortality worldwide (17,18).
Based on the cellular and molecular levels, targeted therapy for
combining drugs with the specific target could kill tumor cells,
but rarely threatened normal tissues and cells. Gene therapy aims
to insert exogenous normal genes into target cells to compensate
for genetic defects and disease-related abnormalities. The
combination of targeted therapy with gene therapy acts as a
powerful target gene for killing human hepatoma carcinoma
cells.
The two most widely used suicide gene systems are
the CD/5-FC system and the HSVtk/GCV system (19). In the CD/5-FC system, the cytosine
deaminase gene (CD) of some bacteria and fungi generates the
CD enzyme, which converts cytosine to uracil; non-toxic
5-fluorocytosine is transformed into cytotoxic 5-fluorouracil
(5-FU), thereby killing tumor cells (20). HSVtk efficiently
phosphorylates non-toxic gancyclovir (GCV) to produce
phosphorylated products that lead to an arrest of DNA synthesis and
cell death (21,22). However, normal cells could be
killed as a result of lack of specific suicide gene, therefore,
improved efficiency of targeted gene therapy is essential. It was,
therefore, imperative to construct a novel recombinant vector, to
be transfected into tumor cells with a targeted ability to kill
cells. Previous studies have demonstrated that some biochemical
markers were tied to hepatoma carcinoma cells, such as AFP,
vascular endothelial growth factor (VEGF) (23,24).
AFP is a major serum protein produced by fetal
hepatocytes. It is not detectable in normal adult cells, however,
in HCC cells, the AFP gene is highly expressed (25,26).
Therefore, AFP sequences could be used to regulate expression of
cytotoxic genes, as in the HSVtk/GCV system, through use of the AFP
promoter; the system has been reported to have a limited effect on
hepatoma cells (27). The
HSVtk/GCV system was a successful suicide gene therapy strategy for
HCC (28,29).
The results of the present study demonstrated that
the novel plasmid pcDNA3.1-pAFP-TK, driven by the AFP promoter, was
constructed successfully. The plasmid pcDNA3.1-pAFP-TK was
transfected into HL-7702, HeLa, and HepG2 cells. RT-PCR and western
blot demonstrated that HSVtk was effectively expressed in
HepG2 cells transfected with the plasmid pcDNA3.1-pAFP-TK, whereas
HSVtk gene expression was not detected in HL-7702 and HeLa
cells. Furthermore, MTT assays indicated that, with increasing of
GCV doses, the HepG2 cells viability significantly decreased; cell
viability was significantly affected in HepG2 cells transfected
with pcDNA3.1-pAFP-TK by 5 µg GCV, and when GCV was increased to 80
µg, it was evident that that cell viability was severely
suppressed. In addition, flow cytometry demonstrated that in HepG2
cells treated with HSVtk/GCV suicide gene system, cell apoptosis
rates and mitochondrial membrane potential reduction rates were
increased dramatically in comparison with the control group.
Caspase-3 staining demonstrated that activated caspase-3 increased
significantly in the HepG2 cells with the HSVtk/GCV suicide gene
system, supporting the cell apoptosis results, whereas increased
activated caspase-3 was not observed in the control group.
Therefore, the present study has constructed a novel
plasmid vector driven by the human AFP promoter, which may has
become an effective approach in overcoming the restrictions of
current technologies.
Acknowledgements
The present study was funded by grants from the
National Natural Science Foundation of China (grant nos. 30901821
and 81172136) and Natural Science Foundation of Shanxi (grant no.
2015011113).
References
|
1
|
Bosch FX, Ribes J, Diaz M and Cléries R:
Primary liver cancer: Worldwide incidence and trends.
Gastroenterology. 127:(5 Suppl 1). S5–S16. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Levin B and Amos C: Therapy of
unresectable hepatocellular carcinoma. N Engl J Med. 332:1294–1296.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Arii S, Yamaoka Y, Futagawa S, Inoue K,
Kobayashi K, Kojiro M, Makuuchi M, Nakamura Y, Okita K and Yamada
R: Results of surgical and nonsurgical treatment for small-sized
hepatocellular carcinomas: A retrospective and nationwide survey in
Japan. The liver cancer study group of Japan. Hepatology.
32:1224–1229. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rongrui L, Na H, Zongfang L, Fanpu J and
Shiwen J: Epigenetic mechanism involved in the HBV/HCV-related
hepatocellular carcinoma tumorigenesis. Curr Pharm Des.
20:1715–1725. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Llovet JM, Di Bisceglie AM, Bruix J,
Kramer BS, Lencioni R, Zhu AX, Sherman M, Schwartz M, Lotze M,
Talwalkar J, et al: Design and endpoints of clinical trials in
hepatocellular carcinoma. J Natl Cancer Inst. 100:698–711. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cheng AL, Kang YK, Chen Z, Tsao CJ, Qin S,
Kim JS, Luo R, Feng J, Ye S, Yang TS, et al: Efficacy and safety of
sorafenib in patients in the Asia-Pacific region with advanced
hepatocellular carcinoma: A phase III randomised, double-blind,
placebo-controlled trial. Lancet Oncol. 10:25–34. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liver Cancer Study Group of Japan: Primary
liver cancer in Japan. Clinicopathologic features and results of
surgical treatment. Ann Surg. 211:277–287. 1990.PubMed/NCBI
|
|
8
|
Schmitz V, Qian C, Ruiz J, Sangro B,
Melero I, Mazzolini G, Narvaiza I and Prieto J: Gene therapy for
liver diseases: Recent strategies for treatment of viral hepatitis
and liver malignancies. Gut. 50:130–135. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dachs GU, Dougheay GJ, Stratford IJ and
Chaplin DJ: Targeting gene therapy to cancer: A review. Oncol Ras.
9:313–325. 1997.
|
|
10
|
Ferrara F, Staquicini DI, Driessen WH,
D'Angelo S, Dobroff AS, Barry M, Lomo LC, Staquicini FI, Cardó-Vila
M, Soghomonyan S, et al: Targeted molecular-genetic imaging and
ligand-directed therapy in aggressive variant prostate cancer. Proc
Natl Acad Sci USA pii: 201615400. 2016. View Article : Google Scholar
|
|
11
|
Anderson WF: Gene therapy for cancer. Hum
Gene Ther. 5:1–2. 2008. View Article : Google Scholar
|
|
12
|
Huber BE, Richards CA and Krenitsky TA:
Retroviral-mediated gene therapy for the treatment of
hepatocellular carcinoma: All innovative approach for therapy. Proc
Natl Acad Sci USA. 88:8039–8043. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yashiyasu K and Ayumi T: Gene therapy of
hepatoma: Bystander effects and non-apoptotic cell death induced by
thymidine kinase and ganciclovir. Cancer Lett. 96:105–110. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma WJ, Wang HY and Teng LS: Correlation
analysis of preoperative serum alpha-fetoprotein (AFP) level and
prognosis of hepatocellular carcinoma (HCC) after hepatectomy.
World J Surg Oncol. 11:2122013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Su H, Chang JC, Xu SM and Kan YW:
Selective killing of AFP-positive hepatocellular carcinoma cells by
adeno-associated virus transfer of the herpes simplex virus
thymidine kinase gene. Hum Gene Ther. 7:463–470. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cahill BA and Braccia D: Current treatment
for hepatocellular carcinoma. Clin J Oncol Nurs. 8:393–399. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Farazi PA and DePinho RA: Hepatocellular
carcinoma pathogenesis: From genes to environment. Nat Rev Cancer.
6:674–687. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Varela M, Sala M, Llovet JM and Bruix J:
Treatment of hepatocellular carcinoma: Is there an optimal
strategy. Cancer Treat Rev. 29:99–104. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huber BE, Austin EA, Richards CA, Davis ST
and Good SS: Metabolism of 5-fluorocytidineto5-fluorouracic in
human colorectal tumor cells transduced with the cytosine deaminase
gene: Significant antitumor effect when only a small percentage of
tumor cells express cytosine deaminase. Proc Natl Acad Sci USA.
91:8302–8306. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang XP, Liu L, Wang P and Ma SL: Human
sulfatase-1 improves the effectiveness of cytosine deaminase
suicide gene therapy with 5-Fluorocytosine treatment on
hepatocellular carcinoma cell line HepG2 in vitro and in vivo. Chin
Med J (Engl). 128:1384–1390. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tsuchiya K, Asahina Y, Matsuda S, Muraoka
M, Nakata T, Suzuki Y, Tamaki N, Yasui Y, Suzuki S, Hosokawa T, et
al: Changes in plasma vascular endothelial growth factor at 8 weeks
after sorafenib administration as predictors of survival for
advanced hepatocellular carcinoma. Cancer. 120:229–237. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sadeghi M, Lahdou I, Oweira H, Daniel V,
Terness P, Schmidt J, Weiss KH, Longerich T, Schemmer P, Opelz G
and Mehrabi A: Serum levels of chemokines CCL4 and CCL5 in
cirrhotic patients indicate the presence of hepatocellular
carcinoma. Br J Cancer. 113:756–762. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qu L, Wang Y, Gong L, Zhu J, Gong R and Si
J: Suicide gene therapy for hepatocellular carcinoma cells by
surviving promoter-driven expression of the herpes simplex virus
thymidine kinase gene. Oncol Rep. 29:1435–1440. 2013.PubMed/NCBI
|
|
24
|
Yu BF, Wu J, Zhang Y, Sung HW, Xie J and
Li RK: Ultrasound-targeted HSVtk and Timp3 gene delivery for
synergistically enhanced antitumor effects in hepatoma. Cancer Gene
Ther. 20:290–297. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tilghman SM and Belayew A: Transcriptional
control of the murine albumin/alpha-fetoprotein locus during
development. Proc Natl Acad Sci USA. 79:5254–5257. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sakai M, Morinaga T, Urano Y, Watanabe K,
Wegmann TG and Tamaoki T: The human alpha-fetoprotein gene.
Sequence organization and the 5′flanking region. J Biol Chem.
260:5055–5060. 1985.PubMed/NCBI
|
|
27
|
Kanai F, Shiratori Y, Yoshida Y, Wakimoto
H, Hamada H, Kanegae Y, Saito I, Nakabayashi H, Tamaoki T, Tanaka
T, et al: Gene therapy for alpha-fetoprotein-producing human
hepatoma cells by adenovirus-mediated transfer of the herpes
simplex virus thymidine kinase gene. Hepatology. 23:1359–1368.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gerolami R, Uch R, Faivre J, Garcia S,
Hardwigsen J, Cardoso J, Mathieu S, Bagnis C, Brechot C and Mannoni
P: Herpes simplex virus thymidine kinase-mediated suicide gene
therapy for hepatocellular carcinoma using HIV-1-derived lentiviral
vectors. J Hepatol. 40:291–297. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sakai Y, Kaneko S, Nakamoto Y, Kagaya T,
Mukaida N and Kobayashi K: Enhanced anti-tumor effects of herpes
simplex virus thymidine kinase/ganciclovir system by codelivering
monocyte chemoattractant protein-1 in hepatocellular carcinoma.
Cancer Gene Ther. 8:695–704. 2001. View Article : Google Scholar : PubMed/NCBI
|