Introduction
Inflammatory bowel disease (IBD) is characterized by
chronic intestinal inflammation that results from an abnormal
immune response to environmental factors in genetically susceptible
individuals. IBD includes ulcerative colitis (UC) and Crohns
disease (CD) (1). The incidence of
IBD is increasing worldwide (2);
however, its etiology and pathogenesis are not fully understood.
Trefoil factor 3 (TFF3) belongs to the trefoil factor family and
contains one trefoil domain of 59 amino acids, with a molecular
weight of ~6.6 kDa (monomer) or 13 kDa (dimer). TFF3 serves an
important role in intestinal mucosal damage and mucosal healing
(3). TFF3 promotes epithelial
restitution resulting in mucosal protection by interacting with
mucin, and enhances the structural integrity of the mucosal
barrier. TFF3 can reconstruct the epithelial barrier by stimulating
epithelial cell migration and proliferation, and by influencing the
expression of tight junction proteins. Furthermore, it maintains
the integrity of the epithelial barrier by inhibiting apoptosis and
regulating immune responses. The physiological functions of TFF3
have been well characterized; however, the regulatory mechanisms
controlling TFF3 remain unclear.
MicroRNAs (miRNAs/miR) are small 22-nucleotide,
noncoding single-stranded RNA molecules, which bind to the 3′
untranslated region (3′UTR) of their target mRNAs, interfering with
RNA expression and inducing either mRNA degradation or
translational repression (4).
Numerous miRNA functions have been described in various
autoimmune-associated conditions, including psoriasis, rheumatoid
arthritis, lupus and asthma (5–7). A
previous study observed differential miRNA expression in patients
with active UC and CD (8). In
addition, previous studies have revealed that miRNAs exert powerful
gene regulatory functions, which can affect the function of
Toll-like receptors (9), regulate
the expression of inflammatory cytokines, enhance the permeability
of tight junctions, increase apoptosis in epithelial cells and
regulate the occurrence and development of IBD (10). Ye et al (11) demonstrated that tumor necrosis
factor-α increased the expression of miR-122, which degraded tight
junction proteins and increased the intestinal permeability of a
filter-grown Caco-2 monolayer. Identification of miRNAs that
regulate TFF3 expression may improve understanding regarding the
pathogenesis of IBD and lead to novel approaches for the diagnosis
and treatment of this disease. The present study aimed to
investigate if miR-7-5p binds to the 3′UTR and regulates the
expression of TFF3, and to explore the expression of miR-7-5p and
TFF3 in IBD tissues.
Materials and methods
Bioinformatics prediction
Bioinformatics prediction software (Targetscan
version 6.2; http://www.targetscan.org/vert_61/) was used to
predict the miRNAs that would bind to the 3′UTR of TFF3.
Cell culture experiments
The human colonic epithelial cell line LS174T was
obtained from American Type Culture Collection (Manassas, VA, USA).
The cell line was grown to near confluence in Dulbecco's modified
Eagle's medium (DMEM) supplemented with 10% fetal bovine serum
(both from Hyclone; GE Healthcare Life Sciences, Logan, Utah, USA),
50 U/ml penicillin and 50 mg/ml streptomycin at 37°C. The cells
were subcultured following partial digestion with 0.25% trypsin and
0.9 mmol/l EDTA in Ca2+-free and Mg2+-free
phosphate-buffered saline (PBS).
Plasmids, transfection and
dual-luciferase reporter assay
miR-7-5p mimic, miR-7-5p inhibitor, mimic negative
control (NC) and inhibitor NC were all purchased from Guangzhou
RiboBio Co., Ltd., (Guangzhou, China). The inhibitor sequences
contained nucleotides with the 2′O methyl ribose modification (m).
The sequences were as follows: miR-7-5p mimic,
5′-UGGAAGACUAGUGAUUUUGUUGU-3′; miR-7-5p mimic NC,
5′-UUUGUACUACACAAAAGUACUG-3′; miR-7-5p inhibitor, 5′-mAm CmA mAm
CmA mAm AmA mUm CmA mCm UmA mGm UmC mUm UmC mCmA-3′; miR-7-5p
inhibitor NC, 5′-mCm AmG mUm AmC mUm UmU mUm GmU mGm UmA mGm UmA
mCm AmA mA-3′. The 3′UTR of TFF3 was obtained from human genomic
DNA (from LS174T cells) using reverse transcription-polymerase
chain reaction (RT-PCR) (42°C 60 min, 70°C 15 min) with the
following primers: Forward, 5′-CCCAAGCTTGGCACCCACGTCACAGGA-3′, and
reverse, 5′-GGACTAGTAACAAAACCCAGGAATAG-3′, respectively. The TFF3
3′UTR was cloned downstream from the luciferase gene in the
pMIR-Reporter luciferase vector (Promega Corporation, Madison, WI,
USA). Mutant vectors with altered predicted miR-7-5p binding sites
were constructed using a site-directed mutagenesis kit (Beyotime
Institute of Biotechnology, Shanghai, China) and the following
primer: 5′-GCAAGCCAATAAAACTGCTGTCCAAAGTGGTCCTTTA-3′. The TFF3 gene
with wild-type 3′UTR (TFF3-WtUTR) or mutant 3′UTR (TFF3-MutUTR) was
inserted into the pMIR-Report luciferase vector. HEK293 cells
(acquired from Shanghai Institute for Biological Sciences, Chinese
Academy of Sciences, Shanghai, China) were seeded at a density of
2×105 cells/well in 24-well plate and cotransfected, respectively
with 200 ng pMIR-TFF3-WtUTR and pMIR-TFF3-MutUTR together with 100
ng pRL-TK-Renilla-luciferase plasmid (Promega Corporation) and 100
nM miR-7-5p mimic or miRNA negative control using Lipofectamine®
3000 (Thermo Fisher Scientific, Inc., Waltham, MA, USA) according
to the manufacturer's protocol. Cells were harvested 48 h post
transfection, and luciferase activity was measured using a
dual-luciferase reporter system (Promega Corporation). The
pRL-TK-Renilla-luciferase plasmid was cotransfected as the internal
Renilla control. Each transfection was performed in triplicate.
In vitro transfection of miR-7-5p
mimic and inhibitor
The effects of miR-7-5p on TFF3 were determined by
western blotting, post-transfection of miR-7-5p into LS174T cells.
The transfection solution was prepared in two microcentrifuge
tubes, one containing DMEM (25 µl) and Lipofectamine® 3000 (5 µl),
the other containing miR-7-5p mimic, miR-7-5p inhibitor, miR-7-5p
mimic NC or miR-7-5p inhibitor NC (10 µl; 100 nM), DMEM (25 µl) and
P3000 (5 µl; Invitrogen; Thermo Fisher Scientific, Inc.). The
contents of the two tubes were mixed together, incubated for 5 min
at room temperature, then added to the LS174T cells, which had been
grown to 60–80% confluence in 6-well plates. Following 48 h culture
at 37°C, the cells were harvested and prepared for the next
experiment.
Western blot analysis
LS174T cells were seeded in 6-well plates and grown
to 60–80% confluence. The cells were transfected with miR-7-5p
mimic, miR-7-5p inhibitor, miR-7-5p mimic NC or miR-7-5p inhibitor
NC for 48 h. Following incubation, the cells were scraped from the
culture plates and lysed with radioimmunoprecipitation assay lysis
buffer [50 mM Tris (pH 7.4), 150 mM NaCl, 1% Triton X-100, 1%
sodium deoxycholate, 0.1% SDS, inhibitors including sodium
orthovanadate, sodium fluoride, EDTA and leupeptin, and
proteinase/phosphatase inhibitors] (Beyotime Institute of
Biotechnology, Shanghai, China). Equal quantities of protein (40
mg) were separated by 15% SDS-PAGE and the proteins were blotted
onto a polyvinylidene fluoride membrane (EMD Millipore, Billerica,
MA, USA). The membrane was blocked in 5% non-fat milk, followed by
incubation at 4°C overnight with primary antibodies against TFF3
(rabbit monoclonal antibody; dilution, 1:1,000; ab108599; Abcam,
Cambridge, MA, USA) and GAPDH (rabbit monoclonal antibody;
dilution, 1:5,000; ab9485; Abcam). After washing in TBS-Tween-20
(0.1%), membranes were incubated with the secondary antibody
[horseradish peroxidase-labeled goat anti-rabbit immunoglobulin G
(H+L); dilution, 1:2,000; A0208; Beyotime Institute of
Biotechnology] at room temperature for 2 h. Bands were visualized
using the enhanced chemiluminescence method (Pierce™ ECL Western
Blotting Substrate; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. Semi-quantification was performed
using ImageJ v1.48u software (National Institutes of Health,
Bethesda, Maryland, USA).
Immunohistochemistry and hematoxylin
and eosin (H&E) staining
Ulcerative colonic tissue specimens (0.2-cm3) were
obtained from children with IBD (5 CD patients and 3 UC patients)
by colonoscopy, and normal colonic tissues were obtained from
patients who had undergone intestinal surgery for other diseases.
Patients were recruited from October-December 2015 in the pediatric
digestive internal medicine ward and the pediatric surgical ward of
Shengjing Hospital of China Medical University (Shenyang, China).
Normal control colon tissues were collected from 8 patients. The
inclusion criteria was as follows: i) Diagnosis with intestinal
polyps or congenital megacolon; ii) pathology revealed normal
intestinal tissue, with no inflammation and ulcers. The exclusion
criteria was as follows: i) Diagnosis with intestinal perforation,
necrosis, or destruction of the mucosal structure; ii) diagnosis
with a tumor, due to the impact on miRNA levels in these patients.
The clinical parameters of the patients were as follows: IBD
patients, age 3.5–14 years; male: Female=7:1; UC: CD=3:5. The age
of the control patients ranged 33 months-10 years; male:
Female=5:3; 7 patients were diagnosed with colonic polyps, 1
patient was diagnosed with congenital megacolon. The present study
was approved by the ethics committee of the Affiliated Shengjing
Hospital of China Medical University (approval no. 2015PS281K;
Shenyang, China). Written informed consent was obtained from the
relatives of the children.
Immunohistochemical staining was performed using a
two-step immunohistochemistry kit (ZSGB-BIO, Beijing, China). The
samples were fixed in 4.0% formaldehyde at room temperature for 24
h, embedded in paraffin and sectioned at 5.0-µm. The sections were
dewaxed (xylene for 10 min, xylene for 20 min), rehydrated [graded
ethanol (100, 95, 85, 70%) for 5 min, respectively], and treated
with 3.0% H2O2 to block endogenous
peroxidases. Antigens were recovered in microwave-heated citrate
acid, and the sections were incubated with primary TFF3 antibody
(dilution, 1:200; ab108599; Abcam) overnight at 4°C. The sections
were recovered at 37°C for 20 min, then incubated with the
secondary antibody (biotin labeled goat anti-rabbit IgG; dilution,
1:50; SP-9001; ZSGB-BIO, Beijing, China) at room temperature for 30
min. The color was developed using 5% diaminobenzidine for 5 min at
room temperature and nuclei were stained with hematoxylin (Beyotime
Institute of Biotechnology) for 10 min at room temperature.
Staining was observed with an Eclipse Ci Plus optical microscope
(Nikon Corporation, Tokyo, Japan). PBS was substituted for the
primary antibody as negative control both in the IBD group and the
NC group.
For H&E staining, samples were washed in
ice-cold PBS, fixed in 4% paraformaldehyde for 24 h at room
temperature and embedded in paraffin. Consecutive colon sections
(4-µm) were stained with 0.1–0.5% H&E at room temperature for 4
min for pathomorphological examination by optical microscope.
RNA extraction and quantitative
(q)-PCR
Ulcerative and normal colonic tissues were obtained
as aforementioned. Total miRNA was extracted using a mirVana™ miRNA
Isolation kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Complementary DNA was synthesized from 10
ng total RNA, using a TaqMan® miRNA reverse transcription kit
(4366596; Thermo Fisher Scientific, Inc.). The reaction mixture
contained the following: 100 nM dNTPs with dTTP, 0.15 µl;
MutiScribe™ Reverse Transcription 50 U/µl, 1 µl; 10X reverse
buffer, 1.5 µl; RNase inhibitor 20 U/µl, 0.19 µl; nuclease-free
water, 4.16 µl; total RNA (1–10 ng), 5 µl; 5X RT primer, 3 µl. The
thermocycling conditions were: 16°C 30 min, 42°C 30 min, 85°C 5
min, hold 4°C. The expression levels of miR-7-5p were quantified
using a TaqMan® small RNA assay kit (PN4427975; Thermo Fisher
Scientific, Inc.). The qPCR reaction mixture contained the
following; TaqMan Universal PCR Master MixII noUNG+
(4440049; Thermo Fisher Scientific, Inc.) 10 µl; nuclease-free
water, 7.67 µl; Taqman Small RNA assay (20X), 1.0 µl; product from
RT reaction, 1.33 µl. The following thermocycling conditions were
employed: Optimal AmpErase UNG activity 50°C 2 min; enzyme
activation 95°C 10 min; 40 cycles of, denaturation at 95°C for 15
sec and extension at 60°C for 60 sec. Quantification was performed
using the 2−ΔΔCq method (12), with U6 snRNA as an endogenous
control.
Statistical analysis
All values were expressed as the mean ± standard
error of the mean. Statistical analyses were performed using SPSS
13.0 (SPSS, Inc., Chicago, IL, USA). Differences between groups
were analyzed by one-way analysis of the variance followed by
Bonferroni post hoc analyses as appropriate. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-7-5p may regulate TFF3 protein
expression
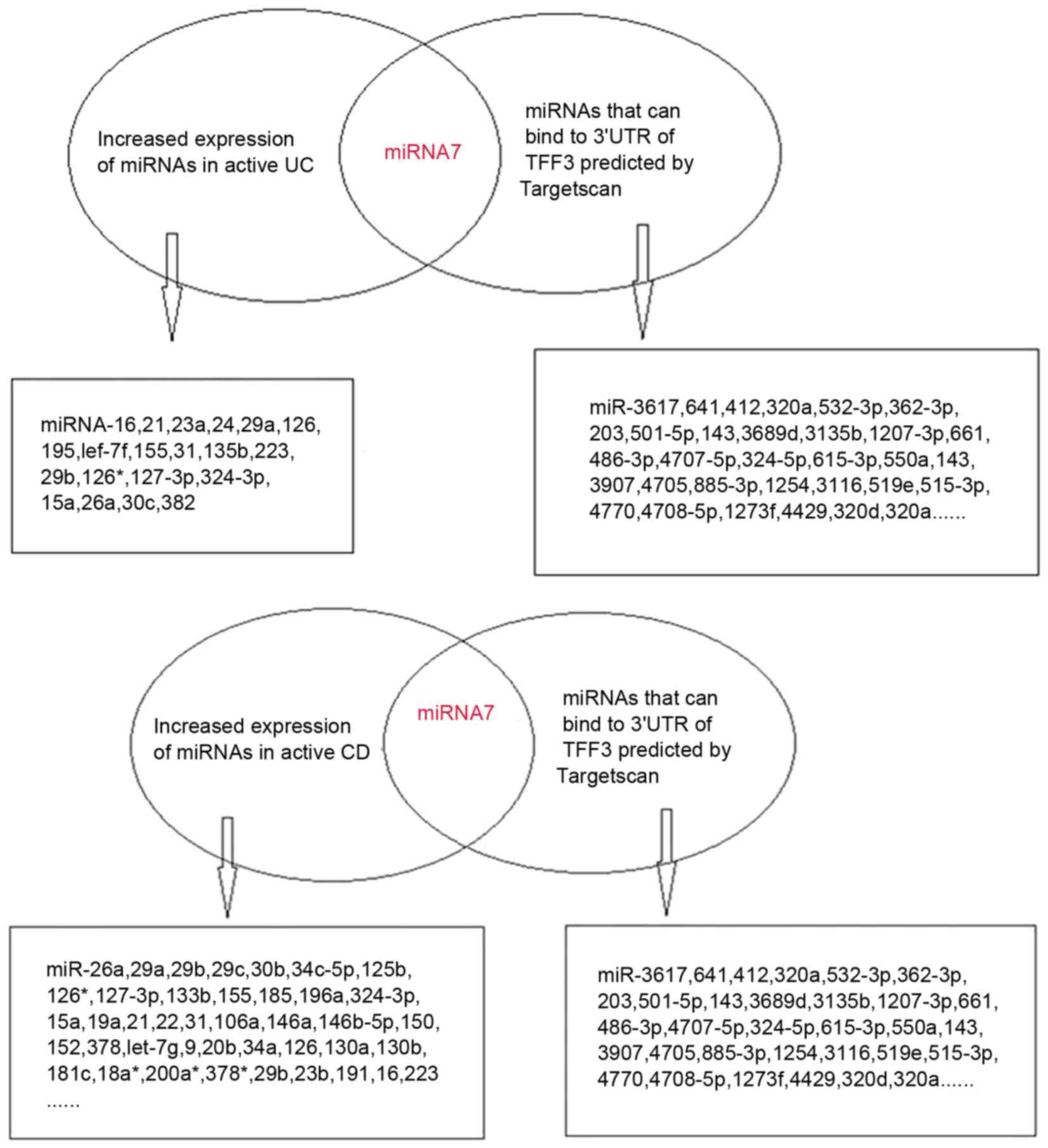
The bioinformatics prediction software Targetscan
v6.2 identified three high-likelihood miRNAs and 123 miRNAs with
relatively lower probabilities, which were able to bind to the
3′UTR of TFF3. Based on previous literature, there were eight
miRNAs that were associated with IBD, including miR-7, miR-320a,
miR-375, miR-532-3p, miR-362-3p, miR-203, miR-501-5p and miR-143
(8,13–16).
Expression of miR-320a was decreased in IBD (8); miR-203, miR-501-5p and miR-143 were
increased in intestinal dysplasia tissues (13–15);
and miR-532-3p and miR-362-3p expression was elevated in the
peripheral blood of patients with CD (16). In addition, miR-7 exhibited
increased expression in active UC and active CD tissues (8).
TargetScan-predicted and literature-identified
miRNAs are presented in Fig. 1.
From these findings, miR-7-5p was hypothesized to be a modulatory
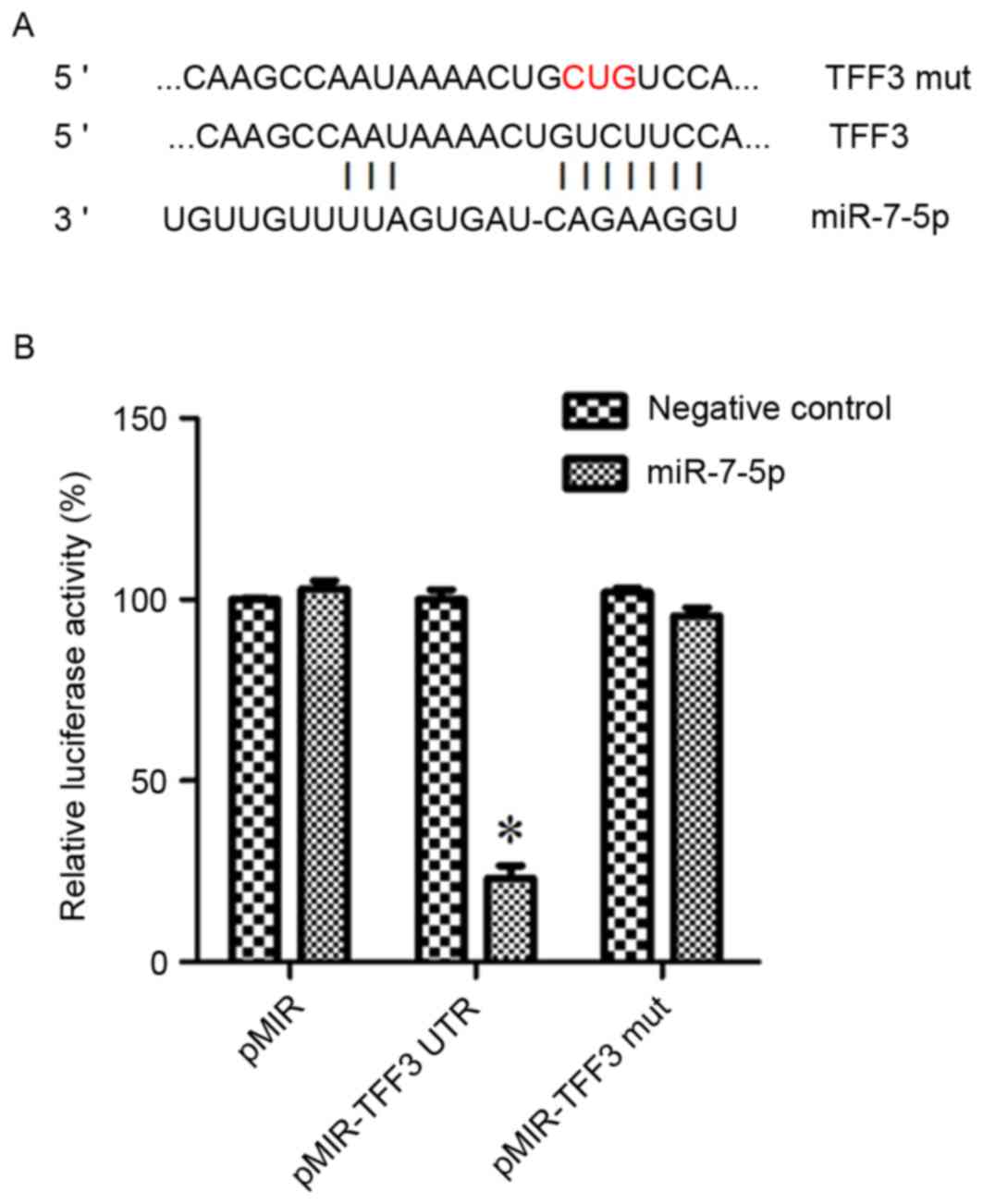
miRNA of the TFF3 protein. Fig. 2A
demonstrates the binding site of miR-7-5p on the 3′UTR of TFF3.
TFF3 is a target of miR-7-5p in HEK293
cells
A dual-luciferase reporter assay was performed to
determine whether TFF3 was a direct target of miR-7-5p in HEK293
cells. The results of the dual-luciferase reporter assays
demonstrated that post-transfection with pMIR-TFF3-WtUTR in
miR-7-5p-transfected cells, there was a significant decrease in
Renilla/firefly luciferase activity; however, there was no
difference among the other groups (Fig. 2B). These findings indicated that
TFF3 may be a target of miR-7-5p.
TFF3 protein expression is negatively
regulated by miR-7-5p in LS174T cells
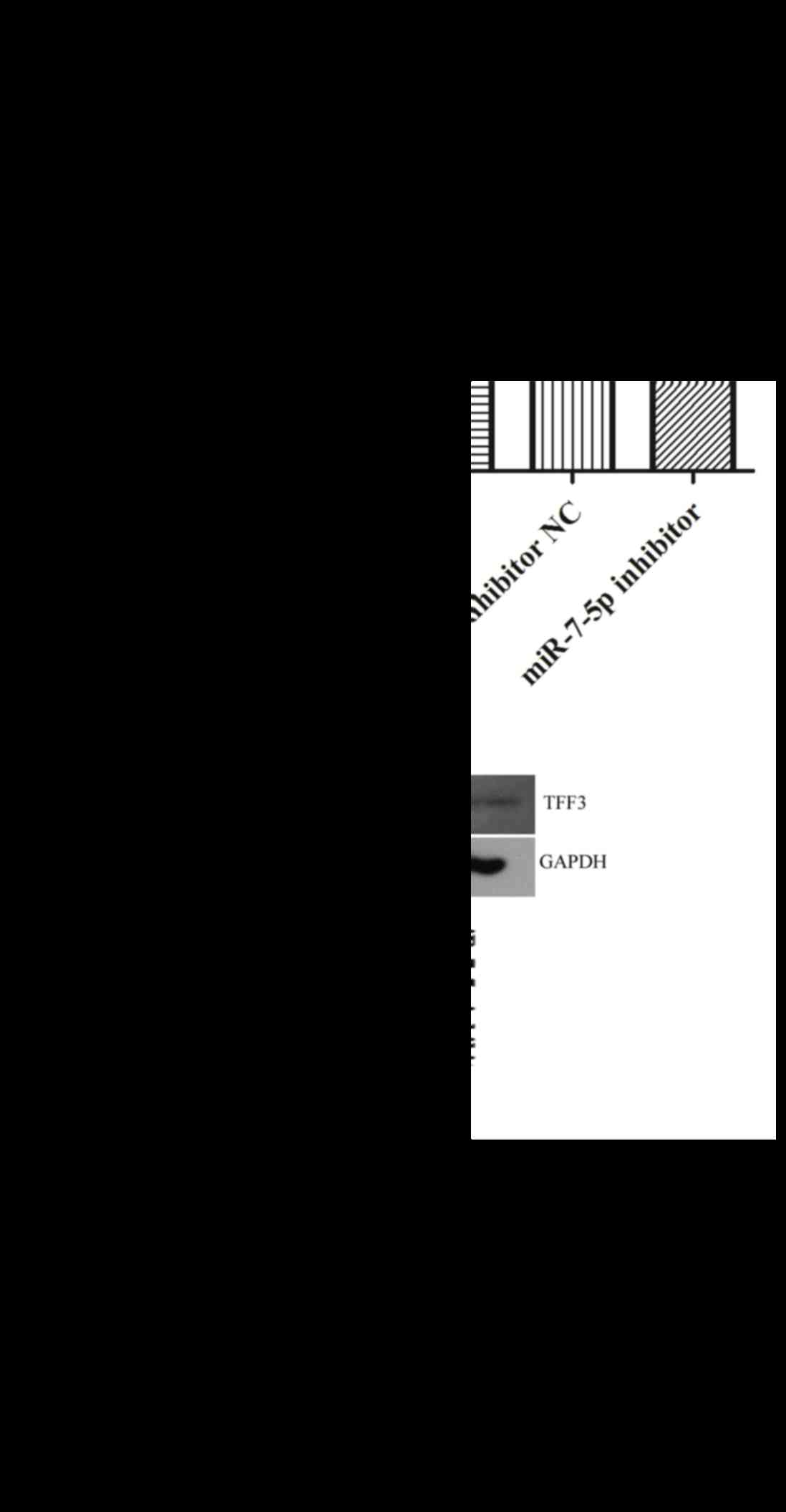
The protein expression levels of TFF3 were evaluated
in LS174T cells where miR-7-5p was overexpressed or suppressed
(Fig. 3). As presented in Fig. 3B, overexpression of miR-7-5p
significantly inhibited the protein expression levels of TFF3.
Conversely, suppression of miR-7-5p expression, via transfection
with the miR-7-5p inhibitor, significantly increased the protein
expression levels of TFF3 in LS174T cells. The relative expression
of miR-7-5p is depicted in Fig.
3A. These findings indicated that miR-7-5p may negatively
regulate TFF3 protein expression at the post-transcriptional
level.
TFF3 expression is decreased in
tissues from children with IBD
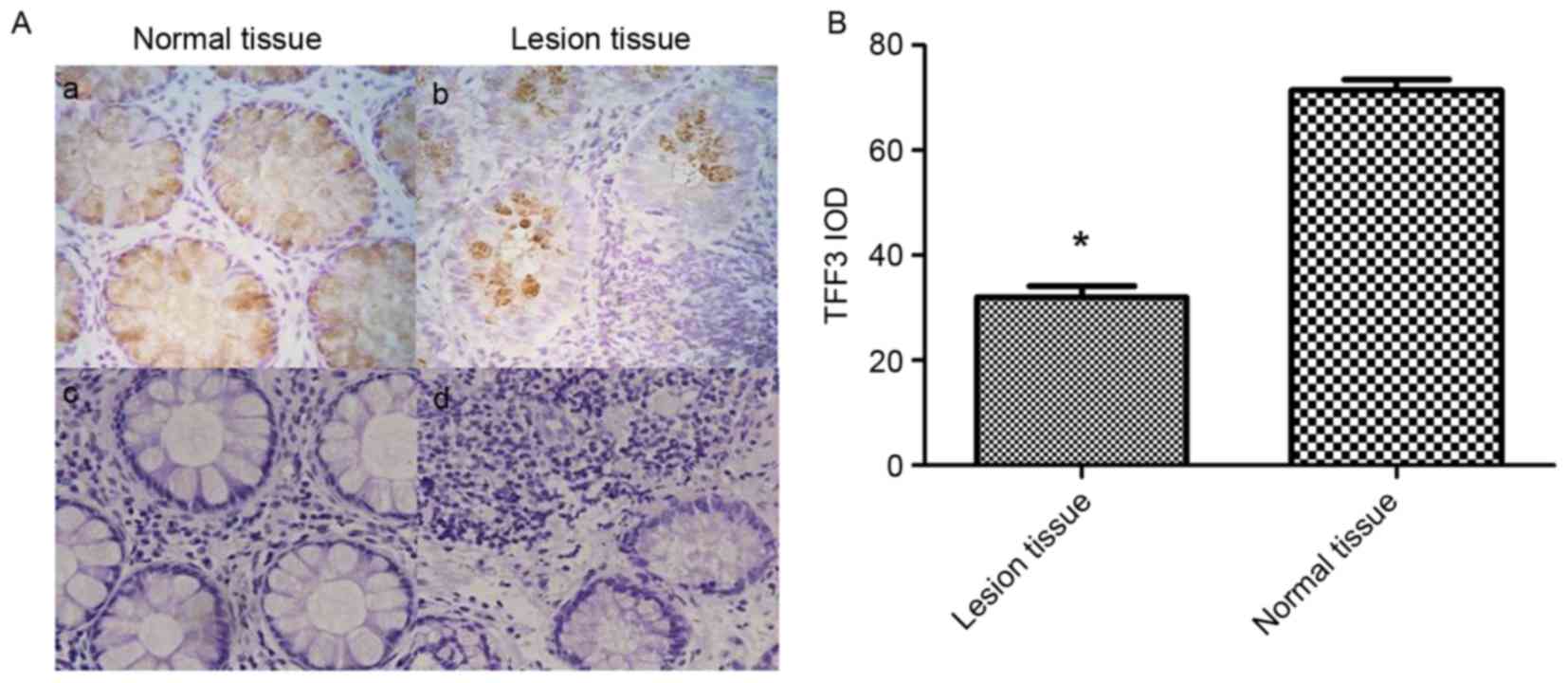
TFF3 expression in IBD tissues from children was
examined by immunohistochemistry (Fig.
4). As presented in Fig. 4Aa and
Ab, when compared to normal colonic tissues, the relative
expression of TFF3 was decreased in IBD lesional tissues. The
integrated optical density of TFF3 staining in normal colonic
tissues was significantly higher compared with in the IBD lesional
tissues (71.5226±4.34 vs. 32.0481±4.76; P<0.05; Fig. 4B). Normal bowel structures were
observed in the H&E stain of normal colon tissue (Fig. 4Ac). In contrast, a large number of
inflammatory cells, irregular intestinal shape and unevenly
distributed goblet cells were present in the H&E stain of IBD
lesional tissue (Fig. 4Ad).
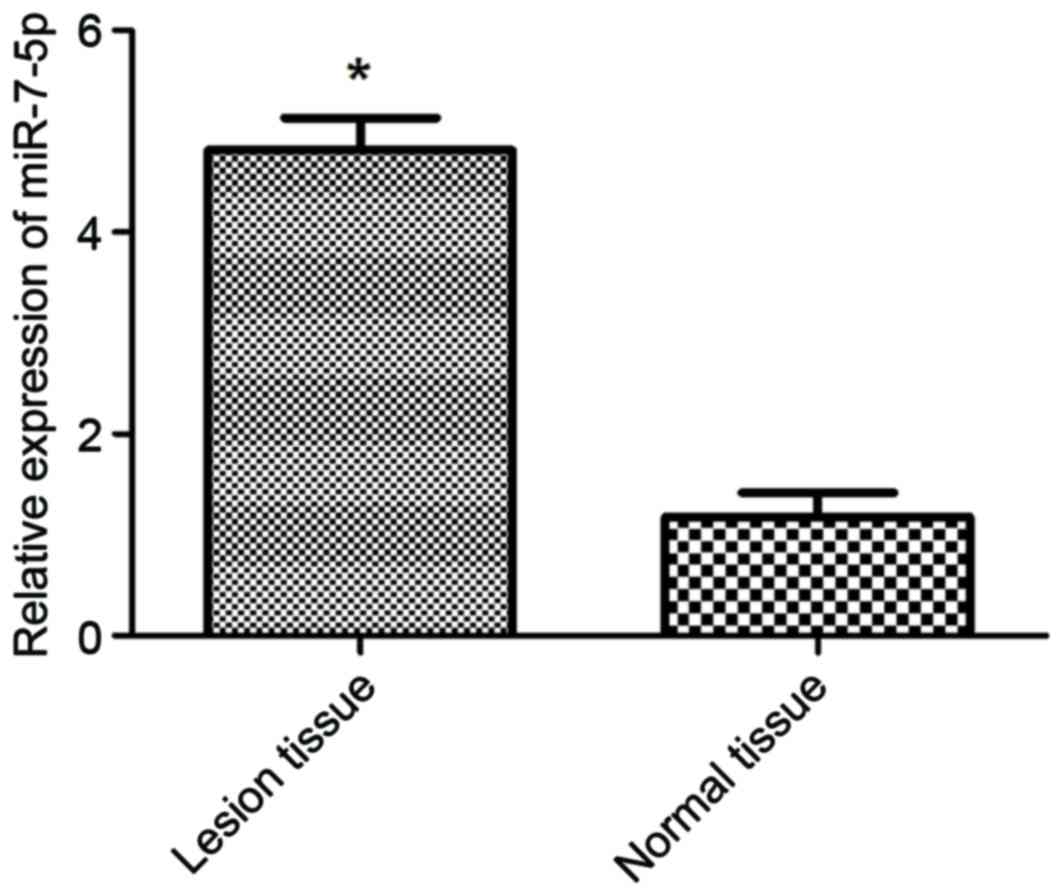
miR-7-5p expression is increased in
tissues from children with IBD
The expression levels of miR-7-5p were detected in
healthy control tissues and IBD lesional tissues (Fig. 5). The expression levels of miR-7-5p
were significantly increased in IBD lesional tissues compared with
in normal colonic tissues (P<0.05). These results indicated that
miR-7-5p expression in IBD lesional tissues was negatively
associated with TFF3 protein expression.
Discussion
The incidence of childhood IBD has been reported to
increase annually. A Scottish study, which was conducted over 40
years, demonstrated that the incidence of IBD in pediatric patients
(<16 years old) increased from 4.45/100,000 between 1990 and
1995, to 7.82/100,000 between 2003 and 2008 (17). In Shanghai, a 10-year study in
patients with IBD reported that the incidence of IBD increased from
0.5 per million in 2001 to 6.0 per million in 2010 (18). Younger patients have a particularly
difficult time dealing with IBD; therefore, it is of great
importance to understand the molecular regulatory mechanisms
contributing to the pathogenesis and treatment of IBD.
TFF3 is a small peptide secreted by goblet cells
that is highly expressed in intestinal mucosa. It can enhance
restitution in intestinal epithelial cells and sustain mucosal
integrity (19,20). Mice overexpressing intestinal TFF3
display increased resistance to intestinal damage and ulceration
(21), whereas mice with TFF3 gene
deletion have a higher susceptibility to gastrointestinal injury
(22). TFF3, in conjunction with
mucin glycoproteins, can protect gastrointestinal mucosa from
various insults (20). Our
previous studies have demonstrated that TFF3 protects the
intestinal epithelium against platelet activating factor-induced
disruption, by restricting the rearrangement of the F-actin
cytoskeleton and tight junctions, thereby decreasing mucosal
permeability (23,24). Given the potential of TFF3 therapy
for treating IBD, the regulatory mechanisms controlling TFF3
expression are of great interest.
Bioinformatics analysis by Targetscan demonstrated
that miR-7-5p can bind to the 3′UTR of TFF3 mRNA. miR-7 is a
well-studied miRNA, and numerous studies have revealed that miR-7
influences cancer cell metastasis, proliferation and apoptosis by
targeting various mRNAs (25–27).
Fang et al (26)
demonstrated that miR-7 inhibited tumor growth and cell metastasis
by targeting the phosphoinositide 3-kinase/Akt pathway in
hepatocellular carcinoma, and Xu et al (27) reported that miR-7 inhibited
colorectal cancer cell proliferation and induced apoptosis by
targeting X-ray repair cross complementing 2. In addition, Nguyen
et al (28) revealed that
miR-7 modulated cluster of differentiation 98 expression during
intestinal epithelial cell differentiation. To the best of our
knowledge, the target of miR-7 in IBD has not been previously
reported. Therefore, the present study evaluated the expression
levels of miR-7-5p in lesional tissues from patients with IBD and
matched normal colonic tissues. The results demonstrated that
miR-7-5p expression was markedly increased in patients with IBD
compared with in control patients. This result was consistent with
a previous study by Pekow et al (29). In addition, Fasseu et al
(8) demonstrated that miR-7 was
overexpressed in inflamed colonic tissues from patients with UC or
CD. The present study also demonstrated that TFF3 expression was
decreased in lesional tissues from patients with IBD compared with
in normal colonic tissue, and was negatively associated with
miR-7-5p expression. The present study identified TFF3 as a target
of miR-7-5p in HEK293 cells, and further demonstrated that miR-7-5p
negatively regulated the expression of TFF3 at a
post-transcriptional level in LS174T cells, indicating that the
effects of TFF3 on IBD may be mediated by miR-7-5p. The functional
implications of the modulation of TFF3 by miR-7-5p require further
study.
In conclusion, the present study demonstrated
differential expression of TFF3 and miR-7-5p in patients with IBD,
and revealed that TFF3 is a novel target of miR-7-5p. In LS174T
cells, miR-7-5p modulated protein expression of TFF3 at a
post-transcriptional level. Further functional studies are required
to fully understand the role of TFF3 in IBD and its regulation by
miR-7-5p.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China Youth Foundation
(grant no. 81400585) and the Natural Science Foundation of Liaoning
Province, China (grant no. 2014021042).
References
|
1
|
Madsen JR, Laursen LS and Lauritsen K:
Chronic inflammatory bowel disease-current status. Ugeskr Laeger.
154:2243–2250. 1992.(In Danish). PubMed/NCBI
|
|
2
|
Ng SC, Zeng Z, Niewiadomski O, Tang W,
Bell S, Kamm MA, Hu P, de Silva HJ, Niriella MA, Udara WS, et al:
Early course of inflammatory bowel disease in a population-based
inception cohort study from 8 countries in Asia and Australia.
Gastroenterology. 150:82–95. e3; quiz e13-e14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kjellev S: The trefoil factor family-small
peptides with multiple functionalities. Cell Mol Life Sci.
66:1350–1369. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stanczyk J, Pedrioli DM, Brentano F,
Sanchez-Pernaute O, Kolling C, Gay RE, Detmar M, Gay S and Kyburz
D: Altered expression of MicroRNA in synovial fibroblasts and
synovial tissue in rheumatoid arthritis. Arthritis Rheum.
58:1001–1009. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tang Y, Luo X, Cui H, Ni X, Yuan M, Guo Y,
Huang X, Zhou H, de Vries N, Tak PP, et al: MicroRNA-146A
contributes to abnormal activation of the type I interferon pathway
in human lupus by targeting the key signaling proteins. Arthritis
Rheum. 60:1065–1075. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tan Z, Randall G, Fan J, Camoretti-Mercado
B, Brockman-Schneider R, Pan L, Solway J, Gern JE, Lemanske RF,
Nicolae D and Ober C: Allele-specific targeting of microRNAs to
HLA-G and risk of asthma. Am J Hum Genet. 81:829–834. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fasseu M, Tréton X, Guichard C, Pedruzzi
E, Cazals-Hatem D, Richard C, Aparicio T, Daniel F, Soulé JC,
Moreau R, et al: Identification of restricted subsets of mature
microRNA abnormally expressed in inactive colonic mucosa of
patients with inflammatory bowel disease. PLoS One. 5:pii:
e131602010. View Article : Google Scholar
|
|
9
|
Sheedy FJ, Palsson-McDermott E, Hennessy
EJ, Martin C, O'Leary JJ, Ruan Q, Johnson DS, Chen Y and O'Neill
LA: Negative regulation of TLR4 via targeting of the
proinflammatory tumor suppressor PDCD4 by the microRNA miR-21. Nat
Immunol. 11:141–147. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
McKenna LB, Schug J, Vourekas A, McKenna
JB, Bramswig NC, Friedman JR and Kaestner KH: MicroRNAs control
intestinal epithelial differentiation, architecture, and barrier
function. Gastroenterology. 139:1654–1664, e1. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ye D, Guo S, Al-Sadi R and Ma TY: MicroRNA
regulation of intestinal epithelial tight junction permeability.
Gastroenterology. 141:1323–1333. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bansal A, Lee IH, Hong X, Anand V, Mathur
SC, Gaddam S, Rastogi A, Wani SB, Gupta N, Visvanathan M, et al:
Feasibility of microRNAs as biomarkers for Barrett's Esophagus
progression: A pilot cross-sectional, phase 2 biomarker study. Am J
Gastroenterol. 106:1055–1063. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Olaru AV, Selaru FM, Mori Y, Vazquez C,
David S, Paun B, Cheng Y, Jin Z, Yang J, Agarwal R, et al: Dynamic
changes in the expression of MicroRNA-31 during inflammatory bowel
disease-associated neoplastic transformation. Inflamm Bowel Dis.
17:221–231. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kanaan Z, Rai SN, Eichenberger MR, Barnes
C, Dworkin AM, Weller C, Cohen E, Roberts H, Keskey B, Petras RE,
et al: Differential microRNA expression tracks neoplastic
progression in inflammatory bowel disease-associated colorectal
cancer. Hum Mutat. 33:551–560. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu F, Guo NJ, Tian H, Marohn M, Gearhart
S, Bayless TM, Brant SR and Kwon JH: Peripheral blood microRNAs
distinguish active ulcerative colitis and Crohn's disease. Inflamm
Bowel Dis. 1:241–250. 2011. View Article : Google Scholar
|
|
17
|
Henderson P and Wilson DC: The rising
incidence of paediatric-onset inflammatory bowel disease. Arch Dis
Child. 97:585–586. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang XQ, Zhang Y, Xu CD, Jiang LR, Huang
Y, Du HM and Wang XJ: Inflammatory bowel disease in Chinese
children: A multicenter analysis over a decade from Shanghai.
Inflamm bowel Dis. 19:423–428. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun Z, Liu H, Yang Z, Shao D, Zhang W, Ren
Y, Sun B, Lin J, Xu M and Nie S: Intestinal trefoil factor
activates the PI3K/Akt signaling pathway to protect gastric mucosal
epithelium from damage. Int J Oncol. 45:1123–1132. 2014.PubMed/NCBI
|
|
20
|
Kindon H, Pothoulakis C, Thim L,
Lynch-Devaney K and Podolsky DK: Trefoil peptide protection of
intestinal epithelial barrier function: Cooperative interaction
with mucin glycoprotein. Gastroenterology. 109:516–523. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Marchbank T, Cox HM, Goodlad RA, Giraud
AS, Moss SF, Poulsom R, Wright NA, Jankowski J and Playford RJ:
Effect of ectopic expression of rat trefoil factor family 3
(intestinal trefoil factor) in the jejunum of transgenic mice. J
Biol Chem. 276:24088–24096. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mashimo H, Wu DC, Podolsky DK and Fishman
MC: Impaired defense of intestinal mucosa in mice lacking
intestinal trefoil factor. Science. 274:262–265. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu LF, Teng X, Guo J and Sun M: Protective
effect of intestinal trefoil factor on injury of intestinal
epithelial tight junction induced by platelet activating factor.
Inflammation. 35:308–315. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu LF, Xu C, Mao ZQ, Teng X, Ma L and Sun
M: Disruption of the F-actin cytoskeleton and monolayer barrier
integrity induced by PAF and the protective effect of ITF on
intestinal epithelium. Arch Pharm Res. 34:245–251. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ma J, Fang B, Zeng F, Pang H, Zhang J, Shi
Y, Wu X, Cheng L, Ma C, Xia J and Wang Z: Curcumin inhibits cell
growth and invasion through up-regulation of miR-7 in pancreatic
cancer cells. Toxicol Lett. 231:82–91. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fang Y, Xue JL, Shen Q, Chen J and Tian L:
MicroRNA-7 inhibits tumor growth and metastasis by targeting the
phosphoinositide 3-kinase/Akt pathway in hepatocellular carcinoma.
Hepatology. 55:1852–1862. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu K, Chen Z, Qin C and Song X: miR-7
inhibits colorectal cancer cell proliferation and induces apoptosis
by targeting XRCC2. Onco Targets Ther. 7:325–332. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nguyen HT, Dalmasso G, Yan Y, Laroui H,
Dahan S, Mayer L, Sitaraman SV and Merlin D: MicroRNA-7 modulates
CD98 expression during intestinal epithelial cell differentiation.
J Biol Chem. 285:1479–1489. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pekow JR and Kwon JH: MicroRNAs in
inflammatory bowel disease. Inflamm Bowel Dis. 18:187–193. 2012.
View Article : Google Scholar : PubMed/NCBI
|