Introduction
In human assisted reproduction technologies (ARTs),
in vitro maturation (IVM) of oocytes had emerged as an
important field, especially when applied to women with a diagnosis
of polycystic ovary syndrome (1,2).
However, embryos produced from IVM oocytes differ greatly from
their in vivo counterparts in multiple aspects, including
developmental competence and subsequent pregnancy rate (3). A lack of the required maturation
factors (growth factors and others) and proper hormonal milieu are
likely to cause incomplete cytoplasmic maturation in IVM.
Additionally, the procedure of IVM involves several steps,
including the isolation, handling and culture of oocytes, which may
exert more environmental stress on oocytes and early embryos
compared with oocytes matured in vivo. Oxidative stress is
one of the most harmful stress factors. In vivo, oocytes and
embryos produce endogenous reactive oxygen species (ROS) that have
an important role as second messengers in cellular functions
through activation of cell signaling cascades (4). However, excessive ROS can lead to
serious consequences, including DNA fragmentation, enzymatic
inactivation and cell death (5).
Progress in embryo developmental competence was obtained by
optimizing culture mediums and IVM protocols. Supplementation of
basic culture medium with serum and gonadotropins, amino acids,
epidermal growth factor, vascular endothelial growth factor,
cysteamine and other factors improved embryo developmental
competence following in vitro fertilization
(IVF)/intracytoplasmic sperm injection (ICSI) of oocytes matured
in vitro.
N-acetyl-5-methoxytryptamine (melatonin), one of the
most effective antioxidants, is a pleiotropic molecule with an
important role in animal reproductive activities (6) and reducing oxidative damage and may
improve the survival environments of cells (7). Melatonin is a derivative of
tryptophan predominantly produced in the pineal gland of
vertebrates and a potent free radical scavenger and antioxidant.
Takasaki et al (8) reported
that oral melatonin can improve the quality of oocytes and may be
useful as a new auxiliary drug for the treatment of infertility.
When melatonin was added to semen extender or culture medium, sperm
viability, oocyte competence and blastocyst development in
vitro were significantly improved (9). Kim et al (10) reported that melatonin
supplementation may be used to improve the clinical outcomes of
typical IVM IVF-embryo transfer. Carr et al (11) demonstrated the long-term safety of
melatonin use. They reported open label follow-up data of 41 cases
of the safety of melatonin use in 50 children. They concluded that
there were no reported late-onset adverse effects.
The majority of the literature supports a
protective/beneficial role for melatonin; however, some controversy
remains. Li et al (12)
demonstrated that exogenous melatonin had no effect on development
of cryopreserved metaphase II oocytes in mice.
To examine the potential beneficial effects of
melatonin, immature oocytes from superovulation cycles were used in
the present study, which was generally referred to as
‘rescue IVM’; these were used instead of immature oocytes
from typical IVM cycles because this was a pilot study and this may
also be applied on human oocytes. Different to immature oocytes in
typical IVM cycles, the immature oocytes in ‘rescue IVM’
were inhibited by the dominant follicle during development.
Normally, these immature oocytes are considered to be of no value
and are discarded. However, several studies have demonstrated their
value (13,14). In the current study, germinal
vesicle (GV) or metaphase I stage (MI) oocytes were incubated with
different concentrations of melatonin (10−11,
10−9, 10−7, 10−5 mol/l).
The advent of commercial microarrays and
high-fidelity RNA amplification techniques have made it possible to
profile gene expression in rarely available human oocytes and
embryos to search for developmental regulators. In the current
study, the transcriptomic profiles of the blastocysts developed
from in vivo matured oocytes and in vitro matured
oocytes with or without melatonin treatment were compared in an
attempt to identify a molecular basis for the effect of melatonin
on oocyte development.
Materials and methods
Experimental patients and ethics
The current study was approved and reviewed by the
Ethics Committee of Anhui Medical University (Hefei, China;
approval no. 2015012). All patients in the study had provided
informed consents for this research. A total of 200 women
undergoing ICSI cycles at the Reproductive Medicine Center of the
First Affiliated Hospital of Anhui Medical University (Hefei,
China) were enrolled from December 2014 to September 2015. The
enrolled patients met the following criteria: i) Patients were
<35 years old; ii) cycles were stimulated with standard long
pituitary downregulation protocol as previously described (15); iii) patients had undergone ICSI
cycles; and iv) no chromosomal abnormalities. No significant
differences were detected in sperm parameters and major clinical
characteristics among the five groups that received varying MT
concentrations (Table I).
 | Table I.Clinical characteristics among groups
treated with varying melatonin concentrations. |
Table I.
Clinical characteristics among groups
treated with varying melatonin concentrations.
| Melatonin treatment
group (mol/l) | Age (years) | Body mass
index | Total Gn dose
(IU) | Gn using days |
|---|
| 0 | 27.0±2.62 | 20.42±1.52 | 1,730±400.10 | 11.2±1.69 |
|
10−5 | 27.3±3.62 | 21.85±3.33 | 1,862±541.98 | 12.1±2.33 |
|
10−7 | 27.5±3.59 | 22.41±2.88 | 2,104±533.11 | 12.4±2.67 |
|
10−9 | 28.4±3.03 | 21.16±2.71 | 1,982±453.28 | 10.9±1.37 |
|
10−11 | 27.6±3.37 | 22.99±2.31 | 2,280±420.49 | 12.8±2.65 |
Human oocyte collection and gamete
manipulation
Controlled ovarian stimulation was achieved using a
standard long pituitary downregulation protocol with
gonadotropin-releasing hormone agonists and recombinant follicle
stimulating hormone (FSH; Gonal-F; Merck KGaA, Darmstadt, Germany)
for treatment of infertility. Follicular development was monitored
by vaginal ultrasound. When at least three follicles reached 18 mm
in diameter, 10,000 IU human chorionic gonadotropin (Livzon
Pharmaceutical Co., Ltd., Shanghai, China) was administered. At ~36
h later, oocyte cumulus complexes (OCC) were collected under
ultrasound guidance. The day prior to oocyte retrieval, serum was
collected from each of the patients. The oocytes were from ICSI
cycles and denuded so as to judge meiotic status as soon as
possible. The immature oocytes at the GV or MI stage were donated
for the research.
Rescue IVM with or without melatonin
treatment
The immature oocytes were placed in pre-equilibrated
in vitro maturation (IVM) medium which was TCM-199
(Sigma-Aldrich; Merck KGaA) medium supplemented with 0.075 IU/ml
human recombinant FSH, 0.1 mg/ml 17β-estradiol, 0.22 mM pyruvic
acid, 0.6 g/l penicillin and streptomycin, 0.5 IU/ml human
chorionic gonadotropin, 20% (v/v) patient serum with different
concentrations of melatonin (0, 10−11, 10−9,
10−7,10−5 mol/l; Sigma-Aldrich; Merck KGaA)
and incubated at 6% CO2, 5% O2 for ~24 h at
37°C. Oocytes with a visible polar body in the perivitelline space
were regarded as matured. The ICSI procedure was carried out with
fresh sperm from different samples. No significant differences were
detected in sperm parameters, which included concentration and
motility among the different groups.
The embryos developed to the blastocyst stage were
vitrified on day 5. The quality of the blastocyst was assessed
based on Gardner's grading system (16).
Blastocyst thawing for total RNA
extraction, amplification, and quality measurement
Blastocysts developed from in vivo maturation
oocytes, and in vitro maturation oocytes with (IVM-anti
group) or without (IVM group) a certain concentration of melatonin
treatment were thawed. Patients that had given birth to healthy
babies donated surplus frozen blastocysts for the research, which
were developed from in vivo maturation oocytes in ICSI
cycles. The three groups of blastocysts, which had similar quality,
were pooled in TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Total RNA was extracted from
each group using the All-Prep DNA/RNA Micro kit (Qiagen, Inc.,
Valencia, CA, USA) according to the manufacturer's instructions.
The purified RNA was then amplified in two rounds and
digoxigenin-labeled according to the NanoAmp™ RT-IVT Labeling kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The quality
and quantity of extracted and amplified RNA were measured using the
NanoDrop ND-1000 spectrophotometer (Thermo Fisher Scientific, Inc.,
Pittsburgh, PA, USA). RNA integrity was assessed by standard
denaturing agarose gel electrophoresis.
RNA labeling and array
hybridization
Sample labeling and array hybridization were
performed according to the Agilent One-Color Microarray-Based Gene
Expression Analysis protocol (Agilent Technologies, Inc., Santa
Clara, CA, USA). Briefly, total RNA from each group was linearly
amplified and labeled with Cy3-UTP. The labeled cRNAs were purified
by RNeasy Mini kit (Qiagen, Inc.). The concentration and specific
activity of the labeled cRNAs (pmol Cy3/µg cRNA) were measured
using the NanoDrop ND-1000. Each labeled cRNA (1 µg) was fragmented
by adding 11 µl 10X blocking agent and 2.2 µl 25X fragmentation
buffer, then heated at 60°C for 30 min, and lastly 55 µl 2X GE
hybridization buffer was added to dilute the labeled cRNA from
Agilent Gene Expression Hybridization kit (Agilent Technologies,
Inc.). Hybridization solution (100 µl) was dispensed into the
gasket slide and assembled to the gene expression microarray slide
(cat no. G4845A; Agilent Technologies, Inc.). The slides were
incubated for 17 h at 65°C in an Agilent Hybridization Oven. The
slides were washed and the arrays were scanned by a G2565BA Agilent
scanner. Three independent repetitions were performed, and a total
of nine chips were used in this study.
Microarray data analysis
Genes in three groups of blastocysts were compared
in pairs. Agilent Feature Extraction software (version 11.0.1.1)
was applied to analyze the acquired array images. Quantile
normalization and subsequent data processing were performed with
using the GeneSpring GX v12.1 software package (Agilent
Technologies, Inc.). Following quantile normalization of the raw
data, genes that at least three out of nine samples had flags in
Detected (‘All Targets Value’) within the analysis software were
selected for further analysis. Data were progressively filtered as
follows: Measurements for each condition with 80% confidence were
removed; less precise measurements based on control strength were
removed; probes recorded as absent in all samples were removed and
the measurements for probes representing the positive and fiducial
controls were removed. Differentially expressed genes with
statistical significance between the two groups were identified
through fold change filtering (fold change ≥2.0; P≤0.05).
Hierarchical clustering was performed using the R scripts (17). Biological functional analysis of
the differentially expressed genes was done using Gene Ontology
(GO; www.geneontology.gov) and the Kyoto
Encyclopedia of Genes and Genomes (KEGG; www.genome.jp/kegg/pathway.html) pathway database.
Validation of important genes derived
from microarray analyses by reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The levels of relevant mRNAs derived from microarray
analyses including the products of certain important genes involved
in a specific pathway, were determined by RT-qPCR using a One-Step
SYBR PrimeScript RT-PCR kit (Takara Bio, Inc., Tokyo, Japan). The
primers used in this study are presented in Table II. Briefly, 2 mg total RNA was
converted to cDNA according to the manufacturer's protocol. PCR was
performed in a total reaction volume of 25 µl, including 10 µl SYBR
Premix Ex Taq (2X), 0.5 µl ROX reference dye II (50X) ×3, 2 µl
cDNA, 8 µl double-distilled water, 1 µl 10 mmol/l PCR forward and
reverse primer. The RT-qPCR was set at an initial denaturation step
of 10 min at 95°C, and 95°C (5 sec), 63°C (30 sec), and 72°C (30
sec) in a total of 40 cycles with a final extension step at 72°C
for 5 min. RT-qPCR was used to quantify the mRNA levels of
methionine adenosyltransferase 2A (MAT2A),
catechol-O-methyltransferase (COMT), period circadian clock 1
(PER1), serine/threonine kinase 4 (STK4), cyclin G1 (CCNG1),
interleukin 4 induced 1 (IL4I1), and forkhead box P3 (FOXP3) with
GAPDH mRNA as the endogenous reference gene using the
2−∆∆Cq method to perform the normalization (18). Melting curve analysis was performed
to confirm the quantitative PCR products. All experiments were
carried out in three repetitions. Statistical comparisons were
performed using the Student's t-test and the SPSS 16.0 statistical
software (P≤0.05 was considered statistically significant).
| Table II.Primer sequences. |
Table II.
Primer sequences.
| Gene | Bidirectional
primer sequences | Size (bp) |
|---|
| GAPDH | F:
5′GGGAAACTGTGGCGTGAT3′ | 299 |
|
| R:
5′GAGTGGGTGTCGCTGTTGA3′ |
|
| MAT2A | F:
5′TGGAGACCAGGGCTTAATGTTTG3′ | 114 |
|
| R:
5′TTACGGCGTAGTTCTGCCAGTTT3′ |
|
| PER1 | F:
5′TACCAGCCATTCCGCCTAAC3′ | 196 |
|
| R: 5′
CAGCCCTTTCATCCACATCC3′ |
|
| STK4 | F:
5′TGAAACTGAAACGCCAGGAAT3′ | 120 |
|
| R:
5′TGCCCATCTCATCACCCACT3′ |
|
| CCNG1 | F:
5′ATGACAAGCCTGAGAAGGTAAA3′ | 142 |
|
| R:
5′TGTGGGAAGACTGATAGTTGATAG3′ |
|
| IL4I1 | F: 5′
CCCTCAAAGACCTCAAGGCACT3′ | 168 |
|
| R: 5′
CCTCGGCGAAGCTGAGATAGA3′ |
|
| COMT | F:
5′GCTGAAGAAGAAGTATGATGTGGA3′ | 220 |
|
| R:
5′CAGGAACGATTGGTAGTGTGTG3′ |
|
| FOXP3 | F:
5′AGGAAAGGAGGATGGACGAA3′ | 124 |
|
| R:
5′GGCAGGCAAGACAGTGGAA3′ |
|
Statistical analysis
Data were expressed as the mean ± standard error,
Statistical comparisons were performed using the Student's t test
or a one-way analysis of variance followed by post-hoc Tukey's
honest significant difference test and the SPSS 16.0 statistical
software (SPSS, Inc., Chicago, IL, USA). P≤0.05 was considered to
indicate a statistically significant difference.
Results
Clinical characteristics of the
patients, effect of melatonin on maturation of human oocyte and
embryo development
A total of 314 GV oocytes and 320 MI oocytes were
collected from 200 patients younger than 35-years-old that had
undergone ICSI cycle, and they were randomly distributed in the
control group (no melatonin) and four other groups of varying MT
concentrations. There was no significant difference in the ratio of
GV/MI in each group. Maturation rate (no. of matured oocytes/total
no. of oocytes), fertilization rate (no. of fertilized oocytes/no.
of matured oocytes), cleavage rate (no. of cleaved oocytes/no. of
fertilized oocytes), blastocyst rate (no. of blastocysts/no. of
cleaved oocytes) and high quality blastocyst rate (no. of high
quality blastocysts/no. of blastocysts) in the five groups are
presented in Table III. The
ratio of high quality blastocysts was significantly higher
(P<0.05) in 10−5 mol/l melatonin-treated group
compared with the others groups. The fertilization rate, cleavage
rate, and the blastocyst rate in 10−5 mol/l
melatonin-treated group were also higher than others groups, but
had no statistically significance. As for maturation rate, there
were also no significant differences among these groups.
| Table III.Embryo development in groups of
varying MT concentrations. |
Table III.
Embryo development in groups of
varying MT concentrations.
| MT treatment group
(mol/l) | Total no. of
oocytes (GV/MI) | Total no. of
matured oocytes maturation rate %) | Total no. of
fertilized oocytes (fertilization rate %) | Total no. of
cleaved oocytes (cleavage rate %) | Total ho. of
blastocysts (Blastocyst rate %) | Total ho. of high
quality blastocyst (high quality blastocyst rate %) |
|---|
| 0 | 135 (70/65) | 101 (76.1±2.2) | 80
(84.8±1.9) | 73 (86.4±2.6) | 14 (23.3±3.5) | 2
(11.1±3.3)a |
|
10−5 | 159 (66/93) | 112 (78.9±2.9) | 101 (94.0±1.5) | 93 (97.9±1.4) | 26 (29.1±3.6) | 14
(54.4±4.7)b |
|
10−7 | 103 (48/55) | 79
(81.1±2.5) | 71
(90.7±2.2) | 64 (82.4±3.6) | 11 (14.0±2.4) | 3
(27.3±4.6)a |
|
10−9 | 117 (64/53) | 93
(79.7±3.2) | 82
(89.7±2.1) | 78 (92.1±2.1) | 14 (16.3±2.8) | 3
(21.1±4.2)a |
|
10−11 | 120 (66/54) | 82
(69.6±3.7) | 68
(85.4±2.4) | 66 (94.2±2.2) | 11 (14.6±2.3) | 2
(15.4±3.2)a |
Comparative gene expression for human
blastocysts from oocytes matured in vitro and in vivo
For each group, [IVM group, IVM-anti group (treated
with 10−5 mol/l melatonin) and in vivo
maturation] 9 blastocysts matured from oocytes were thawed for
total RNA extraction. For each group, 9 blastocysts were assigned
to three PCR tubes filled with 400 µl TRIzol reagent on average, so
as to be replicated three times. In order to clarify a gene prolife
that may better reflect the differences between oocytes matured
in vivo and in vitro, a cut-off of >2-fold
difference was applied. The three groups were compared pairwise. In
blastocysts from the IVM group compared with the in vivo
group, of 15,827 analyzed genes, 412 genes were identified as
differentially expressed (P<0.05), of which 180 were
upregulated, and 232 genes were downregulated. For the IVM-anti
group compared with the IVM group, of 15,827 analyzed genes, 821
genes were identified as differentially expressed (P<0.05), of
which 587 were upregulated, and 234 were downregulated. For the
IVM-anti group compared with the in vivo group, of 15,827
analyzed genes, 1,312 genes are identified as differentially
expressed (P<0.05), of which 877 genes were upregulated, and 435
were downregulated. GenBank accession numbers, false discovery rate
(FDR), regulation direction, biological processes and P-values of
several representative differentially expressed genes are listed in
Table IV. Supervised hierarchical
clustering analysis was performed based on the data of
differentially expressed genes. In clustering maps (Fig. 1), genes from different groups were
clearly distinguished, illustrating that the microarray data
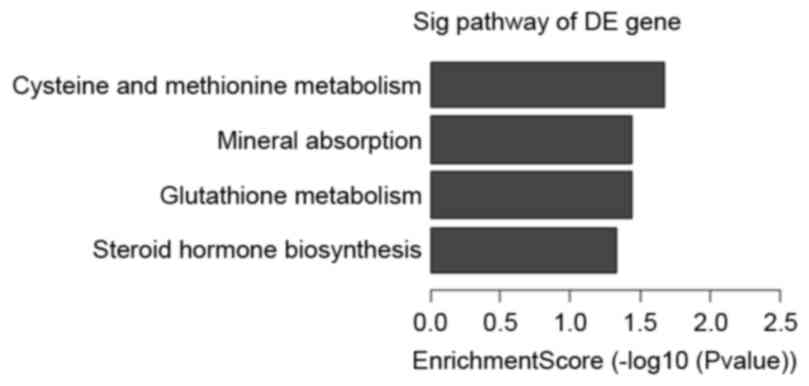
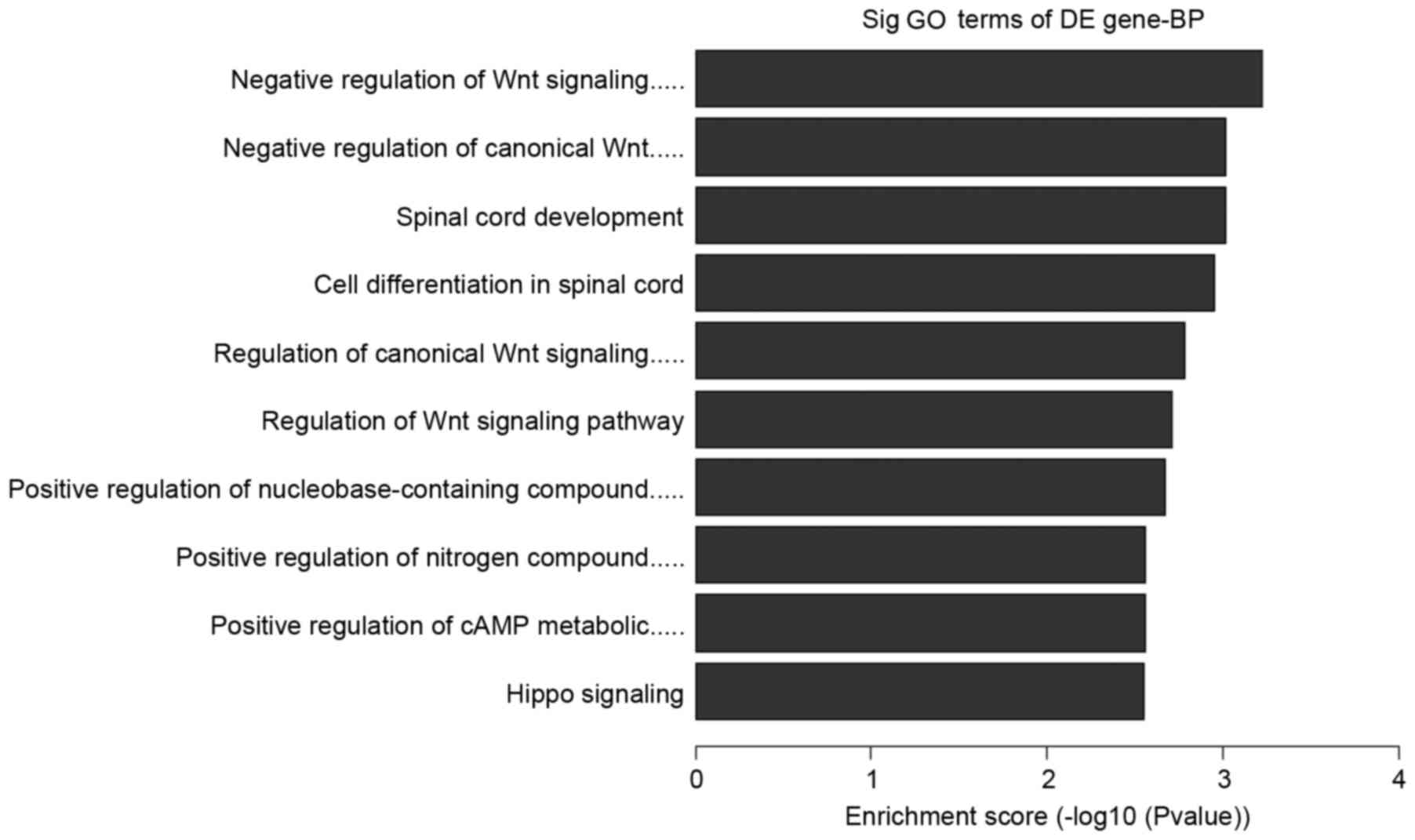
represent the genomic transcripts well. Significantly enriched GO
and KEGG pathways among blastocysts of the three groups are
demonstrated in Tables V–VII. The certain expression changes were
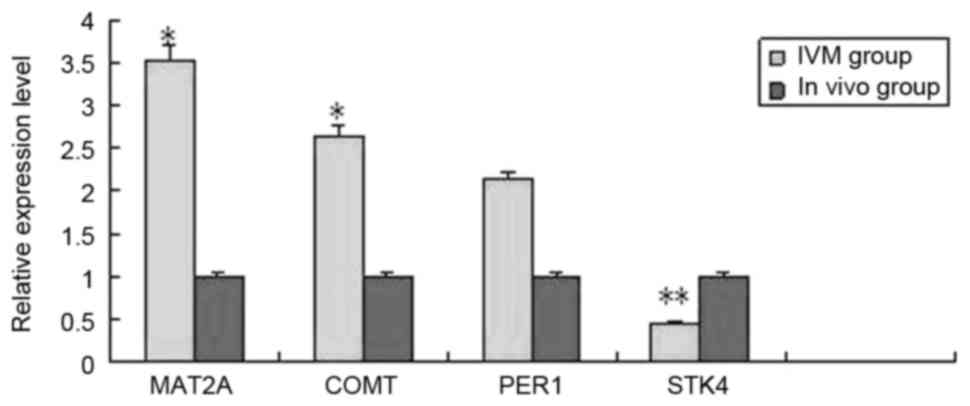
confirmed by RT-qPCR. MAT2A, COMT, PER1, STK4 were analyzed by
RT-qPCR in the IVM group and in vivo group (Fig. 2) MAT2A, COMT and STK4 were
significantly differentially expressed in the RT-qPCR analysis and
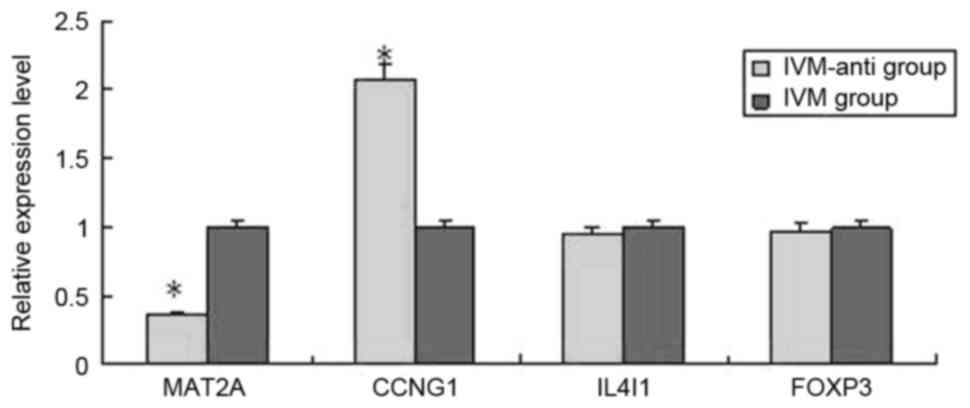
were upregulated in IVM group. CCNG1, IL4I1, MAT2A, FOXP3 were
analyzed by RT-qPCR in IVM-anti group and IVM group (Fig. 3). MAT2A and CCNG1 were
significantly differentially expressed. MAT2A were downregulated
and CCNG1 were upregulated in IVM-anti group.
| Table IV.Differentially expressed genes in the
IVM, IVM-anti and in vivo groups. |
Table IV.
Differentially expressed genes in the
IVM, IVM-anti and in vivo groups.
| A, IVM group vs.
in vivo group |
|---|
|
|---|
| Gene symbol | GenBank
accession | Regulation
direction | FDR | P-value | Biological
processes |
|---|
| MAT2A | NM_005911 | Up | 0.99997 | 0.03405 |
S-adenosylmethionine synthetase |
| COMT | NM_000754 | Up | 0.70388 | 0.00733 |
Catechol-O-methyltransferase |
| ZNF335 | NM_022095 | Up | 0.96627 | 0.01522 | Methylation |
| PER1 | NM_002616 | Up | 0.99997 | 0.01977 | Cation channel
sperm-associated protein 1 |
| FOXO3 | NM_001455 | Up | 0.99997 | 0.02969 | Transcription
factors |
| STK4 | NM_006282 | Down | 0.97854 | 0.01614 |
Phosphorylating |
| NPHP4 | NM_015102 | Down | 0.99997 | 0.03147 | Chromosome and
associated proteins |
|
| B, IVM-anti group
vs. IVM group |
|
| Gene symbol | GenBank
accession | Regulation
direction | FDR | P-value | Biological
processes |
| CCNE1 | NM_001238 | Up | 0.47303 | 0.01166 | G1/S-specific
cyclin E1 |
| CCNG1 | NM_004060 | Up | 0.30861 | 0.00296 | Cyclin G1 |
| MAT2A | NM_005911 | Down | 0.58916 | 0.03081 |
S-adenosylmethionine synthetase |
| IL4I1 | NM_152899 | Down | 0.60742 | 0.04633 | L-amino-acid
oxidase |
| ATP8 | AK128101 | Down | 0.44454 | 0.00934 | Mitochondrial
biogenesis |
|
| C, IVM-anti group
vs. in vivo group |
|
| Gene symbol | GenBank
accession | Regulation
direction | FDR | P-value | Biological
processes |
| DUSP4 | NM_001394 | Up | 0.10966 | 0.00199 | Dual specificity
MAP kinase phosphatase |
| LRG1 | NM_052972 | Up | 0.12045 | 0.00249 | DNA repair and
recombination proteins |
| ATP5J | NM_001003703 | Down | 0.36829 | 0.03604 | ATP synthase |
| COX6B | NM_144613 | Down | 0.13547 | 0.00334 | Cytochrome c
oxidase subunit 6b |
| Table V.Significantly enriched Gene Ontology
and pathways of blastocysts between in IVM group vs. in vivo
group. |
Table V.
Significantly enriched Gene Ontology
and pathways of blastocysts between in IVM group vs. in vivo
group.
| GO/Pathway ID | Regulation
direction | Count | Description | Gene symbols |
|---|
| GO:0015711 | Up | 9 | Organic anion
transport | PQLC2, PRAF2,
SLC38A6, GRM7, SLC26A6, ABCB11, SLC15A4, MFSD10, SCP2 |
| GO:0000096 | Up | 3 | Sulfur amino acid
metabolic process | GCLM, COMT,
MAT2A |
| GO:0034754 | Up | 4 | Cellular hormone
metabolic process | SCP2, HSD17B3,
DHRS9, COMT |
| GO:0032259 | Up | 5 | Methylation | ZNF335, CMTR2,
COMT, MAT2A, PRDM15 |
| GO:0043523 | Up | 4 | Regulation of
neuron apoptotic process | GCLM, LRP1,
BCL2L11, FOXO3 |
| hsa00270 | Up | 2 | Cysteine and
methionine metabolism | MAT2A, SRM |
| hsa00480 | Up | 2 | Glutathione
metabolism | GCLM, SRM |
| hsa04978 | Up | 2 | Mineral
absorption | MT1A, SLC26A6 |
| hsa00140 | Up | 2 | Steroid hormone
biosynthesis | COMT, HSD17B3 |
| GO:0030178 | Down | 6 | Negative regulation
of Wnt signaling pathway | G3BP1, MAD2L2,
NPHP4, SHH, STK4, GNB2L1 |
| GO:0090090 | Down | 5 | Negative regulation
of canonical Wnt signaling pathway | G3BP1, MAD2L2,
NPHP4, SHH, STK4 |
| GO:0035329 | Down | 3 | Hippo
signaling | NPHP4, STK4,
TEAD3 |
| GO:0021510 | Down | 5 | Spinal cord
development | SHH, MDGA2, MDGA1,
EVX1, NEUROG3 |
| Table VII.Significantly enriched Gene Ontology
and pathways of blastocysts in IVM-anti group vs. IVM group. |
Table VII.
Significantly enriched Gene Ontology
and pathways of blastocysts in IVM-anti group vs. IVM group.
| GO/pathway ID | Regulation | Count | Description | Gene symbol |
|---|
| GO:0010942 | Up | 20 | Positive regulation
of cell death | ADIPOQ, BCL2L11,
VAV3, TRIM35, NCSTN, NOTCH2, ADAMTSL4 |
| GO:0043549 | Up | 30 | Regulation of
kinase activity | CDK5R2, CCNE1,
CCNG1, GNAI2, MOS, NTRK3, THBS1, DUSP4 |
| hsa04115 | Up | 5 | p53 signaling
pathway | CCNE1, CCNG1, CD82,
STEAP3, THBS1 |
| hsa00270 | Down | 2 | Cysteine and
methionine metabolism | IL4I1, MAT2A |
| hsa05016 | Down | 5 | Huntington's
disease | ATP8, NDUFV3,
POLR2G, POLR2J, SDHC |
Detailed analysis of associated GO and
KEGG pathways
Further analysis to determine which of the
functional categories assigned by GO and the KEGG pathways were
overrepresented at the 95% confidence level with a minimum overlap
set to two genes. GO analysis and KEGG pathway analysis were
performed using the standard enrichment computation method. The
results are presented in Figs.
4–7.
Discussion
In the majority of superovulation cycles, a small
portion of oocytes remains immature after human chorionic
gonadotropin stimulation. These oocytes are routinely abandoned
because clinical outcomes of these oocytes are flawed (19). In order to make good use of these
oocytes, melatonin was added to the IVM system. The aim of our
study was to evaluate the effect of melatonin in maturation medium
for human ‘rescue IVM’.
As an effective antioxidant, melatonin is known to
be associated with regulation of various dynamic physiological
functions, and also is present in human preovulatory follicular
fluid and may be associated with reproduction. The beneficial
effects of melatonin are concentration dependent. The current study
demonstrated that 10−5 mol/l was the appropriate
concentration of melatonin.
In a previous study, improved maturation and
embryonic development were observed in IVM supplemented with
10−5 mol/l melatonin (10), which is consistent with the results
of the present study. In another study on bovine oocyte maturation
in vitro, the most effective melatonin concentrations ranged
from 10−9-10−7 M; however, as the
concentration of melatonin increased from
10−9-10−3 M, the cleavage rate and the
blastocyst rate declined dramatically (20). This was not surprising as high
concentrations of melatonin inhibit human leukemia cell division
(21).
Consistent with the findings of the current study,
Takasaki et al reported that oral melatonin supplementation
had a beneficial effect on fertilization and embryo quality
(8). In a prospective,
longitudinal, cohort study, treatment with myo-inositol and
melatonin improved ovarian stimulation protocols and pregnancy
outcomes in infertile women with poor oocyte quality (22). Batıoğlu et al (23) demonstrated that melatonin was
likely to improve oocyte and embryo quality in women undergoing IVF
or ICSI (23). Rodriguez-Osorio
et al (24) demonstrated
that melatonin had a positive effect on porcine embryo cleavage
rates and blastocyst cell numbers. Taken together, these data
suggest that melatonin supplementation may have a beneficial effect
on embryo quality. The melatonin in follicular fluid may be
synthesized by the oocytes (25)
and used to protect them from oxidative stress or other
environmental insults by reducing the oxidation of macromolecules
(26). The quality of the oocyte
and the embryo are critical for ART. Poor oocyte or embryo quality
is directly associated with low developmental potential of the
embryo.
Regulation of gene expression has an important role
in embryonic development. Transcripts accumulated throughout the
human oocyte growth phase control development until a
transcriptionally active embryonic genome is established at the 4-
to 8-cell stage of embryo development (27). To further clarify the mechanisms by
which melatonin mediates its beneficial effects on the quality of
embryo, whole gene expression of human blastocysts from in
vivo matured blastocysts, and ‘rescue IVM’ blastocysts
with or without melatonin treatment were profiled. In addition, the
most differentially expressed genes (MAT2A, COMT, PER1, CCNG1), but
also the less differentially expressed genes (PER1, FOXP3 and
IL4I1) were selected for RT-qPCR validation. These genes were
selected out of the numerous differentially expressed genes as they
are involved in the significantly enriched GO and pathways of
blastocysts, which include methylation, cation channel
sperm-associated protein 1, negative regulation of Wnt signaling
pathway, p53 signaling pathway, L-amino-acid oxidase and regulation
of neuron apoptotic process, which have an important role in
regulating embryonic growth.
In vitro and in vivo
The large-scale analysis of the transcriptome
revealed significant differences in mRNA expression levels.
Firstly, a number of the genes that were differentially expressed
when comparing the IVM group and in vivo group (PQ loop
repeat containing 2, PRA1 domain family member 2, solute carrier
family 38 member 6, glutamate metabotropic receptor 7, COMT, MAT2A,
spermidine synthase, metallothionein 1A, STK4 and others) were
involved in organic anion transport, cysteine and methionine
metabolism, methylation, regulation of apoptotic process, mineral
absorption, steroid hormone biosynthesis, Wnt signaling pathway.
These changes likely reflected differences in restructuring of the
cells of the blastocyst during maturation between the in
vitro and in vivo conditions. Many genes that were
involved in stress response and metabolism also varied between
in vitro and in vivo (glutamate-cysteine ligase
modifier subunit, LDL receptor related protein 1, BCL2 like 11 and
forkhead box O3) suggesting sub-optimal oxidative and nutritive
conditions in vitro compared with in vivo.
Cysteine and methionine metabolism pathway genes
were upregulated in the IVM group compared with the vivo
group. Genes, such as MAT2A, involved in this process were
upregulated. Cysteine and methionine metabolism is an epigenetic
pathway associated with DNA methylation that has an important role
in regulating embryonic growth and establishment/maintenance of
genomic imprints (28). DNA
methylation differences between in vitro- and in
vivo-conceived children were previously reported to be
associated with ART procedures rather than infertility (29). The potential for ARTs to adversely
affect the epigenetic status of the developing embryo had been
widely discussed; particularly its potential role in increasing the
relative risk of imprinting defect syndromes, such as
Beckwith-Wiedemann and Angelman syndr`omes (30). Incomplete methylation of maternal
loci during oocyte development may have serious consequences during
development and is of particular concern for IVM.
By contrast, Kuhtz et al (31) demonstrated that human in
vitro oocyte maturation was not associated with increased
imprinting error rates at the KCNQ1 opposite strand/antisense
transcript 1, small nuclear ribonucleoprotein polypeptide N,
paternally expressed 3 and maternally expressed 3 genes (31). Pliushch et al (32) reported that in vitro
maturation of oocytes was not associated with altered
deoxyribonucleic acid methylation patterns in children from in
vitro fertilization or intracytoplasmic sperm injection
(32).
The sample size of microarray analysis in the
present study was relatively small. The MAT2A and COMT genes, which
are associated with DNA methylation, were selected for RT-qPCR
validation comparing expression in the IVM group and in vivo
group; the expression of MAT2A and COMT were significantly
different in these groups. However, whether IVM had an effect on
epigenetics should be clarified in further research.
Melatonin treatment
To the best of our knowledge, establishing a mature
epigenetic status in the genome is part of the maturation process
of the oocyte and is essential for embryo development following
fertilization. Certain reports have suggested that melatonin may
regulate the genome methylation status (33–36).
Li et al (6) reported that
melatonin increased imprinted gene expressions of sirtuin 1, AKT
serine/threonine kinase 2, and DNA polymerase γ 2. In the current
study, MAT2A and COMT expression, which is involved in the pathway
of cysteine and methionine metabolism, were significantly
downregulated in the melatonin-treated group compared with no
melatonin, suggesting that melatonin may regulate epigenetic
status; this may be a factor that mediates the effect of melatonin
on the quality of embryos. When comparing the expression between
the IVM with melatonin treatment group and in vivo group, no
differentially expressed genes involved in DNA methylation were
detected. Thus, melatonin may correct DNA methylation changes
caused by the ‘rescue IVM’ procedure.
In the present study, the p53 signaling pathway was
up regulated in the-melatonin treated group compared with no
melatonin treatment. Differentially expressed genes cyclin E1,
CCNG1, damage specific DNA binding protein 2 and protein
phosphatase, Mg2+/Mn2+ dependent 1D were also
up regulated. p53 is known as the guardian of the genome and has a
major role in DNA repair and apoptosis (36). The cells undergo apoptosis in
response to unrepaired DNA via p53-mediated activation (37). The p53 pathway is induced by a
number of stress signals, including oxidative stress, DNA damage
and activated oncogenes. Over the past three decades researchers
have identified p53 as a multi-functional transcription factor. p53
affects highly diverse cellular processes, and is one of the most
important and extensively studied tumor suppressors (38). p53-regulated gene functions
communicate with adjacent cells, repair damaged DNA, and set up
positive and negative feedback loops that enhance or attenuate the
functions of the p53 protein, integrating these stress responses
with other signal transduction pathways (39). It was previously reported that
apoptosis is suppressed by p53 in post-mitotic spermatogenic cells
to avoid excess death of spermatocytes to guarantee the robustness
of spermatogenesis (40).
Therefore, upregulated p53 signaling pathway may be another
mechanism of action by melatonin.
Blastocyst formation is a crucial stage in early
embryo development. Genes regulate the development and
differentiation of the inner cell mass and trophectoderm of the
embryo, which controls the transition from the undifferentiation to
differentiation stage. The results of the current study also
demonstrated that negative regulation of canonical Wnt signaling
pathway was downregulated in the IVM group without melatonin
treatment compared with the in vivo group [differentially
expressed genes were G3BP stress granule assembly factor 1, MAD2L2,
nephrocystin 4, sonic hedgehog, STK4, receptor for activated C
kinase 1], but no difference in the Wnt signaling pathway were
detected between the IVM with melatonin treatment and the in
vivo group. Wnt signaling was first identified for its role in
carcinogenesis, but had since been recognized for its function in
embryonic development range from cell fate determination, cell
cycle, body axis patterning to cell fate specification, cell
proliferation, and cell migration (41). Wnts were highly evolutionarily
conserved in animals (42).
Melatonin may also act via effects on Wnt signaling.
NET working
The mechanisms of network of interaction between
various signaling pathways had been investigated. Several authors
have reported that apoptotic activation of p53 is mediated by
activation of extracellular signal-regulated kinase, p38
mitogen-activated protein kinase and c-Jun N-terminal kinase
pathways (43). Other studies have
determined the effects of microRNAs (miRNAs). miRNAs are a group of
noncoding regulatory RNAs known to influence the stability and
translational efficiency of target mRNAs. Kim et al
(44) reported that p53 and
miRNA-34 re suppressors of the canonical Wnt signaling pathway;
their data provided insight into the mechanisms by which a
p53-miRNA-34 network restrained canonical Wnt signaling cascades in
developing organisms and human cancer. In another study, miRNAs
were reported to be associated with human embryo implantation
defects (45). Melatonin was
demonstrated to exert its biological functions by modulation of
miRNA expression in human breast cancer cells (46). Thus, melatonin may affect miRNAs,
which may regulate the network of interaction between the p53 and
Wnt signaling pathways in in vitro maturation of
oocytes.
In summary, gene expression profiling using whole
human genome arrays and subsequent data analysis provided a
molecular basis for the relative higher quality of IVM blastocysts
treated with melatonin compared with those without melatonin
treatment. Certain genes identified in the current study, including
MAT2A, COMT, CCNG1, and STK4, may be useful as the biomarkers of
IVM safety. The IVM procedure may potentially affect gametes and
embryos by causing disorders of DNA methylation patterns and the
canonical Wnt signaling pathway. Exogenous melatonin treatment
positively influenced the quality of blastocysts, and regulated the
p53 signaling pathway and genes associated with DNA methylation
during ‘rescue IVM’. However, the current study was limited
due to the suboptimal source of the oocytes and limited sample
size. This study reflects what is generally referred to as
‘rescue IVM’ and was not a true reflection of clinical IVM
techniques. Therefore, as a promising IVM supplement, the role of
melatonin requires further investigation.
Acknowledgements
The study has been supported by the Cultivating
Youth Training Program (2015KJ03) in The First Affiliated Hospital
of Anhui Medical University.
References
|
1
|
Fadini R, Dal Canto MB, Renzini MM,
Brambillasca F, Comi R, Fumagalli D, Lain M and De Ponti E:
Predictive factors in in-vitro maturation in unstimulated women
with normal ovaries. Reprod Biomed Online. 18:251–261. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Smitz JE, Thompson JG and Gilchrist RB:
The promise of in vitro maturation in assisted reproduction and
fertility preservation. Semin Reprod Med. 29:24–37. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nogueira D, Sadeu JC and Montagut J: In
vitro oocyte maturation: Current status. Semin Reprod Med.
30:199–213. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Harvey AJ, Kind KL and Thompson JG: REDOX
regulation of early embryo development. Reproduction. 123:479–486.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Reshi ML, Su YC and Hong JR: RNA viruses:
ROS-mediated cell death. Int J Cell Bio. 2014:4674522014.
|
|
6
|
Li Y, Zhang Z, He C, Zhu K, Xu Z, Ma T,
Tao J and Liu G: Melatonin protects porcine oocyte in vitro
maturation from heat stress. J Pineal Res. 59:365–375. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mukherjee D, Roy SG, Bandyopadhyay A,
Chattopadhyay A, Basu A, Mitra E, Ghosh AK, Reiter RJ and
Bandyopadhyay D: Melatonin protects against isoproterenol-induced
myocardial injury in the rat: Antioxidative mechanisms. J Pineal
Res. 48:251–262. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takasaki A, Nakamura Y, Tamura H,
Shimamure K and Morioka H: Melatonin as a new drug for improving
oocyte quality. Reprod Med Biol. 2:139–144. 2003. View Article : Google Scholar
|
|
9
|
Cruz MH, Leal CL, da Cruz JF, Tan DX and
Reiter RJ: Role of melatonin on production and preservation of
gametes and embryos: A brief review. Anim Reprod Sci. 145:150–160.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim MK, Park EA, Kim HJ, Choi WY, Cho JH,
Lee WS, Cha KY, Kim YS, Lee DR and Yoon TK: Does supplementation of
in-vitro culture medium with melatonin improve IVF outcome in PCOS?
Reprod Biomed Online. 26:22–29. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Carr R, Wasdell MB, Hamilton D, Weiss MD,
Freeman RD, Tai J, Rietveld WJ and Jan JE: Long-term effectiveness
outcome of melatonin therapy in children with treatment-resistant
circadian rhythm sleep disorders. J Pineal Res. 43:351–359. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li W, Cheng K, Zhang Y, Meng Q, Zhu S and
Zhou G: No effect of exogenous melatonin on development of
cryopreserved metaphase II oocytes in mouse. J Anim Sci Biotechnol.
6:422015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Isachenko EF and Nayudu PL: Vitrification
of mouse germinal vesicle oocytes: Effect of treatment temperature
and egg yolk on chromosomal normality and cumulus integrity. Hum
Reprod. 14:400–408. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mandelbaum J, Belaїsch-Allart J, Junca AM,
Antoine JM, Plachot M, Alvarez S, Alnot MO and Salat-Baroux J:
Cryopreservation in human assisted reproduction is now routine for
embryos but remains a research procedure for oocytes. Hum Reprod.
13:161–177. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Y, Yang D and Zhang Q: Clinical outcome
of one-third-dose depot triptorelin is the same as half-dose depot
triptorelin in the long protocol of controlled ovarian stimulation.
J Hum Reprod Sci. 5:14–19. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gardner DK, Lane M, Stevens J, Schlenker T
and Schoolcraft WB: Blastocyst score affects implantation and
pregnancy outcome: Towards a single blastocyst transfer. Fertil
Steril. 73:1155–1158. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
R Development Core Team R, . A language
and Environment for Statistical Computing. R Foundation for
Statistical Computing; Vienna: 2008
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tamura H, Takasaki A, Taketani T, Tanabe
M, Kizuka F, Lee L, Tamura I, Maekawa R, Aasada H, Yamagata Y and
Sugino N: The role of melatonin as an antioxidant in the follicle.
J Ovarian Res. 5:52012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tian X, Wang F, He C, Zhang L, Tan D,
Reiter RJ, Xu J, Ji P and Liu G: Beneficial effects of melatonin on
bovine oocytes maturation: A mechanistic approach. J Pineal Res.
57:239–247. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Büyükavci M, Ozdemir O, Buck S, Stout M,
Ravindranath Y and Savaşan S: Melatonin cytotoxicity in human
leukemia cells: Relation with its pro-oxidant effect. Fundam Clin
Pharmacol. 20:73–79. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Unfer V, Raffone E, Rizzo P and Buffo S:
Effect of a supplementation with myo-inositol plus melatonin on
oocyte quality in women who failed to conceive in previous in vitro
fertilization cycles for poor oocyte quality: A prospective,
longitudinal, cohort study. Gynecol Endocrino. 27:857–861. 2011.
View Article : Google Scholar
|
|
23
|
Batıoğlu AS, Sahin U, Gürlek B, Oztürk N
and Unsal E: The efficacy of melatonin administration on oocyte
quality. Gynecol Endocrinol. 28:91–93. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rodriguez-Osorio N, Kim IJ, Wang H, Kaya A
and Memili E: Melatonin increases cleavage rate of porcine
preimplantation embryos in vitro. J Pineal Res. 43:283–288. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sakaguchi K, Itoh MT, Takahashi N, Tarumi
W and Ishizuka B: The rat oocyte synthesises melatonin. Reprod
Fertil Dev. 25:674–682. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gao C, Han HB, Tian XZ, Tan DX, Wang L,
Zhou GB, Zhu SE and Liu GS: Melatonin promotes embryonic
development and reduces reactive oxygen species in vitrified mouse
2-cell embryos. J Pineal Res. 52:305–311. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wynn P, Picton HM, Krapez JA, Rutherford
AJ, Balen AH and Gosden RG: Pretreatment with follicle stimulating
hormone promotes the numbers of human oocytes reaching metaphase II
by in vitro maturation. Hum Reprod. 13:3132–3138. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chiba H, Hiura H, Okae H, Miyauchi N, Sato
F, Sato A and Arima T: DNA methylation errors in imprinting
disorders and assisted reproductive technology. Pediatr Int.
55:542–549. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Song S, Ghosh J, Mainigi M, Turan N,
Weinerman R, Truongcao M, Coutifaris C and Sapienza C: DNA
methylation differences between in vitro- and in vivo-conceived
children are associated with ART procedures rather than
infertility. Clin Epigenetics. 7:412015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Maher ER, Afnan M and Barratt CL:
Epigenetic risks related to assisted reproductive technologies:
Epigenetics, imprinting, ART and icebergs? Hum Reprod.
18:2508–2511. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kuhtz J, Romero S, de Vos M, Smitz J, Haaf
T and Anckaert E: Human in vitro oocyte maturation is not
associated with increased imprinting error rates at LIT1, SNRPN,
PEG3 and GTL2. Hum Reprod. 29:1995–2005. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pliushch G, Schneider E, Schneider T, El
Hajj N, Rösner S, Strowitzki T and Haaf T: In vitro maturation of
oocytes is not associated with altered deoxyribonucleic acid
methylation patterns in children from in vitro fertilization or
intracytoplasmic sperm injection. Fertil Steril. 103:720–727.e1.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schwimmer H, Metzer A, Pilosof Y, Szyf M,
Machnes ZM, Fares F, Harel O and Haim A: Light at night and
melatonin have opposite effects on breast cancer tumors in mice
assessed by growth rates and global DNA methylation. Chronobiol
Int. 31:144–150. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee SE, Kim SJ, Yoon HJ, Yu SY, Yang H,
Jeong SI, Hwang SY, Park CS and Park YS: Genome-wide profiling in
melatonin-exposed human breast cancer cell lines identifies
differentially methylated genes involved in the anticancer effect
of melatonin. J Pineal Res. 54:80–88. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Korkmaz A and Reiter RJ: Epigenetic
regulation: A new research area for melatonin? J Pineal Res.
44:41–44. 2008.PubMed/NCBI
|
|
36
|
Woods DB and Vousden KH: Regulation of p53
function. Exp Cell Res. 264:56–66. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Appella E and Anderson CW:
Post-translational modifications and activation of p53 by genotoxic
stresses. Eur J Biochem. 268:2764–2772. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Stegh AH: Targeting the p53 signaling
pathway in cancer therapy-the promises, challenges and perils.
Expert Opin Ther Targets. 16:67–83. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Levine AJ, Hu W and Feng Z: The P53
pathway: What questions remain to be explored? Cell Death Differ.
13:1027–1036. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen D, Zheng W, Lin A, Uyhazi K, Zhao H
and Lin H: Pumilio 1 suppresses multiple activators of p53 to
safeguard spermatogenesis. Curr Biol. 22:420–425. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Petersen CP and Reddien PW: Wnt signaling
and the polarity of the primary body axis. Cell. 139:1056–1068.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nusse R and Varmus HE: Wnt genes. Cell.
69:1073–1087. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lin T, Mak NK and Yang MS: MAPK regulate
p53-dependent cell death induced by benzo(a)pyrene: Involvement of
p53 phosphorylation and acetylation. Toxicology. 247:145–153. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kim NH, Kim HS, Kim NG, Lee I, Choi HS, Li
XY, Kang SE, Cha SY, Ryu JK, Na JM, et al: p53 and miRNA-34 are
suppressors of canonical Wnt signaling. Sci Signal. 4:ra712012.
|
|
45
|
Revel A, Achache H, Stevens J, Smith Y and
Reich R: MicroRNAs are associated with human embryo implantation
defects. Hum Reprod. 26:2830–2840. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lee SE, Kim SJ, Youn JP, Hwang SY, Park CS
and Park YS: MicroRNA and gene expression analysis of
melatonin-exposed human breast cancer cell lines indicating
involvement of the anticancer effect. J Pineal Res. 51:345–352.
2011. View Article : Google Scholar : PubMed/NCBI
|