Introduction
Persistent pulmonary hypertension of the newborn
(PPHN) is a common, yet deleterious condition that is observed in
the neonatal intensive care unit and has high mortality and
variable morbidity rates (1). The
etiology of PPHN is complex; however, several studies (2,3) have
reported that the main causes of this condition include hypoxia and
constriction of the fetal ductus arteriosus in utero, which
may occur following exposure to non-steroidal anti-inflammatory
drugs (NSAIDs) during the third trimester (4). PPHN is pathologically characterized
by pulmonary vascular remodeling (5), which comprises smooth muscle cell
proliferation and adventitial hypertrophy (6); pulmonary artery smooth muscle cell
(PASMC) proliferation is a major contributor to pulmonary vascular
remodeling (7).
In PASMCs, a rise in cytosolic Ca2+
concentration is considered to be a key stimulus for proliferation
(8,9). A previous study has demonstrated that
of the three main extracellular calcium influx pathways,
voltage-operated calcium entry, receptor-operated calcium entry and
store-operated calcium entry (SOCE), hypoxia was able to enhance
SOCE through store-operated calcium channels (SOCCs) (10). Furthermore, transient receptor
potential cation channels (TRPCs) are the main components of SOCCs
(11). In adult animal models of
hypoxia-induced pulmonary hypertension (PH), high expression levels
of canonical TRPC, subfamily C, member 1 (TRPC1) and TRPC6 were
revealed to contribute to excessive pulmonary vascular remodeling
(11). Whether the elevated
expression of TRPC1 and TRPC6 results in increased PASMC
proliferation and contributes to pulmonary vascular remodeling in
PPHN model rats is unknown.
Peroxisome proliferator-activated receptor γ
(PPAR-γ) is a member of the nuclear receptor hormone superfamily
(12,13) and is considered to serve a vital
role in cell differentiation, development, metabolism, inflammation
and tumorigenesis (13). It is
widely expressed in pulmonary vascular endothelial and smooth
muscle cells in normal human lungs and regulates vascular smooth
muscle cell proliferation (14). A
number of studies have indicated that a decrease of PPAR-γ
expression may contribute to the development of PH (10,15).
In addition, mice with selective deletion of PPAR-γ in PASMCs were
reported to spontaneously develop PH and exhibited increased
muscularization of the distal pulmonary arteries (16). In addition, PPAR-γ deletion was
revealed to induce human PASMC (HPASMC) proliferation in
vitro and PPAR-γ overexpression reduced HPASMC proliferation
in vitro (17). In adult
rat models of PH, treatment with PPAR-γ agonists were demonstrated
to protect against vascular remodeling and PH (18).
Decreased PPAR-γ expression may contribute to PASMC
proliferation and vascular remodeling in adult models of PH.
Whether changes to the level of expression of PPAR-γ contributes to
PPHN in newborn rats remains unclear. Thus, the aim of the present
study was to establish a PPHN rat model and to investigate whether
the PPAR-γ/TRPC pathway is altered in PPHN.
Materials and methods
Animal models
All animal procedures and protocols were approved by
the Committee on the Ethics of Animal Experiments at China Medical
University (Shenyang, China). All surgeries were performed under
chloral hydrate anesthesia, and all efforts were made to minimize
animal suffering.
A total of 10 specific pathogen free healthy female
Sprague-Dawley (SD) rats (weight, 250–300 g; age, 7–8 weeks old)
were provided by the Experimental Animal Center of Shengjing
Hospital of China Medical University (Shenyang, China). All rats
had free access to food and water, and were housed at 25–27°C and
50–70% humidity with a 12-h light/dark cycle. The female rats were
mated overnight; this was considered day 0 of gestation. A
combination of hypoxic conditions and treatment with the NSAID
indomethacin was used to establish a rat model of PPHN, as
previously described (19)
Briefly, on gestation day 19, pregnant rats were randomly assigned
to either the PPHN model or the Control group (n=5/group). Firstly,
indomethacin (0.5 mg) was dissolved in 10 ml of 38% alcohol
(sterile distilled water and ethanol; cat. no. I7378;
Sigma-Aldrich, Merck KGaA, Darmstadt, Germany). During days 19 to
21 of gestation, pregnant rats in the PPHN group were subjected to
hypoxic conditions (oxygen concentration, 10±0.5%) and treated with
indomethacin (0.5 mg/kg) by intraperitoneal injection twice a day.
However, during days 19 to 21 of gestation the pregnant rats in the
Control group were housed under standard normoxic conditions
(oxygen concentration, 21%) and treated with isotonic saline by
intraperitoneal injection twice a day. On day 22, the fetuses were
born by cesarean section from both groups. In both groups, 50
fetuses were randomly selected, sacrificed and samples were
collected as described below.
Preparation of the lungs and
hearts
Lungs and hearts were isolated from the newborn rats
and prepared for further examination. The inferior lobe of the
right lung was fixed in 4% paraformaldehyde for 24 h at 4°C,
embedded in paraffin and sectioned (4–5 µm) prior to staining with
hematoxylin and eosin, or immunohistochemical analysis. The
remaining lung tissue was stored at −70–80°C for western blot
analysis and mRNA detection. Following collection of fetal hearts,
the left atrium, right atrium and free large vessels were removed.
In addition, the right ventricle (RV) free wall was removed from
the left ventricle (LV) and septum (SV) under a stereomicroscope.
Then the weight of the RV, LV and septum were measured. The RV
hypertrophy index (RVHI) was calculated to evaluate the PPHN model,
which was assessed by the ratio of the weight of the RV to that of
the LV plus SV: RVHI = RV/(LV + SV).
Hematoxylin and eosin (H&E)
staining, immunohistochemistry and measurement of vascular
remodeling
The paraffin-embedded sections were stained with
H&E and immunohistochemical staining. Briefly, sections of lung
(4–5 µm thick) were deparaffinized in graded alcohol solutions and
xylene, and stained with H&E using a H&E Staining kit (cat.
no. G1120; Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China). Immunohistochemical analysis was performed to
identify the α smooth muscle actin (α-SMA) content present in the
paraffin-embedded lung tissue samples. The α-SMA antibody can be
used for marking vascular smooth muscle, thus, in the present
study, medial thickness and medial area occupied by smooth muscle
was determined by positive α-SMA staining; circular areas of
positive α-SMA staining can also indirectly represent the smooth
muscle of the pulmonary artery or distal pulmonary arterioles
(20,21).
Briefly, sections of lung (4–5 µm thick) were
deparaffinized in graded alcohol solutions and xylene. Following
the manufacturer's instructions (immunohistochemistry kit; cat. no.
SP-9002; Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd.;
OriGene Technologies, Inc., Beijing, China), all sections were then
blocked with 3% H2O2 (37°C, 20 min) and goat
serum (40 min at room temperature). Subsequently, sections were
incubated overnight at 4°C with the α-SMA primary antibody (1:100;
mouse anti-rat α-SMA; cat. no. ab7817; Abcam, Cambridge, UK). The
negative control tissues were incubated with phosphate-buffered
saline instead of the primary antibody. The next day, sections were
incubated with the biotin-labeled goat anti-mouse IgG secondary
antibody (20 min at 37°C) and then incubated with a horseradish
peroxidase marker (20 min at 37°C). The paraffin sections were
developed with 3′-diaminobenzidine and counterstained with 10%
hematoxylin for 3 min at room temperature; they were then
dehydrated in a gradient of alcohol, treated with xylene and
mounted using neutral balsam. A light microscope was used for image
acquisition, and the deposition of brown particles indicated a
positive result. A total of 3 paraffin-embedded sections were
randomly selected from 10 random rats/group and 5 pulmonary
arterioles with a diameter of 50 to 100 µm from each rat were then
selected. The pulmonary artery medial wall thickness/external
diameter ratio (WT %) and pulmonary artery medial wall
cross-sectional area/total vessel area (WA %) were determined from
the images by analysis using Image-Pro Plus v6.0 software (Media
Cybernetics; Rockville, MD, USA) to assess medial wall
hypertrophy.
Plasma B-type natriuretic peptide
(BNP) concentration analysis by ELISA
Blood (~2 ml) was collected from 10newborn rats per
group following decapitation, and placed into an EDTA
anticoagulation tube. Blood samples were centrifuged at 1,600 × g
for 20 min at 4°C, and plasma was removed and stored at −80°C for
further study. Plasma BNP concentrations were measured using the
Rat BNP 32 ELISA kit (cat. no. ab108815; Abcam), according to the
manufacturer's protocol. The plates were read at a wavelength of
450 nm (Tecan Sunrise Microplate reader; Tecan Group Ltd.,
Männedorf, Switzerland) and the level of BNP was determined for
each sample from a standard curve. In the present study, 10 samples
from each group were used and experiments on each sample were
repeated 3 times.
Western blot analysis
Samples (80 mg) were obtained from 8 different
rats/group. Briefly, to each lung sample 600 µl
radioimmunoprecipitation lysis buffer (Beyotime Institute of
Biotechnology, Shanghai, China) and 6 µl PMSF (Beyotime Institute
of Biotechnology) were added. Samples were then agitated using
ultrasonication, left for 10 min then centrifuged at 14,000 × g and
4°C for 30 min. The supernatants were then collected. The total
protein extracted from lung tissue was quantified using the
bicinchoninic acid assay. A total of 50 µg of each protein extract
was separated by 8% SDS-PAGE and transferred onto polyvinylidene
difluoride membranes (EMD Millipore, Billerica, MA, USA). The
membranes were incubated for 2–3 h in 5% bovine serum albumin to
block nonspecific binding. Following blocking, the membranes were
incubated with primary antibodies against PPAR-γ (1:400; rabbit
polyclonal; cat. no. ab19481; Abcam), TRPC1 (1:200; rabbit
polyclonal; cat. no. T8276; Sigma-Aldrich; Merck KGaA), TRPC6
(1:200, rabbit polyclonal; cat. no. T6442; Sigma-Aldrich; Merck
KGaA) and GAPDH (1:5,000; mouse monoclonal; cat. no. ab9484;
Abcam), diluted in Tris-buffered saline + 1% Tween-20 (TBST)
overnight at 4°C. Membranes were washed in TBST and incubated for 2
h at room temperature with horseradish peroxidase-conjugated goat
anti-rabbit or goat anti-mouse antibodies (1:2,000; cat. nos.
ZB-2301 or ZB-2305, respectively; Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd.; OriGene Technologies, Inc.) and visualized
using enhanced chemiluminescence reagents (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). Densitometric analysis of
the protein bands was performed using Image-Pro Plus software v6.0
(Media Cybernetics, Inc.) and then normalized to GAPDH.
Total RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Eight samples (70 mg) from each group were obtained
and total RNA was extracted using TRIzol reagent (Takara
Biotechnology Co., Ltd., Dalian, China), according to the
manufacturer's protocol. RNA purity and concentration were
determined according to the OD 260/280 nm ratio. mRNA was reverse
transcribed into cDNA using the PrimeScript RT Reagent kit (Takara
Biotechnology Co., Ltd.). PCR was performed with a 20 µl final
volume reaction mixture SYBR-Green PCR reagent kit (Takara
Biotechnology Co., Ltd.) according to the manufacturer's
instructions, on the Applied Biosystems 7500 Real-Time PCR system
(Applied Biosystems 7500; Thermo Fisher Scientific, Inc., Waltham,
MA, USA). The cDNA PCR conditions were as follows: 1 cycle of 95°C
for 30 sec, then 40 cycles of 95°C for 5 sec and 60°C for 34 sec.
The gene expression levels were calculated with the 2-∆∆Cq method
(22). GAPDH served as an internal
control. Primer sequences are provided in Table I.
 | Table I.Primer sequences used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Primer sequence
(5′→3′) |
|---|
| PPAR-γ | F:
ACCACAGTTGATTTCTCC AG |
|
| R:
TGTTGTAGAGCTGGGTCTTT |
| TRPC1 | F:
AGCCTCTTGACAAACGAGGA |
|
| R:
ACCTGACATCTGTCCGAACC |
| TRPC6 | F:
TACTGGTGTGCTCCTTGCAG |
|
| R:
GAGCTTGGTGCCTTCAAATC |
| GAPDH | F:
AGACAGCCGCATCTTCTTGT |
|
| R:
CTTGCCGTGGGTAGAGTCAT |
Statistical analysis
Data were presented as the mean ± standard
deviation. Statistical analysis was performed by Student's t-test,
using the SPSS version 17.0 software (SPSS, Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Pulmonary vascular remodeling
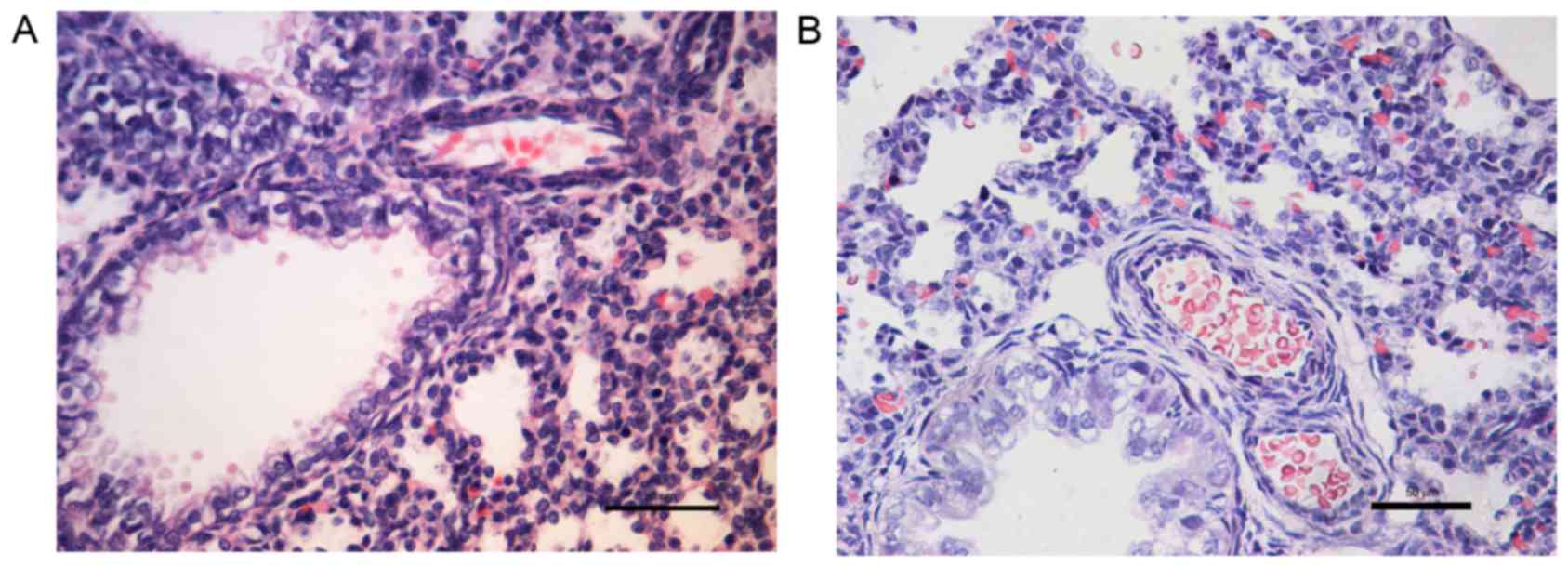
H&E staining revealed that when compared with
the control group (Fig. 1A), the
small pulmonary artery wall was thicker in the PPHN group (Fig. 1B).
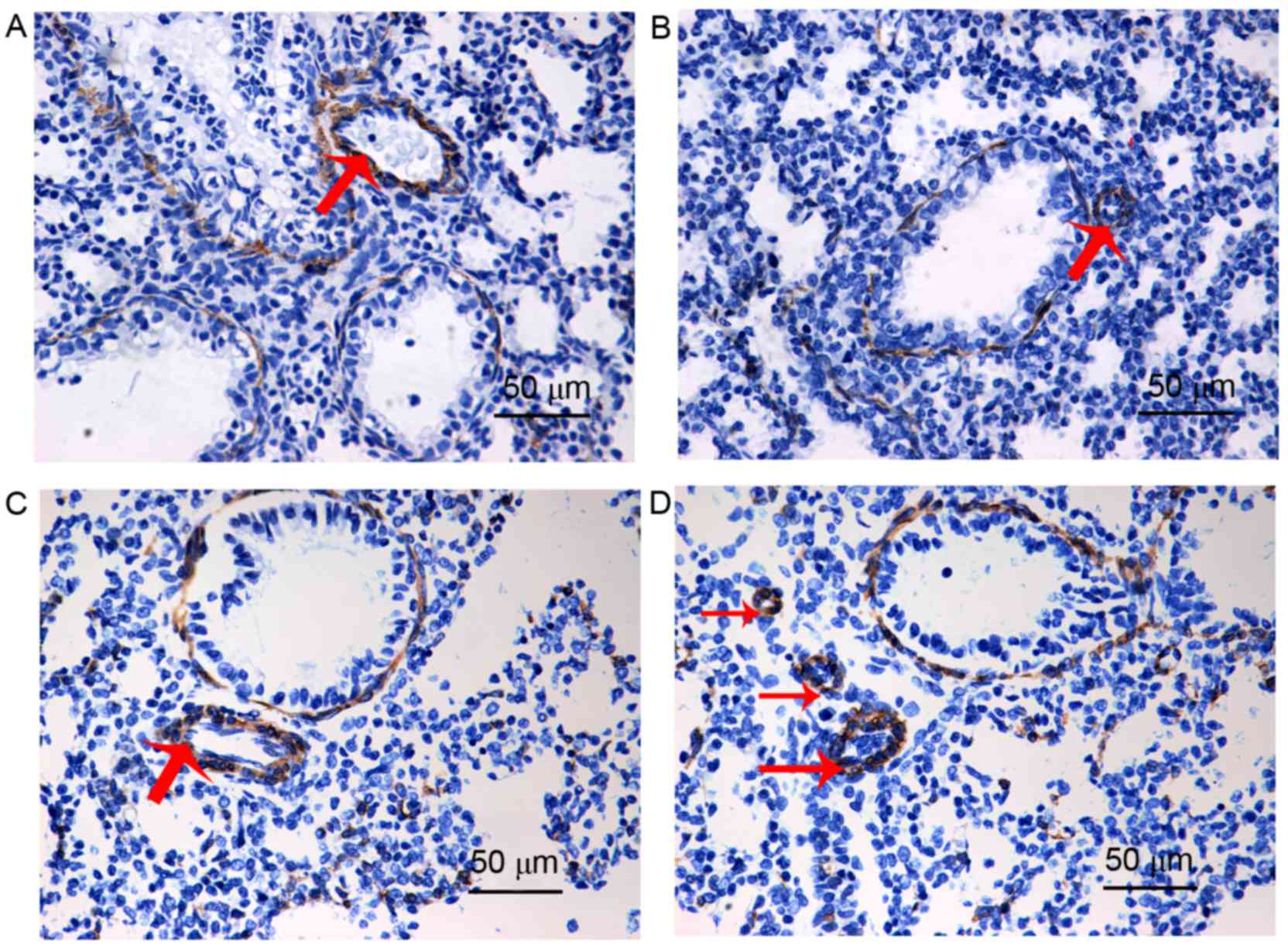
The α-SMA antibody can be used to mark vascular
smooth muscle, thus, in the present study, circular areas of
positive α-SMA staining indirectly indicated the smooth muscle of
the pulmonary artery or distal pulmonary arterioles. When compared
with the control group (Fig. 2A),
the pulmonary arterial medial wall was thicker in the PPHN group
(Fig. 2C). In addition, when
compared with the control group (Fig.
2B), the distal pulmonary arterioles were greatly increased in
the PPHN group (Fig. 2D), which
indicated that muscularization of distal pulmonary arterioles was
increased in the PPHN group, when compared with the control
group.
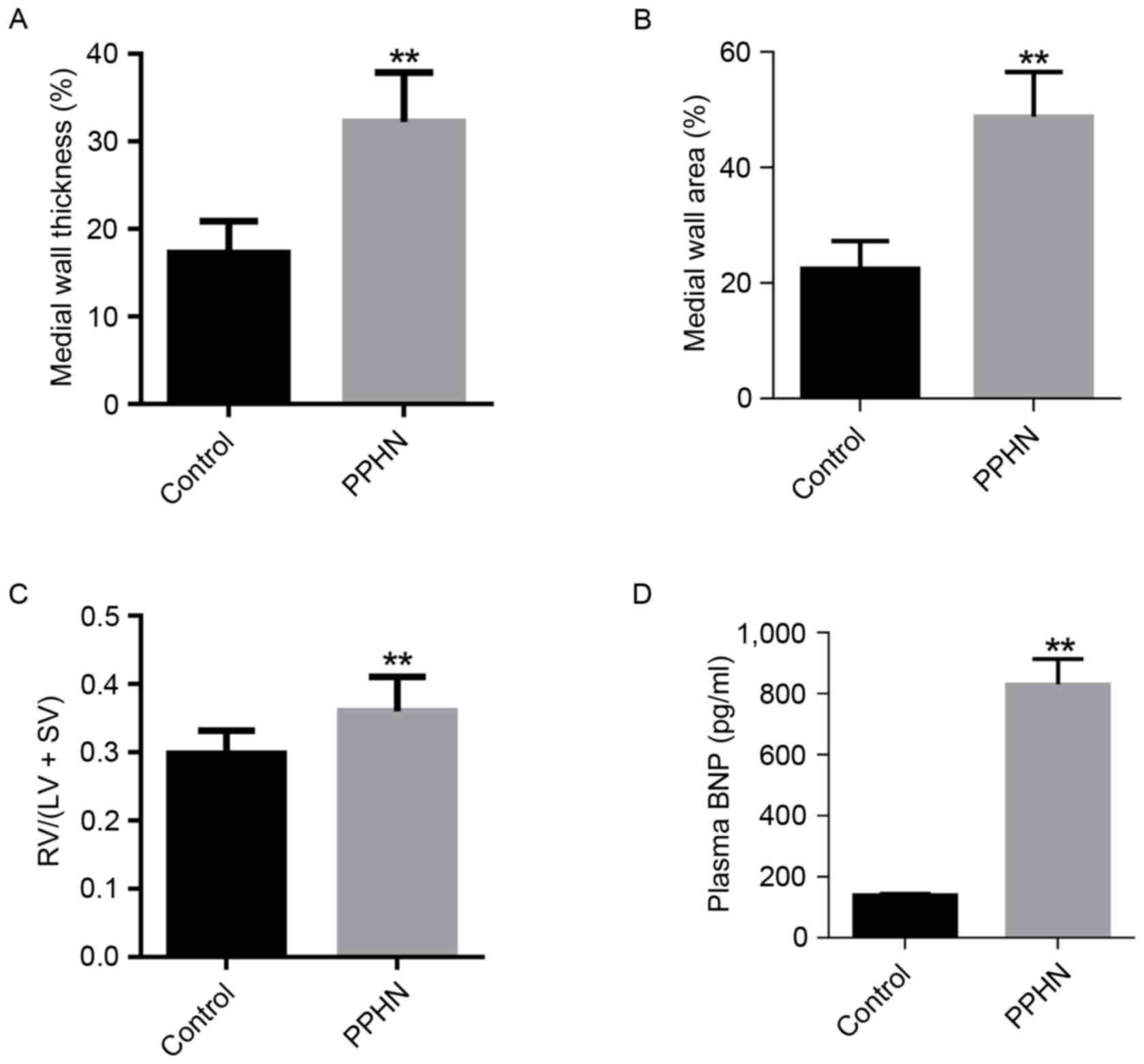
The small pulmonary arteries (50–100 µm in diameter)
were compared, and the WT % in the PPHN group was higher than that
of the control group (P<0.01; Fig.
3A). In addition, the WA % of the PPHN group was also
significantly greater than that of the control group (P<0.01;
Fig. 3B).
Right ventricle hypertrophy
Compared with the control group, the RVHI [RV/(LV +
SV)] was significantly increased in the PPHN group (P<0.01;
Fig. 3C).
Plasma BNP levels
Plasma BNP concentration was measured to indirectly
reflect the pulmonary arterial pressure. In the PPHN group, the
plasma BNP level was 872±35 pg/ml, whereas the plasma BNP
concentration was 139±7 pg/ml in the control group (P<0.01;
Fig. 3D).
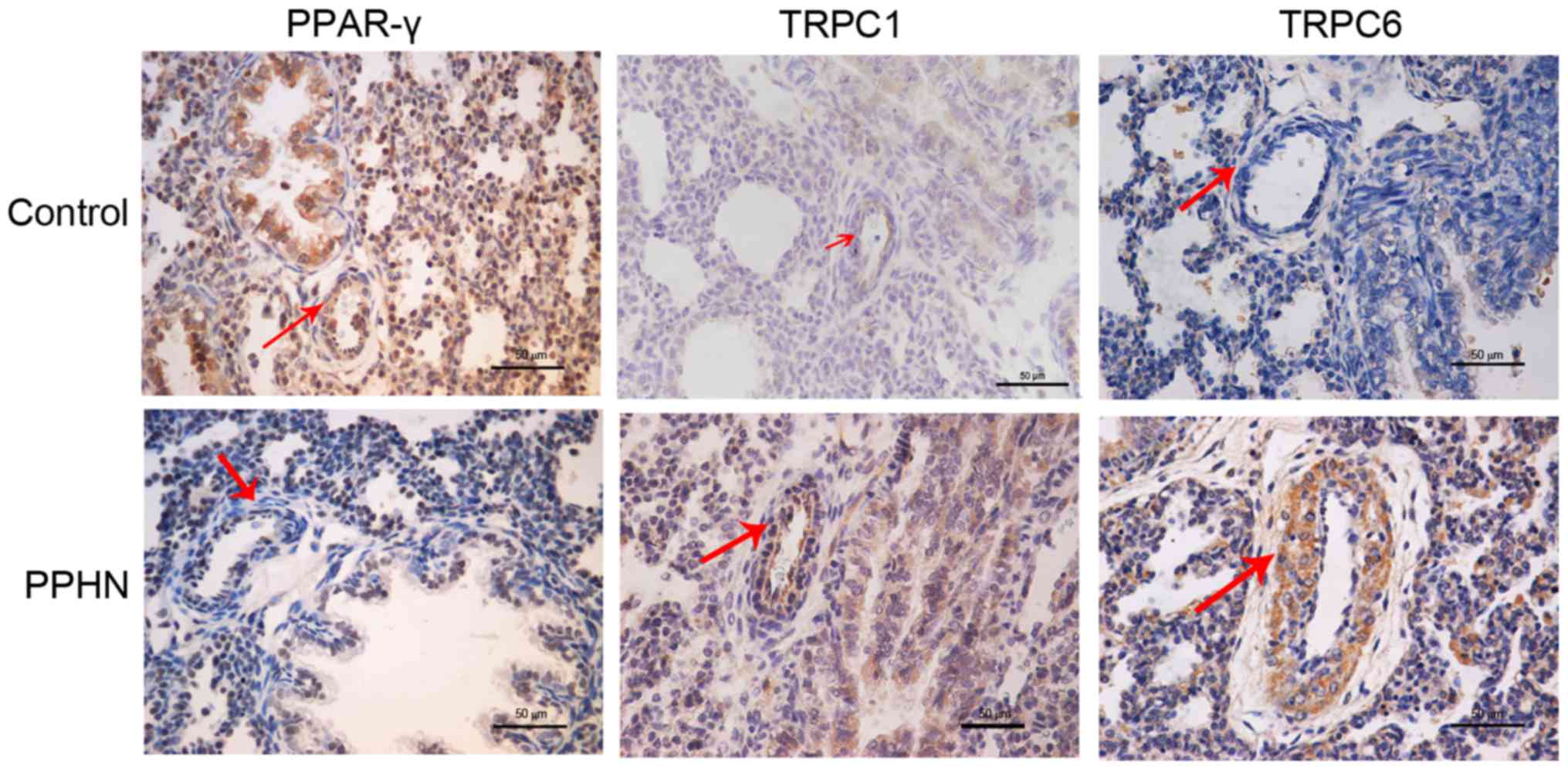
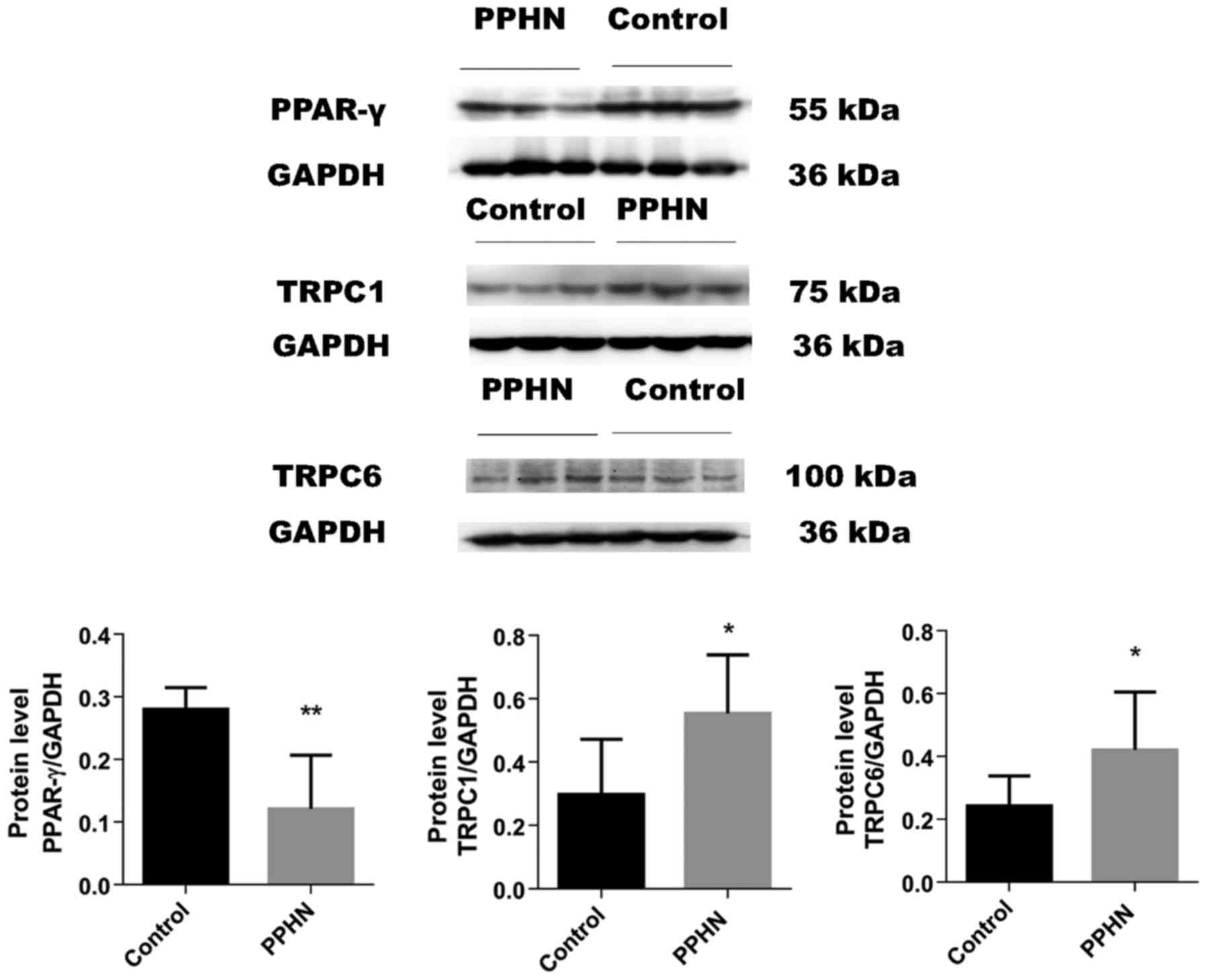
PPAR-γ, TRPC1 and TRPC6 protein
localization in lung tissues
PPAR-γ, TRPC1 and TRPC6 protein expression and
localization in lung tissues were examined by immunohistochemical
staining (Fig. 4). PPAR-γ protein
expression was observed in the cytoplasm and nuclei of pulmonary
vascular endothelial cells and PASMCs. PPAR-γ expression in the
PPHN group was notably lower compared with the control group. TRPC1
and TRPC6 protein expressions were observed in the cytoplasm of
PASMCs, and the expression of both proteins appeared to be greatly
increased in the PPHN group compare with rats in the control
group.
The results of western blot analysis demonstrated
that the protein expression level of PPAR-γ was significantly
decreased in the PPHN group compared with the control group
(Fig. 5; P<0.05). In the PPHN
group, the expression levels of TRPC1 and TRPC6 proteins in lung
tissues greatly increased (Fig. 5;
P<0.05).
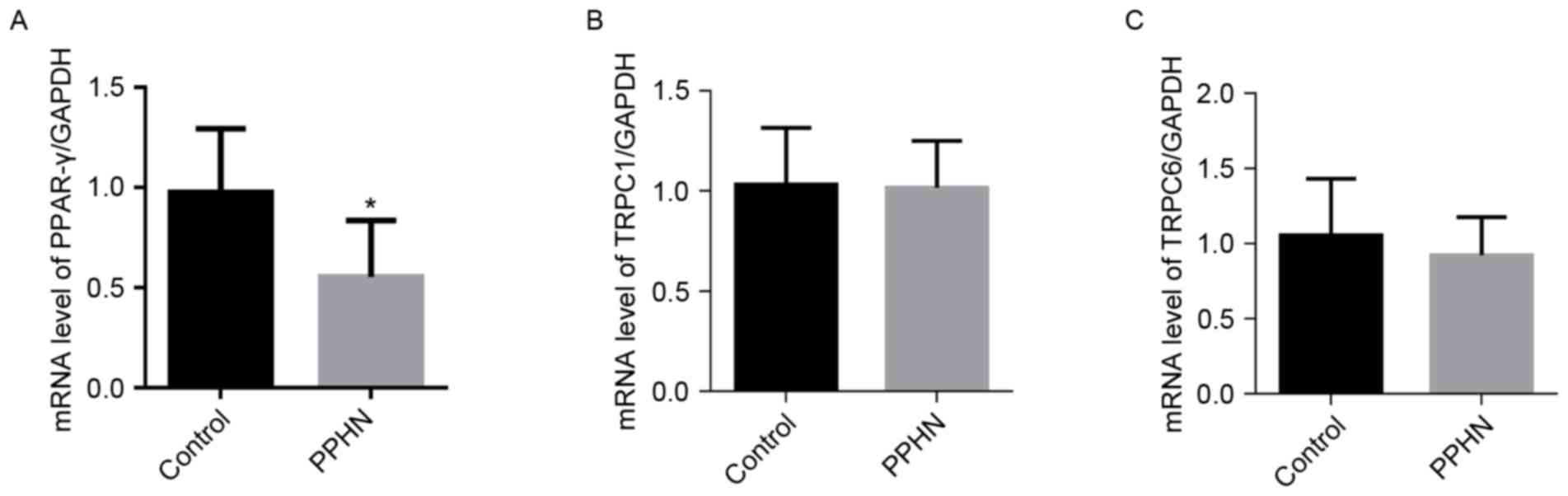
mRNA expression levels of PPAR-γ,
TRPC1 and TRPC6 in lung tissues
The mRNA expression level of PPAR-γ in the PPHN
group was significantly lower than the levels detected in the
control group (Fig. 6A;
P<0.01). However, no significant differences were identified in
the mRNA expression levels of TRPC1 and TRPC6 in the PPHN group
compared with the control group (Fig.
6B and C; P>0.05).
Discussion
PPHN is a severe cardiopulmonary disorder that is
characterized by pulmonary vasoconstriction and late pulmonary
vascular remodeling, which results in significantly increased
pulmonary arterial pressure and right ventricular hypertrophy
(23,24). PPHN mainly affects full-term
infants and near full-term infants, but may also affect premature
infants (25), and the prevalence
of PPHN is 0.43–6.8 per 1,000 births, with a mortality rate of
10–20% (19). Furthermore, PPHN
has long-term complications, including neurodevelopmental,
cognitive and hearing abnormalities (26). Therefore, there is a necessity and
urgency to elucidate the underlying mechanism for the pathogenesis
of PPHN.
Indomethacin is widely used to treat patent ductus
arteriosus in human neonates (27,28).
Previous studies have demonstrated that a partial ligation of the
arterial duct in utero was able to induce PPHN in sheep
(29,30). PH has also been reported to be
induced by continuous hypoxia (31–33).
In addition, a previous report demonstrated that the combination of
indomethacin and hypoxia was able to successfully established PPHN
in rats (21). The present study
used a similar concept, which combined exposure to hypoxic
conditions with indomethacin treatment to establish a PPHN rat
model in prenatal rats during days 19–21 of gestation. The results
demonstrated that WA % and WT % were markedly increased in the PPHN
group compared with rats in the control group. These results
suggested that the PASMCs were significantly increased and that
pulmonary vascular remodeling was present in the PPHN group. In
addition, the expression of α-SMA in fibroblasts was greatly
increased in the PPHN group compared with the control group. This
increase in α-SMA content may indicate that, when compared with the
control group, distal muscularization of the pulmonary arteries was
increased in the PPHN group.
BNP is used to determine the severity of right
ventricular failure and outcomes from PH in humans (34). In the present study, plasma BNP
concentrations were significantly higher in rats in the PPHN group
compared with the control group, suggesting that the PPHN group may
have an enhanced degree of PH. To further examine the state of PH
in the PPHN group RV/(LV + SV) was calculated, which is used to
evaluate right ventricle hypertrophy. Overall, the PPHN group
exhibited a significant increase in pulmonary artery pressure and
in the thickness of pulmonary artery medial smooth muscle, as well
as right ventricular hypertrophy. These results indicated that the
PPHN rat model was successfully established.
A number of previous studies have demonstrated that
the loss of PPAR-γ expression may be closely related to PH. One
study reported that mice with a targeted deletion of PPAR-γ in
PASMCs developed PH in addition to increased muscularization of the
distal pulmonary arteries (16).
In addition, the PPAR-γ agonist rosiglitazone was revealed to
ameliorate PH in rat and mouse models. For example, rosiglitazone
was reported to attenuate the development of PH and pulmonary
vascular remodeling in hypoxia-induced PH model rats (35). Another study demonstrated that
treatment with PPAR-γ agonists was able to attenuate
hypoxia-induced pulmonary vascular remodeling and PH by suppressing
oxidative and proliferative signals in mice (36). Furthermore, treatment of PH with
pioglitazone, another PPAR-γ agonist, was reported to decrease
pulmonary arterial systolic pressure, medial wall thickness and
muscularization of small pulmonary arteries (15). The present study revealed that the
expression of PPAR-γ in newborn rat lung tissues was decreased in
the PPHN group, leading to the hypothesis that PPAR-γ may serve a
protective role in the pathophysiology of PPHN.
Over the past several years, a number of studies
have demonstrated that increased concentration of intracellular
Ca2+ is a major stimulus for PASMC proliferation and
migration (11). A previous study
has also indicated that the hypoxia-enhanced SOCE through the SOCC
is highly associated with the elevated concentration of
Ca2+ (11). In PASMCs
of hypoxia-induced PH and monocrotaline-induced PH (37), the canonical TRPCs are important
aspects of the SOCC, in which TRPC1, TRPC4 and TRPC6 are
significantly expressed in the rat pulmonary artery and PASMCs
(38). Chronic hypoxia was
reported to increase the expression of TRPC1 and TRPC6 (38), but not TRPC4 expression, in PASMCs
isolated from rats as well as in cultured PASMCs (39). Based on these reports, the present
study hypothesized that TRPC1 and TRPC6 were related to the
pathophysiology of PPHN. To test this hypothesis, the expression
levels of TRPC1 and TRPC6 were examined in PPHN model rats.
Although no differences were observed in the mRNA expression levels
of TRPC1 and TRPC6, the protein expression levels of TRPC1 and
TRPC6 were significantly increased in the lung tissues isolated
from the PPHN group. The inconsistency of mRNA and protein
expression in TRPC1 and TRPC6 may be due to posttranscriptional
modifications (40). These results
indicated that upregulated expression levels of TRPC1 and TRPC6
protein may account for the pathogenesis of PPHN.
A recent report demonstrated that PPAR-γ may inhibit
chronic hypoxia-induced PH by downregulating the expression of
TRPC1 and TRPC6 (10). In
addition, sildenafil was reported to attenuate PH as indicated by a
reduction in the levels of SOCE and downregulation of TRPC1 and
TRPC6 expression in rat PASMCs via the cyclic GMP/protein kinase
G/PPAR-γ axis (41). Another study
reported that PPAR-γ inhibited hypoxia-induced SOCE by
downregulating caveolin-1 expression or downregulating TRPC1 and
TRPC6 expression (11), which
suggested that PPAR-γ may inhibit PPHN by downregulating the
expression of TRPC1 and TRPC6 or caveolin-1.
In conclusion, the present study established a PPHN
rat model and observed altered expressions of PPAR-γ, TRPC1 and
TRPC6 in the pulmonary artery located in the lungs of newborn rats
with PPHN, suggesting that these proteins may be important
mediators of PPHN. It was also speculated that PPAR-γ may inhibit
PPHN by upregulating the expression of TRPC1 and TRPC6. Further
investigations are required to verify these results and to
elucidate the underlying mechanisms for the pathogenesis of
PPHN.
Acknowledgements
The present study was supported by The National
Natural Science Foundation of China (grant nos. 81471489 and
81571479).
Glossary
Abbreviations
Abbreviations:
|
BNP
|
B-type natriuretic peptide
|
|
PASMC
|
pulmonary artery smooth muscle
cell
|
|
PH
|
pulmonary hypertension
|
|
PPAR-γ
|
peroxisome proliferator-activated
receptor γ
|
|
PPHN
|
persistent pulmonary hypertension of
the newborn
|
|
TRPC
|
transient receptor potential cation
channel
|
References
|
1
|
Jain A and McNamara PJ: Persistent
pulmonary hypertension of the newborn: Advances in diagnosis and
treatment. Semin Fetal Neonatal Med. 20:262–271. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Keith IM, Tjen-A-Looi S, Kraiczi H and
Ekman R: Three-week neonatal hypoxia reduces blood CGRP and causes
persistent pulmonary hypertension in rats. Am J Physiol Heart Circ
Physiol. 279:H1571–H1578. 2000.PubMed/NCBI
|
|
3
|
Afolayan AJ, Eis A, Alexander M,
Michalkiewicz T, Teng RJ, Lakshminrusimha S and Konduri GG:
Decreased endothelial nitric oxide synthase expression and function
contribute to impaired mitochondrial biogenesis and oxidative
stress in fetal lambs with persistent pulmonary hypertension. Am J
Physiol Lung Cell Mol Physiol. 310:L40–L49. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Steinhorn RH: Neonatal pulmonary
hypertension. Pediatr Crit Care Med. 11 2 Suppl:79–84. 2010.
View Article : Google Scholar
|
|
5
|
Postolow F, Fediuk J, Nolette N, Hinton M
and Dakshinamurti S: Thromboxane promotes smooth muscle phenotype
commitment but not remodeling of hypoxic neonatal pulmonary artery.
Fibrogenesis Tissue Repair. 8:202015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sluiter I, Reiss I, Kraemer U, Krijger Rd,
Tibboel D and Rottier RJ: Vascular abnormalities in human newborns
with pulmonary hypertension. Expert Rev Respir Med. 5:245–256.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo SJ, Wang T, Jia LQ, Li DD, Shen YC, Xu
D and Wen FQ: TRAM-34 attenuates hypoxia induced pulmonary artery
smooth muscle cell proliferation. Eur Rev Med Pharmacol Sci.
19:3515–3521. 2015.PubMed/NCBI
|
|
8
|
Wan J, Yamamura A, Zimnicka AM, Voiriot G,
Smith KA, Tang H, Ayon RJ, Choudhury MS, Ko EA, Wang J, et al:
Chronic hypoxia selectively enhances L- and T-type
voltage-dependent Ca2+ channel activity in pulmonary
artery by upregulating Cav1.2 and Cav3.2. Am J Physiol Lung Cell
Mol Physiol. 305:L154–L164. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yamamura A, Yamamura H, Zeifman A and Yuan
JX: Activity of Ca -activated Cl channels contributes to regulating
receptor- and store-operated Ca entry in human pulmonary artery
smooth muscle cells. Pulm Circ. 1:269–279. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Lu W, Yang K, Wang Y, Zhang J, Jia
J, Yun X, Tian L, Chen Y, Jiang Q, et al: Peroxisome
proliferator-activated receptor gamma inhibits pulmonary
hypertension targeting store-operated calcium entry. J Mol Med
(Berl). 93:327–342. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang K, Lu W, Jiang Q, Yun X, Zhao M,
Jiang H and Wang J: Peroxisome proliferator-activated receptor
γ-mediated inhibition on hypoxia-triggered store-operated calcium
entry. A caveolin-1-dependent mechanism. Am J Respir Cell Mol Biol.
53:882–892. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nawar NN, Mohamed AH, Adieb N and Emad M:
The effect of maternal Naja nigricollis envenomation on the
placenta. An experimental study. Biol Struct Morphog. 2:13–17.
1989.PubMed/NCBI
|
|
13
|
Zhang D, Wang G, Han D, Zhang Y, Xu J, Lu
J, Li S, Xie X, Liu L, Dong L and Li M: Activation of PPAR-γ
ameliorates pulmonary arterial hypertension via inducing heme
oxygenase-1 and p21(WAF1): An in vivo study in rats. Life Sci.
98:39–43. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takano H and Komuro I: Peroxisome
proliferator-activated receptor gamma and cardiovascular diseases.
Circ J. 73:214–220. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Behringer A, Trappiel M, Berghausen EM,
Ten Freyhaus H, Wellnhofer E, Odenthal M, Blaschke F, Er F,
Gassanov N, Rosenkranz S, et al: Pioglitazone alleviates cardiac
and vascular remodelling and improves survival in monocrotaline
induced pulmonary arterial hypertension. Naunyn Schmiedebergs Arch
Pharmacol. 389:369–379. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hansmann G, de Jesus Perez VA, Alastalo
TP, Alvira CM, Guignabert C, Bekker JM, Schellong S, Urashima T,
Wang L, Morrell NW and Rabinovitch M: An antiproliferative
BMP-2/PPARgamma/apoE axis in human and murine SMCs and its role in
pulmonary hypertension. J Clin Invest. 118:1846–1857. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bijli KM, Kleinhenz JM, Murphy TC, Kang
BY, Adesina SE, Sutliff RL and Hart CM: Peroxisome
proliferator-activated receptor gamma depletion stimulates Nox4
expression and human pulmonary artery smooth muscle cell
proliferation. Free Radic Biol Med. 80:111–120. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xie X, Wang G, Zhang D, Zhang Y, Zhu Y, Li
F, Li S and Li M: Activation of peroxisome proliferator-activated
receptor γ ameliorates monocrotaline-induced pulmonary arterial
hypertension in rats. Biomed Rep. 3:537–542. 2015.PubMed/NCBI
|
|
19
|
Xu XF, Ma XL, Shen Z, Wu XL, Cheng F and
Du LZ: Epigenetic regulation of the endothelial nitric oxide
synthase gene in persistent pulmonary hypertension of the newborn
rat. J Hypertens. 28:2227–2235. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fabris VE, Pato MD and Belik J:
Progressive lung and cardiac changes associated with pulmonary
hypertension in the fetal rat. Pediatr Pulmonol. 31:344–353. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu XF, Gu WZ, Wu XL, Li RY and Du LZ:
Fetal pulmonary vascular remodeling in a rat model induced by
hypoxia and indomethacin. J Matern Fetal Neonatal Med. 24:172–182.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Stayer SA and Liu Y: Pulmonary
hypertension of the newborn. Best Pract Res Clin Anaesthesiol.
24:375–386. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dakshinamurti S: Pathophysiologic
mechanisms of persistent pulmonary hypertension of the newborn.
Pediatr Pulmonol. 39:492–503. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cabral JE and Belik J: Persistent
pulmonary hypertension of the newborn: Recent advances in
pathophysiology and treatment. J Pediatr (Rio J). 89:226–242. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nair J and Lakshminrusimha S: Update on
PPHN: Mechanisms and treatment. Semin Perinatol. 38:78–91. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lemmers PM, Benders MJ, D'Ascenzo R,
Zethof J, Alderliesten T, Kersbergen KJ, Isgum I, de Vries LS,
Groenendaal F and van Bel F: Patent ductus arteriosus and brain
volume. Pediatrics. 137:pii: e201530902016. View Article : Google Scholar
|
|
28
|
Louis D, Torgalkar R, Shah J, Shah PS and
Jain A: Enteral feeding during indomethacin treatment for patent
ductus arteriosus: Association with gastrointestinal outcomes. J
Perinatol. 36:544–548. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dodson RB, Morgan MR, Galambos C, Hunter
KS and Abman SH: Chronic intrauterine pulmonary hypertension
increases main pulmonary artery stiffness and adventitial
remodeling in fetal sheep. Am J Physiol Lung Cell Mol Physiol.
307:L822–L828. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Konduri GG, Bakhutashvili I, Eis A and
Afolayan A: Antenatal betamethasone improves postnatal transition
in late preterm lambs with persistent pulmonary hypertension of the
newborn. Pediatr Res. 73:621–629. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang K, Lu W, Jia J, Zhang J, Zhao M, Wang
S, Jiang H, Xu L and Wang J: Noggin inhibits hypoxia-induced
proliferation by targeting store-operated calcium entry and
transient receptor potential cation channels. Am J Physiol Cell
Physiol. 308:C869–C878. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun H, Xia Y, Paudel O, Yang XR and Sham
JS: Chronic hypoxia-induced upregulation of
Ca2+-activated Cl channel in pulmonary arterial
myocytes: A mechanism contributing to enhanced vasoreactivity. J
Physiol. 590:3507–3521. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Peng G, Wang J, Lu W and Ran P: Isolation
and primary culture of rat distal pulmonary venous smooth muscle
cells. Hypertens Res. 33:308–313. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jone PN, Patel SS, Cassidy C and Ivy DD:
Three-dimensional echocardiography of right ventricular function
correlates with severity of pediatric pulmonary hypertension.
Congenit Heart Dis. 11:562–569. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim EK, Lee JH, Oh YM, Lee YS and Lee SD:
Rosiglitazone attenuates hypoxia-induced pulmonary arterial
hypertension in rats. Respirology. 15:659–668. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nisbet RE, Bland JM, Kleinhenz DJ,
Mitchell PO, Walp ER, Sutliff RL and Hart CM: Rosiglitazone
attenuates chronic hypoxia-induced pulmonary hypertension in a
mouse model. Am J Respir Cell Mol Biol. 42:482–490. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Guo Q, Huang JA, Yamamura A, Yamamura H,
Zimnicka AM, Fernandez R and Yuan JX: Inhibition of the
Ca(2+)-sensing receptor rescues pulmonary hypertension in rats and
mice. Hypertens Res. 37:116–124. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yun X, Chen Y, Yang K, Wang S, Lu W and
Wang J: Upregulation of canonical transient receptor potential
channel in the pulmonary arterial smooth muscle of a chronic
thromboembolic pulmonary hypertension rat model. Hypertens Res.
38:821–828. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang J, Weigand L, Lu W, Sylvester JT,
Semenza GL and Shimoda LA: Hypoxia inducible factor 1 mediates
hypoxia-induced TRPC expression and elevated intracellular
Ca2+ in pulmonary arterial smooth muscle cells. Circ
Res. 98:1528–1537. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lotfi K, Karlsson K, Fyrberg A, Juliusson
G, Jonsson V, Peterson C, Eriksson S and Albertioni F: The pattern
of deoxycytidine- and deoxyguanosine kinase activity in relation to
messenger RNA expression in blood cells from untreated patients
with B-cell chronic lymphocytic leukemia. Biochem Pharmacol.
71:882–890. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang J, Yang K, Xu L, Zhang Y, Lai N,
Jiang H, Zhang Y, Zhong N, Ran P and Lu W: Sildenafil inhibits
hypoxia-induced transient receptor potential canonical protein
expression in pulmonary arterial smooth muscle via cGMP-PKG-PPARγ
axis. Am J Respir Cell Mol Biol. 49:231–240. 2013. View Article : Google Scholar : PubMed/NCBI
|