Introduction
Spinal cord injury poses a serious threat to human
health. Spinal cord injury that occurs as a result of an external
force, leads to primary nerve tissue damage and the rapid
occurrence of secondary injury due to the activation of complex
cascade reactions, which prevent the successful repair of spinal
cord injury (1–5). Surgical decompression treatment is
used during the early stage of clinical treatment for spinal cord
injury. If cell transplantation using neural stem cells (NSCs),
olfactory ensheathing cells (OECs) and bone marrow stromal cells
(BMSCs) were to be simultaneously combined with this method, the
treatment may demonstrate beneficial effects, and may be useful in
clinical practice. Previous studies investigating the
pathophysiology of spinal cord injury have demonstrated that the
formation of the cystic cavity following spinal cord injury is an
important factor in the prevention of axonal regeneration (6,7).
Therefore, effective treatments aim to prevent or reduce the
formation of the cystic cavity following spinal cord injury,
improve the micro-environment of the spinal cord injury sites and
protect the damaged nerve cells, which is conducive to axonal
regeneration (3,8,9).
Cell transplantation using NSCs, OECs and BMSCs is considered to be
one of the most promising methods for the treatment of spinal cord
injury. If the transplanted cells survive, proliferate in the site
of spinal cord injury and integrate in the host tissues, it may be
possible to protect the damaged spinal cord tissue, promote axonal
regeneration and enhance functional recovery (1,8–11).
Bone marrow stromal cells (BMSCs) are easy to
acquire and culture, proliferate well in vitro and possess
clinical potential. In response to different inducing factors or
environmental effects, BMSCs differentiate into neuron-like cells
(12–16). Previous studies have demonstrated
that BMSCs efficiently differentiate into neuron-like cells in the
presence of factors secreted by OECs (17–19).
BMSCs have been used in the treatment and repair of nervous system
injuries, and were observed to promote axonal regeneration by
varying degrees, which led to improved motor function (3,4,19–21).
The primary aim of the present study was to determine whether the
application of transplanted OEC-induced BMSC neural-like cells
increases the recovery of injured spinal cord function, improve
cell survival in the harsh environment following spinal cord
injury, and promotes axonal regeneration.
Materials and methods
Culture and purification of OECs and
preparation of conditioned medium
A total of 62 adult male Sprague-Dawley rats
(weight, 200–220 g; age, 6 weeks) were obtained from the
Experimental Animal Center of North China University of Science and
Technology (Tangshan, China). They were individually housed in
clear cages in a controlled environment with constant temperature
of 23.8°C and humidity of 50±10%. The animal room was on a 12:12 h
light:dark cycle and with food and water available ad
libitum. The present study was performed in strict accordance
with the recommendations outlined in the Guide for the Care and Use
of Laboratory Animals (13). The
animal use protocol was reviewed and approved by the Institutional
Animal Care and Use Committee of North China University of Science
and Technology. A rat (250 g) was sacrificed by cervical
dislocation and the olfactory bulbs were separated under sterile
conditions, the pia mater was discarded, and the olfactory
nerve layer and the olfactory bulb granular layer were separated.
The granular layer was washed twice with D-Hank's buffer, placed in
a 37°C incubator, and digested with 0.125% trypsin (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) for 25 min. Digestion was
terminated with a trypsin terminator (0.2 mM/l; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) for 8 min at 37°C and the mixture
was centrifuged at 71.6 × g for 5 min at 37°C, before it was washed
once with serum-free Dulbecco's modified Eagle's medium (DMEM)/F12
medium (Sigma-Aldrich; Merck KGaA). Finally, single cell
suspensions were produced using DMEM/F12 medium containing 20%
fetal calf serum (FCS; Sigma-Aldrich; Merck KGaA), seeded in
plastic culture flasks and cultured in an incubator at 37°C and 5%
CO2. In accordance with the modified Nash differential
adherent method (2,3), the cells were aspirated and
transferred onto polylysine (Sigma-Aldrich; Merck KGaA)-coated
plastic flasks following 18–20 h of culture. Cytarabine (Ara-C;
Sigma-Aldrich; Merck KGaA) was then added 18–20 h later. Cells were
incubated with 3–5 µM/l Ara-C for 24–48 h, based on the number of
fibroblasts as counted using light microscopy. The medium was
refreshed every 2–3 days, with one-third of the medium replaced in
the first 6 days, followed by 50% of the medium thereafter. The
replacement medium contained 100 ml 20% FCS. Once the purified OECs
had reached 70–80% confluence, (following ~9–12 days), the culture
medium was discarded and the cells were washed with fresh medium
(DMEM/F12 containing 15% FCS) twice before fresh medium was added.
The culture supernatant was aspirated following 24 h and
centrifuged at 670.8 × g for 20 min at 37°C. The supernatant was
then collected and used as OEC conditioned medium in subsequent
experiments.
BMSC culture and induced labeling
prior to transplantation
A rat (weight, 150 g) was sacrificed by cervical
dislocation, and the complete bilateral femur and tibia were
obtained under sterile conditions. Both sides of the metaphysis
were successively cut to expose the bone marrow cavity, and the
cavity was washed with DMEM medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) 2–3 times with a 2 ml syringe.
Collected fluid was filtered with a 200 mesh filter, before an
equal volume of 0.84% NH4Cl solution was added (3). The mixture was centrifuged at 850
r/min for 10 min at 37°C and the supernatant was discarded. Cells
were resuspended in DMEM/F12 (Gibco; Thermo Fisher Scientific,
Inc.) containing 15% FCS, inoculated in culture flasks and placed
into a humidified incubator at 37°C and 5% CO2. The
medium was refreshed following 72 h, and then once every three
days. When the cells had reached ~80% confluence, they were
digested with 0.25% trypsin (Sigma-Aldrich; Merck KGaA) for 10 min
at 37°C and subcultured. BMSCs were separated from monocytes and
lymphocytes, as the latter cell types adhere to the bottom of the
culture flask while BMSCs are less adherent and are removed more
easily. The cells were cultured until passage 3, at which point
they were adherent and four culture flasks were selected at random.
The medium in two flasks was replaced with OEC conditioned medium
to induce the BMSCs for 48 h, and all four flasks of cells were
stained with 10 µg/ml Hoechst 33342 (Sigma-Aldrich; Merck KGaA) for
30 min at 37°C in order to label cell nuclei prior to
transplantation (2,4). The cells were subsequently washed
with 0.01 M phosphate-buffered saline (PBS) 3–5 times and digested
with trypsin for 10 min at 37°C. Cells were then collected by
centrifugation at 80.8 × g for 5 min at 37°C, and resuspended to a
final concentration of ~1×105/µl for further use.
Animal grouping, modeling, and cell
transplantation
Healthy male Sprague-Dawley rats (n=60) were
anesthetized with 1% sodium pentobarbital (30 mg/kg) by
intraperitoneal injection. Under anesthesia, the animals were fixed
on to the spinal cord injury instrument. Conventional skin
preparation was performed as follows: The area was disinfected and
towels were positioned around the T8 segment. The dorsal skin was
successively cut and the muscles were separated on both sides of
the spinous process, which was successively opened. A T7-9
laminectomy was then performed under the operating microscope to
expose the T8 segment of the spinal cord. Each rat underwent spinal
cord compression injury with a plastic spinal cord compression
plate with a thickness of 0.5 mm and a breadth of 2.8 mm. The
compression plate was attached to a copper rod giving a total
weight of 20 g on the spinal cord. The compression plate was
lowered down ventrally at a rate of 0.5 mm/min to the bottom of the
vertebral canal and remained there for 5 min. Following the
successful preparation of models, the animals were divided at
random into groups A, B and C (20 rats/group), which were injected
with DMEM, BMSCs and induced BMSCs, respectively, into the spinal
cord injury sites. The injection methods were as follows: The
spinal cord injury center sites were selected and a point for
injection with a depth of 1.7 mm was selected. The injection volume
was 2 µl and the injection speed was 0.2 µl/min, with the needle
slowly drawn back. The needle was retained for 5 min following
injection. The redundant cells adversely flowed along the needle
passage, so the process did not aggravate the spinal cord injury.
The fascia and skin were sutured following surgery, and
postoperative artificial assistance for drainage was provided twice
per day in the first week and then once per day thereafter.
Postoperative conventional insulation, anti-infection (penicillin
4×106 U/day for 3 days after surgery; Hoechst-Huabei
Pharmaceuticals Co. Ltd., Shijiazhuang, China) and other paraplegia
care were performed to prevent the formation of bedsores.
Evaluation of motor function in the
lower extremities
Basso, Beattie and Bresnahan (BBB) ratings were used
for rats in all groups, in order to assess motor function of the
lower limbs (2–5), and the double-blind method was used
for assessment. BBB scores range between 0 (complete paralysis) and
21 (normal). Early Stage (0–7): Isolated joint movements with
little or no hind limb movement; Intermediate Stage (8–13):
Intervals of uncoordinated walking; and, Late Stage (14–21):
Fore and hind limb coordination. Each rat was assessed for at least
4 min once a week following injury for a total of 8 weeks.
Histological examination of the cavity
area of spinal cord injury
At 2 weeks following surgery, 4 rats were randomly
selected from each group. Following excess anesthesia (1% sodium
pentobarbital; Sigma-Aldrich; Merck KGaA) by intraperitoneal
injection, specimens were fixed with 4% paraformaldehyde via the
left ventricle using conventional perfusion methods. Samples
(~15-mm in length) were obtained from the injured segment of spinal
cord. Following dehydration in 25% sucrose at 4°C, continuous
longitudinal frozen sections (~14-µm in thickness) were prepared.
The remaining rats in each group underwent the same procedure at 4
weeks. Following hematoxylin (0.5%) and eosin (1%) (H&E)
staining for 5 min at room temperature, the injured area of the
spinal cord was observed under a light microscope. Observation and
calculation of the damaged cavity area, the target volume area, and
average area/section of regions of interest in the lesioned spinal
cord were estimated from coronal sections using the Cavalieri
method (2,6).
Immunohistochemical staining
Selected frozen tissue sections of spinal cord
injury (SCI) tissue at 2 weeks and 4 weeks following SCI underwent
immunofluorescence staining. The sections were blocked with 20%
donkey serum (Sigma-Aldrich; Merck KGaA) for 2 h at 37°C and washed
in 0.01 M PBS. The sections were then incubated with primary
antibodies against the following in a refrigerator at 4°C
overnight: Neurofilament (NF; N5264; 1:1,000), growth associated
protein-43 (GAP-43; HPA013392; 1:1,000) and neuron-specific nuclear
protein (NeuN; SAB4300883; dilution, 1:1,000) (all from
Sigma-Aldrich; Merck KGaA). The following day, the sections were
washed 3 times with 0.01 M PBS, and incubated for 24 h at 4°C with
the following fluorescence-labeled secondary antibodies:
fluorescein isothiocyanate (FITC)-labeled donkey anti-mouse IgG
(A24501; 1:400) and Texas Red-labelled donkey anti-rabbit IgG
(PA1-28590; 1:800) (both from Molecular Probes; Thermo Fisher
Scientific, Inc.). The sections were then washed with PBS, mounted
with glycerol and observed under a fluorescence microscope, where
NF and GAP-43 double immunofluorescence was used. The relationship,
similarity and difference in the expression of NF and GAP-43 were
observed using confocal microscopy and analyzed using Adobe
Photoshop CS2 (Adobe Systems, Inc., San Jose, CA, USA).
Statistical analysis
The resulting measurement data from all assays were
normally distributed and expressed as the mean ± standard
deviation. Comparisons among multiple groups were performed using
two-way analysis of variance. Student-Newman-Keuls was used as a
post hoc test and the results between 4 and 8 weeks in each group
was compared using the Student's t-test. SPSS statistical software
(version, 19.0; IBM SPSS, Armonk, NY, USA) was used for data
processing. P<0.05 was considered to indicate a statistically
significant difference.
Results
Primary culture of BMSCs and
purification of OECs
BMSCs began to adhere and exhibit a stretched
morphology at 48 h following inoculation (data not shown). The
majority of cells exhibited fusiform growth, and cell colony
formation was observed (data not shown). Following the medium
change, cells began to proliferate rapidly, and when cells reached
80% confluence, fusion passage culture was performed. BMSCs were
isolated by changing the medium to remove non-adherent cells in
passage culture. In the third passage culture, the morphology of
BMSCs was more consistent, demonstrating fusiform growth and strong
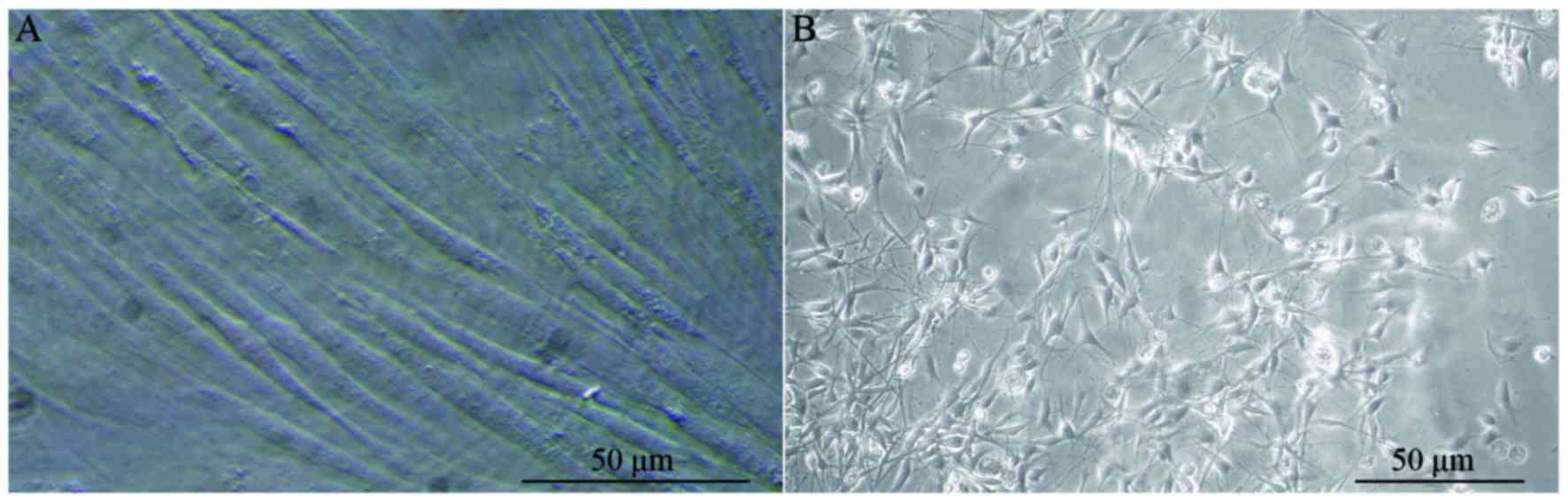
refraction (Fig. 1A). Primary
cultures of OECs were purified using the Nash differential adherent
method and cytarabine pure culture. The growth state of the cells
was observed under a light microscope. The majority of cells were
bipolar and fusiform, most nuclei were oval and distributed in the
center of cells, the cell bodies were clearly visible under the
light microscope and the axons appeared elongated (Fig. 1B).
Transplantation of induced BMSCs
improved motor function following spinal cord injury
The lower limbs of all rats with SCI by extrusion
lost all motor function 24 h following injury and the muscle
strength was 0. The lowest score of motion in BBB ratings was 0,
while the maximum was 21. The hind limb motor function of rats in
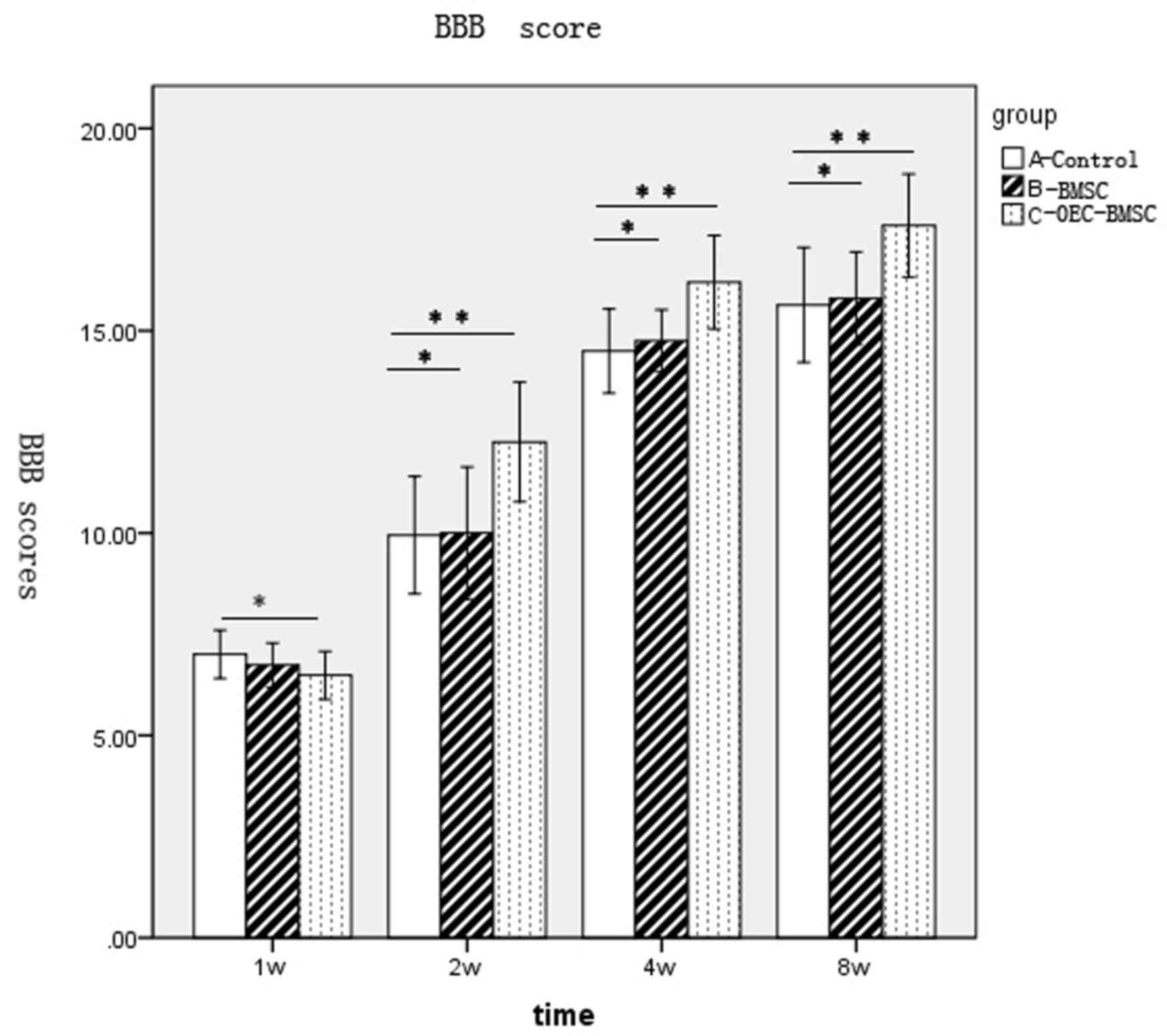
group A appeared to recover at 1 week following surgery (Fig. 2). Functional recovery of behavior
in each group was faster at 2–4 weeks following surgery, with the
difference between BBB scores among groups increasing over time
(Fig. 2). However, behavioral
recovery in each group reduced during 4–8 weeks following surgery,
and entered a relatively stable plateau. No significant difference
in the BBB scores between groups B and A was observed; however, a
significant difference between groups C and groups A and B was
observed at 2, 4 and 8 weeks (Fig.
2).
Spinal cord injury area was reduced
following transplantation of BMSCs
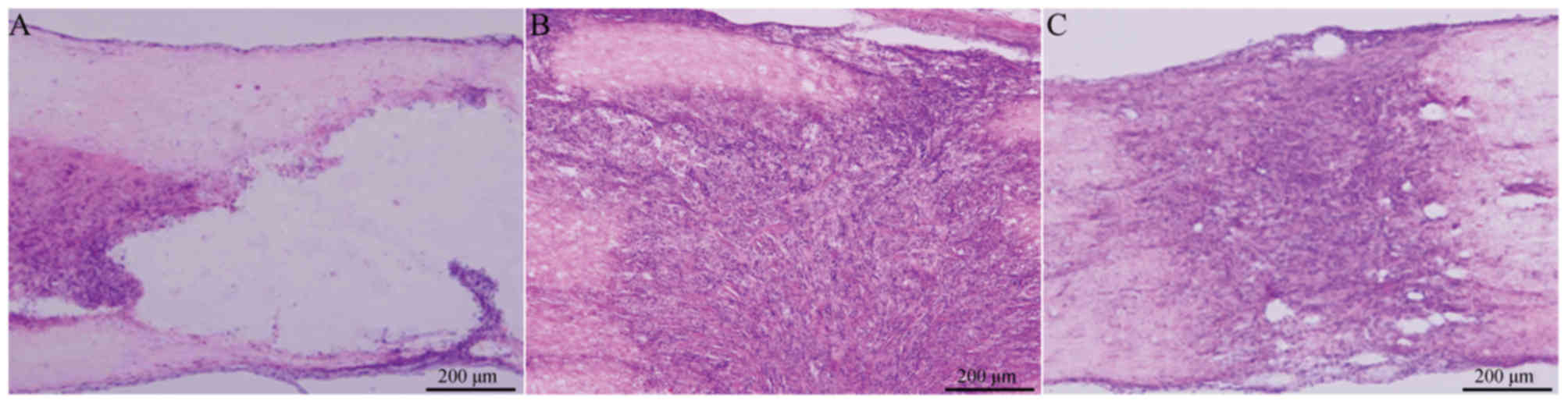
Cavity formation following spinal cord injury was
visualized by H&E staining at 2 weeks following surgery. Frozen
sections were stained with HE, observed under a light microscope
and the cavity area was calculated. The cavity area of group A was
visibly larger than the cavity area of group B or C, where
proliferating BMSCs filled the damaged region, cell reproductive
capacity was increased, and no evident cavity or necrotic cavity
was detected (Fig. 3). The areas
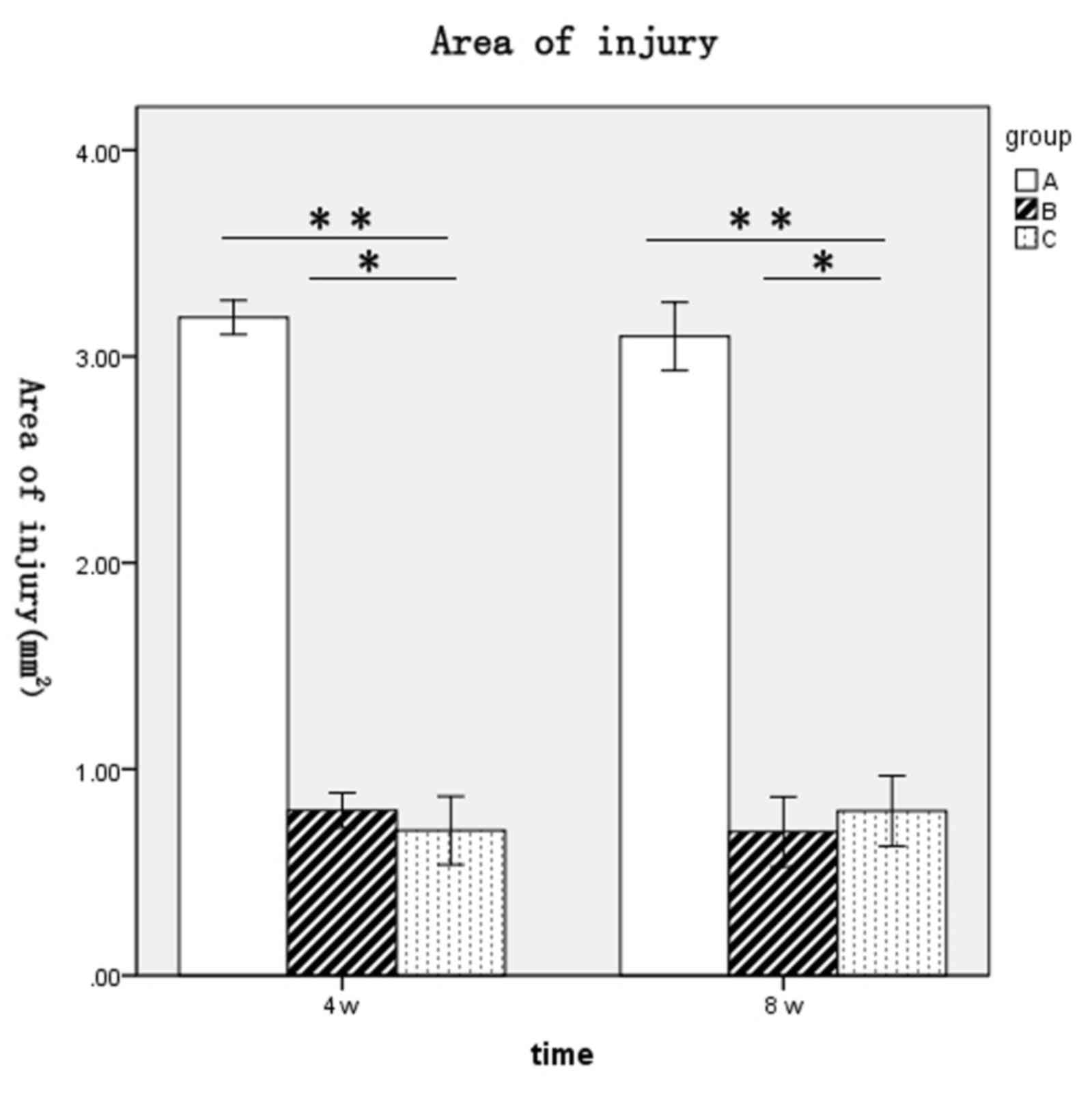
of spinal cord injury for rats in each group at 4 and 8 weeks
following surgery were as follows: Group A, 3.19±0.1 and 3.1±0.2
mm2, respectively; group B, 0.8±0.1 and 0.7±0.2
mm2, respectively; and group C, 0.7±0.2 and 0.8±0.2
mm2, respectively (Fig.
4). The spinal cord injury areas of rats in groups B and C were
significantly lower when compared with group A at 4 and 8 weeks
post-surgery (P<0.01 and P<0.01, respectively; Fig. 4). No significant difference between
groups B and C at either time point was observed (Fig. 4).
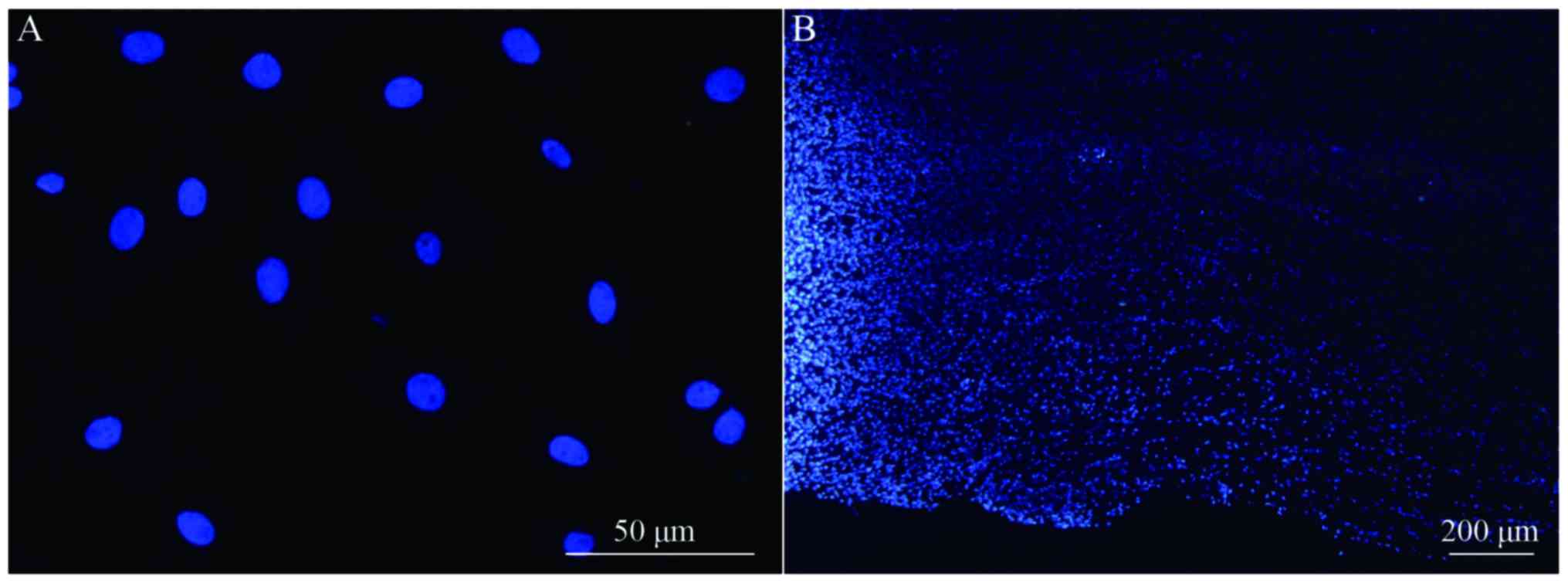
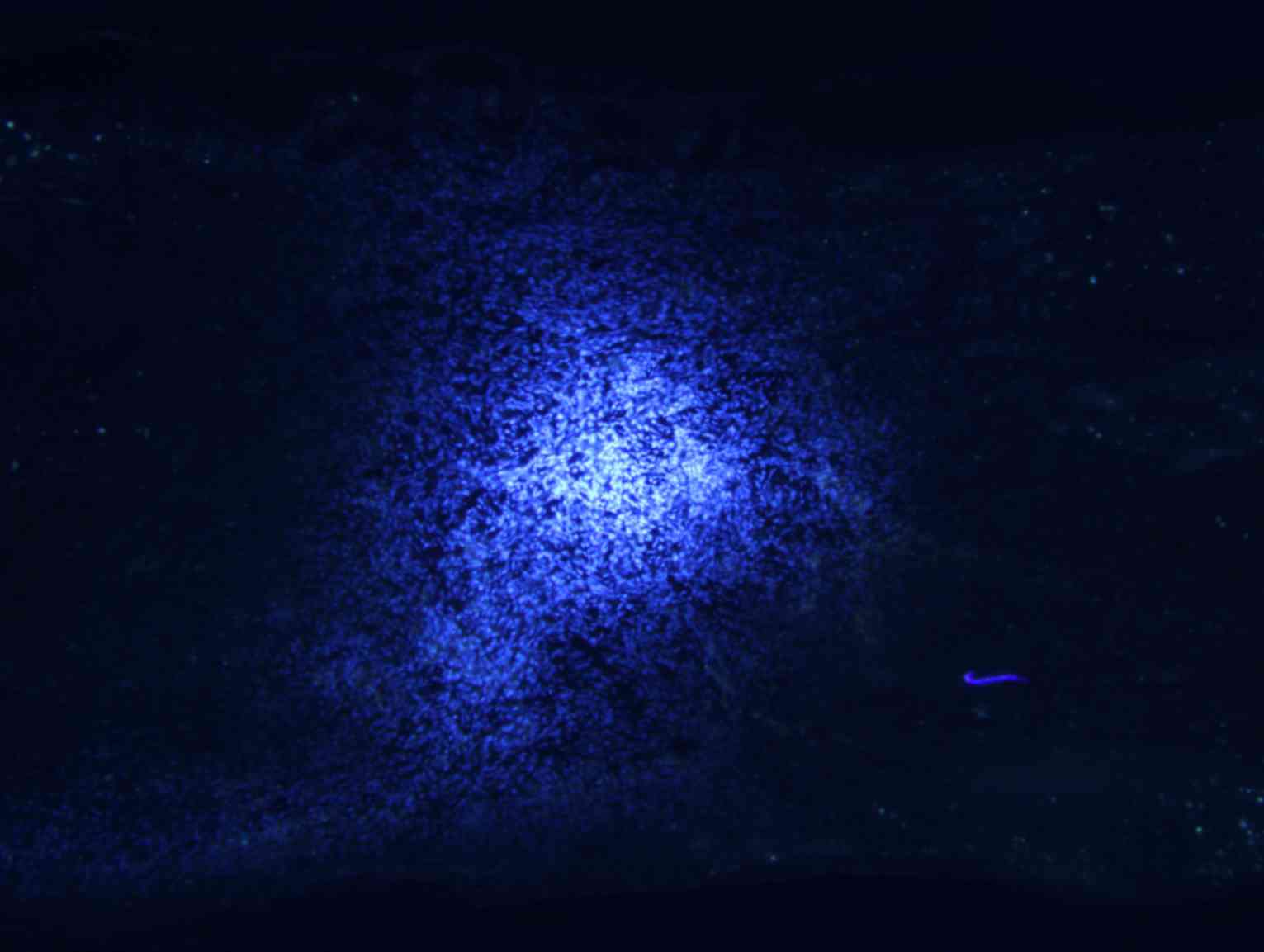
Transplanted BMSCs survive and migrate
in vivo
Longitudinal spinal cord sections from rats in
groups A, B and C were obtained at 2, 4 and 8 weeks following
surgery, and fluorescence microscopy analysis revealed a large
number of Hoechst 33342-labeled cells at the spinal cord injury
site in groups B and C. The majority of nuclei exhibited a soft
uniform blue fluorescence, indicating that transplanted BMSCs
survive in rats in vivo (Fig.
5A). The transplanted cells accumulated in the spinal cord
transplantation sites, and migrated to the spinal cord parenchyma
on both sides of the injury area (Fig.
5B).
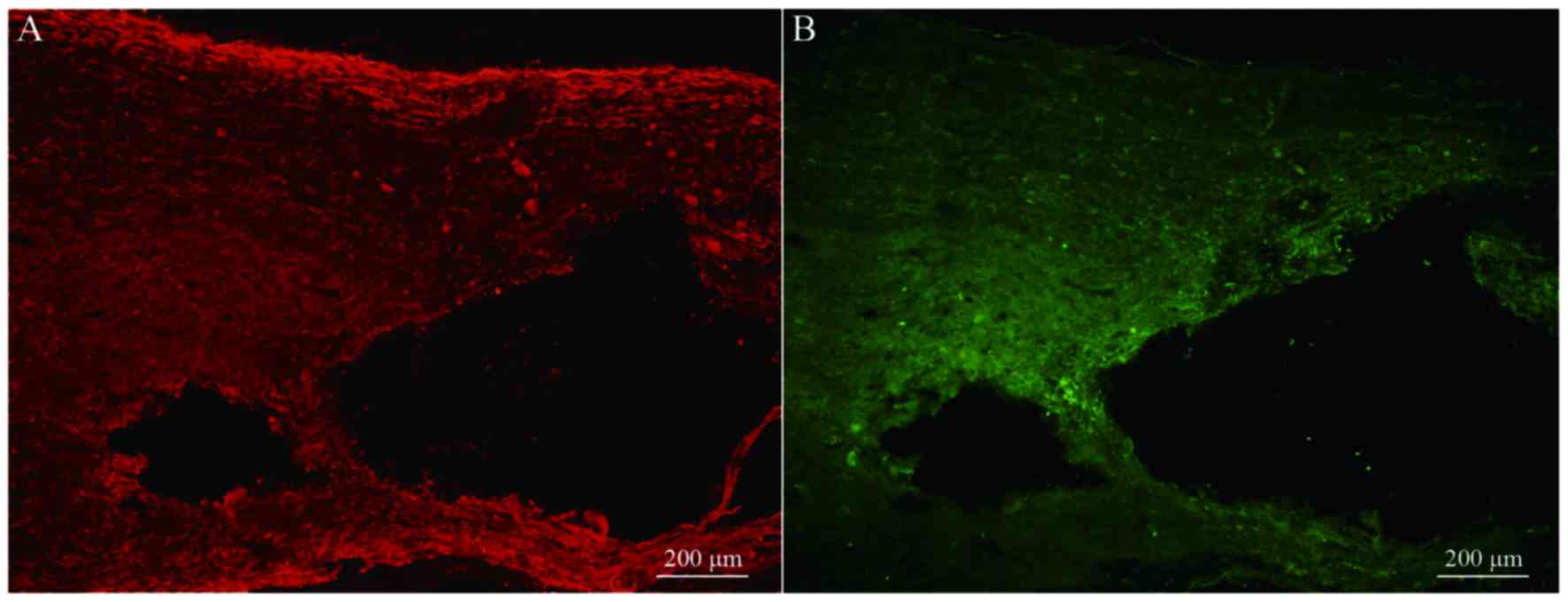
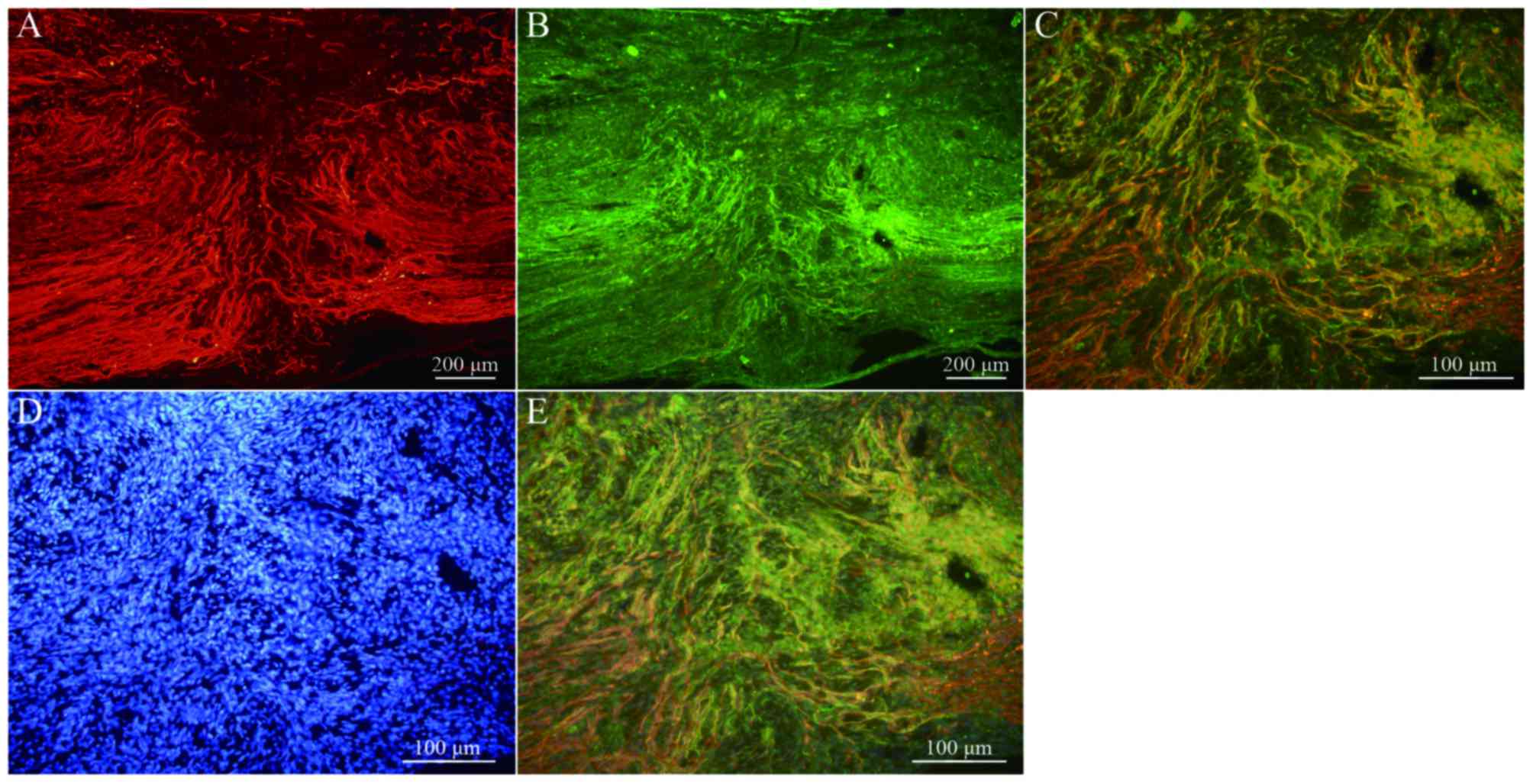
Transplanted BMSCs differentiate into
neuron-like cells in vivo
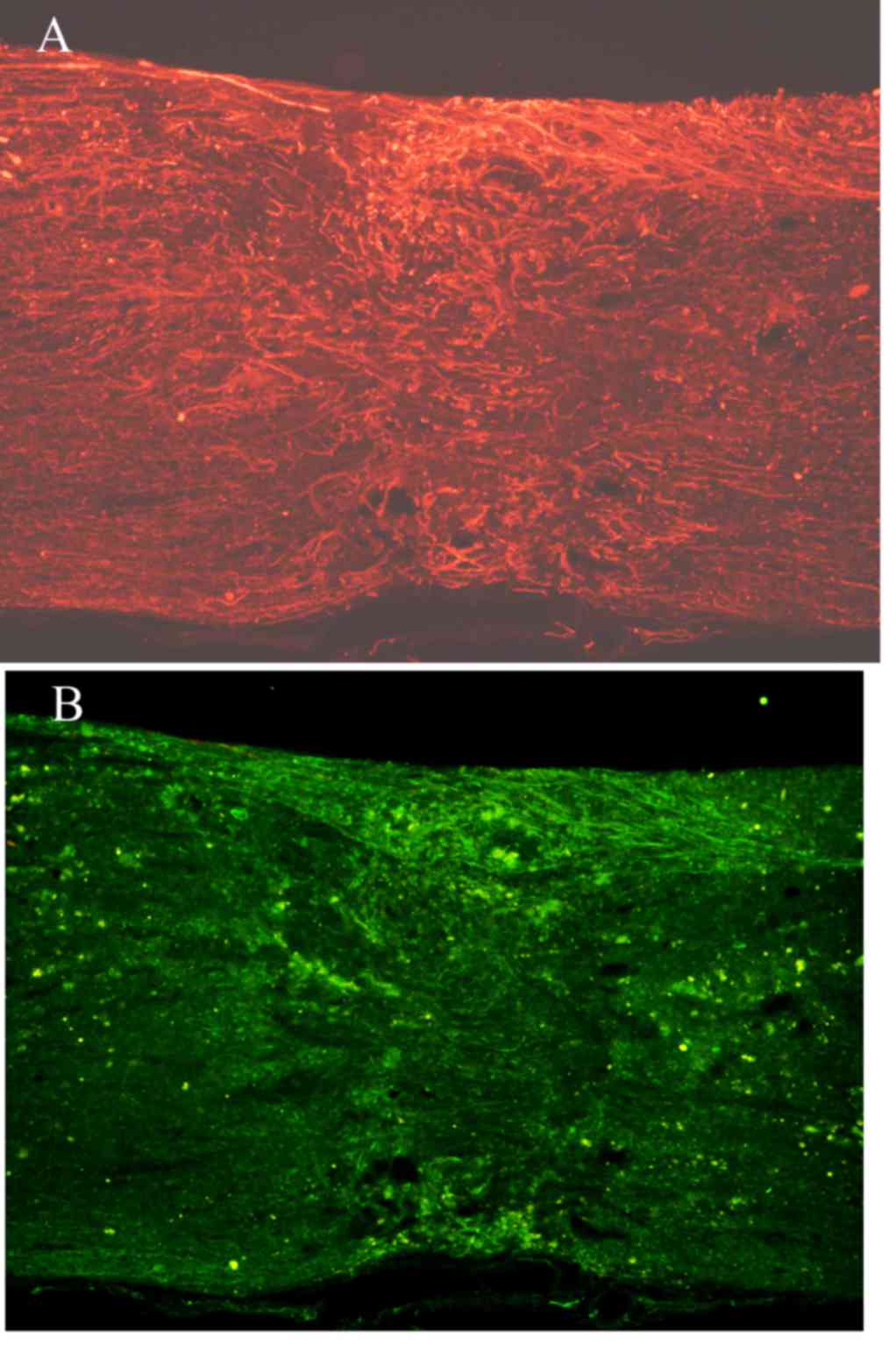
Immunofluorescence staining of NF and GAP-43 in
frozen tissue sections revealed that, while few NF or GAP-43
positive nerve fibers were observed in group A (Fig. 6A and B), continuous NF and GAP-43
immunoreactive nerve fibers in spinal cord injury sites were
observed in group C (Fig. 7A and
B), which exhibited a consistent directional pattern of
arrangement. NF and GAP-43 dual immunofluorescence results revealed
that these proteins were distributed in the damaged area (Fig. 7C). In addition, Hoechst
33342-labeled BMSC nuclei aligned with the NF and GAP-43 dual
immunofluorescence-stained neurons (Fig. 7D and E). This indicated that OEC
conditioned medium-induced BMSCs promoted the regeneration of nerve
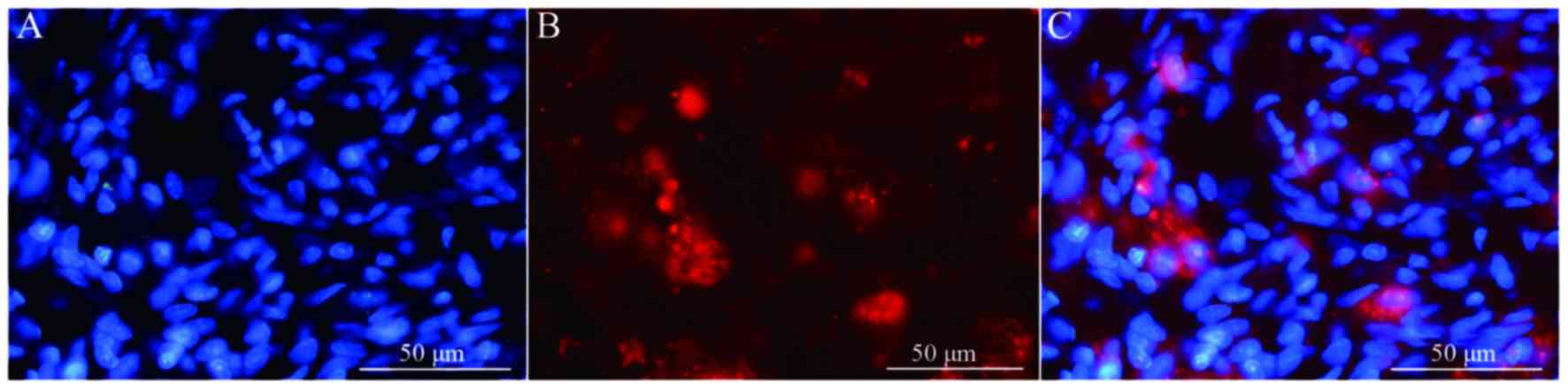
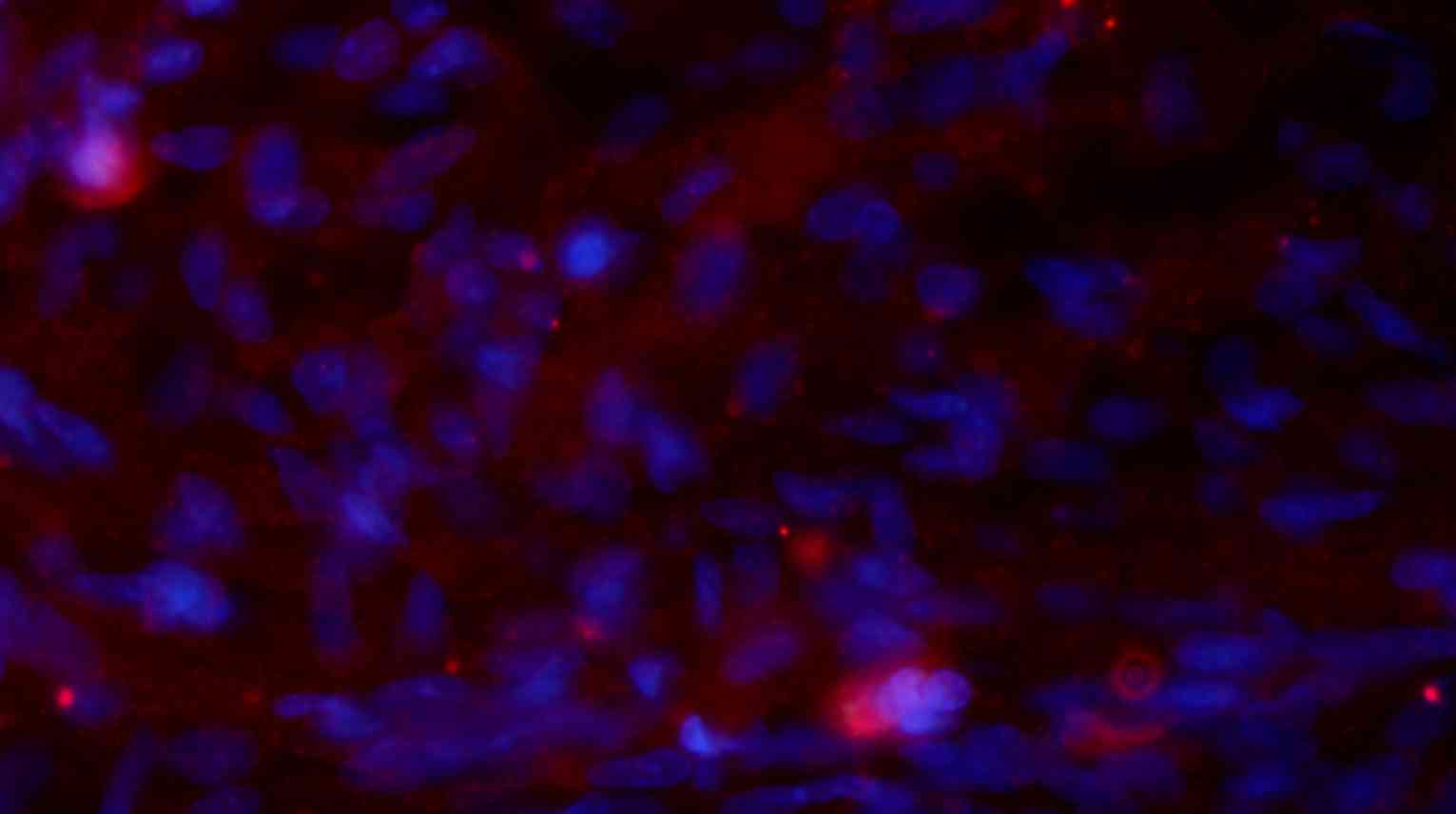
fibers. In addition, when Hoechst 33342-labeled cells were
transplanted in vivo (Fig.
8A) the expression of NeuN was partially positive (Fig. 8B and C), indicating that induced
BMSCs are able to differentiate in vivo. Fig. 9 demonstrates NF and GAP-43 staining
of the spinal cord injury area in the BMSC group 4 weeks after
injury. Fig. 10 demonstrates
surviving and migrating bone marrow stromal Hoechst 33342-labeled
cells in the spinal cord injury area of a group B rat 2 weeks after
injury and Fig. 11 is a merged
Hoechst 33342 and neuronal cell marker neuron-specific nuclear
protein images of a group B rat 2 weeks after injury.
Discussion
Complex alterations in the local microenvironment
following spinal cord injury generate challenging conditions for
efficient repair (2,7). Investigating the application of cell
transplantation for the treatment of spinal cord injury has been a
recent focus of research. In the bone marrow stromal hematopoietic
microenvironment, BMSCs are involved in supporting and regulating
the process of bone marrow hematopoiesis. As specific stem cells,
BMSCs are readily available, proliferate rapidly, exhibit weak
immunogenicity and are autologous. Notably, BMSCs demonstrate a
multi-differentiation ability (28,29).
Previous experimental studies have demonstrated that BMSCs
differentiate into bone cells, muscle cells, neurons and glial
cells under different induction conditions (30,31).
OECs are located in the olfactory bulb and the olfactory mucosa,
and possess lifetime division potentials and regeneration
characteristics. A previous study noted that OEC conditioned medium
successfully induces BMSCs in vitro to differentiate into
neural-like cells (32). In the
present study, OEC conditioned medium-induced BMSCs were
transplanted and used to treat spinal cord injury in a rat model.
OEC conditioned medium-induced BMSCs transplantation was observed
to significantly reduce cavity formation and the extent of damage
following spinal cord injury. In addition, the induced BMSCs
protected original nerve fibers of the spinal cord, promoted axonal
regeneration and expressed the NeuN neuronal marker in vivo.
Furthermore, these induced BMSCs improved lower extremity motor
function in rats, indicating that OEC conditioned medium-induced
BMSCs promoted repair of spinal cord injury in a certain context.
However, the mechanisms underlying how these nerve fibers and cells
established contact with the host in vivo, and performed
their physiological functions requires further study.
In the present study, the functional recovery of the
OEC conditioned medium-induced BMSC transplantation group was more
apparent than in the normal BMSC transplantation group. This result
may be due a number of factors. For instance, prior to
transplantation, a large number of BMSCs in the OEC conditioned
medium-induced group may be induced into neurons or nerve cells,
and a higher number of nerve cells in the spinal cord injury site
survived following transplantation, which provided an important
basis for the regeneration and repair of spinal cord injury. In
addition, prior to transplantation of OEC conditioned
medium-induced BMSCs, undifferentiated BMSCs may have been able to
proliferate rapidly following transplantation, thus reducing the
damage caused by the formation of the cavity, which protected the
host, transplanted nerve cells and nerve fibers, and provided
dependent media for axonal regeneration. Furthermore, BMSCs have
been previously demonstrated to secrete nutritional factors,
including brain-derived neurotrophic factor and vascular
endothelial growth factor, which may have protected neurons and
promoted axonal regeneration in the present study (33). Finally, some undifferentiated BMSCs
could differentiate into neurons in the spinal cord following
transplantation, due to a variety of factors mentioned above
(3). This provides an explanation
as to why the lower extremity motor function recovery of rats in
the OEC conditioned medium-induced BMSC transplantation group was
improved when compared with the non-induced BMSC transplantation
group.
In conclusion, the results of the present study
demonstrated that OEC conditioned medium-induced BMSCs survived
in vivo, reduced the formation of spinal cord injury
cavities, promoted the regeneration of nerve fibers and enabled
partial recovery of motor function in a rat model of spinal cord
injury. However, although a greater number of axons were observed
in the damaged region following transplantation of induced BMSCs,
most of these axons were unordered (Fig. 7). The present study investigated
the state of the transplanted BMSCs over the course of 8 weeks, and
did not determine the effect of induced BMSC transplantation on the
final outcome of spinal cord injury. Therefore, the present study
is a limited primary investigation and further studies are
required.
Acknowledgements
The present study was funded by the Hebei Medical
Research Topic Program (grant no. 20150508).
References
|
1
|
Bonner JF and Steward O: Repair of spinal
cord injury with neuronal relays: From fetal grafts to neural stem
cells. Brain Res. 1619:115–123. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Taoka Y and Okajima K: Spinal cord injury
in the rat. Prog Neurobiol. 56:341–358. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ankeny DP, McTigue DM and Jakeman LB: Bone
marrow transplants provide tissue protection and directional
guidance for axons after contusive spinal cord injury in rats. Exp
Neurol. 190:17–31. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Qiu XC, Jin H, Zhang RY, Ding Y, Zeng X,
Lai BQ, Ling EA, Wu JL and Zeng YS: Donor mesenchymal stem
cell-derived neural-like cells transdifferentiate into
myelin-forming cells and promote axon regeneration in rat spinal
cord transaction. Stem Cell Res Ther. 6:1052015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Collyer E, Catenaccio A, Lemaitre D, Diaz
P, Valenzuela V, Bronfman F and Court FA: Sprouting of axonal
collaterals after spinal cord injury is prevented by delayed axonal
degeneration. Exp Neurol. 261:451–461. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Plunet WT, Streijger F, Lam CK, Lee JH,
Liu J and Tetzlaff W: Dietary restriction started after spinal cord
injury improves functional recovery. Exp Neurol. 213:28–35. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Slotkin JR, Pritchard CD, Luque B, Ye J,
Layer RT, Lawrence MS, O'Shea TM, Roy RR, Zhong H, Vollenweider I,
et al: Biodegradable scaffolds promote tissue remodeling and
functional improvement in non-human primates with acute spinal cord
injury. Biomaterials. 123:63–76. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Deng LX, Walker C and Xu XM: Schwann cell
transplantation and descending propriospinal regeneration after
spinal cord injury. Brain Res. 1619:104–114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakano N, Nakai Y, Seo TB, Homma T, Yamada
Y, Ohta M, Suzuki Y, Nakatani T, Fukushima M, Hayashibe M and Ide
C: Effects of bone marrow stromal cell transplantation through CSF
on the subacute and chronic spinal cord injury in rats. PLoS One.
8:e734942013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tharion G, Indirani K, Durai M, Meenakshi
M, Devasahayam SR, Prabhav NR, Solomons C and Bhattacharji S: Motor
recovery following olfactory ensheathing cell transplantation in
rats with spinal cord injury. Neurol India. 59:566–572. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wright KT, Uchida K, Bara JJ, Roberts S,
El Masri W and Johnson WE: Spinal motor neurite outgrowth over
glial scar inhibitors is enhanced by coculture with bone marrow
stromal cells. Spine J. 14:1722–1733. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun T, Ye C, Zhang Z, Wu J and Huang H:
Cotransplantation of olfactory ensheathing cells and Schwann cells
combined with treadmill training promotes functional recovery in
rats with contused spinal cords. Cell Transplant. 22 Suppl
1:S27–S38. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tropel P, Platet N, Platel JC, Noël D,
Albrieux M, Benabid AL and Berger F: Functional neuronal
differentiation of bone marrow-derived mesenchymal stem cells. Stem
Cells. 24:2868–2876. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Z, He B, Zhang RY, Zhang K, Ding Y,
Ruan JW, Ling EA, Wu JL and Zeng YS: Electroacupuncture promotes
the differentiation of transplanted bone marrow mesenchymal stem
cells pre-induced with neurotrophin-3 and retinoic acid into
oligodendrocyte-like cells in demyelinated spinal cord of rats.
Cell Transplant. 24:1265–1281. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Akiyama Y, Radtke C and Kocsis JD:
Remyelination of the rat spinal cord by transplantation of
identified bone marrow stromal cells. J Neurosci. 22:6623–6630.
2002.PubMed/NCBI
|
|
16
|
Woodbury D, Schwarz EJ, Prockop DJ and
Black IB: Adult rat and human bone marrow stromal cells
differentiate into neurons. J Neurosci Res. 61:364–370. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Barzilay R, Melamed E and Offen D:
Introducing transcription factors to multipotent mesenchymal stem
cells: Making transdifferentiation possible. Stem Cells.
27:2509–2515. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ni WF, Yin LH, Lu J, Xu HZ, Chi YL, Wu JB
and Zhang N: In vitro neural differentiation of bone marrow stromal
cells induced by cocultured olfactory ensheathing cells. Neurosci
Lett. 475:99–103. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yui S, Fujita N, Chung CS, Morita M and
Nishimura R: Olfactory ensheathing cells (OECs) degrade neurocan in
injured spinal cord by secreting matrix metalloproteinase-2 in a
rat contusion model. Jpn J Vet Res. 62:151–162. 2014.PubMed/NCBI
|
|
20
|
Ninomiya K, Iwatsuki K, Ohnishi Y, Ohkawa
T and Yoshimine T: Intranasal delivery of bone marrow stromal cells
to spinal cord lesions. J Neurosurg Spine. 23:111–119. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Torres-Espín A, Redondo-Castro E,
Hernández J and Navarro X: Bone marrow mesenchymal stromal cells
and olfactory ensheathing cells transplantation after spinal cord
injury-a morphological and functional comparison in rats. Eur J
Neurosci. 39:1704–1717. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kastenmayer RJ, Moore RM, Bright AL,
Torres-Cruz R and Elkins WR: Select agent and toxin regulations:
Beyond the eighth edition of the guide for the care and use of
laboratory animals. J Am Assoc Lab Anim Sci. 51:333–338.
2012.PubMed/NCBI
|
|
23
|
Nash HH, Borke RC and Anders JJ: New
method of purification for establishing primary cultures of
ensheathing cells from the adult olfactory bulb. Glia. 34:81–87.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
da Silva C Balao, Macías-García B,
Rodriguez A Morillo, Bolaños JM Gallardo, Tapia JA, Aparicio IM,
Morrell JM, Rodriguez-Martínez H, Ortega-Ferrusola C and Peña FJ:
Effect of Hoechst 33342 on stallion spermatozoa incubated in KMT or
Tyrodes modified INRA96. Anim Reprod Sci. 131:165–171. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sedý J, Urdzíková L, Jendelová P and
Syková E: Methods for behavioral testing of spinal cord injured
rats. Neurosci Biobehav Rev. 32:550–580. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Howard CV and Reed MG: Unbiased
stereology: Three-dimensional measurement in microscopy. J Anat.
194:153–157. 1999.
|
|
27
|
Vawda R, Soubeyrand M, Zuccato JA and
Fehlings MG: Spinal cord injury and regeneration: A critical
evaluation of current and future therapeutic strategies. Pathobiol
Hum Dis. 8:593–638. 2014. View Article : Google Scholar
|
|
28
|
Xu Y, Xiong F, Liu L and Zhang C: Rat bone
marrow stromal cells could be induced into Schwann cell
precursor-like cells in vitro. Neurosci Lett. 488:229–233. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Robey PG, Kuznetsov SA, Ren J, Klein HG,
Sabatino M and Stroncek DF: Generation of clinical grade human bone
marrow stromal cells for use in bone regeneration. Bone. 70:87–92.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chiu LH, Lai WF, Chang SF, Wong CC, Fan
CY, Fang CL and Tsai YH: The effect of type II collagen on MSC
osteogenic differentiation and bone defect repair. Biomaterials.
35:2680–2691. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sanchez-Ramos J, Song S, Cardozo-Pelaez F,
Hazzi C, Stedeford T, Willing A, Freeman TB, Saporta S, Janssen W,
Patel N, et al: Adult bone marrow stromal cells differentiate into
neural cells in vitro original research article. Exp Neurol.
164:247–256. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ni WF, Yin LH, Lu J, Xu HZ, Chi YL, Wu JB
and Zhang N: In vitro neural differentiation of bone marrow stromal
cells induced by cocultured olfactory ensheathing cells. Neurosci
Lett. 475:99–103. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Neuhuber B, Himes B Timothy, Shumsky JS,
Gallo G and Fischer I: Axon growth and recovery of function
supported by human bone marrow stromal cells in the injured spinal
cord exhibit donor variations. Brain Res. 1035:73–85. 2005.
View Article : Google Scholar : PubMed/NCBI
|