Introduction
Stem cells reside in adult mammalian tissue, where
they maintain normal tissue homeostasis and participate in tissue
repair and regeneration in response to damage (1,2).
Mesenchymal stem cells (MSCs) are multipotent stem cells that may
be isolated from bone marrow and other tissues (3,4).
These cells exhibit the ability to differentiate into a wide
variety of tissue cell types. Furthermore, MSCs exhibit
site-specific differentiation, responding to environmental cues and
adapting their functions to diverse biomolecular contexts,
therefore, MSCs are considered a reliable cell source for stem cell
transplantation and are currently being tested in numerous ongoing
clinical trials (5,6). Previous studies have reported that
MSC function declines with age and that MSC dysfunction influences
the effects of autologous MSC transplantation in these individuals
(7–9). The authors previously demonstrated
that MSC aging may be induced by old rat serum (10). Therefore, investigation of the
factors that affect MSC aging is of primary concern.
Numerous studies have demonstrated that glucose is
an important regulator of cell senescence (11,12).
As a systemic milieu (13), blood
glucose is important in the functioning of stem cells (14). A previous study indicated that
hyperglycemia impairs bone marrow hematopoietic function and alters
the hematopoietic niche (15).
Diabetes has previously been reported to alter chemokine expression
in MSCs (16). MSCs cultured in
medium containing high glucose (HG) concentrations exhibit
premature senescence, genomic instability and telomere alterations
(17–19) however, the molecular promotive
mechanisms of HG in stem cell aging remain to be elucidated.
It has previously been demonstrated that
phosphatidylinositol 3-kinase (PI3K)/Akt and mammalian target of
rapamycin (mTOR) are associated with stem cell aging (20,21).
The preliminary study indicated that activated Akt/mTOR is an
important mediator in MSC aging (22). Notably, it has additionally been
reported that high glucose activates the PI3K/Akt signaling pathway
in podocytes (23), vascular
smooth muscle cells (24) and
vascular endothelial cells (25).
Therefore, the present study aimed to investigate if HG induces MSC
aging via the Akt/mTOR pathway. It has previously been demonstrated
that HG promotes cellular aging, however the molecular mechanisms
by which coenzyme Q10 modulates reactive oxygen species generation
and stem cell aging remain to be elucidated. The results of the
present study demonstrated that HG induced MSC senescence and
Akt/mTOR signaling mediated this effect.
Materials and methods
Isolation and culture of MSCs
Sprague-Dawley (SD) rats (weight, 10.0–15.0 g; age,
7 days; male, n=30; female, n=30; total n=60) were obtained from
Zhejiang Medical Academy of Science (Hangzhou, China; permit number
SCXK (Zhejiang) 2008–0033) and housed separately at 20–25°C under a
12-h light/dark cycle and fed ad libitum with a normal diet.
The investigation was permitted by the Law of the People's Republic
of China on the Protection of Wildlife and the protocol was
approved by the Ethical Committee of the Zhejiang University City
College (Hangzhou, China). SD rats were sacrificed by cervical
dislocation sacrificed by cervical dislocation and immersed in 75%
alcohol and disinfectant for 3 min. The rats were subsequently
transferred to a new dish and the whole skin was removed from the
hind limbs and forelimbs. The femurs and tibias were removed from
the SD rats and bone marrow was flushed out using 10 ml PBS with
100 U/ml heparin in a syringe. The cells were centrifuged at 110 ×
g at room temperature for 8 min. The cell pellet was resuspended in
2.5 ml Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (FBS; Hangzhou Sijiqing Biological Engineering
Materials Co., Ltd., Hangzhou, China) and plated in a 25
cm2 plastic flask (Corning Incorporated, Corning, NY,
USA) to allow the MSCs to adhere at 37°C with 5% CO2 in
a humidified atmosphere. Following a 3 day period, the medium was
replaced and the nonadherent cells were discarded. The medium was
completely replaced every 3 days thereafter. The cells became ~80%
confluent at 7–10 days following seeding. The adherent cells were
released from the dishes with 0.25% trypsin (Hyclone; GE Healthcare
Life Sciences, Logan, UT, USA) and seeded into fresh culture
flasks. All the experiments described below were performed using
MSCs from the third to the fifth passages.
Treatment methods of MSCs
In the control groups, MSCs were cultured for 14
days in DMEM supplemented with 10% FBS. In the glucose treatment
groups: MSCs were incubated in the culture medium containing 5.5,
11.0 or 22.0 mM glucose for 14 days. In the high glucose control
group, MSCs were treated with 22.0 mM glucose for 14 days to induce
cellular senescence. In the MK-2206 treatment group, the cells were
incubated in the culture medium containing 0.1, 1.0 or 10.0 nM
MK-2206 and 22.0 mM glucose for 14 days. The medium was completely
replaced every 3 days. The cells of all of the above groups were
cultured in a humidified incubator at 37°C and 5%
CO2.
Senescence-associated β-galactosidase
(SA-β-gal) staining
SA-β-gal staining was performed using a
Senescence-associated β-Galactosidase Staining kit (Beyotime
Institute of Biotechnology, Haimen, China) following the
manufacturer's protocol. The treatment methods for the MSCs in each
group were the same as the aforementioned. The cells were fixed in
4% (v/v) formaldehyde for 5 min and were then stained with SA-β-gal
staining solution at pH 6.0 for 12 h. The SA-β-gal-positive cells
exhibited a blue coloration. The number of positive cells were
counted under a phase-contrast microscope. The experiment was
repeated five times in each group.
Sulforhodamine B (SRB) assay
Briefly, cells were fixed with 10% trichloroacetic
acid solution for 1 h, wells were rinsed five times with tap water
and then cells were stained with 0.4% SRB solution (100 µl per
well) for 20 min at room temperature. The wells were then rinsed
with 1% acetic acid to remove unbound dye and were left to air dry.
The SRB dye was then solubilized by placing 100 µl unbuffered
Tris-based solution in each well, and the absorbance was measured
at a wavelength of 515 nm using a multiscan spectrum. The
experiment was repeated five times in each group.
Western blot analysis
To assay the expression of phosphorylated
(p)-16INK4a, p53, p21, p-mTOR, p-Akt, p-glycogen
synthases kinase (GSK)-3β and β-catenin, the total cellular protein
was extracted from MSCs from different treatment groups. The cells
were first washed in cold-buffered PBS and lysed in
radioimmunoprecipitation assay buffer (150 mM NaCl, 1% Triton
X-100, 0.5% NaDOD, 0.1% SDS and 50 mM Tris, pH 8.0). Following
centrifugation (9660 × g for 5 min) at 4°C, the protein supernatant
was transferred into different tubes. The protein concentration of
the samples was determined using a bicinchoninic acid protein assay
(Beyotime Institute of Biotechnology). A 40 µg sample of the total
protein was resolved using 8–12% SDS-PAGE and transferred onto
polyvinylidene difluoride (EMD Millipore, Billerica, MA, USA)
membranes. The membranes were blocked with 5% nonfat milk at room
temperature for 1 h in Tris-buffered saline containing Tween-20
(TBST) and incubated overnight at 4°C with the following Primary
antibodies: rabbit anti-p16INK4a (1:1,000; cat. no.
sc-1661; Santa Cruz Biotechnology, Inc., Dallas, TX, USA), mouse
anti-p53 (1:2,000; cat. no. 554147; BD Biosciences, Franklin Lakes,
NJ, USA), mouse anti-p21 (1:1,000; cat. no. sc-6246; Santa Cruz
Biotechnology, Inc.), rabbit anti-p-mTOR (1:2,000; Ser2448; cat.
no. 2971S; Cell Signaling Technology, Inc., Danvers, MA, USA),
rabbit anti-p-Akt (1:2,000; Ser473; cat. no. 4058S; Cell Signaling
Technology, Inc.), rabbit anti-p-GSK-3β (1:2,000; Ser9; cat. no.
5558; Cell Signaling Technology, Inc.), mouse anti-β-catenin
(1:1,000; cat. no. sc-7963; Santa Cruz Biotechnology, Inc.) and
mouse anti-β-actin (1:5,000; cat. no. 612657; BD Biosciences), were
incubated overnight with the membranes at 4°C. Membranes were
incubated with horseradish peroxidase (HRP)-conjugated anti-rabbit
or anti-mouse secondary antibodies (1:2,000; cat. nos. 7074 and
7076; Cell Signaling Technology, Inc.) for 1 h at room temperature,
and proteins were detected by enhanced chemiluminescence (cat. no.
RPN2106; GE Healthcare Life Sciences, Little Chalfont, UK). β-Actin
was used as the internal control to normalize the loading
materials.
Statistical analysis
All data are presented as the mean ± standard
deviation. Statistical significance was determined using one-way
analysis of variance to compare data from different experimental
groups, followed by the Student-Newman-Keuls multiple comparison
test. SPSS software, version 11.5 (SPSS, Inc., Chicago, IL, USA)
was used to process data. P<0.05 was considered to indicate a
statistically significant difference.
Results
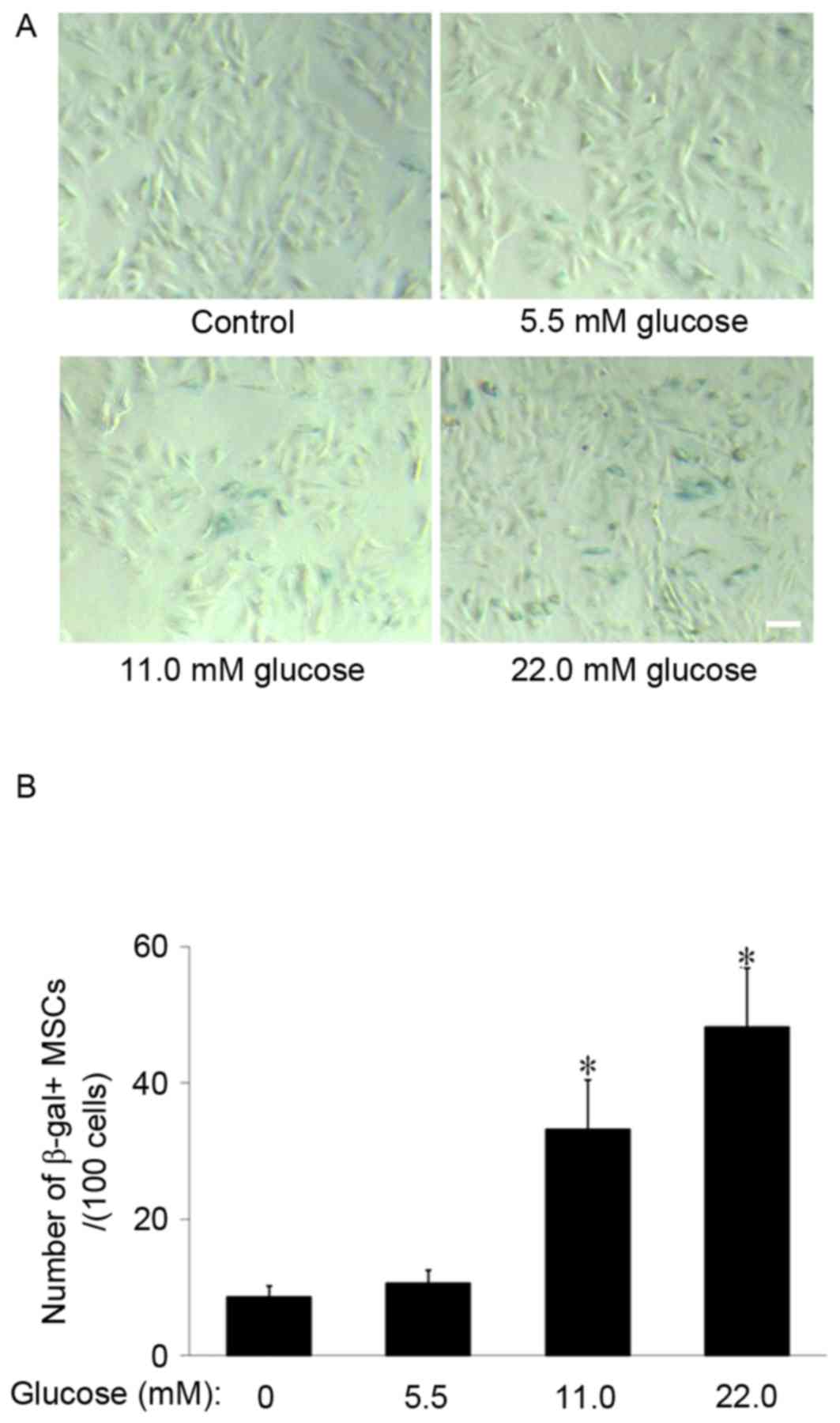
HG promotes MSCs senescence
SA-β-gal staining was used to observe the effect of
HG on MSCs aging. The results demonstrated that SA-β-gal-positive
cells appeared larger and of a flatter shape and notably, few
SA-β-gal-positive cells were observed to be present in the control
group. However, in the 11.0 and 22.0 mM glucose groups, the number
of SA-β-gal-positive cells increased (Fig. 1A). The cell counting indicated that
the number of SA-β-gal-positive cells in the 11.0 mM and 22.0 mM
glucose group (33.2±7.3 and 48.2±8.6/100 cells, respectively) were
significantly increased compared with the control group (8.6±1.7;
P<0.01; Fig. 1B), which
suggested that HG promoted MSCs senescence.
HG induces the expression of
p16INK4a, p53 and p21 in MSCs
To investigate the effects of HG on the expression
of senescence-associated proteins, the present study examined
p16INK4a, p53 and p21 expression levels via western blot
analysis. The results demonstrated that, following cell culture in
11.0 or 22.0 mM glucose for 14 days, the expression levels of
p16INK4a, p53 and p21 were increased compared with the
control group (Fig. 2). The
results suggested that p53/p21 and p16INK4a acted as
potential mediators in MSC senescence, induced by HG.
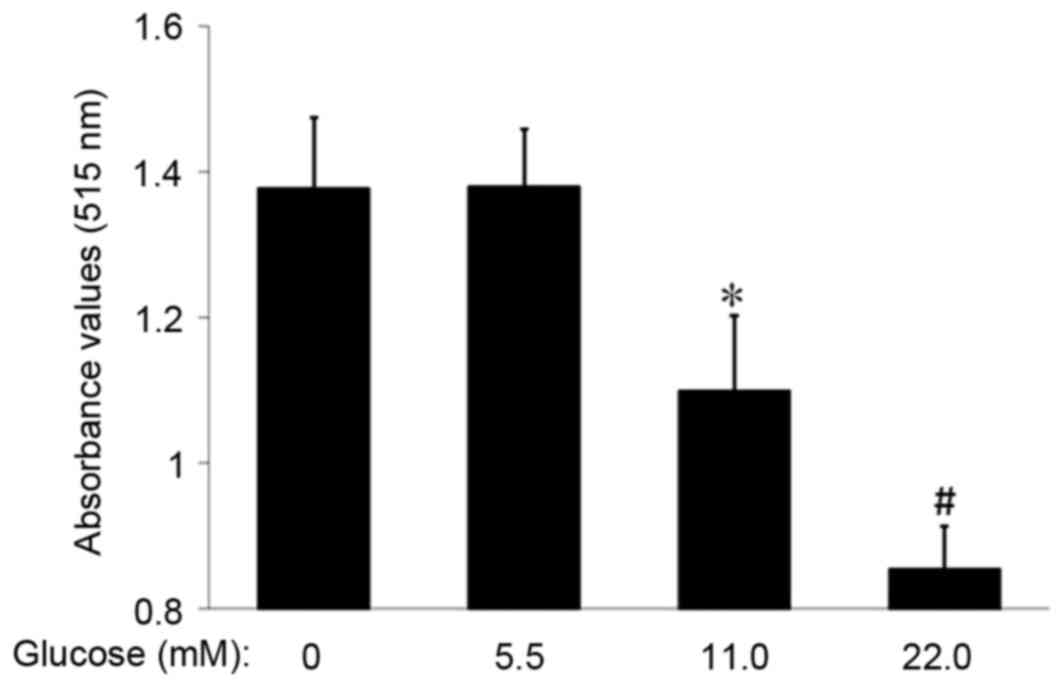
HG inhibits MSC proliferation
The proliferation rate of MSCs in each group was
examined via the SRB assay. Following MSC incubation with 11.0 or
22.0 mM glucose for 14 days, the absorbance value in the 11.0 mM
and 22.0 mM glucose groups was decreased compared with the control
(1.10±0.10 or 0.85±0.06 vs. 1.38±0.10; P<0.05 or P<0.01;
Fig. 3). These data suggested that
HG inhibited the proliferation of MSCs.
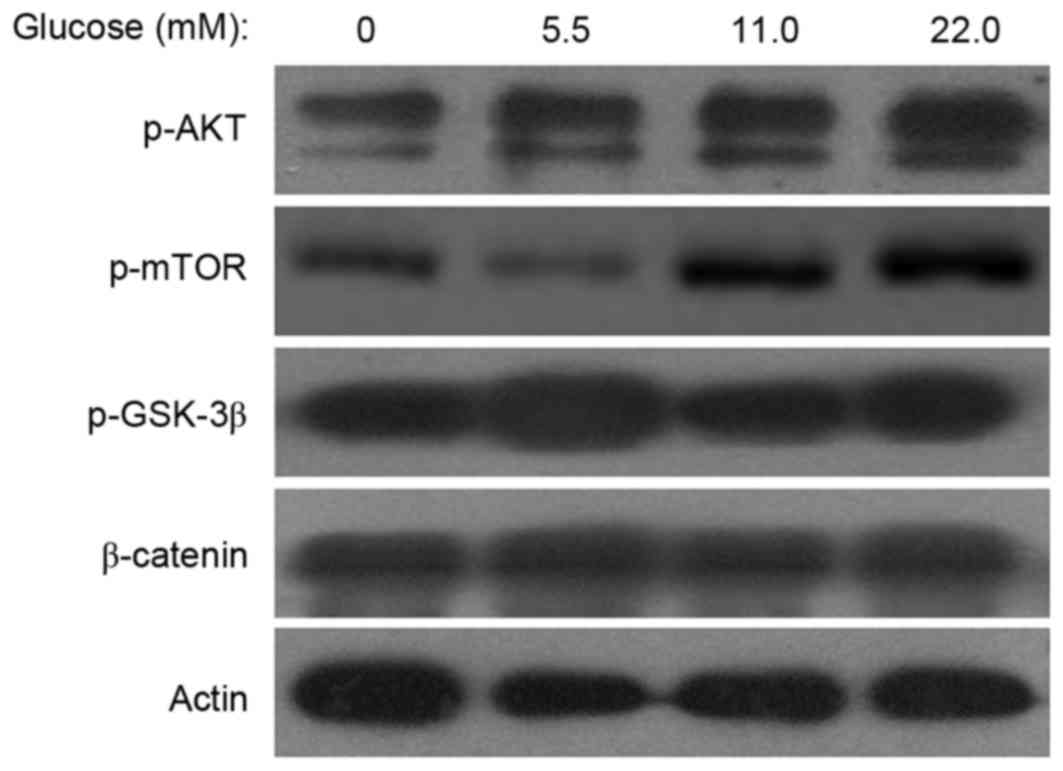
HG increases the expression of p-Akt
and p-mTOR in MSCs
To investigate the mechanisms of MSC senescence
induced by HG, the expression levels of p-mTOR, p-Akt, p-GSK-3β and
β-catenin were examined. Western blot analyses indicated that the
p-mTOR and p-Akt expression levels were increased in the 11.0 mM or
22.0 mM glucose groups compared with the control group and HG did
not affect the expression of p-GSK-3β and β-catenin (Fig. 4). These results indicated that HG
activated the Akt/m-TOR signaling pathway in MSCs.
Akt/mTOR signaling is important in MSC
senescence induced by HG
To further define the role of Akt/mTOR signaling in
MSC senescence promoted by HG, MK-2206, an Akt-specific small
molecule inhibitor, was used to inhibit the Akt/mTOR signaling.
SA-β-gal staining demonstrated that the number of SA-β-gal-positive
cells in the 0.1, 1.0 and 10.0 nM MK-2206 groups decreased in a
dose-dependent manner, compared with the HG control group (Fig. 5A). The cell count revealed that the
number of SA-β-gal-positive cells in the 1.0 and 10.0 nM MK-2206
groups (27.1±4.0 and 13.2±3.7/100 cells) was significantly
decreased compared with the glucose control group (44.3±7.7/100
cells; P<0.05 or P<0.01; Fig.
5B). Western blot analyses indicated that MK-2206 inhibited the
expression of p-Akt, and furthermore the enhancing effects of HG on
p53 and p16INK4a expression levels were reversed by
incubation with MK-2206 (Fig. 5C).
These data indicated that the Akt/mTOR signaling pathway was
important in the MSC senescence induced by HG.
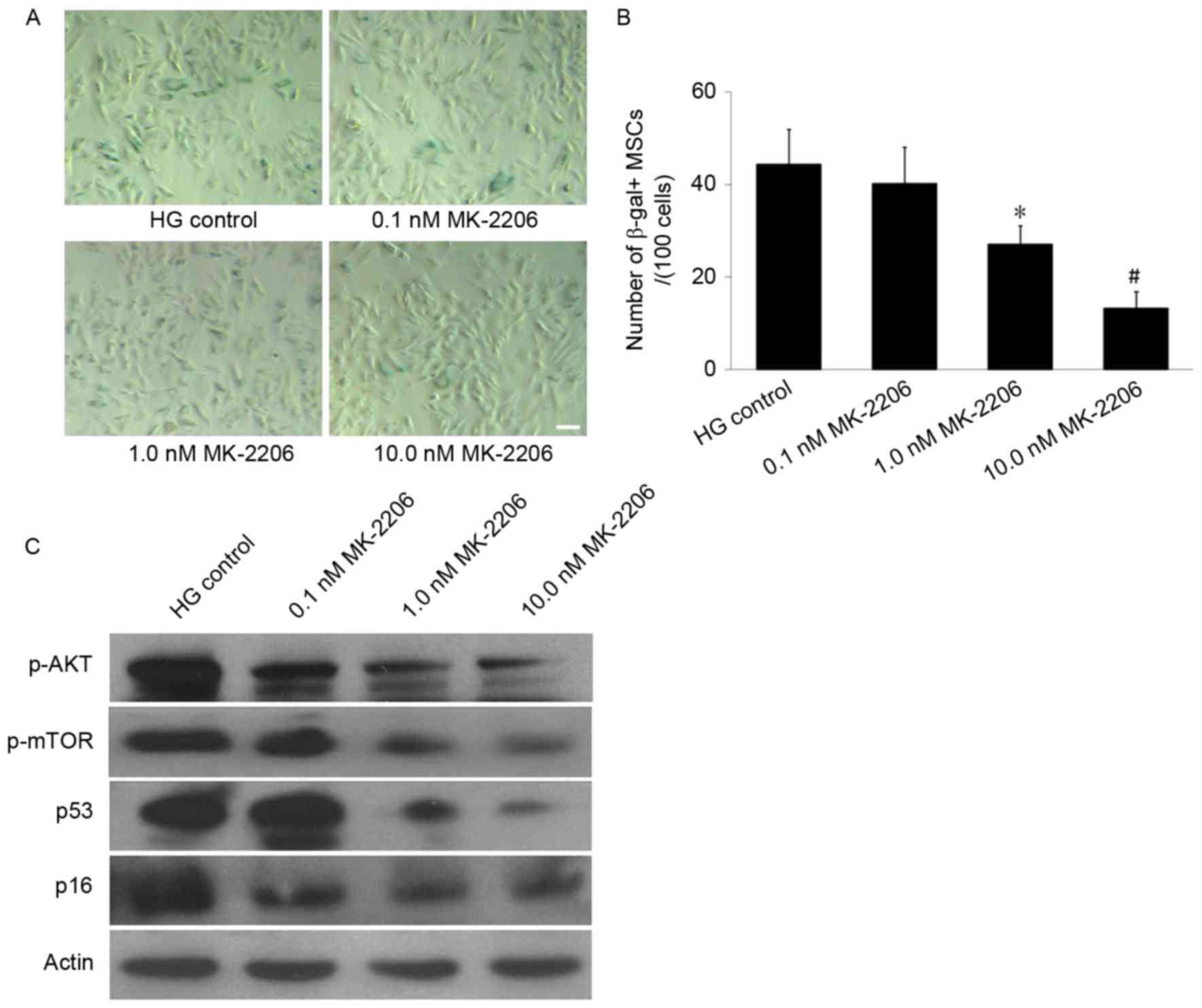 | Figure 5.Effects of differing concentrations
of MK-2206 on HG-induced MSCs senescence. (A) SA-β-gal staining. In
the 1.0 and 10.0 nM MK-2206 groups, the number of SA-β-gal-positive
cells was decreased compared with that in the HG control group.
Scale bar, 25 µm. (B) Quantification of SA-β-gal-positive cells.
The total number of SA-β-gal-positive cells among 100 random cells
was counted using phase-contrast microscopy. The results
demonstrated that the number of SA-β-gal-positive MSCs/100 cells in
the 1.0 nM and the 10.0 nM MK-2206 groups was decreased compared
with HG control group. (C) Western blot analysis. The p-Akt and
p-mTOR expression levels were decreased in the 1.0 and 10.0 nM
MK-2206 groups compared with the HG control group. Furthermore, the
p53 and p16INK4a expression was decreased in the 1.0 and
10.0 nM MK-2206 groups compared with HG control group. β-actin
served as the internal control. *P<0.05, #P<0.01
vs. HG control; n=5. HG, high glucose; SA-β-gal-positive,
senescence-associated β-galactosidase-positive; MSCs, mesenchymal
stem cells; p-, phosphorylated; Akt, protein kinase B; mTOR,
mammalian target of rapamycin signaling. |
Discussion
MSCs are of interest for potential future clinical
applications as they exhibit numerous advantages for cell therapy,
including multilineage differentiation, homing, immune modulation
and wound-healing effects (26).
However, increasing studies have demonstrated the aging of MSCs
affects their clinical application (27,28).
It is important to identify the specific factors and regulatory
mechanisms associated with MSC aging. Hyperglycemia due to diabetes
mellitus and metabolic syndrome is an increasingly occurring health
issue that may result in stem cell dysfunction (29,30).
Previous reports indicate that hyperglycemia impairs bone marrow
hematopoietic functionality and alters the hematopoietic niche
(31) and MSCs cultured in
HG-medium have been demonstrated to exhibit premature senescence
and telomere alterations (17,18,32).
However, due to the rapid rise in diabetes prevalence, further
experimental evidence is necessary to identify the association
between HG and stem cell senescence, particularly regarding MSCs.
The present study investigated the effects of HG on MSC senescence
and proliferation and the results demonstrated that HG (22.0 mM)
increased expression of p53, p21 and p16INK4a, which was
associated with development of cell senescence and inhibition MSC
proliferation.
It has previously been demonstrated that HG is a
primary factor in stem cell aging, however, the mechanisms by which
HG induces stem cell aging remain to be elucidated. It has been
previously demonstrated that the Akt/mTOR and Wnt/β-catenin
signaling pathways are important in MSC senescence (10,22,33),
and further studies indicated that the Akt/mTOR or Wnt/β-catenin
signaling pathways may be activated by HG in mesangial (34) or dendritic cells (35). Therefore, the present study
hypothesized that HG may promote MSC senescence via the Akt/mTOR or
Wnt/β-catenin signaling pathways. The results demonstrated that the
expression of p-Akt and p-mTOR were significantly increased in the
22.0 mM glucose group compared with control group, however these
effects did not appear to have been demonstrated on observation of
phosphorylated GSK-3β and β-catenin levels. To further define the
role of Akt/mTOR signaling in MSC senescence induced by HG, the
present study used MK-2206, which is an Akt-specific small molecule
inhibitor, to inhibit Akt/mTOR signaling and examine if inhibition
of Akt alone was sufficient to reverse the promotive effect of HG
on MSC senescence. The results indicated that MK-2206 significantly
decreased the total number of SA-β-gal-positive cells and the
expression of p53 and p16INK4a was additionally
decreased in the 1.0 and 10.0 nM MK-2206 groups compared with the
HG control group. These results indicated that the Akt/mTOR
signaling pathway acted as a primary mediator of the MSC senescence
induced by HG.
In conclusion, the present study demonstrated that
HG increased the number of SA-β-gal-positive MSCs and the levels of
p53, p21 and p16INK4a. A high concentration of glucose
promoted MSC aging and inhibited MSC proliferation. The Akt/mTOR
signaling pathway may act as the primary mediator of HG-induced MSC
senescence.
MSCs may be induced to form spheroid islet-like
clusters containing insulin producing cells (IPCs) (36), and MSCs from patients with type 1
and 2 diabetes may differentiate into IPCs (37). Although the high differentiation
potential of embryonic stem cells and induced pluripotent stem
cells surpasses that of MSCs, the latter have remained favorable
for transplantation studies as MSCs are considered to be easily
obtained from adult tissues, and exhibit immunodulatory and
immunosuppressive properties, in addition to nontumorigenic
differentiation potential (38).
Therefore, MSCs have emerged as a better source for the generation
of surrogate β cells (5,39). However, a number of studies have
demonstrated that the hyperglycemic state in patients with diabetes
may impair MSC function (40,41);
the results of the present study indicated that cellular senescence
may be one of the causes of this phenomenon. Further deciphering of
the specific underlying molecular mechanisms of HG involvement in
MSC aging will provide an effective intervention target for
delaying MSC aging and improving the efficacy of stem cell
transplantation in diabetes.
Acknowledgements
The present study was supported by Zhejiang
Provincial Foundation of National Science (grant nos. LY17H250001
and LY13H160030), Scientific and Technological Developing Scheme of
Hangzhou (grant nos. 20130633B33 and 20130633B34), Science Research
Foundation of Zhejiang Health Bureau (grant nos. 2013KYA151 and
2016KYB024), Traditional Chinese Medicine Science and Technology
Project of Zhejiang Province (grant no. 2016ZA024), National
College Students Innovation and Entrepreneurship Training Program
of China (grant no. 201513021011) and Zhejiang Provincial College
Students' Science and Technology Innovation Project (grant no.
2015R401190).
References
|
1
|
Schultz MB and Sinclair DA: When stem
cells grow old: Phenotypes and mechanisms of stem cell aging.
Development. 143:3–14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brunauer R and Kennedy BK: Medicine.
Progeria accelerates adult stem cell aging. Science. 348:1093–1094.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Miura Y: Guest editorial: Human
mesenchymal stromal/stem cell (MSC). Int J Hematol. 103:119–121.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Asumda FZ: Age-associated changes in the
ecological niche: Implications for mesenchymal stem cell aging.
Stem Cell Res Ther. 4:472013. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
de Miguel MP, Fuentes-Julián S,
Blázquez-Martínez A, Pascual CY, Aller MA, Arias J and
Arnalich-Montiel F: Immunosuppressive properties of mesenchymal
stem cells: Advances and applications. Curr Mol Med. 12:574–591.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ankrum J and Karp JM: Mesenchymal stem
cell therapy: Two steps forward, one step back. Trends Mol Med.
16:203–209. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Alt EU, Senst C, Murthy SN, Slakey DP,
Dupin CL, Chaffin AE, Kadowitz PJ and Izadpanah R: Aging alters
tissue resident mesenchymal stem cell properties. Stem Cell Res.
8:215–225. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ting CH, Ho PJ and Yen BL: Age-related
decreases of serum-response factor levels in human mesenchymal stem
cells are involved in skeletal muscle differentiation and
engraftment capacity. Stem Cells Dev. 23:1206–1216. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Khan M, Mohsin S, Khan SN and Riazuddin S:
Repair of senescent myocardium by mesenchymal stem cells is
dependent on the age of donor mice. J Cell Mol Med. 15:1515–1527.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang DY, Wang HJ and Tan YZ:
Wnt/β-catenin signaling induces the aging of mesenchymal stem cells
through the DNA damage response and the p53/p21 pathway. PLoS One.
6:e213972011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang E, Guo Q, Gao H, Xu R, Teng S and Wu
Y: Metformin and resveratrol inhibited high glucose-induced
metabolic memory of endothelial senescence through
SIRT1/p300/p53/p21 pathway. PLoS One. 10:e01438142015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
del Nogal M, Troyano N, Calleros L, Griera
M, Rodriguez-Puyol M, Rodriguez-Puyol D and Ruiz-Torres MP:
Hyperosmolarity induced by high glucose promotes senescence in
human glomerular mesangial cells. Int J Biochem Cell Biol.
54:98–110. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Güemes M, Rahman SA and Hussain K: What is
a normal blood glucose? Arch Dis Child. 101:569–574. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cheng NC, Hsieh TY, Lai HS and Young TH:
High glucose-induced reactive oxygen species generation promotes
stemness in human adipose-derived stem cells. Cytotherapy.
18:371–383. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ferraro F, Lymperi S, Méndez-Ferrer S,
Saez B, Spencer JA, Yeap BY, Masselli E, Graiani G, Prezioso L,
Rizzini EL, et al: Diabetes impairs hematopoietic stem cell
mobilization by altering niche function. Sci Transl Med.
3:104ra1012011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kočí Z, Turnovcová K, Dubský M,
Baranovičová L, Holáň V, Chudíčková M, Syková E and Kubinová S:
Characterization of human adipose tissue-derived stromal cells
isolated from diabetic patient's distal limbs with critical
ischemia. Cell Biochem Funct. 32:597–604. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stolzing A, Coleman N and Scutt A:
Glucose-induced replicative senescence in mesenchymal stem cells.
Rejuvenation Res. 9:31–35. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Parsch D, Fellenberg J, Brummendorf TH,
Eschlbeck AM and Richter W: Telomere length and telomerase activity
during expansion and differentiation of human mesenchymal stem
cells and chondrocytes. J Mol Med (Berl). 82:49–55. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Estrada JC, Torres Y, Benguría A, Dopazo
A, Roche E, Carrera-Quintanar L, Pérez RA, Enríquez JA, Torres R,
Ramírez JC, et al: Human mesenchymal stem cell-replicative
senescence and oxidative stress are closely linked to aneuploidy.
Cell Death Dis. 4:e6912013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu S, Liu S, Wang X, Zhou J, Cao Y, Wang
F and Duan E: The PI3K-Akt pathway inhibits senescence and promotes
self-renewal of human skin-derived precursors in vitro. Aging Cell.
10:661–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Iglesias-Bartolome R, Patel V, Cotrim A,
Leelahavanichkul K, Molinolo AA, Mitchell JB and Gutkind JS: mTOR
inhibition prevents epithelial stem cell senescence and protects
from radiation-induced mucositis. Cell Stem Cell. 11:401–414. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang D, Yan B, Yu S, Zhang C, Wang B,
Wang Y, Wang J, Yuan Z, Zhang L and Pan J: Coenzyme Q10 inhibits
the aging of mesenchymal stem cells induced by D-galactose through
Akt/mTOR signaling. Oxid Med Cell Longev. 2015:8672932015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang XM, Yao M, Liu SX, Hao J, Liu QJ and
Gao F: Interplay between the notch and PI3K/Akt pathways in high
glucose-induced podocyte apoptosis. Am J Physiol Renal Physiol.
306:F205–F213. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Montes DK, Brenet M, Muñoz VC, Burgos PV,
Villanueva CI, Figueroa CD and González CB: Vasopressin activates
Akt/mTOR pathway in smooth muscle cells cultured in high glucose
concentration. Biochem Biophys Res Commun. 441:923–928. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guan G, Han H, Yang Y, Jin Y, Wang X and
Liu X: Neferine prevented hyperglycemia-induced endothelial cell
apoptosis through suppressing ROS/Akt/NF-κB signal. Endocrine.
47:764–771. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wakao S, Kuroda Y, Ogura F, Shigemoto T
and Dezawa M: Regenerative effects of mesenchymal stem cells:
Contribution of muse cells, a novel pluripotent stem cell type that
resides in mesenchymal cells. Cells. 1:1045–1060. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tomé M, Sepúlveda JC, Delgado M, Andrades
JA, Campisi J, González MA and Bernad A: miR-335 correlates with
senescence/aging in human mesenchymal stem cells and inhibits their
therapeutic actions through inhibition of AP-1 activity. Stem
Cells. 32:2229–2244. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Raggi C and Berardi AC: Mesenchymal stem
cells, aging and regenerative medicine. Muscles Ligaments Tendons
J. 2:239–242. 2012.PubMed/NCBI
|
|
29
|
Rachmiel M, Cohen M, Heymen E, Lezinger M,
Inbar D, Gilat S, Bistritzer T, Leshem G, Kan-Dror E, Lahat E and
Ekstein D: Hyperglycemia is associated with simultaneous
alterations in electrical brain activity in youths with type 1
diabetes mellitus. Clin Neurophysiol. 127:1188–1195. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Galindo RJ and Wallia A: Hyperglycemia and
diabetes mellitus following organ transplantation. Curr Diab Rep.
16:142016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kojima H, Kim J and Chan L: Emerging roles
of hematopoietic cells in the pathobiology of diabetic
complications. Trends Endocrinol Metab. 25:178–187. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chang TC, Hsu MF and Wu KK: High glucose
induces bone marrow-derived mesenchymal stem cell senescence by
upregulating autophagy. PLoS One. 10:e01265372015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang DY, Pan Y, Zhang C, Yan BX, Yu SS,
Wu DL, Shi MM, Shi K, Cai XX, Zhou SS, et al: Wnt/β-catenin
signaling induces the aging of mesenchymal stem cells through
promoting the ROS production. Mol Cell Biochem. 374:13–20. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu L, Hu X, Cai GY, Lv Y, Zhuo L, Gao JJ,
Cui SY, Feng Z, Fu B and Chen XM: High glucose-induced hypertrophy
of mesangial cells is reversed by connexin43 overexpression via
PTEN/Akt/mTOR signaling. Nephrol Dial Transplant. 27:90–100. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Montani MS Gilardini, Granato M, Cuomo L,
Valia S, Di Renzo L, D'Orazi G, Faggioni A and Cirone M: High
glucose and hyperglycemic sera from type 2 diabetic patients impair
DC differentiation by inducing ROS and activating Wnt/β-catenin and
p38 MAPK. Biochim Biophys Acta. 1862:805–813. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mehrfarjam Z, Esmaeili F, Shabani L and
Ebrahimie E: Induction of pancreatic β cell gene expression in
mesenchymal stem cells. Cell Biol Int. 40:486–500. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sun Y, Chen L, Hou XG, Hou WK, Dong JJ,
Sun L, Tang KX, Wang B, Song J, Li H and Wang KX: Differentiation
of bone marrow-derived mesenchymal stem cells from diabetic
patients into insulin-producing cells in vitro. Chin Med J (Engl).
120:771–776. 2007.PubMed/NCBI
|
|
38
|
Hashemian SJ, Kouhnavard M and
Nasli-Esfahani E: Mesenchymal stem cells: Rising concerns over
their application in treatment of type one diabetes mellitus. J
Diabetes Res. 2015:6751032015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rekittke NE, Ang M, Rawat D, Khatri R and
Linn T: Regenerative therapy of type 1 diabetes mellitus: From
pancreatic islet transplantation to mesenchymal stem cells. Stem
Cells Int. 2016:37646812016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fadini GP, Sartore S, Schiavon M, Albiero
M, Baesso I, Cabrelle A, Agostini C and Avogaro A: Diabetes impairs
progenitor cell mobilisation after hindlimb ischaemia-reperfusion
injury in rats. Diabetologia. 49:3075–3084. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shin L and Peterson DA: Impaired
therapeutic capacity of autologous stem cells in a model of type 2
diabetes. Stem Cells Transl Med. 1:125–135. 2012. View Article : Google Scholar : PubMed/NCBI
|