Introduction
Thoracic aortic aneurysm (TAA) formation is the 15th
leading cause of death in patients of >65 years, and accounts
for ~20% of aortic aneurysm cases (1). A previous study (2) has revealed that both environmental
(smoking, high blood pressure, high cholesterol) and genetic (a
family history of aneurysms or other genetic syndromes) risk
factors are involved in the disease development. A common
pathogenic feature of TAA is progressive medial degeneration
characterized by degeneration of the extracellular matrix,
destruction of elastic lamina and loss of vascular smooth muscle
cells (SMCs) (3). Although the
surgical repair of lesions has become increasingly effective and
less invasive, an urgent requirement exists to determine the
underlying disease mechanism for early prevention and diversion of
treatment from surgical to medical approaches.
Previous studies have demonstrated stimulation of T
lymphocytes and significant changes in the expression of
extracellular matrix proteolytic system during TAA formation,
specifically an imbalance in the system of matrix metallopeptidases
(MMPs) and endogenous tissue inhibitors of MMPs (TIMPs) (4,5). T
lymphocytes and macrophages increased in the media and adventitia
from Marfan syndrome, familial TAA and sporadic cases, when
compared with control aortas (6).
During aneurysm formation, infiltrating inflammatory cells, for
example, macrophages, SMCs, fibroblasts and endothelial cells of
blood vessels all can produce MMPs, leading to the degradation of
extracellular matrix and remodeling of artery walls (7). Previously, high levels of MMP
expression and activity have been observed in natural and
experimentally induced aneurysms (8,9).
However, the exact mechanism behind these changes remains
elusive.
Tuberous sclerosis complex (TSC) is a genetic
disorder with pleiotropic manifestations caused by heterozygous
mutations in either TSC1 or TSC2. One of the less
investigated complications of TSC is the formation of aneurysms,
which are pathologically characterized by smooth muscle cell (SMC)
proliferation in the aortic media. In vitro SMC studies have
revealed that TSC1/TSC2-regulated mammalian target of rapamycin
complex 1 (mTORC1) signaling pathway plays a pivotal role in SMC
differentiation and proliferation (10,11).
Inhibiting mTORC1 signaling with the macrolide antibiotic,
rapamycin, promotes SMC differentiation through the activation of
the Akt pathway and the induction of contractile protein expression
(10). In contrast, the activation
of the mTORC1 pathway with TSC2 deficiency leads to SMC
proliferation and de-differentiation in vitro and in
vivo, which can be reversed with rapamycin treatment (11).
The role of mTOR, the major component of mTORC, in
the pathogenesis of human TAA has received little attention. To
explore the potential role of the mTOR signaling pathway in TAA
development, a series of animal studies were previously conducted
using a CaCl2-induced TAA rat model (12). The results provided novel evidence
indicating that the activation of the mTOR signaling pathway and
enhanced expression of proinflammatory cytokines are important in
early TAA development. Most important, CaCl2-induced TAA
formation in rats maybe inhibited by rapamycin through suppressing
the mTOR signaling pathway and cytokine expression.
Materials and methods
Animal experiments
The experimental study was conducted in accordance
with the Guide for the Care and Usage of Laboratory Animals
(National Institutes of Health, Bethesda, MD, USA, 2011) and
approved by the Committee for Animal Research of Shanghai Jiao Tong
University (Shanghai, China).
Male Sprague-Dawley rats (n=70; 8-weeks-old; 250–300
g) were kept in a specific-pathogen-free facility by Ruijin
hospital (Shanghai, China). A 12-h light/dark cycle was used and a
temperature of 22°C was maintained in the facility. Rats had free
access to food and water at all times. All rats were supplied by
Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai, China) and
were randomly allocated to TAA group (CaCl2-treated;
n=18), sham group (NaCl-treated; n=18), DMSO group (treated with
CaCl2 and DMSO; n=12) or rapamycin group (treated with
CaCl2 and rapamycin; n=12). The rat TAA model was
established with CaCl2, as previously described
(12). Briefly, all animals were
subjected to orotracheal intubation and mechanical ventilation. The
chest cavity was opened, and the descending thoracic aortic segment
was wrapped with a strip (sterilized medical cotton pad) presoaked
in 0.5 M CaCl2 solution or normal saline for 15 min. The
experimental mortality for the treatment and control groups was
~10–20% due to anesthesia side effects or surgical trauma.
For the rapamycin treatment, 2 mg/kg rapamycin (AG
Scientific, Inc., San Diego, CA, USA) in DMSO (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) or DMSO alone (vehicle) was administered
once intraperitoneally 1 day prior to the injury and continued for
2 weeks daily.
Tissue isolation
A ketamine/xylazine cocktail made with normal saline
was used for deep anesthesia prior to performing terminal tissue
collection. Specifically, 80 mg/kg ketamine (Zhejiang Jiuxu
Pharmaceutical Co., Ltd., Jinhua, China) and 10 mg/kg xylazine
(Selleck Chemicals, Houston, TX, USA) were administered as a single
intraperitoneal injection. All rats were euthanized 2 days or 2
weeks following induction. The thorax was opened beneath the
xiphoid process. The rats' right atrium was incised to provide an
outlet for blood and perfusate. The cardiovascular system was
perfused with normal saline from the left ventricle at a pressure
of 100 mmHg for 10 min, and then the entire aorta was harvested.
Only the part that was treated with the medical cotton pad
(CaCl2 or NaCl) was used in the following experiment.
Aortic tissues were stored at −80°C until protein isolation was
conducted for western blotting analysis. For histological and
immunohistochemical studies, the cardiovascular system was
subjected to perfusion with 4% formalin for 10 min, stored in 4%
formalin solution at 4°C, and then embedded in paraffin. Sections
of 5 µm thickness were cut.
Histological and immunohistochemical
studies
Formalin-fixed, paraffin-embedded sections were
deparaffinized at 56°C, washed with xylene and rehydrated using
serial concentrations of ethanol. For immunohistochemistry
staining, heat-mediated antigen retrieval (98°C for 6 min) was
performed to enhance antigen exposure prior to detection.
H2O2 (0.3%) was used to suppress endogenous
peroxidase activity to reduce background staining at room
temperature for 15 min. Normal goat serum (10%; Fuzhou Maixin
Biotech, Co., Ltd., Fuzhou, China) was used prior to immunostaining
and incubated with membranes at room temperature for 1 h to reduce
non-specific staining. Hematoxylin and eosin (H&E) staining was
performed for morphometric analysis. Sections were stained using a
modified Verhoeff-Van Gieson (VVG) stain kit (Sigma-Aldrich; Merck
KGaA) to evaluate the elastic fiber content following the
manufacturer's instructions. Elastin breaks were quantified in
triplicate by two independent investigators. The disruption of
elastic fibers was graded on a scale of 1–4 (1, normal or
disruption <25%; 2, disruption 25–49%; 3, disruption of 50–75%;
4, disruption >75% or total absence).
The antibodies and dilutions used in the present
study are listed in Table I. All
primary antibodies were diluted and incubated with slides at 4°C
overnight. Biotin-conjugated antibodies (cat. no. BA-1300; 1:200;
Vector Laboratories, Inc., Burlingame, CA, USA) were used as
secondary antibody and were incubated with membranes at room
temperature for 1–2 h. For immunohistochemistry, the positively
stained area was determined using the threshold-based digital
planimetry software (Image-Pro Plus 6.0; Media Cybernetics, Inc.,
Rockville, MD, USA), and expressed as the percentage of the
positive area in the intima-media or adventitia.
 | Table I.Antibodies and dilutions. |
Table I.
Antibodies and dilutions.
|
| Primary
antibodies |
|---|
|
|
|
|---|
| Antibody name | Cat. no. | Company | Species | Dilution |
|---|
| MMP-2 | 87809 | Cell signal | Rabbit | 1:100 |
| MMP-9 | 13667 | Cell signal | Rabbit | 1:100 |
| SM α-actin | A2547 | Sigma | Mouse | 1:200 |
| Calponin | sc70487 | Santa cruz | Mouse | 1:100 |
| OPN | ab8448 | Abcam | Rabbit | 1:200 |
| CD68 | ab125212 | Abcam | Rabbit | 1:200 |
| S100A4 | ab41532 | Abcam | Rabbit | 1:400 |
| TNF-α | 8184 | Cell signal | Rabbit | 1:200 |
| IL-6 | 12912 | Cell signal | Rabbit | 1:400 |
| IL-1β | 12703 | Cell signal | Rabbit | 1:200 |
Smooth muscle cell isolation, culture,
and treatment
SMCs were explanted from the descending thoracic
aortas, as previously described (11). SMCs were subcultured in a complete
SMC medium (Lonza Group, Basel, Switzerland) supplemented with 20%
fetal bovine serum, antibodies, L-glutamine, HEPES, sodium
pyruvate, insulin, recombinant human epidermal growth factor, and
recombinant human fibroblast growth factor) in a 37°C, 5%
CO2-humidified incubator. SMCs from passages 2–6 were used for
experiments, and all experiments were confirmed in at least three
different explants of SMCs.
SMCs were divided into different groups: group 1 was
divided into subgroups treated with different concentrations of
epidermal growth factor (EGF; 0, 1.25, 2.5 and 5.0 µg/ml). Group 2
was divided into four subgroups treated with either saline,
rapamycin (20 nmol/l), EGF (2.5 µg/ml) or rapamycin (20 nmol/l) +
EGF (2.5 µg/ml). Group 3 was divided into four subgroups treated
with either saline, rapamycin (20 nmol/l), siTSC2 (small
interfering RNA targeting TSC2), and rapamycin (20 nmol/l) +
siTSC2.
The TSC2 siRNA and control nonspecific siRNA were
synthesized by GeneChem Co., Ltd. (Shanghai, China). SMCs were
transfected with TSC2 siRNA according to the manufacturer's
instructions. In parallel, untreated cells and cells transfected
with nonspecific siRNA were used as controls. The experiments were
performed in triplicate for each experimental condition.
Western blot analysis
At each time point, the cells were harvested, washed
in ice-cold phosphate-buffered saline (PBS), and lysed with
radioimmunoprecipitation assay lysis buffer (Sigma-Aldrich; Merck
KGaA). The protein was extracted, and its concentration was
determined using the Bio-Rad protein assay. Equal cell lysates (10
µg) were separated by 10% SDS-PAGE with Tris-HCl gel (Ready Gel;
Bio-Rad Laboratories, Inc., Hercules, CA, USA) and
electrotransferred onto polyvinylidene difluoride membranes
(Immobilon-P; EMD Millipore, Billerica, MA, USA). Following
blocking in buffer (5% non-fat milk in T-PBS) for 3 h, the
membranes were incubated overnight at 4°C with a primary antibody
diluted in 5% bovine serum albumin (Sigma-Aldrich; Merck KGaA).
Western blot analysis was performed using primary antibodies
against α-tubulin (cat. no. T8203; 1:2,000; Sigma-Aldrich; Merck
KGaA), mTOR (cat. no. 2972; 1:1,000), phospho-mTOR (cat. no. 5536;
1:500), S6 (cat. no. 2317; 1:1,000), phospho-S6 (cat. no. 5364;
1:500), p70S6k (cat. no. 9202; 1:1,000) and phospho-p70S6k (cat.
no. 9206; 1:500; all from Cell Signaling Technology, Inc., Danvers,
MA, USA), smooth muscle myosin heavy chain (cat. no. ab53219;
1:1,000), osteopontin (OPN; cat. no. ab8448; 1:1,000) and S100A4
(cat. no. ab41532; 1:1,000; all from Abcam, Cambridge, MA, USA).
The membranes were probed with horseradish peroxidase-conjugated
goat anti-mouse and goat anti-rabbit secondary antibody (cat. nos.
115-035-003 and 111-035-003, respectively; Jackson ImmunoResearch
Laboratories, Inc., West Grove, PA, USA). The secondary antibody
was incubated with membranes at room temperature for 1 h, at a
dilution of 1:5,000. Signals were developed using an enhanced
chemiluminescence Western Blotting Detection kit (GE Healthcare
Life Sciences, Chalfont, UK). α-tubulin was used as a control.
Statistical analysis
The morphometric analysis was performed blinded for
the statistician. The results were statistically analyzed using
one-way analysis of variance, with the Dunnett's C post hoc test. A
two-sided probability level of P<0.05 was considered to indicate
a statistically significant difference. All analyses were performed
using the SPSS for Windows software (version, 13.0; SPSS, Inc.,
Chicago, IL, USA).
Results
Molecular alterations in rat TAA induction
Enhanced expression of proinflammatory
cytokines
A CaCl2-induced rat TAA model was
previously established to explore the potential role of mTOR
signaling pathway in the disease development (12). In the present study, male
Sprague-Dawley rats (250–300 g) underwent periarterial exposure of
thoracic aorta to either 0.5M CaCl2 or normal saline
(0.90% NaCl). Strikingly, immunohistochemical assessment of
proinflammatory cytokines [i.e., tumor necrosis factor (TNF)-α,
interleukin (IL)-6 and IL-1β] by computerized planimetry in the
aortic adventitia and aortic media revealed enhanced protein
expression as early as 2 days following TAA induction (Fig. 1A and B). These results confirmed
that the infiltration of inflammatory cells contributes to the
pathogenesis of TAA.
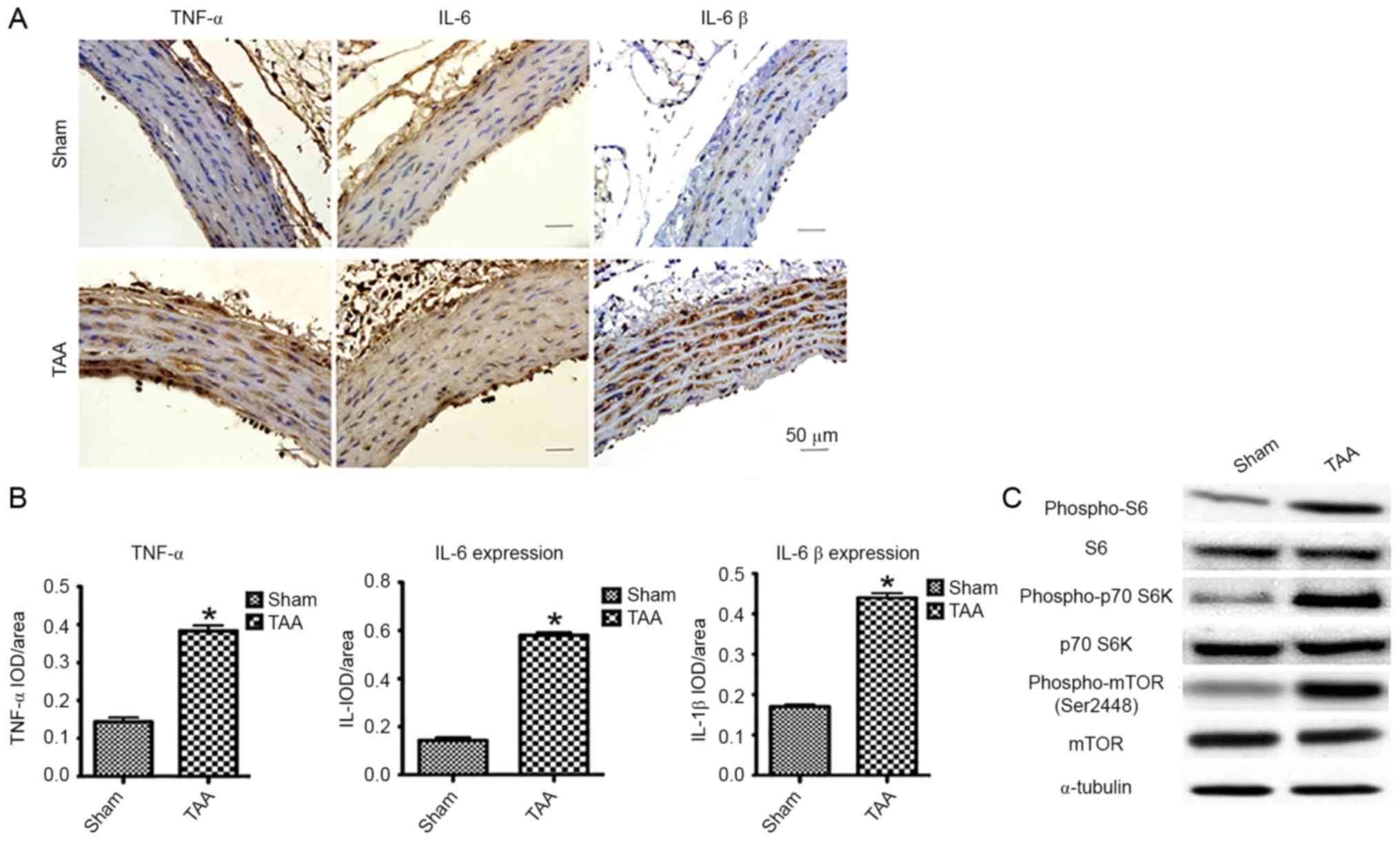 | Figure 1.The effect of proinflammatory
cytokines and mTOR signaling 2 days following TAA induction. (A)
Representative pictures of the immunohistochemical study in aortic
segments 2 days post-TAA induction. The slides were stained with
TNF-α, IL-6 and IL-1β. The anti-rabbit horseradish
peroxidase/diaminobenzidine detection system was used to visualize
the expression (brown staining). Scale bar, 50 µm. Sham:
NaCl-treated group (n=6). TAA: CaCl2-treated group
(n=6). (B) Quantitation of the protein content of TNF-α, IL-6, and
IL-1β was performed by computerized planimetry in the aortic
adventitia and aortic media in immunohistochemically stained
slides. Data are presented as the mean ± standard error of mean,
*P<0.05 vs. the Sham group. (C) Representative western blots
demonstrated the upregulation of phosphorylation of proteins
involved in the mTOR signaling pathway in the
CaCl2-treated segment of aorta as early as 2 days,
including phospho-mTOR, phospho-p70 S6K and phospho-S6. TNF-α,
tumor necrosis factor-α; IL, interleukin; TAA, thoracic aortic
aneurysm; IOD, integral optical density. |
Enhanced mTOR signaling
Besides the enhanced expression of proinflammatory
cytokines, immunoblot analyses revealed a significantly increased
phosphorylation of mTOR (Ser2448) and S6K (Thr389) as early as 2
days following TAA induction (Fig.
1C). The activation of S6K was further confirmed by the
increased phosphorylation of its substrate, S6, at both Ser240 and
Ser244. Interestingly, mTOR signal enhancement was not detected 2
weeks post-induction, suggesting a role of the mTOR signaling
pathway in the early development of TAA.
Dedifferentiation of SMCs
At 2 weeks post-induction, H&E staining was
performed for measuring the aortic diameter in each group,
presenting aneurysmal alteration in the CaCl2-treated
segments (Fig. 2A). In addition,
more severely disrupted elastic fibers were observed in
CaCl2-treated segments than in
CaCl2-untreated segments by VVG staining (Fig. 2A).
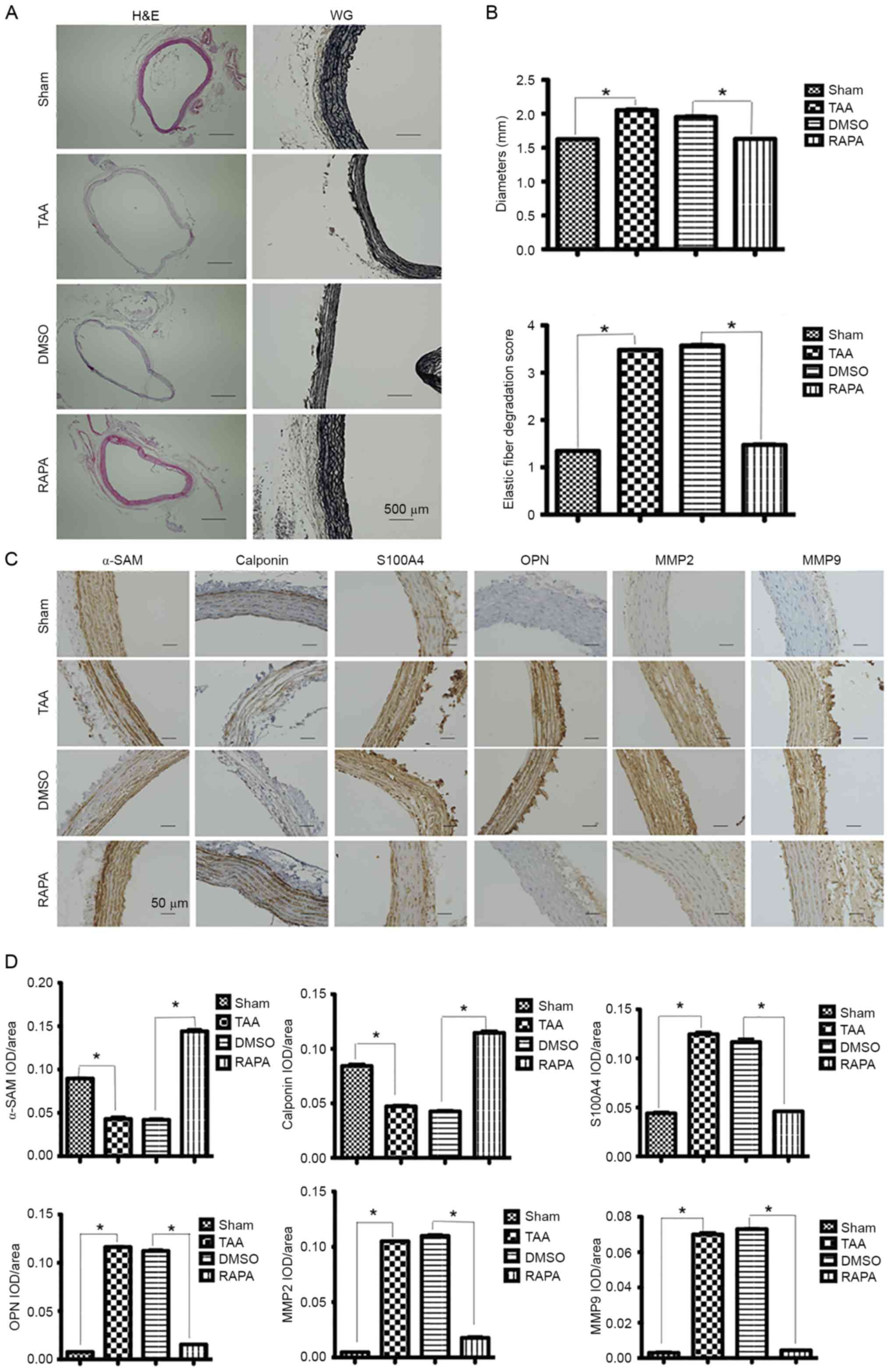 | Figure 2.Expression of contractile proteins
and dedifferentiation markers in aortic segments 2 weeks
post-rapamycin treatment. (A) Representative lower-power
micrographs of the hematoxylin and eosin and Verhoeff-Van Gieson
staining of NaCl-treated (sham group, n=6) and
CaCl2-treated aortas (TAA group, n=6) pretreated with
rapamycin (RAPA group, n=8) and DMSO (DMSO group, n=8). Scale bar,
500 µm. (B) Measurements of external media diameter and grading of
elastin degradation in the four groups (2 weeks post-TAA
induction). Data are presented as the mean ± standard error of
mean, *P<0.05 as indicated. (C) Representative pictures of
immunohistochemical study in aortic segments 2 weeks post-TAA
induction. The slides were stained with αSMA, calponin, S100A4,
OPN, MMP2 and MMP9. The anti-rabbit horseradish
peroxidase/diaminobenzidine detection system was used to visualize
the expression (brown staining). Scale bar, 50 µm. (D) The
quantitation of the protein content of αSMA, calponin, S100A4, OPN,
MMP2 and MMP9. The immunohistologically stained slides were
assessed by computerized planimetry in the aortic adventitia and
aortic media. Data are presented as the mean ± standard error of
mean. *P<0.05 as indicated. TAA, thoracic aortic aneurysm; RAPA,
rapamycin; MMP, matrix metallopeptidase; OPN, osteopontin; αSMA,
α-smooth muscle actin; IOD, integral optical density. |
SMCs contract in response to changes in pulse
pressures through a cyclic interaction between thin and thick
contractile filaments composed of SMC-specific isoforms of α-actin
(α-SMA) and myosin heavy chain, respectively. Heterozygous
mutations in α-SMA, encoded by the ACTA2 gene, predispose TAAs and
acute aortic dissections (13).
Therefore, the present study aimed to determine the expression of
contractile proteins including α-SMA and calponin in aortic SMCs.
Immunohistochemical staining revealed decreased expression of both
proteins in the CaCl2-treated segments. In contrast, an
increased expression of SMC dedifferentiation markers S100A4 and
osteopontin (OPN) was observed in the CaCl2-treated
segments (Fig. 2C).
Enhanced expression of MMP2 and
MMP9
MMPs are a family of zinc-dependent endopeptidases
capable of degrading different extracellular matrix components
including collagens and elastin. Under normal physiological
conditions, the enzymatic activity of MMPs is tightly controlled by
their TIMPs that always keep a balance in the MMP/TIMP system, but
in vascular pathologies (e.g., aneurysm, vasculitis,
atherosclerosis), the increased expression of MMPs has been often
observed (14–16). At 2 weeks post-induction, MMP-2
expression was detected diffusely across the entire layers of
CaCl2-treated segments, which was significantly higher
than the expression across the NaCl-treated segments (Fig. 2C). Similar changes were also
observed for MMP-9 expression (Fig.
2C).
Reversal effects of rapamycin
pretreatment
Pretreatment with rapamycin, a pharmacological
inhibitor of the mTOR signaling pathway, reversed the augmentation
of CaCl2-treated segments (Fig. 2A) in both diameters and elastic
fiber degradation. Fig. 3
demonstrated that, with rapamycin pretreatment (RAPA group), IOD
(integral optical density) values of TNF-α, IL-6 and IL-1β
significantly decreased (P<0.01) compared with those without
treatment (TAA and DMSO groups). As presented in Fig. 3A, in the RAPA and sham groups,
immunostaining-positive cells in the media layer of aorta were
arranged in line with less cytoplasm coloring. In contrast, in the
TAA and DMSO groups, a large number of immunostaining-positive
cells, in a mass, presented deep cytoplasm coloring. Staining with
a macrophage-specific antibody indicated that all aneurysm tissue
sections contained CD68+ macrophages that were
frequently present diffusely in the adventitia of the aortas from
the CaCl2-treated group at 2 weeks, but were rarely
found in the control aortas (Fig.
3A, CD68 staining). IOD values of CD68 in the RAPA group
significantly decreased (P<0.01) compared with those without
treatment (TAA and DMSO groups). The results suggested that
rapamycin pretreatment suppressed the hypersecretion of TNF-α,
IL-6, IL-1β and CD68 and therefore protected the aorta against
proinflammatory stress.
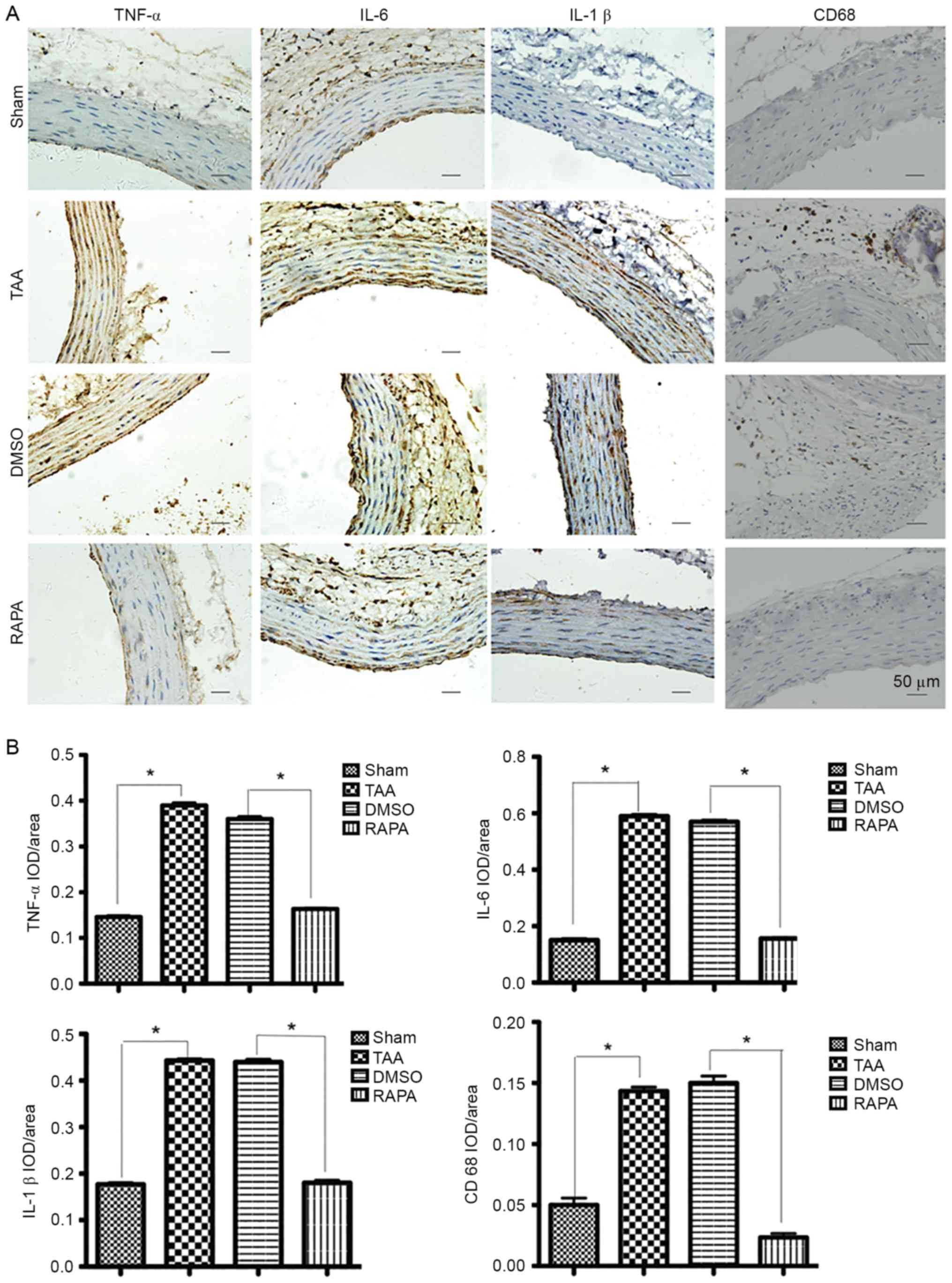 | Figure 3.Representative pictures of
immunohistochemical study in aortic segments at 2 weeks
post-rapamycin treatment. (A) The slides were stained with TNF-α,
IL-6, IL-1β and CD68. The anti-rabbit horseradish
peroxidase/diaminobenzidine detection system was used to visualize
the expression (brown staining). Scale bar, 50 µm. (Sham group,
n=6. TAA group, n=6. RAPA group, n=8. DMSO group, n=8). (B) The
quantitation of the protein content of TNF-α, IL-6, IL-1β and CD68.
The immunohistologically stained slides were assessed by
computerized planimetry in the aortic adventitia and aortic media.
Data are presented as the mean ± standard error of mean. *P<0.05
as indicated. RAPA, rapamycin; TNF, tumor necrosis factor; IL,
interleukin; IOD, integral optical density. |
Immunohistochemical assays of α-actin and calponin
contractile proteins revealed that the expression of both proteins
progressively increased following treatment with rapamycin
(Fig. 2C). In contrast, following
treatment with rapamycin, a significant reduction in expression was
observed for dedifferentiation markers S100A4 and OPN (Fig. 2C).
As described previously, in the
CaCl2-treated segments, MMP-2 significantly increased in
the entire layers of aorta, while the NaCl-treated segments
presented only mild staining (Fig.
2C). MMP-9 protein was also demonstrated at a peak level in the
adventitia and elastic lamellae 2 weeks post-TAA induction.
Following treatment with rapamycin, a significant reduction in
MMP-2 and MMP-9 was observed (Fig.
2C).
Further evidence revealed by in vitro
cell culture studies
To further explore the mechanism underlying the
rapamycin-associated inhibition of TAA formation, rat aortic SMCs
were isolated to conduct the in vitro cell culture assay.
mTOR signaling was induced by treating cells with different
concentrations of EGF (0, 1.25, 2.5 and 5.0 µg/ml). The EGF
treatment of SMCs was indicated to activate mTOR signaling and led
to an increased expression of dedifferentiation marker OPN
(Fig. 4A). However, pretreating
aortic SMCs with rapamycin inhibited EGF-induced mTOR activation,
leading to decreased expression of OPN and S100A4 (Fig. 4B).
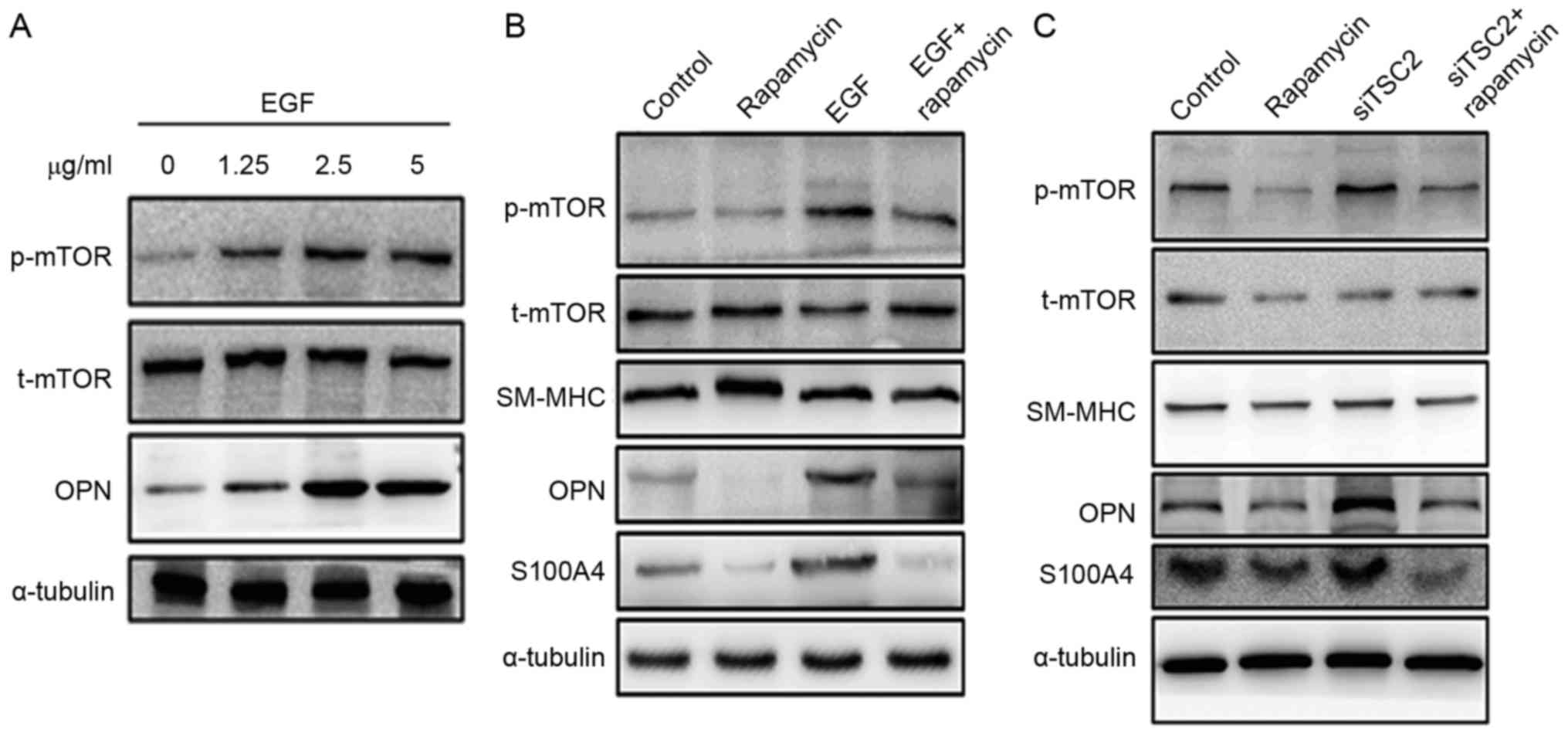 | Figure 4.Expression of the change in mTOR,
αSMA and OPN/S100A4 in aortic SMCs. (A) SMCs explanted from the
aorta of rats were cultured at EGF concentrations of 0, 1.25, 2.5
and 5 µg/ml for 72 h. Western blot analysis demonstrated that EGF
induced mTOR signaling in a dose-dependent manner. In addition, OPN
expression increased. (B) Rapamycin treatment inhibited the
expression of OPN and S100A4. Pretreating SMCs with rapamycin could
inhibit EGF-induced mTOR activation. (C) Downregulation of TSC2
expression with SiTSC2 would activate the mTOR signaling pathway.
Also, the expression of OPN and S100A4 increased when SMCs were
pretreated with siTSC2. On pretreating SMCs with rapamycin, the
activation of mTOR signaling pathway was inhibited. In the siTSC2
and rapamycin-double-treated cells, the expression levels of OPN
and S100A4 were similar to the levels observed in rapamycin-treated
cells. α-tubulin was used as a control. αSMA, α-smooth muscle
actin; OPN, osteopontin; SMC, smooth muscle cell; EGF, epidermal
growth factor; SM-MHC, smooth muscle, myosin heavy chain. |
In another study, siRNA technology was used to
downregulate TSC2 expression (11). TSC2 is a negative regulator of mTOR
signaling (11). Therefore, when
TSC2 was downregulated, the mTOR signaling pathway was activated.
An increased expression of OPN and S100A4 was observed with TSC2
downregulation. However, when the cells were pretreated with
rapamycin, the activation of the mTOR signaling pathway was
inhibited, which is consistent with the fact that mTOR is a
downstream mediator of TSC2. In the siTSC2 and
rapamycin-double-treated cells, the expression levels of OPN and
S100A4 were similar to the levels observed in rapamycin-treated
cells (Fig. 4C).
Discussion
Aortic aneurysms are classified in terms of their
anatomical location, and most commonly occur in the infrarenal
abdominal aorta and thoracic aorta. AAAs primarily affect the
elderly population and are characterized by atherosclerotic changes
with the chronic inflammation of the aortic wall (17). By contrast, TAAs affect a younger
population; the pathology present in the aortic wall of these
patients is medial degeneration, which is described as a lesion
characterized by the triad of loss of SMCs, fragmented and
diminished number of elastic fibers and increased accumulation of
proteoglycans (18). Medial
degeneration that is associated with TAAs and TADs (thoracic aortic
dissections) was originally described by Erdheim as a
noninflammatory lesion (18).
However, more recent previous evidence indicated that T lymphocytes
and macrophages were common features in the aortas of patients with
medial degeneration (19–22). Previous studies have documented an
inflammatory infiltrate in the aortic wall of patients with TAA. In
aortas of patients undergoing the prophylactic repair of TAAs, a
significant increase in the number of CD3+ and
CD68+ cells was observed throughout the aortic media and
adventitia when compared with control aortas (6). Other investigators have also
documented an inflammatory infiltrate in the aortic wall, which was
associated with the IFN-γ production (23). The present study confirmed the
aforementioned findings and further characterized the inflammatory
infiltrate in aortas of the rat TAA model. Staining with the
macrophage-specific antibody indicated that CD68+
macrophages were frequently present in the adventitia of the aortas
from the TAA model, but were rarely found in the control aortas
(Fig. 3A, CD68 staining). In
addition, the results of the present study parallel the results
obtained with a novel mouse model of TAAs, the IL-1 receptor
antagonist-deficient mouse (IL1-Ra−/−) (24). In this mouse model, inflammatory
aortitis of the ascending aorta developed in the deficient mice,
which was not present in the wild-type mice. The medial
infiltration of T lymphocytes and macrophages was observed,
although the phenotype and biological activity of invading T cells
and macrophages in TAAs should be further characterized before
drawing conclusions regarding their relevance to the disease
process (24).
Increased circulating levels of inflammatory
cytokines, such as IL-1, IL-6 and TNF-α, were identified in
patients with AAA (25–28). IL-6 is of major importance for the
aneurysmal process in humans (26). IL-1β protein levels were measured
in human TAA and control aortas; it increased ~20-fold in human
TAAs (29). Genetic deletion of
IL-1β and IL-1R significantly decreased thoracic aortic dilation in
the mice TAA model (29). In the
present TAA rat model, the enhanced expression of proinflammatory
cytokines, such as IL-6, IL-1β, and TNF-α, was observed 2 days
following TAA induction (Fig. 1A and
B).
Contractile proteins and biomarkers associated with
the differentiation status of SMCs were also examined in the
present study, along with extracellular matrix proteolytic
proteins. At 2 days post-TAA induction, the expression of
contractile proteins, including α-SMA and calponin, significantly
decreased compared with the control group (Fig. 2C and D). In contrast, the
expression of biomarkers OPN and S100A4, as well as MMP2 and MMP9,
increased. OPN is a multifunctional glycophosphoprotein with a
regulatory function in bone remodeling and in the synthesis of
collagen fibers (16). Increased
OPN levels may indicate the transition phase from the contractile
to the synthetic form of SMCs in the tunica media, which might lead
to a higher aortic diameter or aortic dissection (30). Based on these observations, OPN
might serve as an early marker for changes leading to SMC
activation in the aortic wall (16,30).
In a previous study, the S100A4/MMP-expressing cells were also
identified as CD68 positive, suggesting an inflammatory property
for the cells (9). S100A4 has been
suggested as a proinflammatory factor due to its contribution to
proliferation, inflammatory angiogenesis, and extracellular matrix
remodeling (31–33). An early observation of increased
MMP2 and MMP9 expression in the present study has presented another
line of evidence supporting a critical role of extracellular matrix
remodeling in TAA development.
So far, the role of mTOR signaling in the
pathogenesis of human TAA has remained elusive and received little
attention. In SMCs, however, mTORC1 signaling is already known to
influence SMC differentiation, and inhibiting mTORC1 signaling with
the macrolide antibiotic rapamycin promotes SMC differentiation
through the activation of the Akt pathway and the induction of
contractile protein expression (10,34).
A previous study on the Tsc2+/− mice demonstrated
that the activation of the mTORC1 pathway with Tsc2 deficiency
leads to SMC proliferation and dedifferentiation in vitro
and in vivo, which can be reversed with rapamycin treatment
(11). In the present study, the
enhanced expression of proinflammatory cytokines and mTOR signaling
were observed as early as 2 days following TAA induction. In
contrast, mTOR enhancement was not detected 2 weeks following
induction, suggesting an important role of the mTOR signaling
pathway in the early development of TAA.
The most exciting observation in the present study
is that rapamycin pretreatment was capable of effectively reversing
the formation of aortic aneurysms augmented likely by
downregulating the expression of proinflammatory molecules.
Inhibiting mTORC1 using rapamycin may also promote differentiation
through the activation of the Akt pathway and the induction of SMC
contractile protein expression (10) (as also presented in the present
study). The effectiveness of rapamycin in vivo is
illustrated by the success of rapamycin-eluting stents in
preventing stent occlusion due to SMC proliferation (35). These clinical studies and the
present data suggest a potential therapeutic use of rapamycin or
rapamycin-eluting stents in treating aortic aneurysms.
In the in vitro cell culture assay used in
the present study, mTOR signaling was induced by treating explanted
vascular SMCs with different concentrations of EGF. SMCs are not
terminally differentiated and maintain a phenotypic plasticity that
allows for transition from quiescent, differentiated cells
expressing a repertoire of proteins required for contractile
function (α-actin, calponin and β-myosin heavy chain) to
proliferating, migrating cells with the loss of contractile protein
expression and increased synthesis of extracellular matrix
proteins. SMCs are differentiated cells in a mature, functional
artery, which dedifferentiate with vascular injury and
environmental cues. In the current study, SMCs of wild-type rats
were explanted from the descending thoracic aorta. The EGF
treatment of SMCs was indicated to activate mTOR signaling and led
to the increased expression of dedifferentiation marker OPN
(Fig. 4A). Increased OPN levels
may indicate the transition from the contractile to the synthetic
form of SMCs in the tunica media, which may lead to a higher aortic
diameter or aortic dissection. Consistent with the animal study, it
was observed that pretreating aortic SMCs with rapamycin could
inhibit EGF-induced mTOR activation, leading to decreased
expression of OPN and S100A4 (Fig.
4B).
As the major effectors of the mTOR signaling
pathway, TSC1 and TSC2 are ubiquitously expressed and form
heterodimers that inhibit the activation of mTOR signaling. The
TSC1/TSC2 complex negatively regulates mTOR through the
GTPase-activating protein activity. siRNA technology was used to
test whether TSC2 functions as a regulator of mTOR expression. When
TSC2 was downregulated, the mTOR signaling pathway was activated.
Consequently, the expression of OPN and S100A4 increased (Fig. 4C). However, when the cells were
pretreated with rapamycin, activation of the mTOR signaling pathway
was inhibited, which is consistent with the fact that mTOR is a
downstream mediator of TSC2. In the siTSC2 and rapamycin-double
treated cells, the expression levels of OPN and S100A4 were similar
to the levels observed in rapamycin-treated cells.
In summary, the current experimental evidence
indicated that the early enhanced mTOR signaling pathway and
increased proinflammatory cytokines in the early stage of TAA
development and mTOR inhibitor rapamycin can inhibit
CaCl2-induced TAA formation. This effect appears to be
because of the decrease in the proinflammatory factors.
Acknowledgements
The present study was supported by the grant from
the National Natural Science Foundation of China (grant nos.
30971211, and 81170284 to Y.C. and 81570226 to J.C.).
References
|
1
|
National Center for Injury Prevention and
Control: WISQARS Leading Causes of Death Reports. 1999–2006.
|
|
2
|
Coady MA, Rizzo JA, Goldstein LJ and
Elefteriades JA: Natural history, pathogenesis, and etiology of
thoracic aortic aneurysms and dissections. Cardiol Clin.
17:615–635. vii:1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El-Hamamsy I and Yacoub MH: Cellular and
molecular mechanisms of thoracic aortic aneurysms. Nat Rev Cardiol.
6:771–786. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Barbour JR, Spinale FG and Ikonomidis JS:
Proteinase systems and thoracic aortic aneurysm progression. J Surg
Res. 139:292–307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ikonomidis JS, Jones JA, Barbour JR,
Stroud RE, Clark LL, Kaplan BS, Zeeshan A, Bavaria JE, Gorman JH
III, Spinale FG and Gorman RC: Expression of matrix
metalloproteinases and endogenous inhibitors within ascending
aortic aneurysms of patients with bicuspid or tricuspid aortic
valves. J Thorac Cardiovasc Surg. 133:1028–1036. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
He R, Guo DC, Sun W, Papke CL, Duraisamy
S, Estrera AL, Safi HJ, Ahn C, Buja LM, Arnett FC, et al:
Characterization of the inflammatory cells in ascending thoracic
aortic aneurysms in patients with Marfan syndrome, familial
thoracic aortic aneurysms, and sporadic aneurysms. J Thorac
Cardiovasc Surg. 136:922–929, 929.e1. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang X, Shen YH and LeMaire SA: Thoracic
aortic dissection: Are matrix metalloproteinases involved?
Vascular. 17:147–157. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jones JA, Barbour JR, Lowry AS, Bouges S,
Beck C, McClister DM Jr, Mukherjee R and Ikonomidis JS:
Spatiotemporal expression and localization of matrix
metalloproteinas-9 in a murine model of thoracic aortic aneurysm. J
Vasc Surg. 44:1314–1321. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cao J, Geng L, Wu Q, Wang W, Chen Q, Lu L,
Shen W and Chen Y: Spatiotemporal expression of matrix
metalloproteinases (MMPs) is regulated by the
Ca2+-signal transducer S100A4 in the pathogenesis of
thoracic aortic aneurysm. PLoS One. 8:e700572013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Martin KA, Merenick BL, Ding M, Fetalvero
KM, Rzucidlo EM, Kozul CD, Brown DJ, Chiu HY, Shyu M, Drapeau BL,
et al: Rapamycin promotes vascular smooth muscle cell
differentiation through insulin receptor
substrate-1/phosphatidylinositol 3-kinase/Akt2 feedback signaling.
J Biol Chem. 282:36112–36120. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cao J, Gong L, Guo DC, Mietzsch U, Kuang
SQ, Kwartler CS, Safi H, Estrera A, Gambello MJ and Milewicz DM:
Thoracic aortic disease in tuberous sclerosis complex: Molecular
pathogenesis and potential therapies in Tsc2+/− mice.
Hum Mol Genet. 19:1908–1920. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Geng L, Wang W, Chen Y, Cao J, Lu L, Chen
Q, He R and Shen W: Elevation of ADAM10, ADAM17, MMP-2 and MMP-9
expression with media degeneration features
CaCl2-induced thoracic aortic aneurysm in a rat model.
Exp Mol Pathol. 89:72–81. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guo DC, Pannu H, Tran-Fadulu V, Papke CL,
Yu RK, Avidan N, Bourgeois S, Estrera AL, Safi HJ, Sparks E, et al:
Mutations in smooth muscle alpha-actin (ACTA2) lead to thoracic
aortic aneurysms and dissections. Nat Genet. 39:1488–1493. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dodd T, Jadhav R, Wiggins L, Stewart J,
Smith E, Russell JC and Rocic P: MMPs 2 and 9 are essential for
coronary collateral growth and are prominently regulated by p38
MAPK. J Mol Cell Cardiol. 51:1015–1025. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dabek J, Glogowska-Ligus J and Szadorska
B: Transcription activity of MMP-2 and MMP-9 metalloproteinase
genes and their tissue inhibitor (TIMP-2) in acute coronary
syndrome patients. J Postgrad Med. 59:115–120. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huusko T, Salonurmi T, Taskinen P,
Liinamaa J, Juvonen T, Pääkkö P, Savolainen M and Kakko S: Elevated
messenger RNA expression and plasma protein levels of osteopontin
and matrix metalloproteinase types 2 and 9 in patients with
ascending aortic aneurysms. J Thorac Cardiovasc Surg.
145:1117–1123. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nordon IM, Hinchliffe RJ, Loftus IM and
Thompson MM: Pathophysiology and epidemiology of abdominal aortic
aneurysms. Nat Rev Cardiol. 8:92–102. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Erdheim J: Medionecrosis aortae
idiopathica cystica. Virchows Arch (Pathol Anat). 276:187–229.
1930. View Article : Google Scholar
|
|
19
|
Girardi LN and Coselli JS: Inflammatory
aneurysm of the ascending aorta and aortic arch. Ann Thorac Surg.
64:251–253. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Biddinger A, Rocklin M, Coselli J and
Milewicz DM: Familial thoracic aortic dilatations and dissections:
A case control study. J Vasc Surg. 25:506–511. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Roth M, Lemke P, Bohle RM, Klovekorn WP
and Bauer EP: Inflammatory aneurysm of the ascending thoracic
aorta. J Thorac Cardiovasc Surg. 123:822–824. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
He R, Guo DC, Estrera AL, Safi HJ, Huynh
TT, Yin Z, Cao SN, Lin J, Kurian T, Buja LM, et al:
Characterization of the inflammatory and apoptotic cells in the
aortas of patients with ascending thoracic aortic aneurysms and
dissections. J Thorac Cardiovasc Surg. 131:671–678. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tang PC, Yakimov AO, Teesdale MA, Coady
MA, Dardik A, Elefteriades JA and Tellides G: Transmural
inflammation by interferon-gamma-producing T cells correlates with
outward vascular remodeling and intimal expansion of ascending
thoracic aortic aneurysms. FASEB J. 19:1528–1530. 2005.PubMed/NCBI
|
|
24
|
Shepherd J and Nicklin MJ: Elastic-vessel
arteritis in interleukin-1 receptor antagonist-deficient mice
involves effector Th1 cells and requires interleukin-1 receptor.
Circulation. 111:3135–3140. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Davis VA, Persidskaia RN, Baca-Regen LM,
Fiotti N, Halloran BG and Baxter BT: Cytokine pattern in aneurysmal
and occlusive disease of the aorta. J Surg Res. 101:152–156. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jones KG, Brull DJ, Brown LC, Sian M,
Greenhalgh RM, Humphries SE and Powell JT: Interleukin-6 (IL-6) and
the prognosis of abdominal aortic aneurysms. Circulation.
103:2260–2265. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schönbeck U, Sukhova GK, Gerdes N and
Libby P: T(H)2 predominant immune responses prevail in human
abdominal aortic aneurysm. Am J Pathol. 161:499–506. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Walton LJ, Franklin IJ, Bayston T, Brown
LC, Greenhalgh RM, Taylor GW and Powell JT: Inhibition of
prostaglandin E2 synthesis in abdominal aortic aneurysms:
Implications for smooth muscle cell viability, inflammatory
processes, and the expansion of abdominal aortic aneurysms.
Circulation. 100:48–54. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Johnston WF, Salmon M, Pope NH, Meher A,
Su G, Stone ML, Lu G, Owens GK, Upchurch GR Jr and Ailawadi G:
Inhibition of interleukin-1β decreases aneurysm formation and
progression in a novel model of thoracic aortic aneurysms.
Circulation. 130 11 Suppl 1:51–59. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lesauskaite V, Epistolato MC, Castagnini
M, Urbonavicius S and Tanganelli P: Expression of matrix
metalloproteinases, their tissue inhibitors, and osteopontin in the
wall of thoracic and abdominal aortas with dilatative pathology.
Hum Pathol. 37:1076–1084. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Boye K and Maelandsmo GM: S100A4 and
metastasis: A small actor playing many roles. Am J Pathol.
176:528–535. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Senolt L, Grigorian M, Lukanidin E, Simmen
B, Michel BA, Pavelka K, Gay RE, Gay S and Neidhart M: S100A4 is
expressed at site of invasion in rheumatoid arthritis synovium and
modulates production of matrix metalloproteinases. Ann Rheum Dis.
65:1645–1648. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Oslejsková L, Grigorian M, Gay S, Neidhart
M and Senolt L: The metastasis associated protein S100A4: A
potential novel link to inflammation and consequent aggressive
behaviour of rheumatoid arthritis synovial fibroblasts. Ann Rheum
Dis. 67:1499–1504. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Martin KA, Rzucidlo EM, Merenick BL,
Fingar DC, Brown DJ, Wagner RJ and Powell RJ: The mTOR/p70 S6K1
pathway regulates vascular smooth muscle cell differentiation. Am J
Physiol Cell Physiol. 286:C507–C517. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Holmes DR Jr, Leon MB, Moses JW, Popma JJ,
Cutlip D, Fitzgerald PJ, Brown C, Fischell T, Wong SC, Midei M, et
al: Analysis of 1-year clinical outcomes in the SIRIUS trial: a
randomized trial of a sirolimus-eluting stent versus a standard
stent in patients at high risk for coronary restenosis.
Circulation. 109:634–640. 2004. View Article : Google Scholar : PubMed/NCBI
|














