Introduction
Lower back pain (LBP) is a common disorder that
induces activity limitation (1).
Degenerative disc disease (DDD) is a common cause of LBP (2). Numerous factors have been reported to
be associated with the pathogenesis of DDD, including mechanical
stress (3), cellular senescence
(4) and extracellular matrix
degradation (5). Among these
mechanisms, the decreased transport of nutrition and waste products
appears to be the most important (6,7). The
intervertebral disc (IVD) is the largest avascular tissue in the
human body, and the metabolic exchange of a mature IVD is largely
dependent on diffusion through the cartilage endplate (CEP)
(8). The CEP is a thin horizontal
layer of hyaline cartilage that separates adjacent vertebrae from
the IVD. The blood vessels in the vertebral bones do not invade the
discs, ending at the interface between the IVD and the vertebral
body (9). As the most important
channel for metabolic exchange, the degeneration of the CEP is
hypothesized to be predominantly responsible for the initiation of
DDD (10).
CEP degeneration is characterized by ossification,
rather than chondrification (11).
Chondrification is important for the physiological functioning of
the CEP, whereas ossification is harmful to the ability of the
cartilage to resist compressive forces and disrupts the transport
properties of the CEP (12).
However, the mechanisms underlying ossification in the CEP remain
unclear.
A previous study demonstrated the presence of
CEP-derived stem cells (CESCs), which may undergo osteogenic and
chondrogenic differentiation (13). The differentiation characteristics
were notable, since the osteogenic and chondrogenic differentiation
fates of CESCs may be responsible for the balance between
ossification and chondrification in the CEP.
Due to its avascular nature, the microenvironment
surrounding the IVD is exposed to hypoxia (14). Hypoxia may influence the
osteogenesis of mesenchymal stem cells (MSCs) (15), indicating that the physiological
hypoxic microenvironment may regulate the osteogenesis of CESCs,
thereby regulating the ossification of the CEP.
During research into the mechanisms through which
hypoxia regulates CESC osteogenesis, it was hypothesized that
alternative splicing (AS) may serve a ‘bridging’ role between
hypoxia and CESC osteogenesis. The hypothesis may be attributed to
numerous observations. Hypoxia may initiate various alternative
splicing (AS) events. For example, the inhibitory Per/Arnt/Sim
domain protein undergoes unique AS under hypoxic conditions to
produce variants in exons 3 and 6, which may establish a novel
negative feedback modulation of the adaptive responses to
hypoxia/ischemia (16).
Furthermore, hypoxia stimulates the generation of the neurotrophin
tyrosine kinase receptor type 1 (TrkA) AS variant TrkAIII, which
tends to form a stress-resistant phenotype that protects against
hypoxia (17). The regulatory
roles of AS in stem cell osteogenic differentiation have also been
reported. For example, the expression of a TATA binding
protein-associated factor 4 (TAF4) variant that does not include
exons 6 and 7 in the hTAF4-TAFH domain may promote the early
osteogenic differentiation of MSCs (18). In addition, the parathyroid
hormone-related protein phenotype becomes selectively increased
during the osteogenic differentiation of MSCs, indicating its
potential to be a molecular marker of stem cell fate (19).
High-throughput screening technology is a powerful
tool that may be used to study the role of gene expression and AS
on a genome-wide scale. For example, an exon microarray was
previously used to investigate the role of hypoxia on the gene
expression profiles and AS events in human umbilical vein
endothelial cells (20). In
addition, microarray technology was used to identify a
hypoxia-associated alternatively spliced laminin-A3 variant that
was correlated with poor prognosis, which indicated a decreased
probability of survival of 59 patients with head and neck cancer
(21).
The present study aimed to investigate the
transcriptional and AS mechanisms during CESC osteogenic
differentiation under normoxic and hypoxic conditions. The CESCs
were isolated and induced to undergo osteogenic differentiation
under normoxic and hypoxic conditions. The samples were extracted
and analyzed using the Human Transcriptome Array 2.0 (HTA 2.0)
system. An analysis of the gene expression profiles and AS events
between the two groups was performed on a genome-wide scale. The
Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes
(KEGG) web-based tools were used to analyze the significantly
enriched biological processes, molecular functions, cellular
components and related signaling pathways of the differentially
expressed genes (DEGs) and alternatively spliced genes (ASGs). To
the best of our knowledge, genome-wide studies regarding the
regulatory role of hypoxia on the transcription and AS events
associated with stem cell osteogenic differentiation have not
previously been performed; therefore, the present study may be
helpful to elucidate the mechanisms underlying the
hypoxia-regulated osteogenic differentiation of CESCs, which is
beneficial for increasing understanding of the mechanisms
underlying CEP ossification.
Materials and methods
Ethics statement
The CEP samples used in the present study were
obtained from patients who experienced disc herniation accompanied
by spondylolisthesis and underwent discectomy procedures at the
Xinqiao Hospital, Third Military Medical University (Chongqing,
China) between January 2014 and June 2014 (Table I). The study procedures were
approved by the Ethics Committee of Xinqiao Hospital, Third
Military Medical University and were performed in accordance with
the Declaration of Helsinki; written informed consent was obtained
from each patient.
 | Table I.Patient information. |
Table I.
Patient information.
| Case no. | Gender | Age, years | Diagnosis | Degenerated disc
level | Surgery type |
|---|
| 1 | Male | 50 |
Spondylolisthesis | L4-L5 | TLIF |
| 2 | Male | 55 |
Spondylolisthesis | L4-L5 | TLIF |
| 3 | Female | 52 |
Spondylolisthesis | L4-L5 | TLIF |
Tissue procurement
The adherent tissue (nucleus pulposus and annulus
fibrosus) was carefully removed from the surgically obtained CEPs
under a sterilized microscope until a thin layer of cartilage
remained, which was washed with sterile 0.1 M PBS. Subsequent to
being mechanically homogenized, small portions of the CEP tissue
were randomly selected for hematoxylin and eosin staining to
eliminate the possibility of pollution from any other residual
impurity.
Cell isolation
The CEP tissues were mechanically cut into pieces
and digested in Dulbecco's modified Eagle's medium (DMEM)/F12
(Hyclone; GE Healthcare Life Sciences, Logan, UT, USA) containing
0.2% collagenase II (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
and 1% fetal calf serum (FCS; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) overnight at 37°C. The suspension was
filtered through a 70-µm cell filter and centrifuged for 5 min at
110 × g at room temperature. Following aspiration of the
supernatant, the pellet was resuspended in DMEM/F12 containing 10%
FCS and 1% penicillin-streptomycin. The cells were transferred to a
25-cm2 cell culture flask and cultured at 37°C and 5%
CO2.
Agarose culture
Following the first passage, the cells were
recultured in an agarose selection solution, which was established
as previously described (13). The
culture dishes (Costar; Corning Incorporated, Corning, NY, USA)
were precoated with a 1% low melting point agarose solution. A
mixture containing 0.5 ml DMEM/F12, 0.5 ml 2% low melting point
agarose solution and 1 ml culture medium, containing ~5×104
suspended CEP cells, was added to the culture dishes. The culture
dishes were transferred to a 37°C humidified incubator containing
5% CO2. The culture medium was changed twice a week.
Following 6 weeks of culturing, the cell aggregates (diameter,
>50 µm) were transferred to a 25-cm2 cell culture flask using a
sterile Pasteur pipette and were cultured in a 37°C humidified
incubator containing 5% CO2. The agarose was gradually
absorbed into the culture medium and the cells grew as adherent
cultures. Cells within passage 3 were used in the present
study.
Induction and oxygen deprivation
For osteogenic differentiation, the cells were
induced in osteogenic induction medium (OIM; HUXMA-90021; Cyagen
Biosciences, Inc., Guangzhou, China) under hypoxic conditions (1%
O2) and normoxic conditions (21% O2). The
medium was changed twice a week over a period of 3 weeks.
Western blotting
In order to evaluate the osteogenesis of CESCs, the
protein expression levels of runt-related transcription factor 2
(RUNX2) and collagen type I (COL1) were examined. RUNX2 is the
master regulator of osteoblast differentiation and maturation,
which is necessary for skeletogenesis (22). COL1, which is the major organic
component of bone, may affect the expression of bone cell
phenotypes (23,24). RUNX2 and COL1 have been recognized
as markers of osteogenic differentiation (25). Cell lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China) was used for the
extraction of total protein. The protein concentration was
determined using a BSA kit (Beyotime Institute of Biotechnology)
and 30 µg protein from each sample was loaded per lane. The
proteins in whole cell lysates were separated by 10% SDS-PAGE and
transferred to a polyvinylidene fluoride membrane (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The membranes were
incubated with rabbit anti-human monoclonal antibody against RUNX2
(12556; 1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA)
and mouse anti-human monoclonal antibody against COL1 (ab90395;
1:1,000; Abcam, Cambridge, UK) overnight at 4°C. Following washing
three times with PBS, the membrane was incubated with horseradish
peroxidase-conjugated horse anti-mouse (7076; 1:5,000) or goat
anti-rabbit polyclonal immunoglobulin G (7074; 1:5,000) (both from
Cell Signaling Technology, Inc.) secondary antibodies for 2 h at
room temperature. Protein expression was measured via
chemiluminescent detection using ECL western blotting regents
(Thermo Fisher Scientific, Inc.). The protein expression levels
were normalized to the rabbit antibody against β-actin (4967;
1:5,000; Cell Signaling Technology, Inc.). The protein expression
data were analyzed using Quantity One software version 4.6.2
(Bio-Rad Laboratories, Inc.).
Alizarin red and alkaline phosphatase
(ALP) staining
In order to identify mineral deposits following the
different treatments, the cells were fixed with 4% paraformaldehyde
(PFA) for 30 min at room temperature, washed 3 times with PBS,
stained with alizarin red (Cyagen Biosciences, Inc.) for 5 min at
room temperature and washed a further 3 times with PBS. For ALP
staining, the cells were fixed with 4% PFA for 30 min at room
temperature, stained with an ALP assay kit (Beyotime Institute of
Biotechnology), according to the manufacturer's protocol, and
washed 3 times with PBS. Subsequently, images of the stained cells
were captured.
Affymetrix HTA 2.0
Total RNA was extracted from the cell samples using
TRIzol extraction (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and hybridized to HTA 2.0 (Affymetrix, Inc.,
Santa Clara, CA, USA). With probes targeting exons and junctions,
HTA 2.0 is able to simultaneously analyze gene expression profiles
and AS events. The microarray was scanned by CapitalBio Corporation
(Beijing, China), according to the manufacturer's protocol. The
image signals of the microarray were saved as DAT files. The
Affymetrix GeneChip Command Console software version 3.2
(Affymetrix, Inc.) converted the DAT files (image signals) into CEL
files (digital signals). The CEL files were transformed to chp
files for quantile normalization, probe set signal integration and
background correction with a Robust Multichip Analysis algorithm
using the Affymetrix Expression Console software version 1.3
(Affymetrix, Inc.). The DEGs and ASGs in the chp files were
subsequently analyzed using the Affymetrix Transcriptome Analysis
Console software version 3.0 (Affymetrix, Inc.). Web-based tools
including the Database for Annotation, Visualization and Integrated
Discovery (david.ncifcrf.gov), KEGG (www.genome.jp/kegg), and Molecule Annotation System
(mas.capitalbiotech.online/mas3) were used to identify the
significantly enriched GO terms and signaling pathways. The
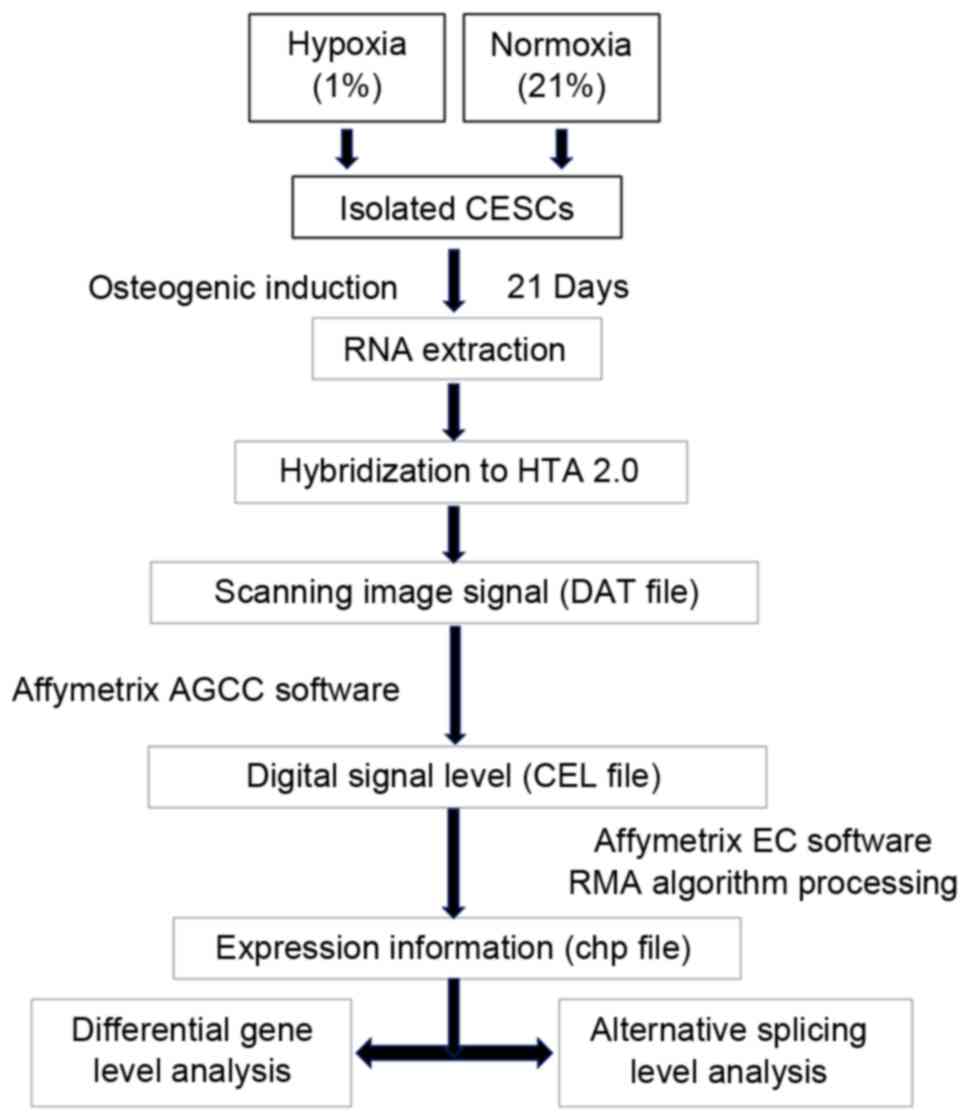
workflow of the present study is presented in Fig. 1.
Criteria for detecting DEGs and
ASGs
The data from the different samples under normoxic
conditions were used as the control level for calculating the fold
changes in gene expression. Fold changes in gene expression of ≤-2
or ≥2 and a p-value of <0.05 were considered to be the threshold
for significant DEGs. A splicing index (SI) model was used to
determine the ASGs. SI, which represents the ratio of the signal
intensity of an exon normalized to that of the target gene between
the two experimental groups, was employed to analyze the level of
exon exclusion/inclusion. The SI value was obtained using the
following formulae:
NI (i, j)A = exoni signal
intensity in condition A/genej signal intensity in
condition A
SI(X,Y)=Log2NI(X,Y)H/NI(X,Y)N
Where NI, normalized intensity; NI(i,
j)A, the signal intensity of the ith exon normalized to
that of the jth gene in the condition A; N, normoxic induction
condition; and H, hypoxic induction condition. The threshold for
ASGs was set as SI (linear) values of ≥2/≤-2 and a p-value of
<0.05.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was obtained from the cell samples using
TRIzol, according to the manufacturer's protocol (Invitrogen;
Thermo Fisher Scientific, Inc.). A total of 1 µg RNA from each
sample was transcribed into cDNA according to the manufacturer's
protocol using the PrimeScript™ RT Master Mix kit (RR047A; Takara
Bio, Inc., Otsu, Japan). A total of 1 µg cDNA from each sample was
used for qPCR with SYBR Premix Ex Taq™ II (RR820A; Takara Bio,
Inc.). The qPCR was a two step-method (without an extension step)
according to the manufacturer's protocols, and performed under the
following conditions: 95°C for 30 sec, followed by 40 cycles at
95°C for 5 sec and 60°C for 34 sec. Following each run,
dissociation curves were generated using temperatures ranging
between 60 and 95°C. The expression levels of each gene were
normalized to the levels of β-actin and analyzed using the
2−∆∆Cq method (26).
The sequences of the specific primers are presented in Table II.
| Table II.Primer sequences. |
Table II.
Primer sequences.
| A, RT-qPCR primers
for DEG validation |
|---|
|
|---|
| Gene symbol | Primer sequence
(5′-3′) |
|---|
| MMP3-F |
AGAAGTGAGCAACTGCAAAAACT |
| MMP3-R |
CTTCCCCGTCACCTCCAATC |
| AKR1C1-F |
CAATTGAAGCTGGCTTCCGC |
| AKR1C1-R |
GACCAACTCTGGTCGATGGG |
| MFGE8-F |
AACAGCATCCCTGACAAGCA |
| MFGE8-R |
GGAAGATCTGCAGCCACTGA |
| CCL2-F |
AGCAGCAAGTGTCCCAAAGA |
| CCL2-R |
GTGTCTGGGGAAAGCTAGGG |
| FGF7-F |
CAGCGTCACAGCAACTGAAC |
| FGF7-R |
TAGTGTAGTGCTCCGGGTGT |
| POSTN-F |
TCCCCGTGACTGTCTATAAGC |
| POSTN-R |
GTGACCTTGGTGACCTCTTCTT |
| GPM6B-F |
GCGAGACCTGCAAACTTGTG |
| GPM6B-R |
GTTGCTCAAGAATCGCCACG |
| IGFBP2-F |
CGAGGGCACTTGTGAGAAGC |
| IGFBP2-R |
CAGTGACCTTCTCCCGGAAC |
| VCAM1-F |
GGACCACATCTACGCTGACA |
| VCAM1-R |
TTGACTGTGATCGGCTTCCC |
| FBLN5-F |
TTGTGAGGAGTCTAGCCAGTTG |
| FBLN5-R |
TGGTTTTGCTTAGCCCTCTTCA |
| β-actin-F |
CAACCGGGAAGGAAATGAATGG |
| β-actin-R |
GCCCAATACGACCAAATCAGAG |
|
|---|
| B,
Semi-quantitative PCR primers for ASG validation |
|
|
|---|
| Gene symbol | Primer sequence
(5′-3′) |
| IFNGR1-F |
CTTTCTCCTACCCCTTGT |
| IFNGR1-R |
CCTGTGGCATGATCTGGT |
| PAX2-F |
CCTCCCCTCCTGTTTCCA |
| PAX2-R |
TGCTGGGTGAAGGTGTCA |
| BCAP29-F |
TTCTAAGGCACAAAATGA |
| BCAP29-R |
GAGGCTAACATAACAAAATT |
| POSTN-F |
AATCCCCGTGACTGTCTA |
| POSTN-R |
ATTGCTTCTTTGTGCTGA |
| FXR1-F |
ACGAAGGACTGATGAAGA |
| FXR1-R |
CTGGAGTACGCTGTAGCT |
| CACNB4-F |
GTTTTACAGCGGTTGATT |
| CACNB4-R |
TGGGGTTTGTAAGTGTCC |
| TUBD1-F |
ACTTGTACCGATCTTCAG |
| TUBD1-R |
CCAAGTTAGCAATGGAAGTGTTAAA |
| GPM6B-F |
GCGAGACCTGCAAACTTGTG |
| GPM6B-R |
GTTGCTCAAGAATCGCCACG |
| MEF2C-F |
ATCTCCGAGTTCTTATTCC |
| MEF2C-R |
TATCCTCCCATTCCTTGT |
| CADM1-F |
GAAATGCCTCAACACGCCGTAC |
| CADM1-R |
ACGACGCCACCGATCACG |
| β-actin-F |
CAACCGGGAAGGAAATGAATGG |
| β-actin-R |
GCCCAATACGACCAAATCAGAG |
ASG validation by semi-quantitative
RT-PCR
Total RNA extraction and cDNA synthesis were
performed as in the aforementioned RT-qPCR method. A total of 2 µl
cDNA from each sample was used to conduct semi-quantitative RT-PCR
with Premix Taq™ (RR901A; Takara Bio, Inc.) and specific primers
that were designed to flank the constitutively expressed exons. The
sequences of the specific primers are presented in Table II. The expression levels of each
ASG were normalized to the expression levels of β-actin. The ASGs
of interest were selected for validation according to the following
criteria: i) A whole exon skip/gain; ii) an increased absolute SI
value; and iii) the first and last AS exons tended to be excluded
due to difficulties in designing primers. The semi-quantitative
RT-PCR was performed using 1.5% agarose gel and Gold Nucleic Acid
Gel Stain (Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China) under the following conditions: 94°C for 30 sec,
followed by 30 cycles at 94°C for 30 sec, 55°C for 30 sec and 72°C
for 60 sec.
Statistical analysis
Data are expressed as the mean ± standard deviation
for each independent experiment. Comparisons were made using an
independent sample t-test to determine the significance between the
two groups. P<0.05 was considered to indicate a statistically
significant difference. All data were analyzed with SPSS version
19.0 (IBM Corp., Armonk, NY, USA).
Results
Hypoxia inhibits the osteogenic
differentiation of CESCs
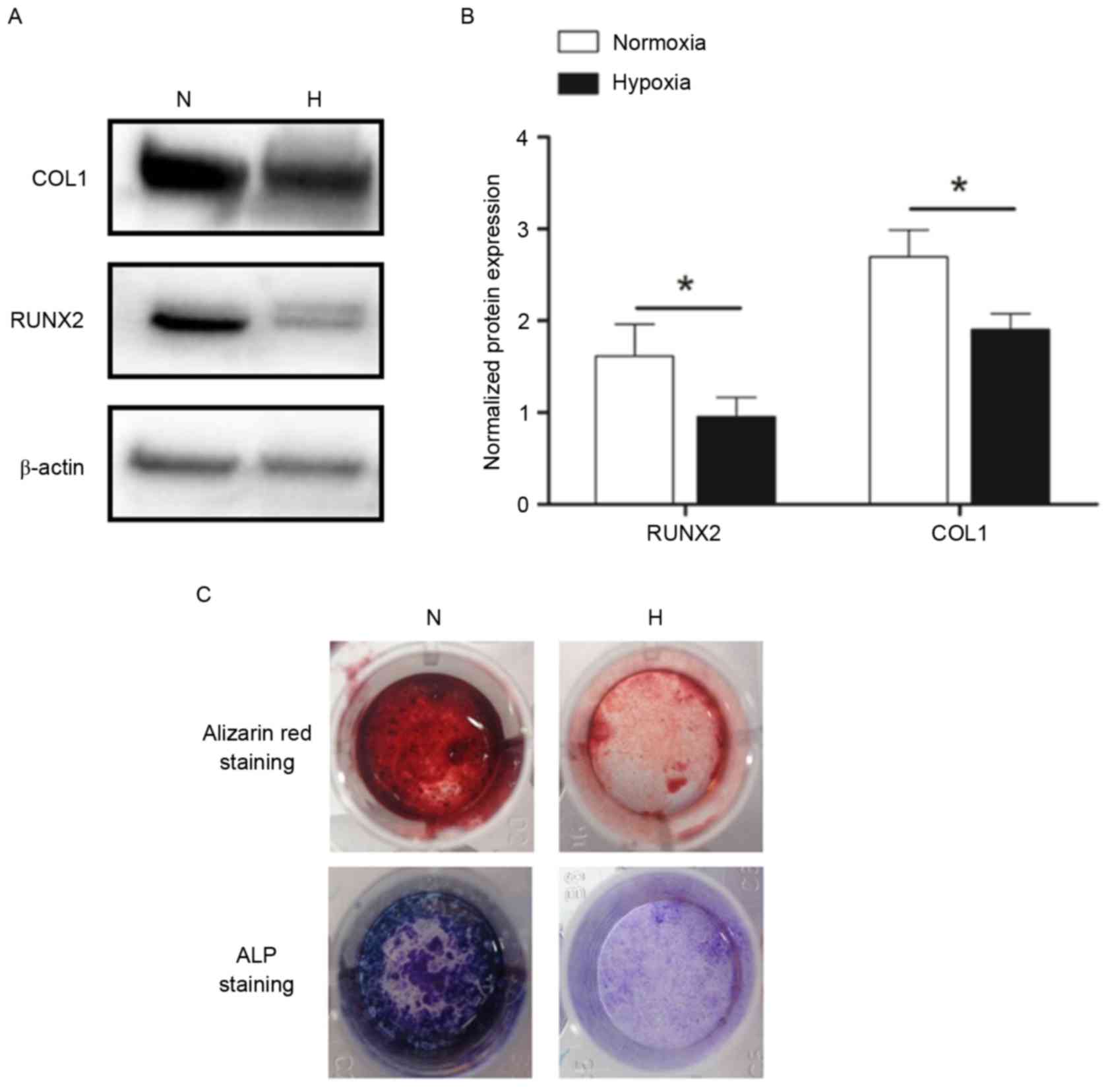
In order to evaluate the effects of hypoxia on
osteogenic differentiation, CESCs were stimulated to differentiate
into the osteogenic lineage under normoxic (21% O2) and
hypoxic (1% O2) conditions for 21 days. The protein
expression levels of osteogenic differentiation markers RUNX2 and
COL1 in the hypoxia group were decreased compared with in the
normoxia group (Fig. 2A and B). In
addition, hypoxia exhibited an inhibitory effect on the functional
mineralization of CESCs (Fig. 2C).
These findings suggested that the osteogenesis of CESCs was
inhibited in the hypoxic microenvironment.
DEG detection, validation and
functional analysis during osteogenic differentiation of CESCs
under normoxic and hypoxic conditions
A comparative genome-wide analysis of the DEGs in
the hypoxia and normoxia groups identified 214 DEGs, of which 95
(44%) were upregulated and 119 (56%) were downregulated. In
addition, 56 (26%) were non-coding transcripts. A total of 10 DEGs
were selected for validation and 8 of them were validated
(consistent tendency). The expression levels of matrix
metalloproteinase 3, aldo-keto reductase family 1 member C2, milk
fat globule EGF-factor 8 protein and C-C motif chemokine ligand 2
were upregulated, whereas the expression levels of fibroblast
growth factor 7 (FGF7), periostin (POSTN), glycoprotein M6B (GPM6B)
and insulin like growth factor binding protein 2 (IGFBP2) were
downregulated in the hypoxia group compared with the normoxia group
(Fig. 3). Two of the genes
[vascular cell adhesion molecule 1 (VCAM1) and fibulin 5 (FBLN5)]
were not differentially expressed (data not presented).
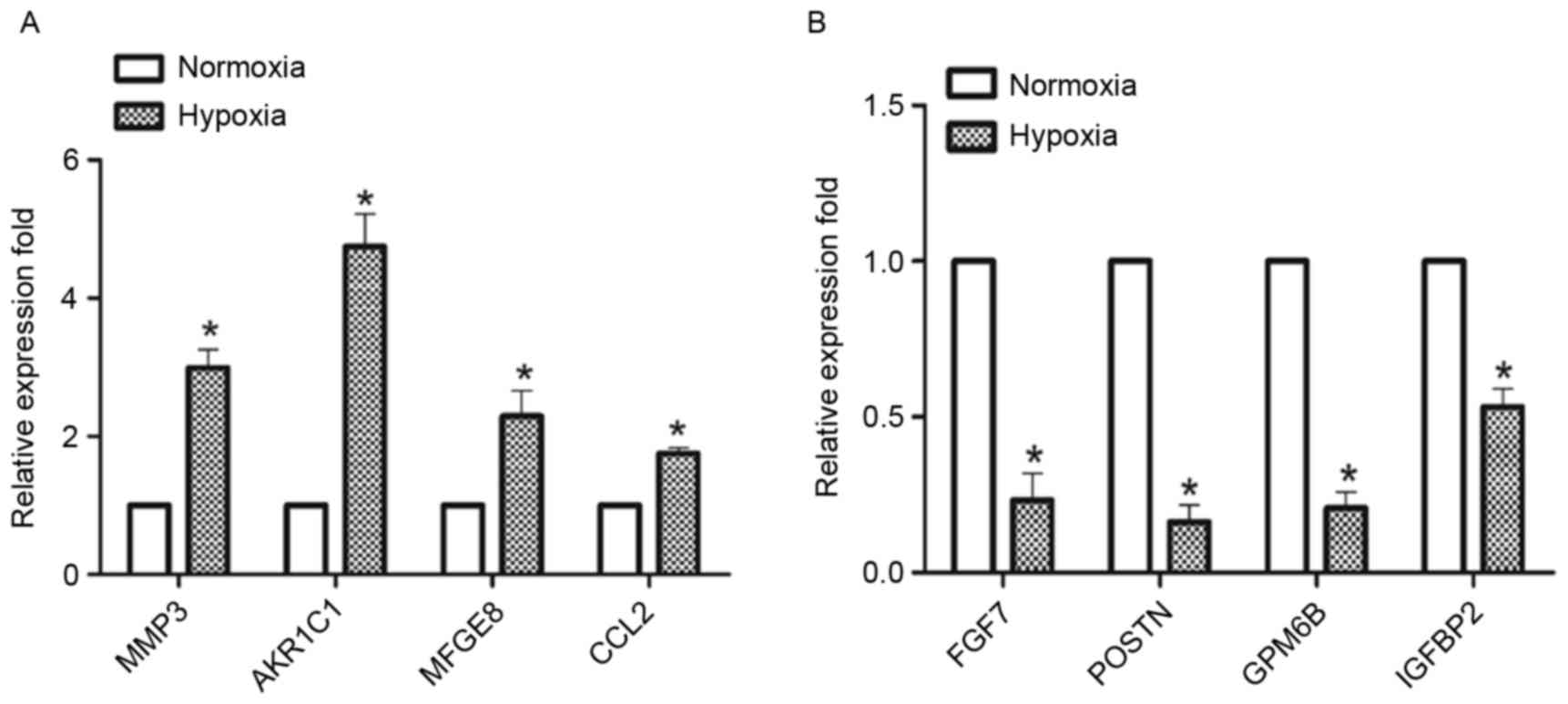 | Figure 3.DEGs in CESCs during osteogenic
differentiation under normoxic and hypoxic conditions were
validated using RT-qPCR. CESCs were induced to differentiate in
osteogenic induction medium under normoxic and hypoxic conditions
for 21 days. A total of 8 of the 10 DEGs were validated by RT-qPCR.
(A) MMP3, AKR1C1, MFGE8, CCL2, and (B) FGF7, POSTN, GPM6B and
IGFBP2. The expression levels of the genes were normalized to
β-actin, and the data are presented as the mean ± standard
deviation; n=3/group. *P<0.05 vs. respective normoxia group.
DEGs, differentially expressed genes; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; CESCs,
cartilage endplate-derived stem cells; MMP3, matrix
metalloproteinase 3; AKR1C1, aldo-keto reductase family 1 member
C1; MFGE8, milk fat globule-EGF factor 8 protein; CCL2, C-C motif
chemokine ligand 2; FGF7, fibroblast growth factor 7; POSTN,
periostin; GPM6B, glycoprotein M6B; IGFBP2, insulin like growth
factor binding protein 2. |
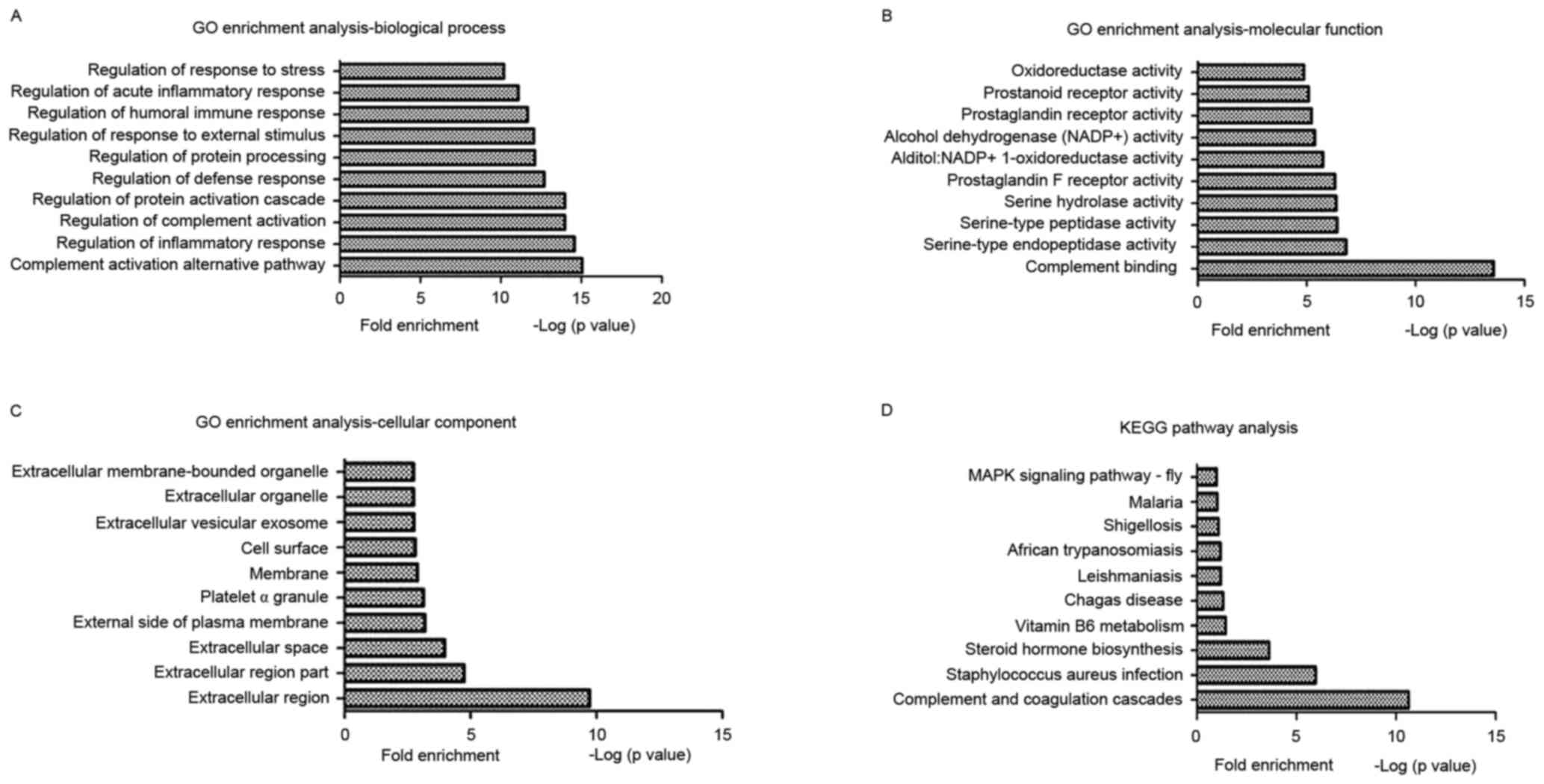
A GO enrichment analysis of the DEGs during
osteogenic differentiation of CESCs under normoxic and hypoxic
conditions was performed to identify the enriched biological
processes, molecular functions and cellular components. The results
of the present study demonstrated that certain important GO terms
were significantly enriched, including complement activation
(alternative pathway), complement binding and the extracellular
region. The top 10 GO functions that were enriched in the DEGs
during the osteogenic differentiation of CESCs under normoxic and
hypoxic conditions are presented in Fig. 4A-C.
The KEGG web-based tool was used to identify the
functional pathways that were significantly enriched in the DEGs.
The results revealed that numerous cellular pathways were involved,
including the complement and coagulation cascades,
Staphylococcus aureus infection and steroid hormone
biosynthesis. The top 10 KEGG pathways that were enriched in the
DEGs during the osteogenic differentiation of CESCs under normoxic
and hypoxic conditions are presented in Fig. 4D.
ASG detection, validation and
functional analysis during the osteogenic differentiation of CESCs
under normoxic and hypoxic conditions
The analysis of genome-wide AS events identified
6,999 AS exons belonging to 1,618 ASGs during the osteogenic
differentiation of CESCs under normoxic and hypoxic conditions. A
total of 3,227 (46%) AS exons with SI values ≥2 were defined as
‘general exon inclusion’ events, whereas the remaining 3,772 (54%)
AS exons were defined as ‘general exon exclusion’ events. Each ASG
exhibited 4.33 (6,999/1,618) AS exons on average, confirming that
numerous AS events may occur in the same gene. POSTN was selected
as a typical example, which exhibited 21 AS exons, indicating a
complex regulatory effect of AS. Notably, 42 of these 1,618 ASGs
were also significant DEGs, which indicated an underlying intrinsic
relationship between AS and gene expression. In order to validate
their accuracy, 10 ASGs were selected for the semi-quantitative
RT-PCR analysis. The data presented in Fig. 5 demonstrated that 7 of the 10
selected ASGs were successfully validated.
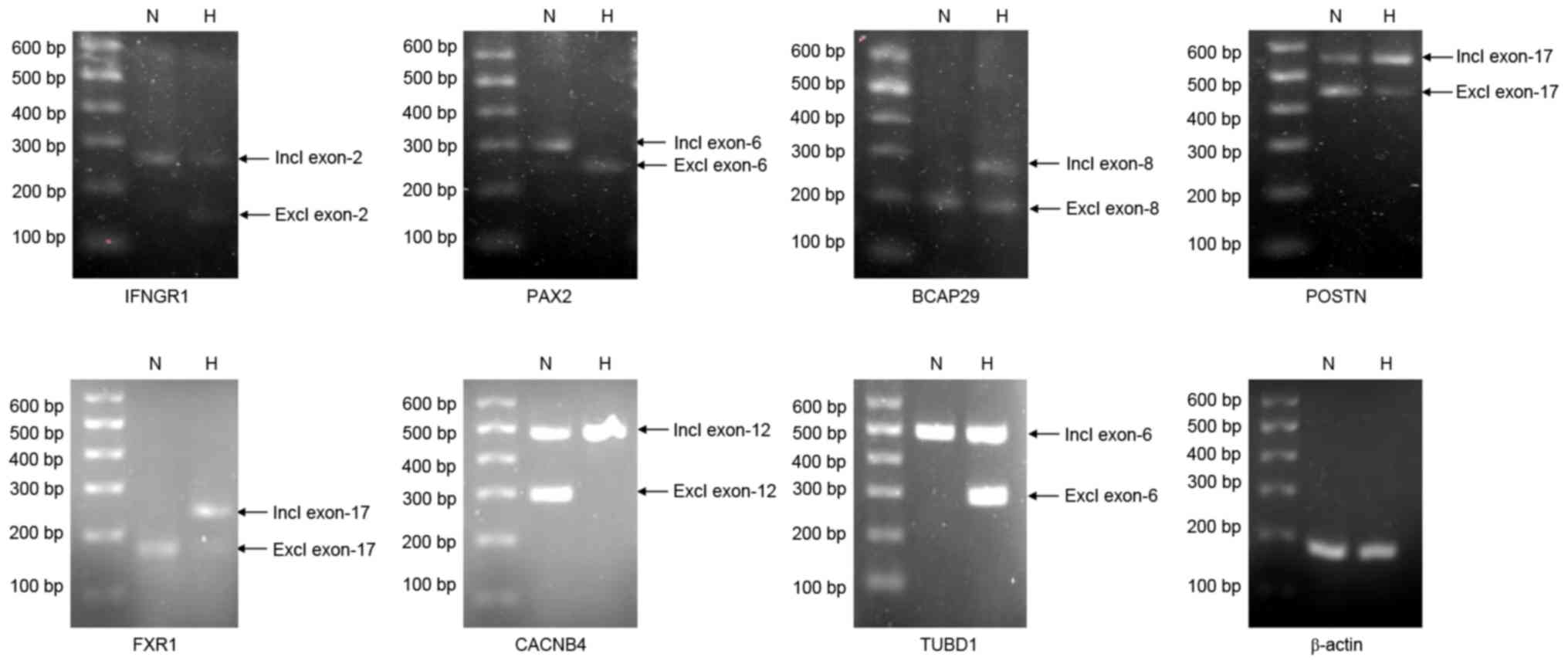 | Figure 5.Alternatively spliced genes during
the osteogenic differentiation of CESCs under normoxic and hypoxic
conditions were validated by semi-quantitative PCR. CESCs were
induced to differentiate in osteogenic induction medium under
normoxic and hypoxic conditions for 3 weeks. A total of 7 of the 10
ASGs were successfully validated by semi-quantitative PCR. β-actin
was used as an internal control. CESCs, cartilage endplate-derived
stem cells; PCR, polymerase chain reaction; N, normoxia; H,
hypoxia; IFNGR1; interferon γ receptor 1; PAX2, paired box 2;
BCAP29, B-cell receptor associated protein 29; POSTN, periostin;
FXR1, FMR1 autosomal homolog 1; CACNB4, calcium voltage-gated
channel auxiliary subunit β 4; TUBD1, tubulin δ 1; Incl, inclusion;
Excl, exclusion. |
A GO enrichment analysis of the ASGs during the
osteogenic differentiation of CESCs under normoxic and hypoxic
conditions was performed to identify the enriched biological
processes, molecular functions and cellular components. The results
of the present study demonstrated that certain GO terms were
regulated by AS in CESCs under hypoxic conditions, including
DNA-dependent regulation of transcription, protein binding and
cytoplasm. The top 10 GO terms of the ASGs during the osteogenic
differentiation of CESCs under normoxic and hypoxic conditions are
presented in Table III.
| Table III.List of top ten enriched GO terms
according to GO analysis of alternatively spliced genes. |
Table III.
List of top ten enriched GO terms
according to GO analysis of alternatively spliced genes.
| GO term | Count | P-value |
|---|
| Biological
process |
|
GO:0006355 regulation of
transcription, DNA-dependent | 133 |
3.69×10−104 |
|
GO:0006350 transcription | 114 |
1.30×10−77 |
|
GO:0007049 cell cycle | 64 |
1.15×10−57 |
|
GO:0006468 protein amino acid
phosphorylation | 57 |
2.01×10−54 |
|
GO:0007165 signal
transduction | 101 |
1.93×10−50 |
|
GO:0055114 oxidation
reduction | 48 |
5.60×10−45 |
|
GO:0051301 cell division | 37 |
6.57×10−45 |
|
GO:0019941
modification-dependent protein catabolism | 46 |
4.68×10−44 |
|
GO:0007067 mitosis | 29 |
7.80×10−34 |
|
GO:0007155 cell adhesion | 42 |
5.84×10−33 |
| Molecular
function |
|
GO:0005515 protein
binding | 504 | 0 |
|
GO:0008270 zinc ion
binding | 206 |
1.30×10−202 |
|
GO:0000166 nucleotide
binding | 184 |
4.57×10−182 |
|
GO:0046872 metal ion
binding | 222 |
1.73×10−171 |
|
GO:0005524 ATP binding | 154 |
6.58×10−169 |
|
GO:0016740 transferase
activity | 130 |
7.67×10−120 |
|
GO:0005509 calcium ion
binding | 82 |
1.78×10−80 |
|
GO:0003677 DNA binding | 99 |
4.12×10−66 |
|
GO:0003723 RNA binding | 62 |
6.92×10−61 |
|
GO:0004674 protein
serine/threonine kinase activity | 47 |
2.39×10−51 |
| Cellular
component |
|
GO:0005737 cytoplasm | 453 | 0 |
|
GO:0005634 nucleus | 455 | 0 |
|
GO:0016020 membrane | 265 |
1.01×10−163 |
|
GO:0016021 integral to
membrane | 212 |
6.83×10−136 |
|
GO:0005829 cytosol | 108 |
1.08×10−117 |
|
GO:0005886 plasma
membrane | 150 |
3.25×10−98 |
|
GO:0005576 extracellular
region | 116 |
2.69×10−90 |
|
GO:0005794 Golgi
apparatus | 69 |
1.30×10−65 |
|
GO:0005783 endoplasmic
reticulum | 69 |
2.45×10−63 |
|
GO:0005739 mitochondrion | 71 |
1.01×10−61 |
The 1,618 ASGs were analyzed with the KEGG web-based
tool to identify the enriched signaling pathways. The results
demonstrated that numerous signaling pathways were significantly
affected, including focal adhesion, ubiquitin-mediated proteolysis
and the mitogen-activated protein kinase signaling pathway. The top
10 KEGG pathways that were enriched in the ASGs during the
osteogenic differentiation of CESCs under normoxic and hypoxic
conditions are presented in Table
IV.
| Table IV.List of top 10 signaling pathways
enriched by Kyoto Encyclopedia of Genes and Genomes analysis of
alternatively spliced genes. |
Table IV.
List of top 10 signaling pathways
enriched by Kyoto Encyclopedia of Genes and Genomes analysis of
alternatively spliced genes.
| Pathway | Count | P-value |
|---|
| Focal adhesion | 31 |
4.68×10−18 |
| Ubiquitin mediated
proteolysis | 25 |
1.16×10−16 |
| MAPK signaling
pathway | 29 |
7.75×10−13 |
| ECM-receptor
interaction | 15 |
1.64×10−10 |
| Regulation of actin
cytoskeleton | 23 |
1.78×10−10 |
| Axon guidance | 18 |
2.00×10−10 |
| Insulin signaling
pathway | 17 |
3.97×10−9 |
| Leukocyte
transendothelial migration | 16 |
4.01×10−9 |
| Complement and
coagulation cascades | 12 |
1.51×10−8 |
| Cell adhesion
molecules | 16 |
1.78×10−8 |
Discussion
CESCs are considered to be MSCs due to their
embryonic mesoderm origin, similar surface immunophenotypes and
similar differentiation capacity compared with other reported MSCs
(12).
The in situ environment of MSCs is frequently
hypoxic: Bone marrow, 4–7% O2; muscle, 1–10%
O2; and adipose tissue, 3.8–9.6% O2 (27,28).
In response to hypoxia, MSCs exhibit altered differentiation fates
according to their different tissues of origin. For example, in
bone marrow MSCs, the hypoxic microenvironment has been
demonstrated to favor chondrogenesis, and inhibit osteogenesis and
adipogenesis (29–31). In adipose MSCs, adipogenesis and
chondrogenesis have been observed to be promoted, whereas
osteogenesis was reduced under hypoxic conditions (15,32).
In muscle MSCs, the decreased oxygen level improved myogenic
differentiation and inhibited adipogenic differentiation (22). In addition, periodontal ligament
MSCs exhibited increased osteogenic differentiation under hypoxic
conditions (33). Therefore, stem
cells derived from various tissues exhibit a tissue-specific
differentiation fate in response to physiological hypoxia, which
may be beneficial for their physiological functions.
In the present study, OIM was used under both
hypoxic and normoxic conditions. Under normoxic conditions, Huang
et al (34) reported that
OIM was able to significantly induce the osteogenic differentiation
of CESCs, compared with normal growth medium. It was observed in
Huang et al (34) study
that the expression of osteogenic differentiation marker genes
(RUNX2, ALP and OC) was decreased in the absence of OIM under
normoxic conditions. In addition, previous studies have
demonstrated that stem cell differentiation may be decreased in
hypoxic conditions compared with normoxic conditions (35,36).
Therefore, in the absence of OIM, osteogenic differentiation occurs
at a low level, and this is physiologically important for CESCs to
maintain their stem cell properties. In the research area of
hypoxia/normoxia, normal growth medium is frequently used to
investigate growth, including proliferation and apoptosis (37,38),
whereas OIM is used to study the potential for osteogenesis
(15,30). The present study focused on the
potential of stem cells to undergo osteogenesis; therefore, OIM was
used in the present study.
Anatomically, the CEPs of healthy, mature juvenile,
adolescent and adult discs are free of blood vessels, whereas the
bone endplate (BEP, part of the vertebral bone) contains extensive
blood spaces for the vertebral blood sinuses. Blood vessels end at
the interface between the CEP and BEP, without invading the CEP
(39). Lee et al (40) reported that all three components of
the IVD (nucleus pulposus, annulus fibrosus and CEP) remained in a
hypoxic microenvironment in a rat model using the 2-nitroimidazole,
EF5, a drug that forms covalent adducts with cellular proteins at
low oxygen concentrations. During the process of degeneration,
blood vessels may invade into the CEP through fissures (39,41,42).
Blood vessel invasion may increase the oxygen levels, which may
disrupt the physiological hypoxic microenvironment of the CESCs.
This destruction of the hypoxic microenvironment may facilitate the
osteogenic differentiation of CESCs, which may initiate the
ossification of the CEP. The ossification of the CEP has been
demonstrated to result in a poor capacity to resist mechanical
stress and poor nutrient exchange, and, therefore, to initiate IVD
degeneration.
The present study sought to elucidate the mechanism
through which hypoxia inhibits the osteogenic differentiation of
CESCs. At present, genome-wide transcription analysis is the
predominant method used to study the mechanism of stem cell
differentiation. Previous studies have used this tool to obtain a
coherent view of the transcriptional and post-transcriptional
alterations during the differentiation process (43–46).
However, to the best of our knowledge, no previous studies have
investigated the AS mechanisms of stem cell differentiation under
hypoxic conditions on a genome-wide scale. AS is considered to be
an intricate regulatory mechanism through which a single pre-mRNA
may produce various mature RNA subtypes, which leads to structural
diversity of the genetic phenotypes without expansion of the genome
(47). Since hypoxia is a source
of AS events, and AS is associated with the regulation of the
osteogenic differentiation of stem cells, it was hypothesized that
AS may be the connection between hypoxia and the chondrogenic
differentiation of CESCs (16–18).
Therefore, the present study used high-throughput screening
technology to identify DEGs and ASGs during the osteogenic
differentiation of CESCs under normoxic and hypoxic conditions, and
analyzed the GO enrichment terms and functional pathways with
relevant web-based bioinformatics tools.
The detected DEGs between the normoxic osteogenic
induction and hypoxic osteogenic induction groups were validated by
qPCR. FGF7, GPM6B and IGFBP2, which were downregulated in the
hypoxia group compared with the normoxia group, were used as
examples. FGF7 may facilitate the dexamethasone-,
β-glycerophosphate- and ascorbic acid-induced osteogenic
differentiation of embryonic stem cells into bone-like nodules and
induce mineralization through the extracellular signal-related
kinase/RUNX2 signaling pathway (48). GPM6B, which encodes a membrane
glycoprotein of the proteolipid protein family, is upregulated
during osteoblast differentiation. GPM6B silencing in MSCs was
observed to lead to decreased mineralization of the extracellular
matrix and reduced ALP activity; a microarray analysis attributed
these alterations to cytoskeleton and matrix vesicle release
(49). IGFBP2 was reported to
trigger the osteogenic differentiation of MSCs by interacting with
integrin α 5 and insulin like growth factor 2 (50). These results suggested that FGF7,
GPM6B and IGFBP2, which were downregulated in the present study,
may be associated with the osteogenesis of CESCs, confirming the
inhibition of osteogenic differentiation by hypoxia.
KEGG and GO analysis of the identified DEGs was
performed in the present study. The results demonstrated that the
complement and coagulation cascades were enriched in the DEGs,
according to the KEGG analysis. In addition, in the GO analysis of
the DEGs, the biological process of complement activation
(alternative pathway) and the molecular function of complement
binding were enriched. The results of the present study indicated
that the complement pathway may be associated with the mechanism
through which hypoxia is able to regulate the osteogenic
differentiation of CESCs.
Due to the avascular nature and low immunogenicity
of the IVD, few studies have investigated the role of immune
signaling in the CEP. However, blood vessel invasion may be
observed in the degenerated CEP, and revascularization may provide
immune signals (30,31). Furthermore, numerous immune signals
have been observed to be involved in a series of non-immune
effects. In the present study, the complement pathway was used as
an example. The complement pathway is known to exhibit roles in
immune surveillance, and has previously been demonstrated to be
involved in non-immune effects, including angiogenesis, clearance
of apoptotic cells, and stem cell recruitment and differentiation
(51). MSCs and osteoblasts
express proteins in the complement cascade (52), including complement C3, C3a
anaphylatoxin chemotactic receptor (C3aR), complement C5, C5a
anaphylatoxin chemotactic receptor (C5aR), and cell surface
markers, including cluster of differentiation (CD)46, CD55 and CD59
(53). C5aR expression was
markedly increased during osteogenic differentiation in MSCs
(54), and osteogenesis was
reported to be accelerated in the presence of C5a and C3a in a
C5aR- and C3aR-specific manner (41). In the present study, the expression
of C3, the precursor of C3a, in the hypoxic group was reduced
compared to the normoxic group, indicating a possible method by
which hypoxia inhibits osteogenesis of CESCs. As previously
mentioned, it can be hypothesized that the complement cascade was
enriched according to GO and KEGG analysis. The precise and
complicated regulation of complement cascade on osteogenesis may
include both enhancement and suppression. Since hypoxia is an
important activator of the complement pathway (55,56),
complement activation may serve a role in regulating the osteogenic
differentiation of CESCs under hypoxic conditions.
A total of 7 out of the 10 chosen ASGs were
successfully validated. In the present study, interferon γ (IFNγ)
receptor 1 (IFNGR1) was used as an example to elucidate the role of
AS. The exclusion of exon 2 of IFNGR1 was reported to be a
characteristic of IFNGR1 deficiency disease, in which patients
exhibited impaired IFNγ-mediated function, leading to
susceptibility to infection (57).
In the present study of the osteogenic differentiation of CESCs,
the exclusion of exon 2 of IFNGR1 was detected in the hypoxia group
compared with in the normoxia group. IFNGR1 is the receptor for
IFNγ; the IFNγ/IFNGR1 complex is responsible for activation of the
IFN pathway. IFNγ was reported to promote osteogenic
differentiation in vitro (58) and in vivo (59), whereas the osteogenesis of
allogeneic MSCs was inhibited by IFNγ (60). Given that exon 2 exclusion may lead
to the change of IFNγ-mediated function, it is reasonable to
consider that exon 2 exclusion may also influence the IFNγ-mediated
regulation of osteogenesis. In conclusion, since exon 2 exclusion
occurs in the hypoxia-regulated osteogenesis system and the immune
system, it may be hypothesized that immune signals may be
associated with immunomodulation, in addition to exhibiting
non-immune effects, including osteogenesis regulation, through AS
under hypoxic conditions. At present, studies on the role of ASG in
hypoxia-regulated osteogenesis remain scarce. However, the results
of the present study may offer useful insights for further studies
into the role of AS in the mechanism of hypoxia-regulated
osteogenesis.
KEGG and GO analyses of the ASGs were performed.
Consistent with the high-throughput analysis of the DEGs, the
complement and coagulation cascades were enriched in the KEGG
analysis. A previous study reported that AS may influence the
complement cascade by producing various protein phenotypes
(61); therefore, it was
hypothesized that AS may be associated with the complement pathway
during the process of hypoxia-regulated osteogenesis. In addition,
the biological process of DNA-dependent regulation of transcription
and the molecular function of nucleotide binding were enriched. A
number of the regulatory molecules that are expressed during the
osteogenic differentiation process, including RUNX2 and osterix,
are transcription factors that function by binding to the promoter
of target genes to promote or inhibit transcription (62). In addition, the cellular components
cytoplasm and nucleus were enriched, which indicated an increased
rate of cytoplasmic/nuclear translocation. A number of molecules
with regulatory effects undergo cytoplasmic/nuclear translocation,
including nuclear factor-κB (63)
and AKT (64). The GO analysis of
the ASGs performed in the present study indicated that AS may be
associated with nuclear signal transduction, which may be the
functional factor that is influenced by AS during the
hypoxia-regulated osteogenesis of CESCs.
CEP chondrocytes (CEPCs) are present in the CEP.
Therefore, further studies are required to analyze the role of
hypoxia in the differentiation/AS regulation of CEPCs. CEPCs are a
type of terminally differentiated chondrocyte. To the best of our
knowledge, few previous studies have focused on the differentiation
properties of terminally differentiated cells. However, hypoxia is
reported to induce dedifferentiation to drive committed cells
towards a pluripotent fate (65);
therefore, hypoxia may serve a role in transforming CEPCs into
CESCs. A previous study demonstrated the regulatory effects of AS
on chondrocytes in response to hypoxia. For example, vascular
endothelial growth factor (VEGF)120 and
VEGF164 are the most abundant splicing variants that are
expressed in chondrocytes in response to decreased oxygen levels
(66). Therefore, hypoxia/normoxia
may regulate AS in CEPCs; however, the specific mechanisms require
further study.
In conclusion, the results of the present study
demonstrated that hypoxia inhibited osteogenesis in CESCs.
Alterations in the gene expression profiles and AS events were
observed on a genome-wide scale. The subsequent GO and KEGG
analyses provided a reference for future mechanistic studies of the
gene expression profiles and AS regulation during the inhibition of
osteogenesis. Notably, the identification of the significance of
the complement pathway in the hypoxia-mediated regulation of
osteogenic differentiation will be of use in improving the
understanding of this physiological phenomenon, and may be
important for the identification of targets for CEP degeneration
therapy.
Acknowledgements
The authors of the present study would like to
acknowledge Dr Yi Zha (CapitalBio Corporation, Beijing, China), for
help with analyzing the microarray data. The present study was
supported by the National Natural Science Foundation of China
(grant nos. 81472076, 81271982 and 81401801).
Glossary
Abbreviations
Abbreviations:
|
IVD
|
intervertebral disc
|
|
LBP
|
lower back pain
|
|
DDD
|
degenerative disc disease
|
|
CEP
|
cartilage endplate
|
|
MSCs
|
mesenchymal stem cells
|
|
CESCs
|
cartilage endplate-derived stem
cells
|
|
DEGs
|
differentially expressed genes
|
|
ASGs
|
alternatively spliced genes
|
|
AS
|
alternative splicing
|
References
|
1
|
Andersson GB: Epidemiological features of
chronic low-back pain. Lancet. 354:581–585. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Freemont AJ: The cellular pathobiology of
the degenerate intervertebral disc and discogenic back pain.
Rheumatology (Oxford). 48:5–10. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stokes IA and Iatridis JC: Mechanical
conditions that accelerate intervertebral disc degeneration:
Overload versus immobilization. Spine (Phila Pa 1976).
29:2724–2732. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Le Maitre CL, Freemont AJ and Hoyland JA:
Accelerated cellular senescence in degenerate intervertebral discs:
A possible role in the pathogenesis of intervertebral disc
degeneration. Arthritis Res Ther. 9:R452007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao CQ, Wang LM, Jiang LS and Dai LY: The
cell biology of intervertebral disc aging and degeneration. Ageing
Res Rev. 6:247–261. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Urban JP, Smith S and Fairbank JC:
Nutrition of the intervertebral disc. Spine (Phila Pa 1976).
29:2700–2709. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Buckwalter JA: Aging and degeneration of
the human intervertebral disc. Spine (Phila Pa 1976). 20:1307–1314.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Holm S, Maroudas A, Urban JP, Selstam G
and Nachemson A: Nutrition of the intervertebral disc: Solute
transport and metabolism. Connect Tissue Res. 8:101–119. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Raj PP: Intervertebral disc:
Anatomy-physiology- pathophysiology-treatment. Pain Pract. 8:18–44.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li FC, Zhang N, Chen WS and Chen QX:
Endplate degeneration may be the origination of the vacuum
phenomenon in intervertebral discs. Med Hypotheses. 75:169–171.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jackson AR, Huang CY and Gu WY: Effect of
endplate calcification and mechanical deformation on the
distribution of glucose in intervertebral disc: A 3D finite element
study. Comput Methods Biomech Biomed Engin. 14:195–204. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Roberts S, Urban JP, Evans H and
Eisenstein SM: Transport properties of the human cartilage endplate
in relation to its composition and calcification. Spine (Phila Pa
1976). 21:415–420. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu LT, Huang B, Li CQ, Zhuang Y, Wang J
and Zhou Y: Characteristics of stem cells derived from the
degenerated human intervertebral disc cartilage endplate. PLoS One.
6:e262852011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Boskey AL: Signaling in response to
hypoxia and normoxia in the intervertebral disc. Arthritis Rheum.
58:3637–3639. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Merceron C, Vinatier C, Portron S, Masson
M, Amiaud J, Guigand L, Chérel Y, Weiss P and Guicheux J:
Differential effects of hypoxia on osteochondrogenic potential of
human adipose-derived stem cells. Am J Physiol Cell Physiol.
298:C355–C364. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Makino Y, Kanopka A, Wilson WJ, Tanaka H
and Poellinger L: Inhibitory PAS domain protein (IPAS) is a
hypoxia-inducible splicing variant of the hypoxia-inducible
factor-3alpha locus. J Biol Chem. 277:32405–32408. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tacconelli A, Farina AR, Cappabianca L,
Desantis G, Tessitore A, Vetuschi A, Sferra R, Rucci N, Argenti B,
Screpanti I, et al: TrkA alternative splicing: A regulated
tumor-promoting switch in human neuroblastoma. Cancer Cell.
6:347–360. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kazantseva J, Kivil A, Tints K, Kazantseva
A, Neuman T and Palm K: Alternative splicing targeting the
hTAF4-TAFH domain of TAF4 represses proliferation and accelerates
chondrogenic differentiation of human mesenchymal stem cells. PLoS
One. 8:e747992013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kazantseva J, Kivil A, Tints K, Kazantseva
A, Neuman T and Palm K: PTHrP in differentiating human mesenchymal
stem cells: Transcript isoform expression, promoter methylation,
and protein accumulation. Biochimie. 95:1888–1896. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hang X, Li P, Li Z, Qu W, Yu Y, Li H, Shen
Z, Zheng H, Gao Y, Wu Y, et al: Transcription and splicing
regulation in human umbilical vein endothelial cells under hypoxic
stress conditions by exon array. BMC Genomics. 10:1262009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Moller-Levet CS, Betts GN, Harris AL,
Homer JJ, West CM and Miller CJ: Exon array analysis of head and
neck cancers identifies a hypoxia related splice variant of LAMA3
associated with a poor prognosis. PLoS Comput Biol. 5:e10005712009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ducy P, Zhang R, Geoffroy V, Ridall AL and
Karsenty G: Osf2/Cbfa1: A transcriptional activator of osteoblast
differentiation. Cell. 89:747–754. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lynch MP, Stein JL, Stein GS and Lian JB:
The influence of type I collagen on the development and maintenance
of the osteoblast phenotype in primary and passaged rat calvarial
osteoblasts: Modification of expression of genes supporting cell
growth, adhesion, and extracellular matrix mineralization. Exp Cell
Res. 216:35–45. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Andrianarivo AG, Robinson JA, Mann KG and
Tracy RP: Growth on type I collagen promotes expression of the
osteoblastic phenotype in human osteosarcoma MG-63 cells. J Cell
Physiol. 153:256–265. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu N, Shi S, Deng M, Tang L, Zhang G, Liu
N, Ding B, Liu W, Liu Y, Shi H, et al: High levels of β-catenin
signaling reduce osteogenic differentiation of stem cells in
inflammatory microenvironments through inhibition of the
noncanonical Wnt pathway. J Bone Miner Res. 26:2082–2095. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Derveaux S, Vandesompele J and Hellemans
J: How to do successful gene expression analysis using real-time
PCR. Methods. 50:227–230. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schiller ZA, Schiele NR, Sims JK, Lee K
and Kuo CK: Adipogenesis of adipose-derived stem cells may be
regulated via the cytoskeleton at physiological oxygen levels in
vitro. Stem Cell Res Ther. 4:792013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Redshaw Z and Loughna PT: Oxygen
concentration modulates the differentiation of muscle stem cells
toward myogenic and adipogenic fates. Differentiation. 84:193–202.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Khan WS, Adesida AB, Tew SR, Lowe ET and
Hardingham TE: Bone marrow-derived mesenchymal stem cells express
the pericyte marker 3G5 in culture and show enhanced chondrogenesis
in hypoxic conditions. J Orthop Res. 28:834–840. 2010.PubMed/NCBI
|
|
30
|
Yang DC, Yang MH, Tsai CC, Huang TF, Chen
YH and Hung SC: Hypoxia inhibits osteogenesis in human mesenchymal
stem cells through direct regulation of RUNX2 by TWIST. PLoS One.
6:e239652011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Martin-Rendon E, Hale SJ, Ryan D, Baban D,
Forde SP, Roubelakis M, Sweeney D, Moukayed M, Harris AL, Davies K
and Watt SM: Transcriptional profiling of human cord blood
CD133+ and cultured bone marrow mesenchymal stem cells
in response to hypoxia. Stem Cells. 25:1003–1012. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim JH, Kim SH, Song SY, Kim WS, Song SU,
Yi T, Jeon MS, Chung HM, Xia Y and Sung JH: Hypoxia induces
adipocyte differentiation of adipose-derived stem cells by
triggering reactive oxygen species generation. Cell Biol Int.
38:32–40. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang QB, Zhang ZQ, Fang SL, Liu YR, Jiang
G and Li KF: Effects of hypoxia on proliferation and osteogenic
differentiation of periodontal ligament stem cells: An in vitro and
in vivo study. Genet Mol Res. 13:10204–10214. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang B, Liu LT, Li CQ, Zhuang Y, Luo G,
Hu SY and Zhou Y: Study to determine the presence of progenitor
cells in the degenerated human cartilage endplates. Eur Spine J.
21:613–622. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
D'Ippolito G, Diabira S, Howard GA, Roos
BA and Schiller PC: Low oxygen tension inhibits osteogenic
differentiation and enhances stemness of human MIAMI cells. Bone.
39:513–522. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Choi JR, Pingguan-Murphy B, Wan Abas WA,
Azmi MA Noor, Omar SZ, Chua KH and Wan Safwani WK: Impact of low
oxygen tension on stemness, proliferation and differentiation
potential of human adipose-derived stem cells. Biochem Biophys Res
Commun. 448:218–224. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Brock M, Haider TJ, Vogel J, Gassmann M,
Speich R, Trenkmann M, Ulrich S, Kohler M and Huber LC: The
hypoxia-induced microRNA-130a controls pulmonary smooth muscle cell
proliferation by directly targeting CDKN1A. Int J Biochem Cell
Biol. 61:129–137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Leszczynska KB, Foskolou IP, Abraham AG,
Anbalagan S, Tellier C, Haider S, Span PN, O'Neill EE, Buffa FM and
Hammond EM: Hypoxia-induced p53 modulates both apoptosis and
radiosensitivity via AKT. J Clin Invest. 125:2385–2398. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nerlich AG, Schaaf R, Wälchli B and Boos
N: Temporo-spatial distribution of blood vessels in human lumbar
intervertebral discs. Eur Spine J. 16:547–555. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lee DC, Adams CS, Albert TJ, Shapiro IM,
Evans SM and Koch CJ: In situ oxygen utilization in the rat
intervertebral disc. J Anat. 210:294–303. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Freemont AJ, Watkins A, Le Maitre C, Baird
P, Jeziorska M, Knight MT, Ross ER, O'Brien JP and Hoyland JA:
Nerve growth factor expression and innervation of the painful
intervertebral disc. J Pathol. 197:286–292. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Walsh DA, McWilliams DF, Turley MJ, Dixon
MR, Fransès RE, Mapp PI and Wilson D: Angiogenesis and nerve growth
factor at the osteochondral junction in rheumatoid arthritis and
osteoarthritis. Rheumatology (Oxford). 49:1852–1861. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Babadagli ME, Tezcan B, Yilmaz ST and
Tufan AC: Matrilin-3 as a putative effector of C-type natriuretic
peptide signaling during TGF-β induced chondrogenic differentiation
of mesenchymal stem cells. Mol Biol Rep. 41:5549–5555. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fernandes AM, Herlofsen SR, Karlsen TA,
Küchler AM, Fløisand Y and Brinchmann JE: Similar properties of
chondrocytes from osteoarthritis joints and mesenchymal stem cells
from healthy donors for tissue engineering of articular cartilage.
PLoS One. 8:e629942013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Herlofsen SR, Bryne JC, Høiby T, Wang L,
Issner R, Zhang X, Coyne MJ, Boyle P, Gu H, Meza-Zepeda LA, et al:
Genome-wide map of quantified epigenetic changes during in vitro
chondrogenic differentiation of primary human mesenchymal stem
cells. BMC Genomics. 14:1052013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Weber M, Sotoca AM, Kupfer P, Guthke R and
van Zoelen EJ: Dynamic modelling of microRNA regulation during
mesenchymal stem cell differentiation. BMC Syst Biol. 7:1242013.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Blencowe BJ: Alternative splicing: New
insights from global analyses. Cell. 126:37–47. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jeon YM, Kook SH, Rho SJ, Lim SS, Choi KC,
Kim HS, Kim JG and Lee JC: Fibroblast growth factor-7 facilitates
osteogenic differentiation of embryonic stem cells through the
activation of ERK/Runx2 signaling. Mol Cell Biochem. 382:37–45.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Drabek K, van de Peppel J, Eijken M and
van Leeuwen JP: GPM6B regulates osteoblast function and induction
of mineralization by controlling cytoskeleton and matrix vesicle
release. J Bone Miner Res. 26:2045–2051. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hamidouche Z, Fromigué O, Ringe J, Häupl T
and Marie PJ: Crosstalks between integrin alpha 5 and IGF2/IGFBP2
signalling trigger human bone marrow-derived mesenchymal stromal
osteogenic differentiation. BMC Cell Biol. 11:442010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Schraufstatter IU, Khaldoyanidi SK and
DiScipio RG: Complement activation in the context of stem cells and
tissue repair. World J Stem Cells. 7:1090–1108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lee DS, Yi TG, Lee HJ, Kim SN, Park S,
Jeon MS and Song SU: Mesenchymal stem cells infected with
Mycoplasma arginini secrete complement C3 to regulate
immunoglobulin production in B lymphocytes. Cell Death Dis.
5:e11922014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Soland MA, Bego M, Colletti E, Zanjani ED,
St Jeor S, Porada CD and Almeida-Porada G: Mesenchymal stem cells
engineered to inhibit complement-mediated damage. PLoS One.
8:e604612013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ignatius A, Ehrnthaller C, Brenner RE,
Kreja L, Schoengraf P, Lisson P, Blakytny R, Recknagel S, Claes L,
Gebhard F, et al: The anaphylatoxin receptor C5aR is present during
fracture healing in rats and mediates osteoblast migration in
vitro. J Trauma. 71:952–960. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Collard CD, Väkevä A, Morrissey MA, Agah
A, Rollins SA, Reenstra WR, Buras JA, Meri S and Stahl GL:
Complement activation after oxidative stress: Role of the lectin
complement pathway. Am J Pathol. 156:1549–1556. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cowell RM, Plane JM and Silverstein FS:
Complement activation contributes to hypoxic-ischemic brain injury
in neonatal rats. J Neurosci. 23:9459–9468. 2003.PubMed/NCBI
|
|
57
|
Altare F, Jouanguy E, Lamhamedi-Cherradi
S, Fondanéche MC, Fizame C, Ribiérre F, Merlin G, Dembic Z,
Schreiber R, Lisowska-Grospierre B, et al: A causative relationship
between mutant IFNgR1 alleles and impaired cellular response to
IFNgamma in a compound heterozygous child. Am J Hum Genet.
62:723–726. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
58
|
Duque G, Huang DC, Macoritto M, Rivas D,
Yang XF, Ste-Marie LG and Kremer R: Autocrine regulation of
interferon gamma in mesenchymal stem cells plays a role in early
osteoblastogenesis. Stem Cells. 27:550–558. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Duque G, Huang DC, Dion N, Macoritto M,
Rivas D, Li W, Yang XF, Li J, Lian J, Marino FT, et al:
Interferon-γ plays a role in bone formation in vivo and rescues
osteoporosis in ovariectomized mice. J Bone Miner Res.
26:1472–1483. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Dighe AS, Yang S, Madhu V, Balian G and
Cui Q: Interferon gamma and T cells inhibit osteogenesis induced by
allogeneic mesenchymal stromal cells. J Orthop Res. 31:227–234.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Olszowski T, Poziomkowska-Gęsicka I,
Jensenius JC and Adler G: Lectin pathway of complement activation
in a Polish woman with MASP-2 deficiency. Immunobiology.
219:261–262. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Choung HW, Lee DS, Lee HK, Shon WJ and
Park JC: Preameloblast-derived factors mediate osteoblast
differentiation of human bone marrow mesenchymal stem cells by
Runx2-Osterix-BSP signaling. Tissue Eng Part A. 22:93–102. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Haddad JJ: Endotoxin-mediated regulation
of nuclear factor-kappaB nuclear translocation and activation in
the hippocampus of the central nervous system: Modulation by
intracerebroventricular treatment with thymulin and the
immunomodulatory role of the IkappaB-alpha/pIkappaB-alpha pathway.
Neuroscience. 164:1509–1520. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Nguyen TL Xuan, Choi JW, Lee SB, Ye K, Woo
SD, Lee KH and Ahn JY: Akt phosphorylation is essential for nuclear
translocation and retention in NGF-stimulated PC12 cells. Biochem
Biophys Res Commun. 349:789–798. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mathieu J, Zhang Z, Nelson A, Lamba DA,
Reh TA, Ware C and Ruohola-Baker H: Hypoxia induces re-entry of
committed cells into pluripotency. Stem Cells. 31:1737–1748. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Cramer T, Schipani E, Johnson RS, Swoboda
B and Pfander D: Expression of VEGF isoforms by epiphyseal
chondrocytes during low-oxygen tension is HIF-1 alpha dependent.
Osteoarthritis Cartilage. 12:433–439. 2004. View Article : Google Scholar : PubMed/NCBI
|