Introduction
Ischemic heart disease (IHD) is a major factor in
terms of influencing worldwide mortality rates. IHD has also become
a significant health problem among the Thai population and has led
to a high mortality rate in the country (1). Moreover, this disease is an important
cause of myocardial infarction (MI), which is related to
atherosclerosis. When the mechanical change of myocardium occurs
after the ischemic condition, cardiomyocytes die and progress to
form the scar tissue that often leads to irreversible defects in
cardiac functions (2). Naturally,
a heart has a limitation for tissue regeneration due to the fact
that cardiomyocytes cannot divide after birth (3–5).
Treatment options for the ischemic condition involve taking
medicine, surgical procedures and stem cell therapy using
mesenchymal stem cell (MSCs) (3).
MSCs are a type of adult stem cells that have the capability of
being differentiated into cardiomyocytes when they were induced by
certain factors such as 5-azacytidine (5-aza) (6–11).
These cells can be isolated from bone marrow, adipose tissue,
amniotic fluid, the umbilical cord, placenta and peripheral blood
(12,13). They have displayed plastic adherent
properties when maintained under standard culture conditions. They
express specific cell surface markers such as CD29, CD44, CD73 and
CD90 and can be differentiated into various mesenchymal linages
(14,15). The most common source of MSCs for
clinical use is human adult bone marrow; however, the
differentiating capacities are low with regard to the age of the
donor and the isolation method is considered an invasive technique
(8). Thus, different fetal tissues
have been studied for their potential an alternative source.
Amniotic fluid (AF) is a new putative source of MSCs which contain
a heterogeneous cell population derived from placental membranes
and fetal origins (15).
Human amniotic fluid (hAF) is a clear and yellowish
fluid that surrounds and protects the fetus in the uterus. It
contains a variety of cells that have generated from fetal origins
(15). Embryonic and fetal cells
from all three germ layers have long been identified in the AF
(16). Previous studies have
isolated and characterized these cells based on their morphology
and have reported that AF colonies consist of adherent
‘spindle’-shaped fibroblast-like cells and ‘round’-shaped
epithelioid cells, but the epithelial cells disappear during the
propagation of mixed primary cell cultures (15). The properties and types of AF cells
vary for different donors and the time needed for gestation and
cultivation. Moreover, these cells can be classified into three
types based on their morphological and growth characteristics:
Epitheloid type (E-type cells) which appear at the beginning of the
cultivation process and decrease during the cultivation process,
amniotic fluid-specific type (AF-type cells) which appear at the
beginning of the cultivation process and persist during the
cultivation process and fibroblast type (F-type cells), which are
believed to have originated from mesenchymal tissues (17). AF cells and amniotic fluid
derived-mesenchymal stem cells (AF-MSCs) are different because AF
cells are non-adhering cells, while AF-MSCs are cells that are
adhering cells and display dividing colonies (17). The AF-MSCs obtained from
amniocentesis in the second-trimester (16th-22nd weeks) of
pregnancy for prenatal diagnosis also express MSCs surface markers
such as hyaluronan receptor (CD44), Thy-1 (CD90), transforming
growth factor-beta receptor endoglin (CD105) and ES marker CD117
(14,18). Moreover, about 90% of the AF-MSCs
express the octamer-binding transcription factor 4 (Oct-4) that is
recognized as the transcription factor of embryonic stem cells and
can express pluripotent stem cells markers; TERT (17). The AF-MSCs represent the
intermediate stage between embryonic stem cells and adult stem
cells (18,19). The AF-MSCs have a high renewal
capacity and can be expanded for over 250 doublings without loss of
chromosomal telomere length (17).
The advantageous consequence of obtaining AF-MSCs is that it is
easy, safe and presents a low risk of destroying embryos.
Certainly, MSCs obtained from AF are recognized for presenting
little to no ethical objections and are useful in cell
transplantation in immunologically matched recipients (17,19).
Materials and methods
The cell samples and preparation
The hAF cell samples were received from the
16th-22nd weeks of gestation by amniocentesis for prenatal
diagnosis from the Human Genetics Laboratory, Department of
Anatomy, Faculty of Medicine, Chiang Mai University. The cell
samples displayed a normal karyotype (46, XX/46, XY). Written
informed consent was obtained after approval by the Ethics
Committee from the Faculty of Medicine, Chiang Mai University, 21st
April 2015 no. ANA-2558-2942.
In this study, the direct adherence method was used
to separate human amniotic fluid derived-mesenchymal stem cells
(hAF-MSCs) (20). Briefly, hAF
cells were centrifuged at 2,035 g for 6 min and then, plated in 25
cm2 flasks (Corning Incorporated, NY, USA) with
expansion medium (BIOAMF-3™ Complete Medium) (Biological
Industries, Kibbutz Beit Haemek, Israel) at 37°C, 5% CO2
and 95% humidity until the colonies of the adherent cells appeared.
After that, the basal growth medium, Dulbecco's Modified Eagle
Medium (DMEM)-high glucose (Gibco, USA) was replaced supplement
with 10% fetal bovine serum (FBS) (Gibco, South America),
gentamycin, Pen Strep (penicillin and streptomycin) (Gibco, USA) in
order to remove non-adherent cells from the supernatant. The medium
was changed every 3 days. After the cells reached 80% confluence,
they were sub-cultured using 0.25% trypsin-EDTA (Gibco, USA) and
re-expanded in a 25 cm2 flask under similar conditions
at a dilution of 1:2. The hAF cell samples at passage 3 were used
in the analysis.
Cell cultivation
The hAF cell samples in passage 3 (n=5) were washed
twice with sterile phosphate-buffered saline (PBS) (Amresco, Ohio,
USA) and trypsinized with 0.25% trypsin-EDTA. Subsequently, hAF
cells were suspended in the basal growth medium (DMEM-high glucose
with 10% FBS) and centrifuged at 2,035 g, for 6 min. After that,
the supernatant liquid was removed and the hAF cells were used in
the experiments.
The hAF cell samples were seeded in 24-well culture
plates (Corning Incorporated, NY, USA) at the density of
1×105 cells. Cells were divided into two groups under
different culture medium conditions. The control group was cultured
with basal growth medium and the cardiogenic induced group was
cultured with cardiogenic induced medium (DMEM-high glucose, 10%
FBS, gentamycin, Pen Strep + 10 µmol/l 5-aza (Sigma-Aldrich, St.
Louis, MO, USA)) (8). After 24 h,
the cardiogenic induced medium was changed to basal growth medium
and the culture medium was changed every 3 days for the duration of
the 21 days of cell culturing.
Flow cytometry
The hAF cell samples in passage 3 (n=3) were used to
determine MSCs markers expression. The cells were trypsinized with
0.25% trypsin-EDTA and centrifuged at 2,035 g for 6 min and then,
they were incubated with monoclonal antibodies; fluorescein
isothiocyanate (FITC)-conjugated mouse anti-human CD34, CD90
(Biolegend, San Diego, USA), mouse anti-human HLA-ABC (Immuno Tools
GmbH, Friesoythe, Germany) and mouse anti-human fibroblast (EDM,
Millipore Crop, UK), as well as phycoerythrin (PE)-conjugated mouse
anti-human CD31, CD117, HLA-DR (Immuno Tools GmbH, Friesoythe,
Germany), mouse anti-human CD44, CD105 (Pierce Biotechnology,
Rockford, USA), mouse anti-CD45 (Biolegend, San Diego, USA) and
mouse anti-human CD73 (Life Technologies, California, USA) for 60
min at 4°C. FITC mouse isotype control and PE mouse isotype control
(Biolegend, San Diego, USA) were used as negative controls. Cell
fluorescence was evaluated using FACscan (Becton Dickinson, Lincon
Park, NJ) and analyzed using the CellQuest Pro 9.0 software (Becton
Dickinson).
Alamar blue assay
This assay is a colorimetric indicator used to
measure living cell proliferation and cytotoxicity evaluation from
the oxidation-reduction of the living cell metabolism through the
conversion of resazurin (oxidised form, blue color) to resorufin
(reduced form, pink color) and to monitor the absorbance of alamar
blue at 540–630 nm. In brief, the cells in passage 3 (n=3) were
cultured in 24-well culture plates of 2×103 cells/well
with basal growth medium for 24 h. Thereafter, the basal growth
medium was removed and 100 µl 10% alamar blue in DMEM was added,
and then the samples were incubated at 37°C, 5% CO2 with
95% humidity for 4 h. After that, the supernatant liquid samples
were evaluated using the colorimetric change from each well, while
hAF cells were continuously cultivated under the same conditions.
Absorbance measurement was done on days 1, 3, 5, 7, 9, 11, 13, 15,
17, 19 and 21 of cell culturing using a spectrophotometer plate
reader (Original Multiskan EK, Thermoscientific, UK) and a sample
consisting of 10% alamar blue solution that was unexposed to the
cells was used as a control.
Immunofluorescence analysis
After 21 days of cells cultivation, the cells were
cultured on coverslips (Thermo Scientific, UK). The basal and
cardiogenic induced groups (n=5) were used to evaluate the
expression of cardiac specific proteins. After fixation for 30 min
at 4°C with 4% paraformaldehyde, the cell membranes were
permeabilized for 5 min with 0.2% triton X-100 (Amresco, Ohio, USA)
in PBS and blocked in 10% AB-serum in 1% bovine serum albumin (BSA)
in PBS for 30 min at 4°C. The cells were incubated with mouse
monoclonal primary antibodies against human GATA4, cardiac troponin
T (cTnT) and Nkx2.5 (Sigma-Aldrich, St. Louis, MO, USA) for 2 h at
4°C. After being washed with PBS, the cells were incubated with
goat anti-mouse secondary antibody conjugated with FITC (Thermo
Scientific, UK) for 60 min at 37°C. Subsequently, the cover-slips
were mounted onto the microscopic slides using anti-fade reagent
with 4′-6-diamidino-2-phenylindole (DAPI) (Invitrogen, USA). The
cells were visualized using a fluorescence microscope Olympus AX70.
Photographs were taken with DP manager and DP controller (Olympus
Life Science, USA).
Immunoenzymatic analysis
After 21 days of cell cultivation, the cells
cultured on coverslips (Thermo Scientific, UK) from the basal and
cardiogenic induced groups (n=5) were used to evaluate the
expression of cardiac specific proteins. After fixation for 30 min
at 4°C with 4% paraformaldehyde, the cells were blocked in 10%
AB-serum in 1% BSA in PBS for 30 min at 4°C, and then incubated
with rabbit monoclonal primary antibody against human connexin43
(Cx43) (Sigma-aldrich, St. Louis, MO, USA) for 2 h at 4°C. After
being washed with PBS, the cells were incubated with mouse
anti-rabbit peroxidase-conjugated secondary antibody (Immuno Tools
GmbH, Friesoythe, Germany) for 60 min at 37°C. Finally, the
immunoreaction was detected by using 3,3′-Diaminobenzidine (DAB)
liquid substrate system (Sigma-aldrich, St. Louis, MO, USA) and
analyzed under the Axiostar plus light microscope (Carl Zeiss,
Germany). Photographs were taken with Canon pc 1049 PowershotG5
(Canon, USA) and were used to determine and normalize the average
level of color intensity in both the basal and cardiogenic induced
groups using iSolution FL Auto ×64.
Reverse transcriptase-quantitative
polymerase chain reaction (RT-qPCR)
After 7 and 21 days of cell cultivation, total
ribonucleic acid (RNA) obtained from the basal and the cardiogenic
induced groups (n=5) were prepared using an Illutra RNAspin Mini
RNA Isolation kit (GE Healthcare, LittleChalfont, UK).
Complementary DNA (cDNA) was synthesized using the iScript™ cDNA
Synthesis kit (Bioline, USA) according to the manufacturer's
instructions. The RT-qPCR was performed using SsoFast™
EvaGreen® Supermix (Bio-Rad, Singapore) on a Chromo4™
Real-Time PCR Detector (Bio-Rad, USA) with gene specific primers
(Table I). The PCR mix contained
SsoFast™ EvaGreen® Supermix (Bio-Rad, Singapore), cDNA
template, primers, and nuclease free water to reach a final volume
of 20 µl. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH)
was used as the internal control gene for normalization. A melt
curve analysis was performed at the end of the reaction and the
profiles were obtained by plotting relative gene expression levels
of the both basal and cardiogenic induced groups.
 | Table I.The primers for RT-qPCR and products
size. |
Table I.
The primers for RT-qPCR and products
size.
| Gene (marker) | Primer
sequence | Size (bp) |
|---|
| GATA4 |
F5′-CTGTGCCAACTGCCACACCA-3′ | 437 |
|
|
R5′-GGCTGACCGAAGATGCGTAG-3′ |
|
| cTnT |
F5′-GGCAGCGGAAGAGGATGCTGAA-3′ | 150 |
|
|
R5′-GAGGCACCAAGTTGGGCATGAACGA-3′ |
|
| Cx43 |
F5′-GAATCCTGCTCCTGG-3′ | 380 |
|
|
R5′-GATGCTGATGATGTAG-3′ |
|
| Nkx2.5 |
F5′-CTGCCGCCGCCAACAAC-3′ | 136 |
|
|
R5′-CGCGGGTCCCTTCCCTACCA-3′ |
|
| GAPDH |
F5′-ATGGGGAAGGTGAAGGTCG-3′ | 70 |
|
|
R5′-TAAAAGCAGCCCTGGTGACC-3′ |
|
Statistical analysis
The data were analyzed using SPSS version 22 (SPSS,
Inc., Chicago, IL). Data are expressed as the mean ± SD. The
significance was analyzed using the Mann-Whitney U test. P<0.05
was considered to indicate a statistically significant
difference.
Results
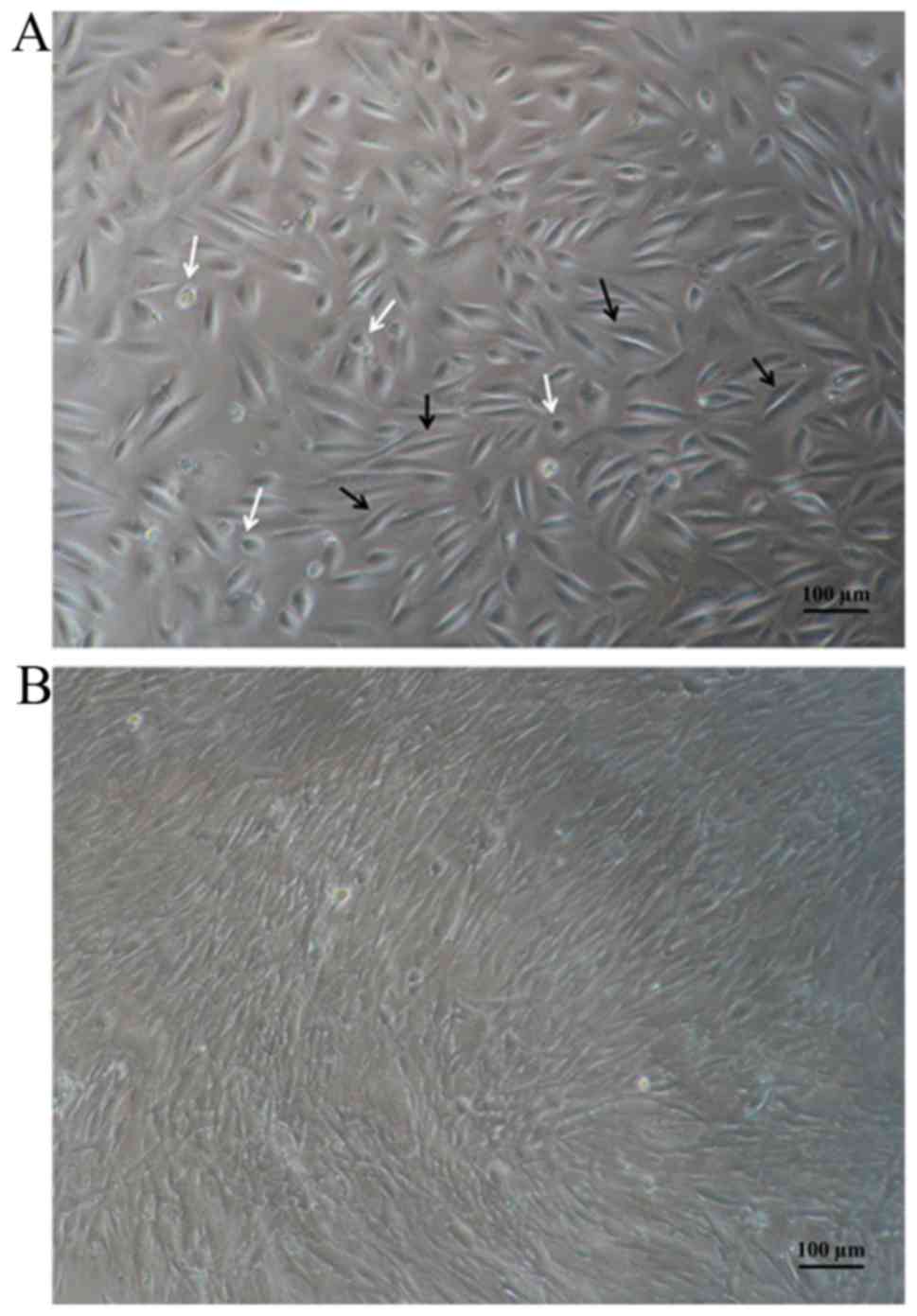
Morphology
The hAF cell samples were cultured with expansion
medium until the colonies of the adherent cells appeared. After
being subcultured, the hAF cells were investigated using inverted
microscopy. Inverted microscopy revealed that the hAF cell samples
were adhered to culture flasks and showed a heterogeneous
population. Moreover, the 3rd passages exhibited fibroblast-like
morphology (Fig. 1).
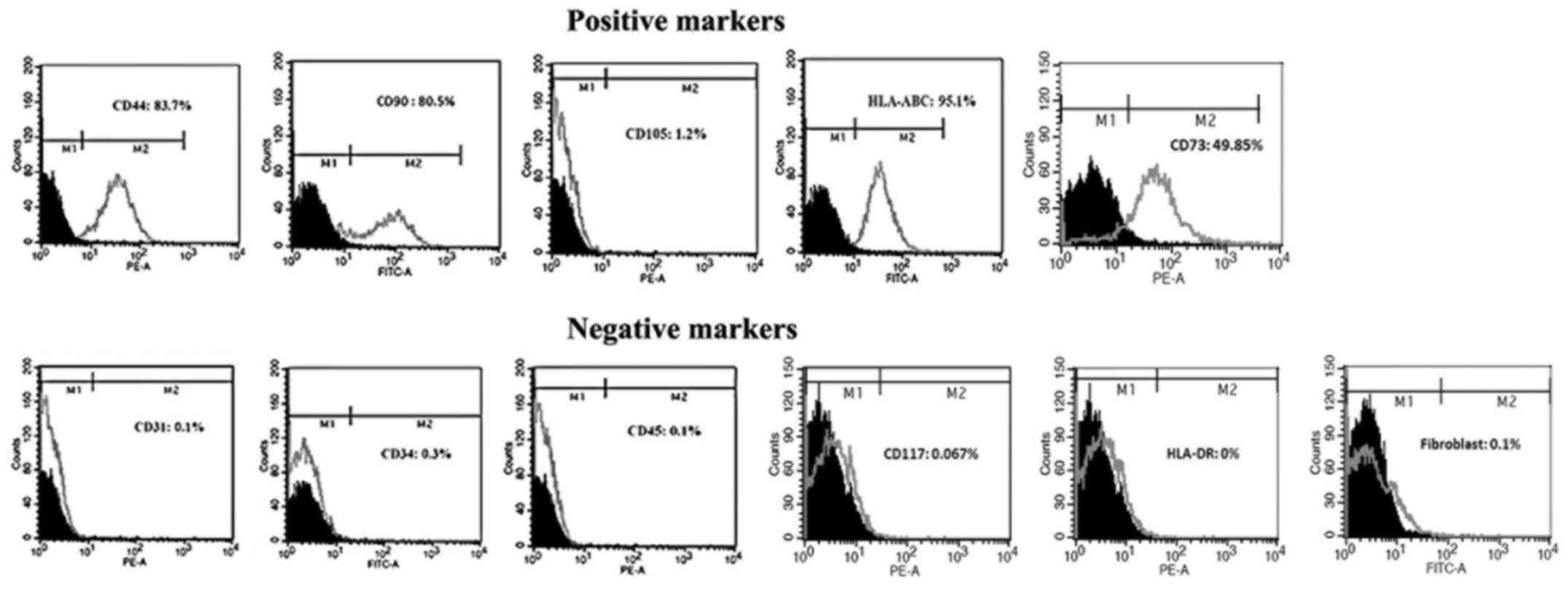
Flow cytometry
The hAF cells were characterized for MSCs surface
markers using flow cytometry analysis. The results showed that the
hAF cells were stained positively for MSCs surface markers CD44
(83.7%), CD90 (80.5%), HLA-ABC (95.1%) and CD73 (49.85%).
Interestingly, the hAF cells were not positively stained with
anti-CD105 (1.2%). As expected, the hAF cells were negatively
stained with platelet endothelial cell adhesion molecule CD31
(0.1%), heamatopoietic cell surface marker CD34 (0.3%) and CD45
(0.1%), amniotic fluid stem cells CD117 (0.067%), HLA-DR (0%) and
fibroblast (0.1%) (Fig. 2).
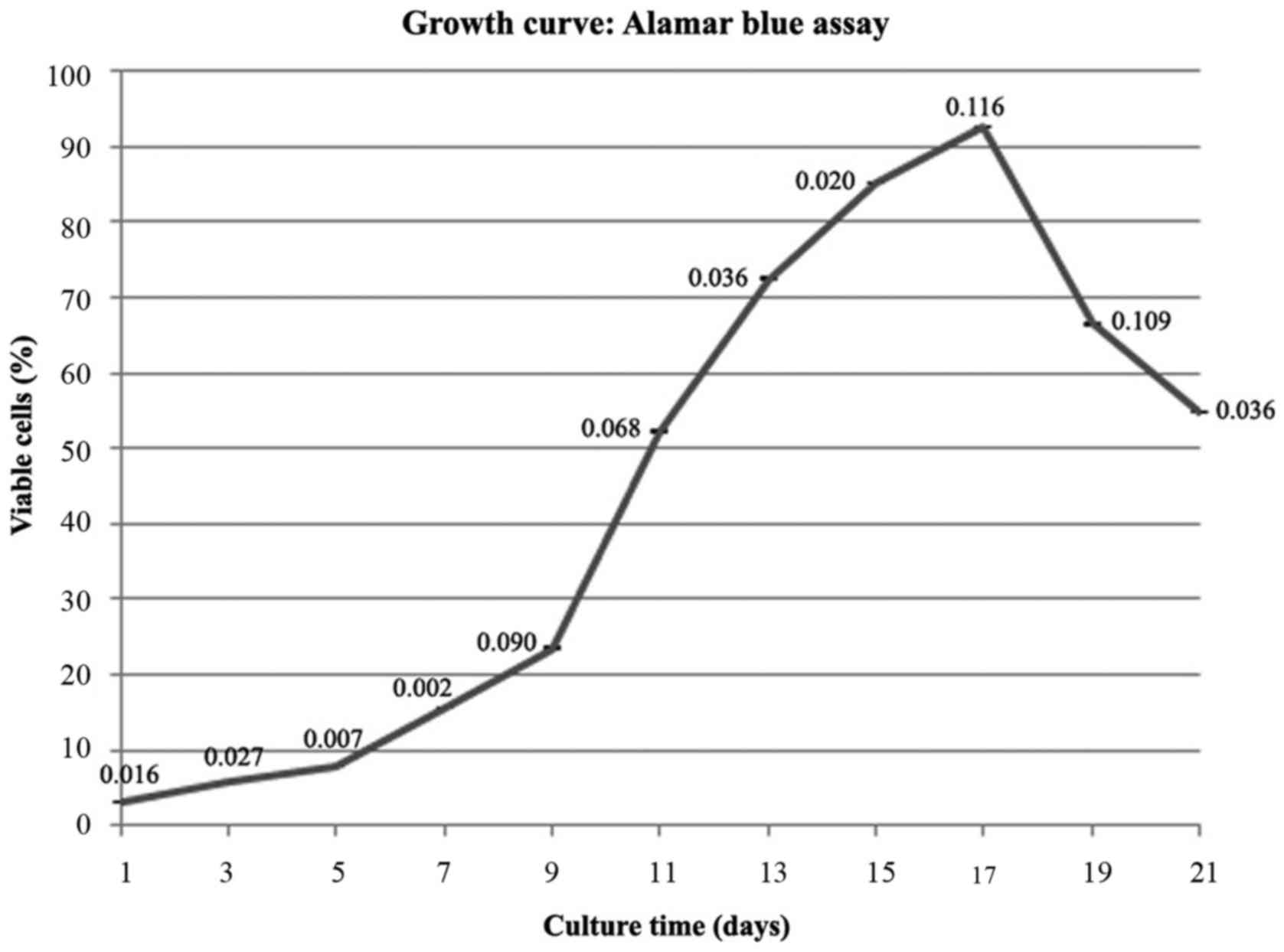
Alamar blue assay
The alamar blue assay was used to evaluate the
ability of cell proliferation. The results showed that the hAF cell
proliferation continuously increased from the early stage of
cultivation (Day 5) following the logarithmic phase. After that,
the cells began to proliferate swiftly and the cell population
doubled over 12 days (day 5-day 17). This event showed an
exponential phase of the cell cycle. The highest cell proliferation
had been observed on day 17 with 30-fold relative to day 1.
Afterward the cell numbers began to decrease (Fig. 3).
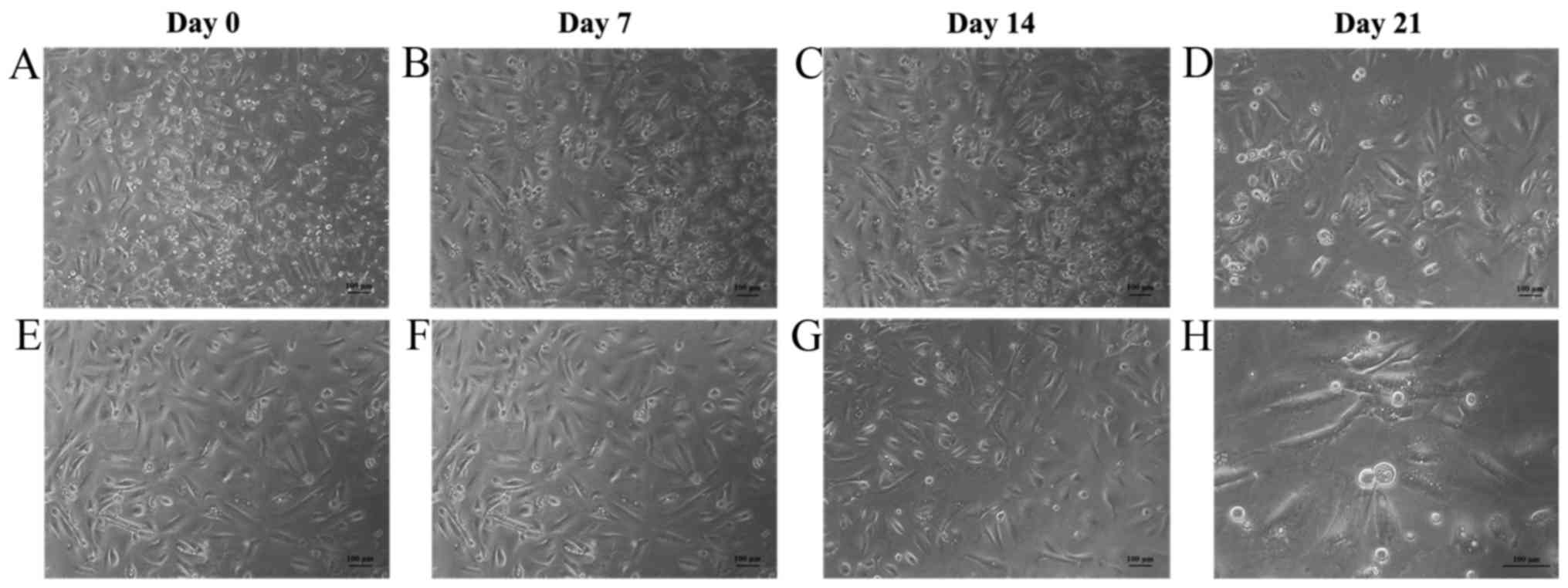
Cardiogenic differentiation
potential
In the hAF-MSCs in both the basal and cardiogenic
induced groups, morphological changes were observed using inverted
microscopy beginning at day 0 until day 21. The cells in the
cardiogenic induced group cultured with 5-aza showed elongated
myotube-like morphology and formed aggregations after 7 days of
culturing. The myotube-like morphology was maintained until 21 days
of cell culture. In the basal group, culture in the basal growth
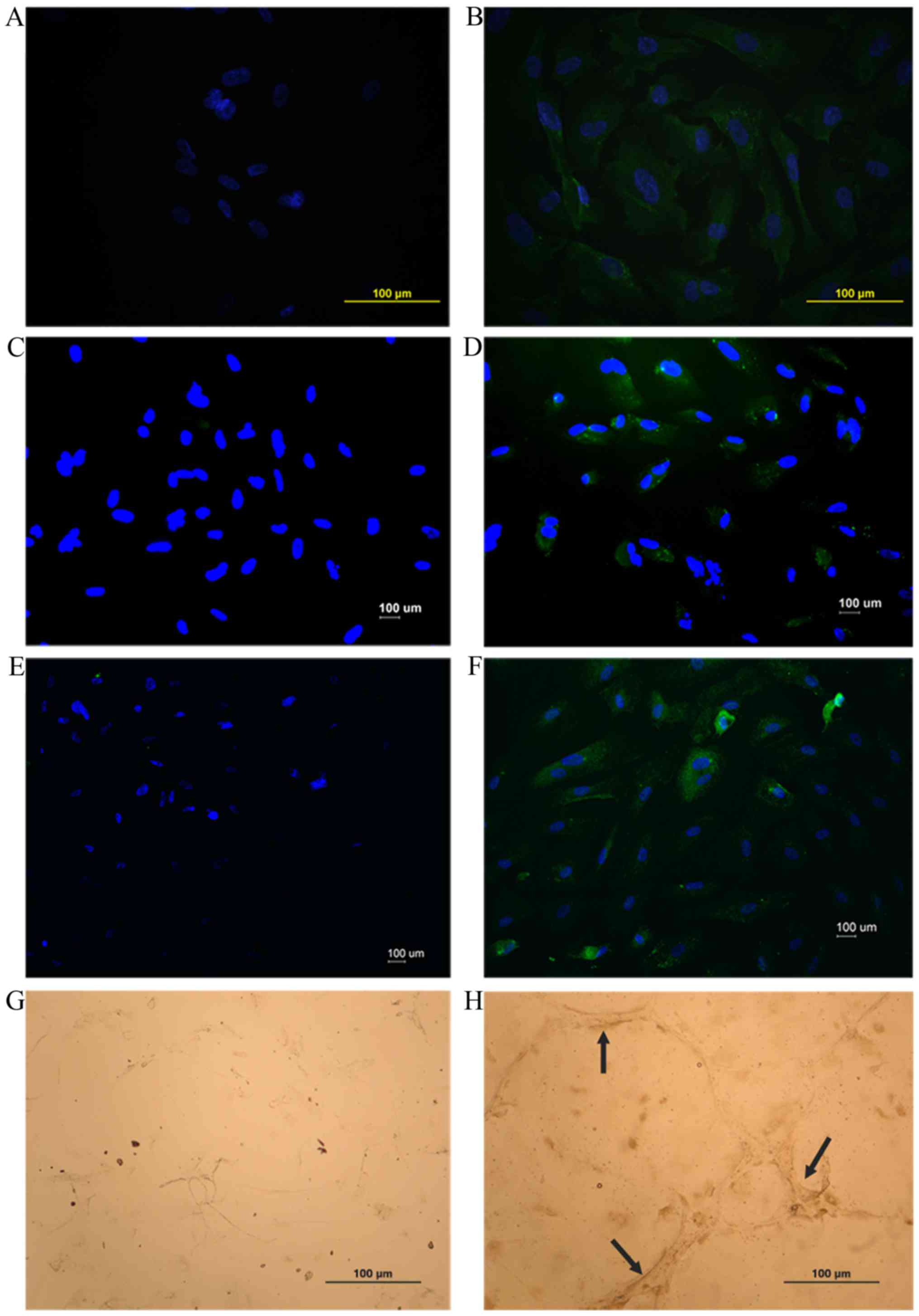
medium was maintained their fibroblastic-like morphology (Fig. 4). Immunofluorescence staining,
immunoenzymatic staining and RT-qPCR were used to determine the
cardiogenic differentiation potential of hAF-MSCs. After 21 days of
cell culturing, the cells that had been cultured on coverslips in
both groups (basal and cardiogenic induced groups) were
investigated for cardiogenic specific proteins (GATA4, cTnT, Nkx2.5
and Cx43). The results of the immunofluorescence staining and
immunoenzymatic staining indicated that the cardiogenic induced
group was strongly positive for the cardiac-specific markers GATA4,
cTnT, Nkx2.5 and Cx43. Interestingly, the results of the
immunofluorescence staining displayed positively stained GATA4,
cTnT and Nkx2.5 in the cardiogenic induced group, but it was not
positive in the basal group. The immunoenzymatic staining displayed
positively stained Cx43 in the cardiogenic induced group (73.5%),
but it was a very weak positive in the basal group (17.8%)
(Fig. 5). Moreover, the
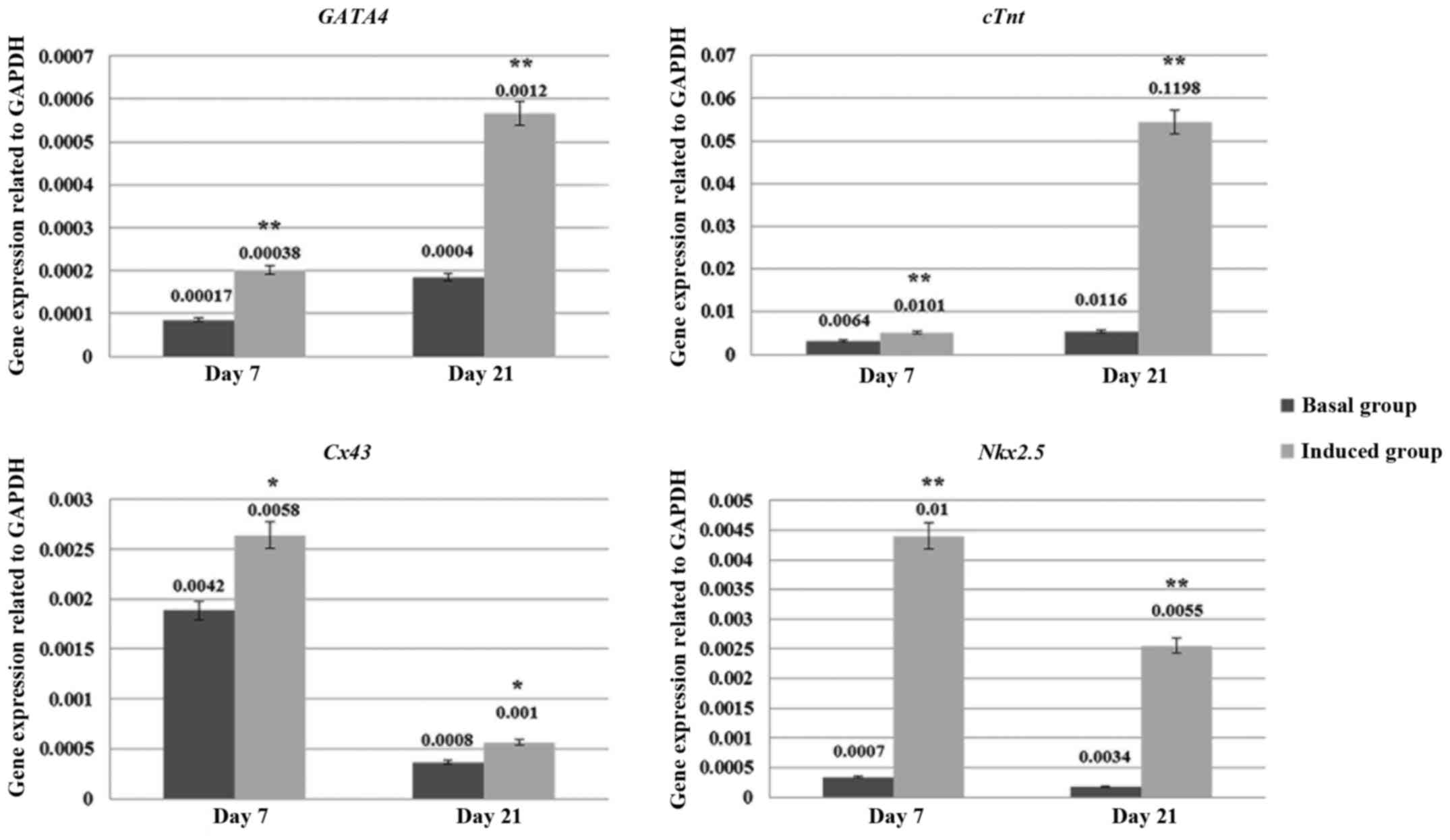
cardiogenic differentiation capacity of these hAF-MSCs was
confirmed by using RT-qPCR. After 7 and 21 days of cell culturing,
the cells in both groups (basal and cardiogenic induced groups)
were evaluated for cardiogenic specific genes (GATA4,
cTnT, Nkx2.5 and Cx43). The results of the RT-qPCR
indicated that the hAF-MSCs in the cardiogenic induced group could
express the cardiogenic specific genes that were related to
cardiogenesis. The cardiogenic induced group at days 7 and 21 had a
significant expression of cardiac specific genes that was greater
than the control group (P<0.05) (Fig. 6).
Discussion
Previous studies have shown that MSCs from bone
marrow can improve cardiac function in myocardial infarction
(21,22) and cardiogenic differentiation from
bone marrow MSCs has long been investigated (23,24).
However, the process of using MSCs obtained from this source has
displayed certain limitations including the invasive technique
needed to obtain MSCs from bone marrow, the decreased capacity of
these cells that correlate with the increased age of the donor
(25) and the obstacle of cell
transplantation therapy by immunological rejection (26). Thus, alternative sources of MSCs
have been studied and AF has recently become a new and interesting
source of MSCs. It contains a heterogenous population of cells in
the placental membrane as well in other areas stemming from its
fetal origin including the fetal skin, the epithelial, digestive
and respiratory tract, along with the urinary tract (15). AF is considered fairly easy to
collect by amniocentesis using ultrasound-guided transabdominal
puncture for prenatal diagnosis with a low risk of damaging the
mother or the embryos and low ethical concerns when compared with
other sources such as embryonic stem cells (17). Previous studies have demonstrated
that AF-MSCs are multipotent stem cells, which are capable of
self-renewal, high expansion rates, a high number of isolated cells
(27), and a low risk of
tumorigenicity (28). In addition,
AF-MSCs are capable of chondrogenic, osteogenic, adipogenic and
myogenic differentiation (14,29–31).
Over the years, MSCs have been isolated from various tissues and
organs including bone marrow, adipose tissue, skin, dental pulp,
placenta and the umbilical cord (32). According to previous studies, it
has been reported that mesenchymal amniocytes proliferate more
rapidly in vitro when compared to fetal and adult stem cells
(33,34). In vitro, MSCs usually grow
as a monolayer culture in a medium supplemented with FBS (35). Furthermore, the hAF-MSCs expressed
both embryonic and adult stem cell markers. The ability of this
cell has been acknowledged at the intermediate stage of embryonic
stem cells and adult stem cells (18,19,36).
These cells may present more advantages than adult stem cells and
show a wide differentiation of properties that can induce
mesenchymal linages (14).
The hAF cells showed heterogenous population
colonies and these cells exhibited fibroblast-like morphology in
the 3rd passage. They adhered to the culture flask, a feature that
resembled previous studies and it was concluded that hAF-MSCs could
easily be expanded without feeder cells, exhibit fibroblastoid
spindle shape morphology (29,31,32)
and could expose plastic adherents when maintained under standard
culture conditions (14).
The present study aimed to characterize hAF-MSCs and
evaluate the cardiogenic differentiation capacity of the hAF-MSCs.
Flow cytometry was used to determine the surface antigens of the
MSCs (Cut). It has been previously reported that hAF-MSCs strongly
expressed MSCs markers such as CD29, CD44, CD73, CD90, CD144,
CD166, and HLA-ABC (37–40). Moreover, hAF-MSCs showed a positive
co-expression of pluripotent markers such as Otc-4 (19), embryonic antigens Nanog and
(SSEA)-4 (12,32,38).
The results of our experiments showed that the hAF cells in passage
3 were strongly positive for MSCs surface markers including the
transmembrane glycoprotein CD44, the hyaluronan receptor which
plays a role in MSCs migration, membrane glycoprotein CD73 that is
often used for MSCs marker, membrane glycoprotein CD90 or Thy-1
that is often used as a marker for several kinds of stem cells such
as keratinocyte stem cells, endometrial stem cells and mesenchymal
stromal cells (32,41,42)
and the HLA-ABC (HLA class I) marker. On the other hand, these
cells were negative for platelet endothelial cell adhesion marker
CD31, hematopoietic cell surface markers including CD34 and CD45,
CD117 or c-kit that found in amniotic fluid stem cells (43), HLA-DR and fibroblast marker.
Furthermore, this investigation found that the MSCs surface antigen
was different. The percentage of positive expression was between
80–95% in each marker and excluded CD105, transforming growth
factor-beta receptor endoglin, or SH2 expression of 1.2%. In
previous studies, it has been reported that the percentage of CD105
expression was varied among the different cell populations in the
amniotic fluid (36). CD105 is a
proliferation-associated and hypoxia-inducible protein that is
abundantly expressed in the angiogenic endothelial cell, while
monocyte and immunohistochemistry studies have revealed that CD105
is strongly expressed in blood vessels of tumor tissues (44). Moreover, the expression of CD105
depends on the serum supplemented in the culture medium, cell
passage, age of gestation, maternal age and source of MSCs
(15,45,46).
All factors influenced the different levels of surface antigen
expression (47). The hAF cells
were strongly positive for HLA-ABC (HLA class I) and negative for
HLA-DR (HLA class II), which suggested that this cell has the
potential for clinical applications. The previous study showed that
cardiomyocytes differentiated from AF-MSCs transplantation and may
be helpful in avoiding unexpected events such as calcification
(48).
In this study, the determination of hAF-MSCs
proliferation using alamar blue assay exposed the continuously
increased from the early cultivation phase up to 5-fold during day
1 to 5 of cell culturing, following a logarithmic phase or the
induction phase of the cell cycle. After that, the cells started to
proliferate swiftly and the cell population doubled from day 5 to
day 17. The highest cell proliferation had been observed on day 17
with 30-fold relative to day 1. This event showed an exponential
phase of the cell cycle. Afterward, the cell numbers began to
decrease. These results indicate that these cells had a
proliferation capacity, which is related to MSCs growth and
proliferation properties. In agreement with previous study, the
hAF-MSCs presented a slow rate of proliferation in the first week
of cell cultivation, after which the proliferation swiftly
increased; this finding was different in AF cells that were derived
from various gestational stages (16,33,47).
The MSCs revealed a multipotency capability
differentiation with regard to the variety of cell types of all
three embryonic germ layers (14,15)
including cardiomyocytes (25). In
this study, the hAF-MSCs were induced to cardiomyocyte-like cells
using 5-aza for 24 h. (Cut) The morphology was observed from day 0
until day 21 of cell culturing in both the basal and cardiogenic
induced groups. From previous studies have been reported that, the
MSCs morphology were changed to myotube-like structure after
induced with 5-aza for 24 h and maintained in standard culture
condition for 7–10 days (7,11).
In this study the cells in the cardiogenic induced group showed the
myotube-like morphology and formed aggregations after 7 days of
culturing. The myotube-like morphology was maintained until day 21
and related to the cardiac specific gene expression on day 7 and
day 21 of cell culturing. After day 21, cells in both the basal and
cardiogenic induced groups were examined for cardiac specific
proteins using immunofluorescence and immunoenzymatic staining.
Expression of the cardiac specific genes was detected using RT-qPCR
on days 7 and 21 of cell culturing. The results showed that the
cardiogenic induced group strongly expressed the cardiac specific
proteins (GATA4, cTnT, Nkx2.5 and Cx43) and genes (GATA4, cTnT,
Nkx2.5 and Cx43) (P<0.05). These were highly
expressed in the cardiogenic induced group, which was different
from the basal group. GATA4 is an important transcription factor in
term of early myocardial development, which is expressed in the
cytoplasm. Moreover, the expression of GATA4 increases the
potential of MSCs to be differentiated into cardiomyocytes
(49). The cTnT is a protein which
functions as a part of the troponin complex of myofibrils only in
the cardiac muscles and it is expressed in the cytoplasm (50,51).
Nkx2.5 is a transcription factor for cardiac development, which
play an important role in early cardiac development and express in
the cytoplasm (51). Cx43 is one
variety of the gap junction proteins, which found in the atrial and
ventricular myocytes (52) and
maintains electrical activity (9).
GATA4, cTnT, Nkx2.5 and Cx43 promote the differentiation of MSCs
into cardiomyocytes and are required for cardiogenic
differentiation (53,54). Moreover, GATA4 and Nkx2.5 are the
greatest transcription factors that have major regulatory roles in
the cardiogenic differentiation of MSCs (9). All of the results in this study
indicated a strong correlation with the early evaluation of
cardiogenic differentiation of AF-MSCs and corresponded with
previous studies, which reported that AF-MSCs can be expanded in
vitro and can be differentiated into cardiomyocytes with a
strong expression of cardiac specific proteins and genes (12,55,56).
5-aza is a DNA methylation inhibitor that is mainly
used in the treatment of myelodysplastic syndrome (MDS) and it can
induce MSCs into cardiomyocytes. In previous studies, it has been
reported that 5-aza induced the up-regulation of phosphorylated
cardiac specific genes through the ERK pathway and these results
suggested that the sustained activation of ERK by 5-aza contributed
to the induction of the differentiation of MSCs into cardiomyocytes
in vitro (57).
Additionally, 5-aza can change the morphology of hAF-MSCs from a
spindle shape to myotube-like morphology at about 7 days of cell
culturing and maintained this until day 21. The results of the
morphological changes can confirm cardiogenic differentiation with
cardiac specific proteins expression on day 21 and genes expression
on day 7 and day 21. (Cut).
In conclusion, our study has demonstrated that AF is
an important source of MSCs that can be obtained from the routine
amniocentesis investigation. These cells are easily isolated and
can be grown in vitro without feeder layers. In addition,
they show the characteristics of multipotency MSCs and demonstrate
the ability of increased proliferation. Nevertheless, they exhibit
the capacity and potential for cardiogenic differentiation. The
data obtained suggests that they are worthy of further
investigation in term of the study of the regenerative medicine in
stem cell transplantation and tissue engineering.
Acknowledgements
This research study was supported by the Faculty of
Medicine Research Fund, Chiang Mai University, Chiang Mai,
Thailand.
References
|
1
|
Kiatchoosakun S, Sutra S and
Thepsuthammarat K: Coronary artery disease in the Thai population:
Data from health situation analysis 2010. J Med Assoc Thai. 95
Suppl 7:S149–S155. 2012.PubMed/NCBI
|
|
2
|
Hansen JT and Lambert DR: Netter's
clinical anatomy. Elsevier; Philadelphia: pp. 5–7. 2010
|
|
3
|
Fukuda K: Development of regenerative
cardiomyocytes from mesenchymal stem cells for cardiovascular
tissue engineering. Artif Organs. 25:187–193. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mescher AL: Junqueira's Basic Histology.
McGraw-Hill; New York: pp. 1842010
|
|
5
|
Passier R and Mummery C: Cardiomyocyte
differentiation from embryonic and adult stem cells. Curr Opin
Biotechnol. 16:498–502. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kadivar M, Khatami S, Mortazavi Y,
Shokrgozar MA, Taghikhani M and Soleimani M: In vitro
cardiomyogenic potential of human umbilical vein-derived
mesenchymal stem cells. Biochem Biophys Res Commun. 340:639–647.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Makino S, Fukuda K, Miyoshi S, Konishi F,
Kodama H, Pan J, Sano M, Takahashi T, Hori S, Abe H, et al:
Cardiomyocytes can be generated from marrow stromal cells in vitro.
J Clin Invest. 103:697–705. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nartprayut K, U-Pratya Y, Kheolamai P,
Manochantr S, Chayosumrit M, Issaragrisil S and Supokawej A:
Cardiomyocyte differentiation of perinatally-derived mesenchymal
stem cells. Mol Med Rep. 7:1465–1469. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shen H, Wang Y, Zhang Z, Yang J, Hu S and
Shen Z: Mesenchymal stem cells for cardiac regenerative therapy:
Optimization of cell differentiation strategy. Stem Cells Int.
2015:5247562015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Singh A, Singh A and Sen D: Mesenchymal
stem cells in cardiac regeneration: A detailed progress report of
the last 6 years (2010–2015). Stem Cell Res Ther. 7:822016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wakitani S, Saito T and Caplan AI:
Myogenic cells derived from rat bone marrow mesenchymal stem cells
exposed to 5-azacytidine. Muscle Nerve. 18:1417–1426. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bai J, Hu Y, Wang YR, Lui LF, Chen J, Su
SP and Wang Y: Comparison of human amniotic fluid-derived and
umbilical cord Wharton's jelly-derived mesenchymal stromal cells:
Characterization and myocardial differentiation capacity. J Geriatr
Cardiol. 9:166–171. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim J, Lee Y, Kim H, Hwang KJ, Kwon HC,
Kim SK, Cho DJ, Kang SG and You J: Human amniotic fluid-derived
stem cells have characteristics of multipotent stem cells. Cell
Prolif. 40:75–90. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dominici M, Le Blanc K, Muller I,
Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A,
Prockop Dj and Horwitz E: Minimal criteria for defining multipotent
mesenchymal stromal cells. The International Society for Cellular
Therapy position statement. Cytotherapy. 8:315–317. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Savickiene J, Treigyte G, Baronaite S,
Valiuliene G, Kaupinis A, Valius M, Arlauskiene A and Navakauskiene
R: Human amniotic fluid mesenchymal stem cells from second- and
third-trimester amniocentesis: Differentiation potential, molecular
signature, and proteome analysis. Stem Cells Int. 2015:3192382015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fauza D: Amniotic fluid and placental stem
cells. Best Pract Res Clin Obstet Gynaecol. 18:877–891. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Antonucci I, Stuppia L, Kaneko Y, Yu S,
Tajira N, Bae EC, Chheda SH, Weinbren NL and Borlongan CV: Amniotic
fluid as a rich source of mesenchymal stromal cells for
transplantation therapy. Cell Transplant. 20:789–795. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rossi B, Merlo B, Colleoni S, Iacono E,
Tazzari PL, Ricci F, Lazzari G and Galli C: Isolation and in vitro
characterization of bovine amniotic fluid derived stem cells at
different trimesters of pregnancy. Stem Cell Rev. 10:712–724. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Prusa AR, Marton E, Rosner M, Bernaschek G
and Hengstschläger M: Oct-4 expressing cells in human amniotic
fluid: A new source for stem cell research. Hum Reprod.
18:1489–1493. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fei X, Jiang S, Zhang S, Li Y, Ge J, He B,
Goldstein S and Ruiz G: Isolation, culture, and identification of
amniotic fluid-derived mesenchymal stem cells. Cell Biochem
Biophys. 67:689–694. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bartunek J, Croissant JD, Winjns W,
Gofflot S, de Lavareille A, Vanderheyden M, Kaluzhny Y, Mazouz N,
Willemsen P, Penicka M, et al: Pretreatment of adult bone marrow
mesenchymal stem cells with cardiomyogenic growth factors and
repair of the chronically infracted myocardium. Am J Physiol Heart
Circ Physiol. 292:H1095–H1104. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hahn JY, Cho HJ, Kang HJ, Kim MH, Chung
JH, Bae JW, Oh BH, Park YB and Kim HS: Pre-treatment of mesenchymal
stem cells with a combination of growth factors enhances gap
junction formatio, cytoprotective effect on cerdiomyocytes, and
therapeutic efficacy for myocardial infraction. J Am Coll Cardiol.
51:933–943. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Khanabdali R, Saadat A, Fazilah M, Bazli
KF, Qazi RE, Khalid RS, Adli DS Hasan, Moghadamtousi SZ, Naeem N,
Khan I, et al: Promoting effect of small molecules in
cardiomyogenic and neurogenic differentiation of rat bone
marrow-derived mesenchymal stem cells. Drug Des Devel Ther.
10:81–91. 2015.PubMed/NCBI
|
|
24
|
Kumar BM, Maeng GH, Lee YM, Kim TH, Lee
JH, Jeon BG, Ock SA, Yoo JG and Rho GJ: Neurogenic and
cardiomyogenic differentiation of mesenchymal stem cells isolated
from minipig bone marrow. Res Vet Sci. 93:749–757. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Macias MI, Grande J, Moreno A, Domínguez
I, Bornstein R and Flores AI: Isolation and characteriazation of
true mesenchymal stem cells derived from human term decidua capable
of multilineage differentiation into all 3 embryonic layers. Am J
Obstet Gynecol. 203:495.e9–e495.e23. 2010. View Article : Google Scholar
|
|
26
|
Dai W and Kloner RA: Myocardial
regeneration by human amniotic fluid stem cells: Challenges to be
outcome. J Mol Cell Cardiol. 42:730–732. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ferdaos N and Nordin N: Human amniotic
fluid cells and their future perspectives. Regenerative Research.
1:14–19. 2012.
|
|
28
|
Rosner M, Dolznig H, Schipany K, Mikula M,
Brandau O and Hengstschläger M: Human amniotic fluid stem cells as
a model for functional studies of genes involved in human genetic
diseases or oncogenesis. Oncotarget. 2:705–712. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chao KC, Yang HT and Chen MW: Human
umbilical cord mesenchymal stem cells suppress breast cancer
tumourigenesis through direct cell-cell contact and
internalization. J Cell Mol Med. 16:1803–1815. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tsai MS, Hwang SM, Tsai YL, Cheng FC, Lee
JL and Chang Y: Clonal amniotic fluid-derived stem cells express
characteristics of both mesenchymal and neural stem cells. Biol
Reprod. 74:545–551. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao F and Ma T: Perfusion bioreactor
system for human mesenchymal stem cell tissue engineering: Dynamic
cell seeding and construct development. Biotechnol Bioeng.
91:482–493. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
De Coppi P, Bartsch G Jr, Siddiqui MM, Xu
T, Santos CC, Perin L, Mostoslavsky G, Serre AC, Snyder EY, Yoo JJ,
et al: Isolation of amniotic stem cell lines with potential for
therapy. Nat Biotechnol. 25:100–106. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kaviani A, Perry TE, Dzakovic A, Jennings
RW, Ziegler MM and Fauza DO: The amniotic fluid as a source of
cells for fetal tissue engineering. J Pediatr Surg. 36:1662–1165.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Trohatou O, Anagnou NP and Roubelakis MG:
Human amniotic fluid stem cells as an attractive tool for clinical
applications. Curr Stem Cell Res Ther. 8:125–132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Almalki SG and Agrawal DK: Key
transcription factors in the differentiation of mesenchymal stem
cells. Differentiation. 92:41–51. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hartmann K, Raabe O, Wenisch S and Arnhold
S: Amniotic fluid derived stem cells give rise to neuron-like cells
without a further differentiation potential into retina-like cells.
Am J Stem Cells. 2:108–118. 2013.PubMed/NCBI
|
|
37
|
Bossolasco P, Montemurro T, Cova L,
Zangrossi S, Calzarossa C, Buiatiotis S, Soligo D, Bosari S, Silani
V, Deliliers GL, et al: Molecular and phenotypic characterization
of human amniotic fluid cells and their differentiation potential.
Cell Res. 16:329–336. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mareschi K, Rustichelli D, Comunanza V, De
Fazio R, Cravero C, Morterra G, Martinoglio B, Medico E, Carbone E,
Benedetto C and Fagioli F: Multipotent mesenchymal stem cells from
amniotic fluid originate neural precursors with functional
voltage-gated sodium channels. Cytotherapy. 11:534–547. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
In't Anker PS, Scherjon SA, Kleijburg-van
der Keur C, Noort WA, Claas FH, Willemze R, Fibbe WE and Kanhai HH:
Amniotic fluid as a novel source of mesenchymal stem cells for
therapeutic transplantation. Blood. 102:1548–1549. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pievani A, Scagliotti V, Russo FM, Azario
I, Rambaldi B, Sacchetti B, Marzorati S, Erba E, Giudici G,
Riminucci M, et al: Comparative analysis of multilineage properties
of mesenchymal stromal cells derived from fetal sources shows an
advantage of mesenchymal stromal cells isolated from cord blood in
chondrogenic differentiation potential. Cytotherapy. 16:893–905.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Barry FP and Murphy JM: Mesenchymal stem
cells: Clinical applications and biological characterization. Int J
Biochem Cell Biol. 36:568–584. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
L Ramos T, Sánchez-Abarca LI, Muntión S,
Preciado S, Puig N, López-Ruano G, Hernández-Hernández Á, Redondo
A, Ortega R, Rodríguez C, et al: MSC surface markers (CD44, CD73,
and CD90) can identify human MSC-derived extracellular vesicles by
conventional flow cytometry. Cell Commun Signal. 14:22016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cananzi M and De Coppi P: CD117(+)
amniotic fluid stem cells: State of the art and future
perspectives. Organogenesis. 8:77–88. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Duff SE, Li C, Garland JM and Kumar S:
CD105 is important for angiogenesis: Evidence and potential
applications. FASEB J. 17:984–992. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Maleki M, Ghanbarvand F, Behvarz M Reza,
Ejtemaei M and Ghadirkhomi E: Comparison of mesenchymal stem cell
markers in multiple human adult stem cells. Int J Stem Cells.
7:118–126. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mark P, Kleinsorge M, Gaebel R, Lux CA,
Toelk A, Pittermann E, David R, Steinhoff G and Ma N: Human
mesenchymal stem cells display reduced expression of CD105 after
culture in serum-free medium. Stem Cells Int. 2013:6980762013.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Carraro G, Garcia OH, Perin L, Filippo RD
and Warburton D: Amniotic fluid stem cells. Amniotic fluid stem
cells, Embryonic stem cells-Differentiation and pluripotent
alternativesEmbryonic Stem Cells-Differentiation and Pluripotent
Alternatives. Kallos MS: IN TECH; Croatia: pp. 1–506. 2011
|
|
48
|
Yoon YS, Park JS, Tkebuchava T, Luedeman C
and Losordo DW: Unexpected severe calcification after
transplantation of bone marrow cells in acute myocardial
infraction. Circulation. 109:3154–3157. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Li H, Zuo S, He Z, Yang Y, Pasha Z, Wang Y
and Xu M: Paracrine factors released by GATA-4 overexpressed
mesenchymal stem cells increase angiogenesis and cell survival. Am
J Physiol Heart Circ Physiol. 299:H1772–H1781. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Oma Y and Harata M: Actin-related proteins
localized in the nucleus: From discovery to novel roles in nuclear
organizeation. Nucleus. 2:38–46. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Stennard FA, Costa MW, Elliott DA, Rankin
S, Haast SJ, Lai D, McDonald LP, Niederreither K, Dolle P, Bruneau
BG, et al: Cardiac T-box factor Tbx20 directly interacts with
Nkx2-5, GATA4, and GATA5 in regulation of gene expression in the
developing heart. Dev Biol. 262:206–224. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Laird DW: The gap junction proteome and
its relationship to diesase. Trends Cell Biol. 20:92–101. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Armiñán A, Gandía C, García-Verdugo JM,
Lledó E, Mullor JL, Montero JA and Sepúlveda P: Cardiac
transcription factors driven lineage-specification of adult stem
cells. J Cardiovasc Transl Res. 3:61–65. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yamada Y, Sakurada K, Takeda Y, Gojo S and
Umezawa A: Single-cell-derived mesenchymal stem cells
overexpressing Csx/Nkx2.5 and GATA4 undergo the stochastic
cerdiomyogenic fate and behave like transient amplifying cells. Exp
Cell Res. 313:698–706. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen J, Lu Z, Cheng D, Peng S and Wang H:
Isolation and characterization of porcine amniotic fluid-derived
multipotent stem cells. PLoS One. 6:e199642011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Connell J Petsche, Camci-Unal G,
Khademhosseini A and Jacot JG: Amniotic fluid-derived stem cells
for cardiovascular tissue engineering applications. Tissue Eng Part
B Rev. 19:368–379. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Qian Q, Qian H, Zhang X, Zhu W, Yan Y, Ye
S, Peng X, Li W, Xu Z, Sun L and Xu W: 5-Azacytidine induces
cardiac differentiation of human umbilical cord-derived mesenchymal
stem cells by activating extracellular regulated kinase. Stem Cells
Dev. 21:67–75. 2012. View Article : Google Scholar : PubMed/NCBI
|