Introduction
Epithelial-mesenchymal transition (EMT) endows cells
with increased motility and invasiveness during the development of
diseases, especially tumor and fibrosis (1,2).
Tumor growth factor (TGF)-β can boost tumor or fibrosis progression
by enhancing proliferation, migration, and invasion partly because
it can induce EMT (3,4). However, the underlying molecular
mechanisms that mediate EMT remain unclear. Thus, enhancing our
understanding of these molecular mechanisms lead to the development
of effective EMT-targeted therapy and improvement of the diagnosis,
treatment, and prognosis of tumor or fibrosis (5,6).
Emerging studies showed that noncoding RNAs
(ncRNAs), such as microRNAs (miRNAs) and long noncoding RNAs
(lncRNAs), participate in many critical biological processes, such
as cell proliferation, apoptosis, and differentiation (7–9).
Furthermore, the regulatory network of ncRNAs participates in the
development of various illnesses; in particular, miRNAs inhibit the
crucial process of target RNA transcription (10–13),
and lncRNAs exhibit facilitative or suppressive effects on the
gene-regulatory network (14–16).
However, the relationships among messenger RNAs (mRNAs), miRNAs,
and lncRNAs remain unclear. A competing endogenous RNA (ceRNA)
mechanism that partly explains their mutual regulation of mRNAs,
miRNAs, and lncRNAs is hypothesized to occur in a disordered
manner. Furthermore, lncRNAs, such as GAS5 and glucocorticoid
receptor (18), GADD7 and TDP-43
(19), and PANDA and NF-YA
(20), that inhibit proteins
through competitive binding miRNA were reported (17). Nevertheless, owing to the generally
low abundance of lncRNAs and frequent promiscuity of protein-RNA
interactions, the extent to which lncRNAs function through this
mechanism is highly debated (21).
Thus, characterizing the interactions among mRNA,
miRNA, and lncRNA can enhance our understanding of disease
pathogenesis. Our previous studies reported different lncRNA
profiles in bleomycin-induced rats (22,23).
However, the rats' genes showed a high degree of difference from
its human homologs at lncRNA, miRNA, and mRNA levels. Thus, their
interactions cannot be analyzed completely. In addition, TGF-β1 is
the prototypic growth factor for the induction of EMT-like changes
in tumor or fibrosis (1,2). Thus, determining what changes can
occur during the interactions among mRNAs, miRNAs, and lncRNAs upon
TGF-β1 stimulation during EMT is important. The A549 cell line
displays properties similar to those of human type II pulmonary
epithelial cells (24) and is thus
often used as the normal model for studying pulmonary fibrosis or
other pulmonary diseases (25–27).
In the present study, the interactions among coexpressed mRNAs,
miRNAs, and lncRNAs were assessed in TGF-β1-activated EMTs of A549
cells. The results demonstrated that networks mediated by
EMT-associated mRNAs, miRNAs, and lncRNAs can be used to accelerate
the discovery of molecular mechanisms and EMT-targeted therapeutics
for pulmonary tumor and fibrosis.
Materials and methods
Cell model
A549 and MRC-5 cell lines were purchased from Cell
Bank of Chinese Academy of Sciences (Shanghai, China). Cells were
maintained in Dulbecco's modified Eagle's medium and advanced
minimum essential medium, respectively, supplemented with 10%
newborn calf serum, 100 U/ml penicillin, and 100 µg/ml streptomycin
at 37 °C under a humidified atmosphere of 5% CO2 and 95%
air. A549 or MRC-5 cells were collected after treatment with 10
ng/ml TGF-β1 for 12, 24, and 48 h as our previously described
(28–30).
Differentially expressed mRNAs, miRNAs
and lncRNAs
Total RNA was harvested and quantified, and its
integrity was verified by denaturing gel electrophoresis. Samples
with a 28S:18S ratios of approximately 2:1 were accepted for
further analysis, which was performed on an Affymetrix Human
Transcriptome Array 2.0 microarray for mRNAs and lncRNAs and
Affymetrix Gene Chip miRNA 4.0 microarray for miRNAs. The detection
of mRNAs, miRNAs, and lncRNAs was performed according to the
manufacturer's instructions. Finally, differentially expressed
mRNAs, miRNAs, and lncRNAs were identified through fold-change
filtering after the quantile normalization of raw data.
Bioinformatics analysis of
differentially expressed mRNA
Gene ontology (GO) analysis was applied to predict
the main functions of the target genes according to the GO project.
Fisher's exact test and χ2 were used to classify the GO
category. Moreover, false discovery rate (FDR) was calculated to
correct the P-value. The smaller the FDR, the smaller the error in
judging the P-value. The standard difference screening was
P<0.01.
Pathway analysis was performed to determine the
significant pathway of the differential genes according to the
Kyoto Encyclopedia of Genes and Genomes (KEGG), Biocarta, and
Reatome. Fisher's exact test and χ2 results were used to
select the significant pathway, and the significance threshold was
defined by P-value and FDR. The standard difference screening was
P<0.05.
MiRNA-target gene network was established on the
basis of GO and KEGG predicted data for the illustration of the
relationship between miRNAs and their target genes.
Quantitative real-time PCR
(qRT-PCR)
Total RNA was isolated with TRIzol reagent
(Invitrogen, Carlsbad, CA, USA). RNA quantity and quality were
measured on a NanoDrop 2000 spectrophotometer (Thermo Fisher
Scientific, Waltham, MA, USA). RNA integrity was assessed by
standard denaturing agarose gel electrophoresis. Complementary DNA
synthesis was performed with a Moloney Murine Leukemia Virus
reverse transcriptase kit (Invitrogen) following the manufacturer's
instructions. qRT-PCR was performed with a SYBR-Green-based PCR
master mix kit (Takara, Shiga, Japan) on a Rotor Gene 3000
real-time PCR system from Corbett Research (Sydney, Australia).
Primers of lncRNAs as the following: linc00941 sense:
5′-GCGGTAGCCTTCTCTGAACTG-3′, antisense: 5′-GTTGCATAACCTGACCTGCC-3′;
AF086191 sense: 5′-GCAGAGTGGAGCCTTCTCAT-3′, antisense:
5′-TATGCAAACTCCCATGTGGC-3′. Quantification of miR-500a-5p,
miR-628-3p, miR-128-3p, miR-223-3p and miR-30a was performed with a
stem-loop real-time PCR miRNA kit bought from Guangzhou RiboBio
Co., Ltd. (Guangzhou, China).
Western blot analysis
Protein concentration was quantified using a
bicinchoninic acid protein assay kit and boiled with the sample
buffer in a water bath for 5 min. Protein samples were separated
with 15% SDS-PAGE gels for 2 h and transferred onto a
polyvinylidene difluoride membrane, which was subsequently blocked
in 5% non-fat milk for 2 h. Blots were probed using the primary
antibodies. The anti-vimentin, a-SMA, E-cadherin and Snail antibody
was from ProteinTech Group. After three times washing with tris
buffered saline tween, the horseradish peroxidase-conjugated
secondary antibodies were added. Antigen-antibody complexes were
visualized by enhanced chemiluminescence.
Transmission electron microscopy (TEM)
observation
Cells were fixed by treatment with fresh 2.5%
glutaraldehyde at 4°C for at least 4 h, post-fixed in 1% osmium
tetroxide for 1.5 h, dehydrated in gradient ethanol, infiltrated
with Epon812, embedded and cultured at 37°C for 12 h, 45°C for 12 h
and 60°C for 24 h. Ultrathin sections prepared with an ultracut E
ultramicrotome were stained with uranyl acetate and lead citrate
and observed using a JEM-1400 TEM (JEOL Ltd., Tokyo, Japan).
Statistical analysis
All data are expressed as the means ± SD.
Statistical analysis was performed using SPSS13.0 software by
one-way ANOVA. P<0.05 was considered to indicate a statistically
significant difference.
Results
Identification of the cell model
EMT is the phenomenon in which epithelial cells
obtain mesenchymal characteristics, including change in shape,
expression of mesenchymal markers, such as collagen and a smooth
muscle actin (a-SMA); loss of epithelial cell marker E-cadherin;
and increase of transcription repressor Snail. Under an optical
microscope, the shapes of the cells changed from irregular to
spindle. In the TEM images, the number of muscle fibers in the
cytoplasm of TGF-β1-activated cells are larger than those of normal
control cells (Fig. 1A).
Meanwhile, the expression levels of collagen I, a-SMA, E-cadherin,
and Snail were significantly increased, whereas that of E-cadherin
expression significantly decreased (Fig. 1B-F). These findings indicated the
successful establishment of the cell model.
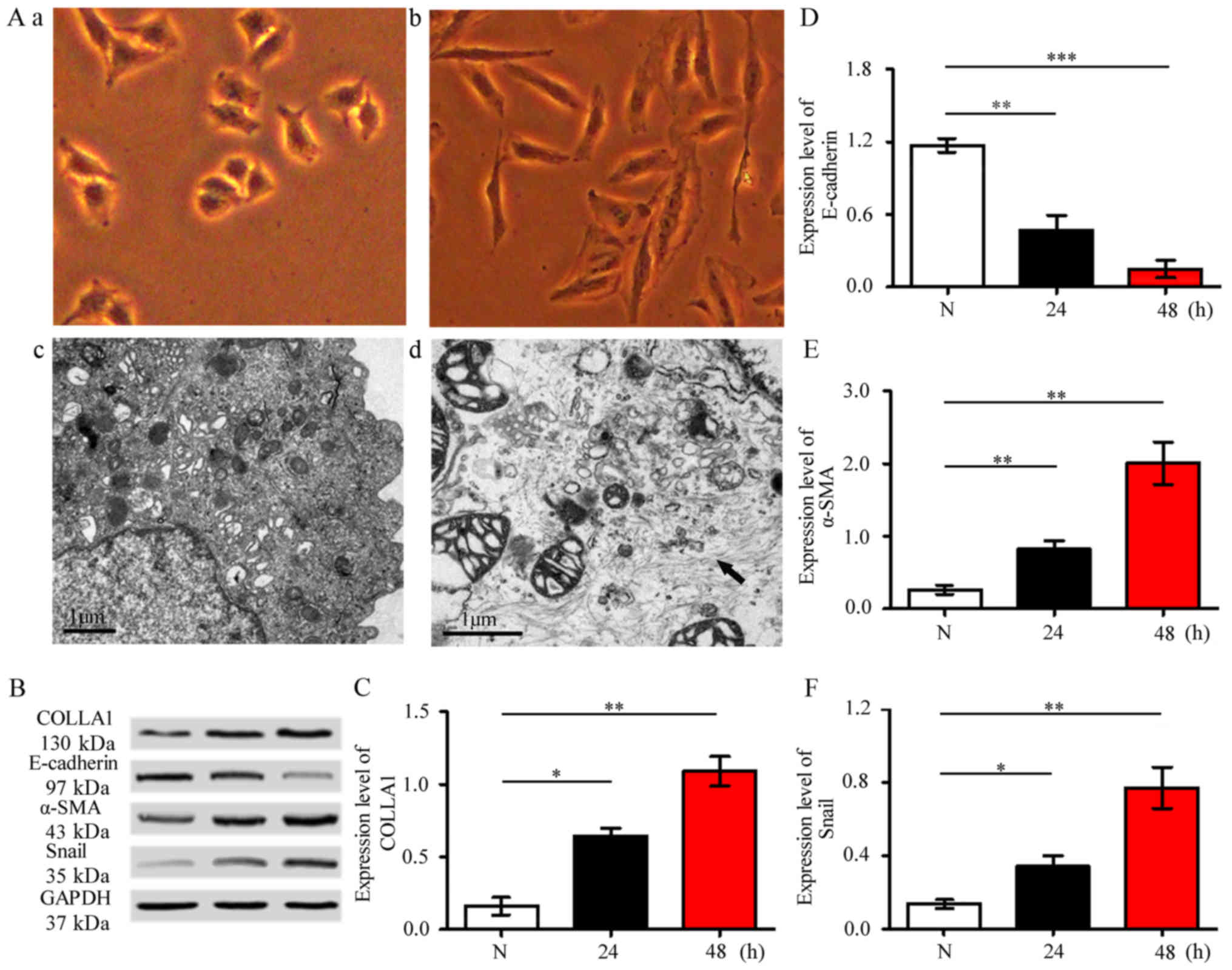 | Figure 1.Identification of cell model. (A)
Optical microscopy observed A549 cell shape changed from irregular
to spindle (a and b), ×400 magnification. TEM image showed more
muscle fibers in the cytoplasm of TGF-β1-activated cells compared
with those in normal controls (c and d). (B-F) Expression levels of
collagen, a-SMA, E-cadherin, and Snail were determined using
western blot analysis in A549 cell. The expression levels of
collagen I, a-SMA, and Snail significantly increased, whereas that
of E-cadherin significantly decreased. Data are expressed as means
± SD, *P<0.05, **P<0.01, ***P<0.001 vs. control. TEM,
transmission electron microscopy; TGF-β1, tumor growth factor-β1;
SMA, smooth muscle actin. |
Differentially coexpressed mRNAs,
miRNAs, and lncRNAs
The expressed profiles were determined by using
Affymetrix Human Transcriptome Array 2.0 microarray for mRNAs and
lncRNAs and Affymetrix Gene Chip miRNA4.0 microarray for miRNAs in
A549 and TGF-β1-activated A549 at 12, 24, and 48 h, respectively.
Hierarchical clustering analysis results showed that the mRNAs,
miRNAs, and lncRNAs of the cell models are differentially expressed
(Fig. 2A-C). Basing on our
previous studies and other published works, we analyzed the data of
TGF-β1-activated A549 only at 24 and 48 h. A total of 242 mRNAs
were differentially coexpressed in TGF-β1-activated 24 and 48 h
cell samples compared with the normal control group. Of these
mRNAs, 139 were upregulated, 103 were downregulated, and 143 miRNAs
were differentially coexpressed in TGF-β1-activated 24 and 48 h
cell samples compared with the normal control group. Among these
miRNAs, 83 were upregulated, 60 were downregulated, and 127 lncRNAs
were differentially coexpressed in TGF-β1-activated 24 and 48 h
cell samples compared with the normal control group. Among which,
71 were upregulated and 56 were downregulated.
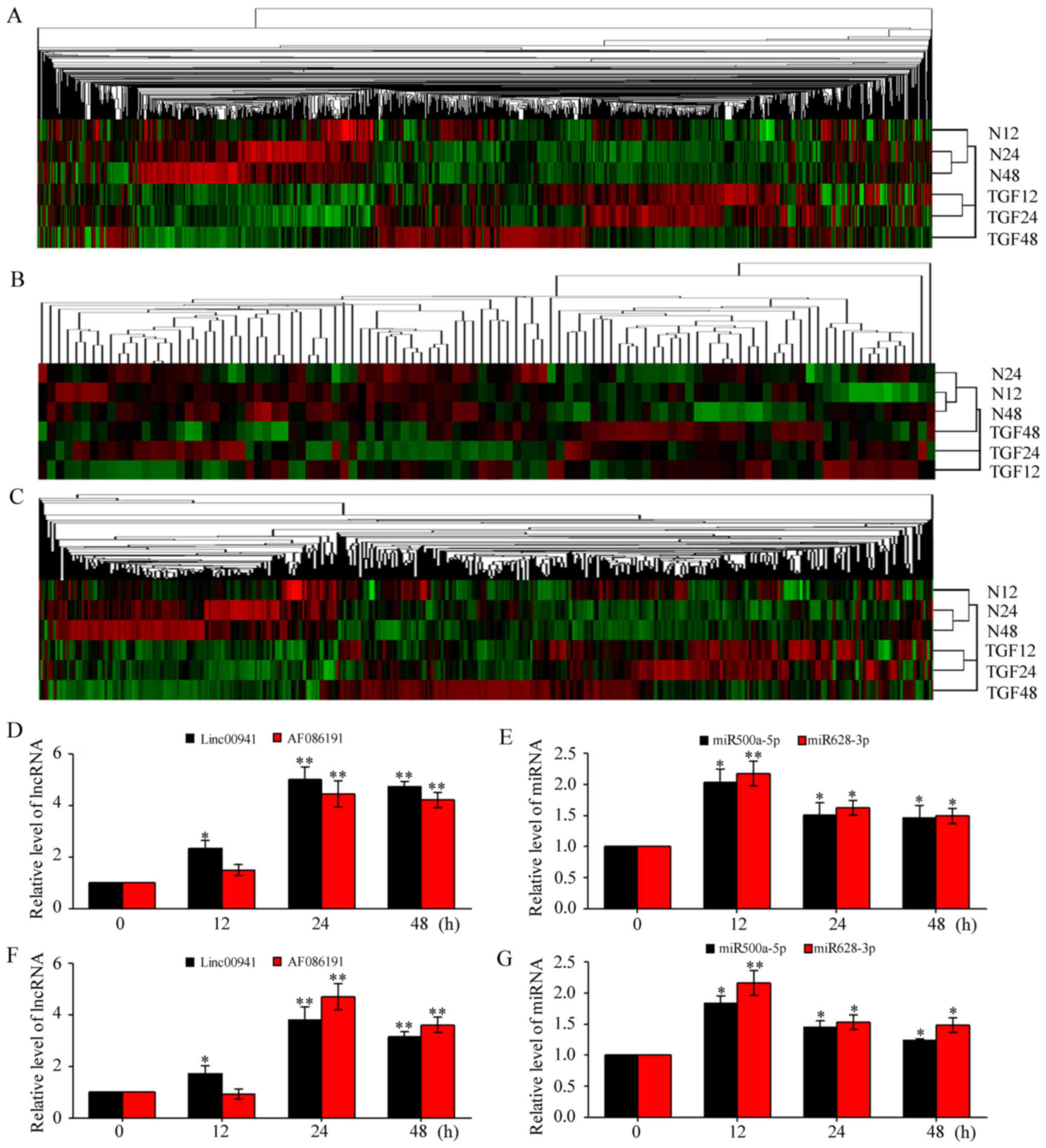 | Figure 2.Hierarchical clustering and
validation of microarray data. (A-C) Hierarchical clustering of
mRNAs (A), miRNAs (B), and lncRNAs (C) in TGF-β1-activated
pulmonary epithelial cells and normal controls at 12, 24, and 48 h.
TGF12-48: TGF-β1-activated 12, 24, and 48 h A549 cell samples.
N12-48: Normal A549 cell controls. The red and green shades
indicate the expression above and below the relative expression,
respectively, across all samples. (D-G) Validation of microarray
data by qRT-PCR analysis. Two lncRNAs (D and F) and two miRNAs (E
and G) were validated in A549 and MRC-5 cell, respectively. U6
served as an internal control for miRNA, and glyceraldehyde
3-phosphate dehydrogenase for lncRNA. The relative expression level
of each miRNA or lncRNA was a normalized one, and data displayed in
histograms are expressed as means ± SD, *P<0.05, **P<0.01 vs.
control. miRNAs, microRNAs; lncRNAs, long noncoding RNAs; TGF,
tumor growth factor. |
To validate the microarray data, we randomly
selected two miRNAs (miR-500a-5p and miR-628-3p) and two lncRNAs
(linc00941 and AF086191) and determined their expression levels
through qRT-PCR. These RNAs were selected because they have
relatively higher differences than other RNAs. As shown in Fig. 2, the two miRNAs and two lncRNAs
were differentially expressed in A549 cell models compared with
those in controls (Fig. 2D and E).
To further validate this result, we tested these ncRNAs on MRC-5
cells derived from human embryonic lung fibroblast cells (Fig. 2F and G). All results are in
agreement with those in A549 cell models (12). As expected, we also identified
previously discovered fibrotic miR-30a (28,29),
thus validating our microarray data.
Analysis of coexpressed mRNAs
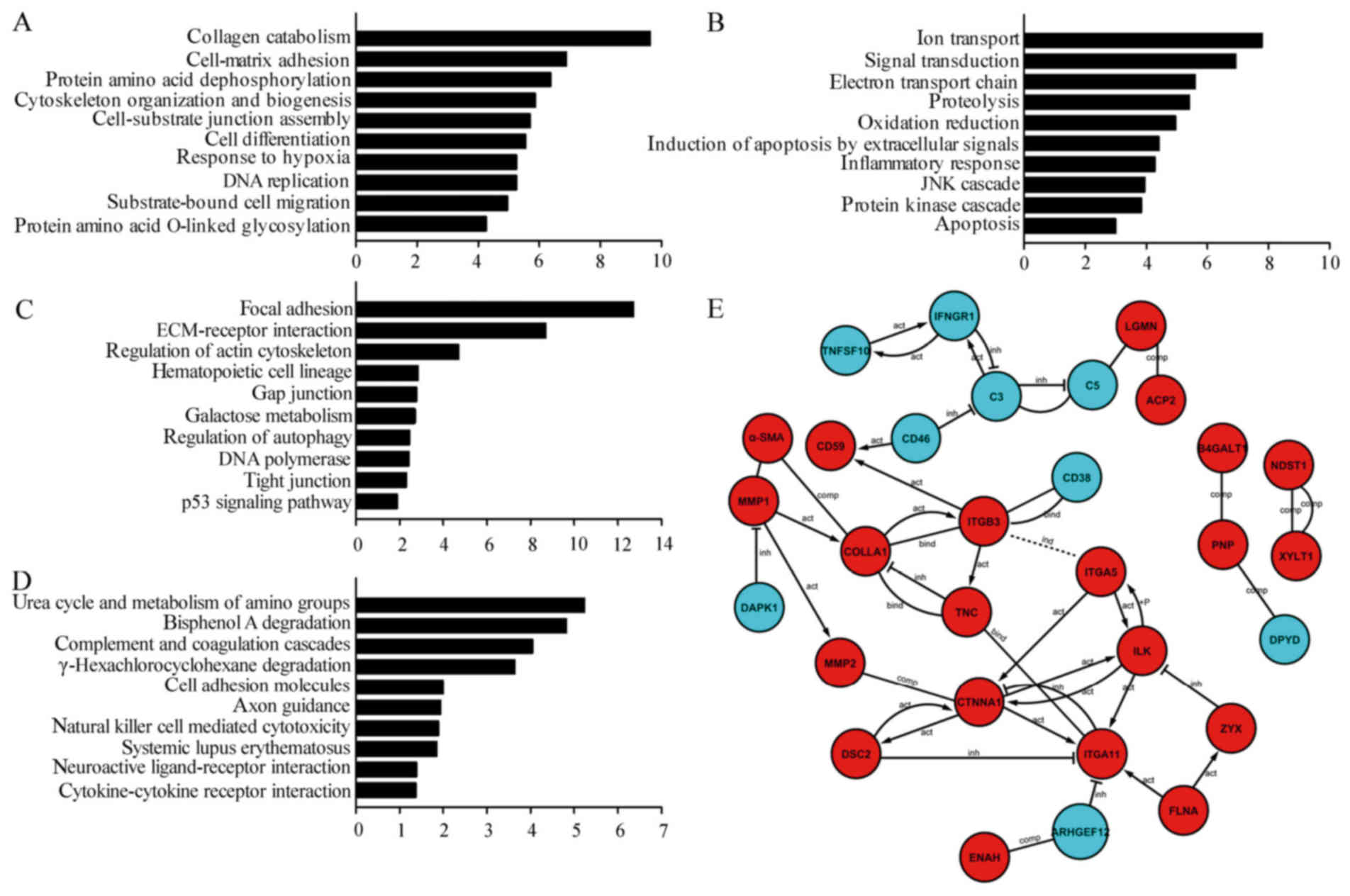
The functional classification of 242 coexpressed
mRNAs in TGF-β1-activated 24 and 48 h cell samples was analyzed
using GO analysis. According to the GO annotation tool,
differentially upregulated coexpressed mRNAs were principally
enriched for GO terms related to collagen catabolism, cell-matrix
adhesion, protein amino acid dephosphorylation, cytoskeleton
organization and biogenesis, cell-substrate junction assembly, cell
differentiation, response to hypoxia, DNA replication,
substrate-bound cell migration, and protein amino acid O-linked
glycosylation (Fig. 3A). At the
same time, we compared the downregulated coexpression mRNAs in
TGF-β1-activated 24 and 48 h cell samples with that in the normal
control group, which were principally enriched for GO terms related
to ion transport, signal transduction, electron transport chain,
proteolysis, oxidation reduction, induction of apoptosis by
extracellular signals, inflammatory response, Jun-N-terminal kinase
cascade, protein kinase cascade, and apoptosis (Fig. 3B). The TGF-β1 induction in
EMT-related genes in pulmonary epithelial cells contributes to the
upregulation or downregulation of the coexpression of these genes
by endowing cells with a specific adhesive, motile, and invasive
potential.
Biological pathways associated with coexpressed
mRNAs in TGF-β1-activated 24 and 48 h cell samples were analyzed
using the KEGG database (Fig. 3C and
D). The 10 upregulated and 10 downregulated pathways listed
were identified and associated with focal adhesion, extracellular
cell matrix (ECM)-receptor nitration, regulation of actin
cytoskeleton, hematopoietic cell lineage, gap junction, galactose
metabolism, autophagy regulation, p53 signaling pathway, DNA
polymerase, tight junction, and cell adhesion molecules.
Furthermore, we further analyzed the interaction
network of the 242 coexpressed mRNAs on the basis of
protein-protein interaction. In total, 21 upregulated mRNAs and
nine downregulated mRNAs were included in the interaction network
(Fig. 3E), which showed four types
of interaction, namely, activated, inhibited, complementary, and
binding interaction, among the coexpressed 30 mRNAs. CTNNA, ITGA11,
and ITGB3 are the network nodes or connections in the interaction
network. In addition, MMP1, MMP2, and COLLA1, which are the
physiological indicators of fibrogenesis during EMT progression,
were also highly and positively expressed.
Coexpressed mRNA-miRNA-lncRNA
interaction network
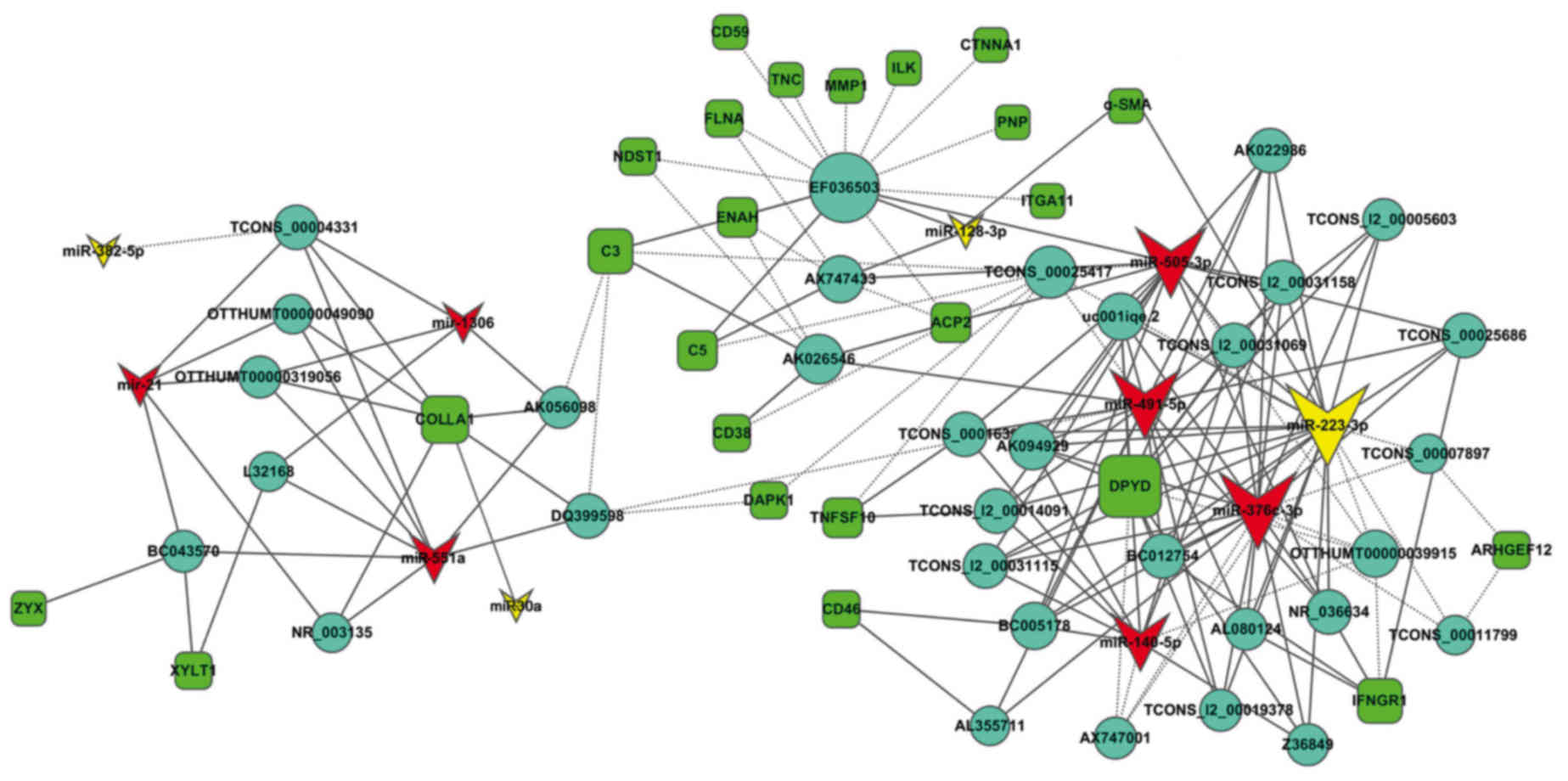
The 30 coexpressed mRNAs were further analyzed.
Among these mRNAs, 24 mRNAs, which can coexpress and interact with
11 miRNAs and 33 lncRNAs, exist. The coexpressed interaction
network indicated that one mRNA might interact with several miRNAs
or lncRNAs. Similarly one miRNA or lncRNA can interact with several
mRNAs or lncRNAs (Fig. 4). The
degree of mRNA-miRNA-lncRNA coexpression were analyzed. An
increased degree indicated an increased coexpression in the
mRNA-miRNA-lncRNA interaction network. The coexpressed degree of
the mRNAs, miRNAs, and lncRNAs in the network nodes and connections
were higher than that of the others (Fig. 5A-C). We further analyzed the lncRNA
category. The data showed that most of differentially coexpressed
lncRNAs are transcribed from the intergenic gene. Some are from
sense-overlapping gene. Meanwhile, a few other unknown sources of
lncRNAs exist (Fig. 5D).
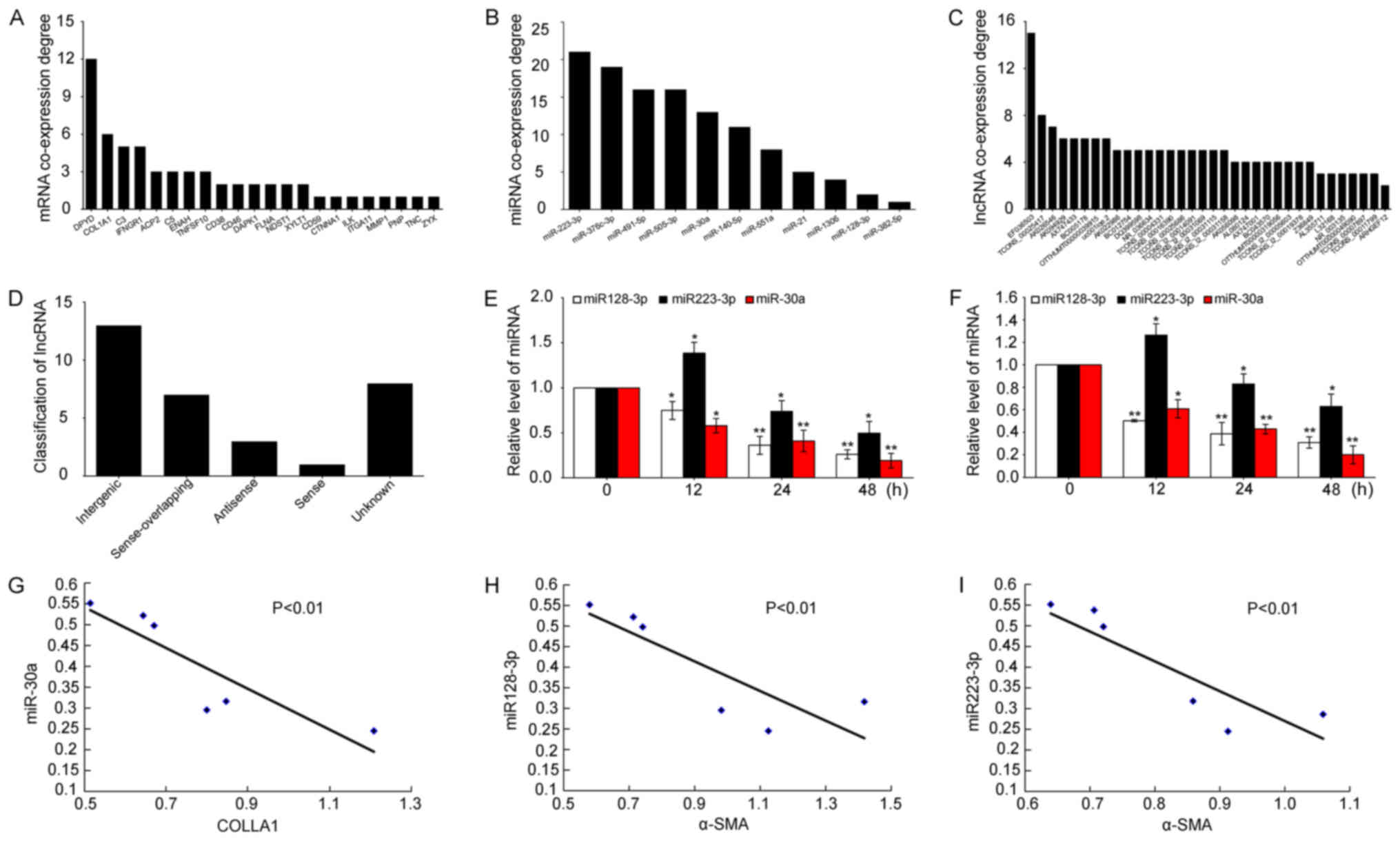 | Figure 5.Analysis of the mRNA-miRNA-lncRNA
interaction network. (A-C) Coexpression degree of mRNA, miRNA, and
lncRNA in TGF-β1-activated A549 cell samples at 24 and 48 h. (D)
Bar diagram shows the 33 lncRNA category. Most differentially
expressed lncRNAs originated from intergenic genes. Some of the
lncRNAs were derived from sense overlapping and few from other
unknown sources. (E) Expression of miR-30a, miR-128-3p, and
miR-223-3p decreased in TGF-β1-activated A549 cell samples at 12,
24, and 48 h, respectively. U6 served as an internal control for
miRNA. (F) Expression of miR-30a, miR-128-3p, and miR-223-3p
decreased in TGF-β1-activated MRC-5 cell samples at 12, 24, and 48
h, respectively. U6 served as an internal control for miRNA. (G-I)
Pearson correlation coefficient showed that miR-30a, miR-128-3p and
miR-223-3p are inversely correlated with collagen I and α-SMA,
respectively. Data are expressed as mean ± SD, *P<0.05,
**P<0.01 vs. control. miRNA, microRNA; lncRNA, long noncoding
RNA; TGF-β1, tumor growth factor-β1. |
In the interaction network, miR-223-3p displays the
highest coexpression degree compared with other miRNAs. miR-30a has
relatively high coexpression degree, whereas miR-128-3p has
relatively low coexpression degree (Fig. 5A). Thus, we selected the three
miRNAs and determined their expression levels by using qRT-PCR to
illuminate the interaction network. As shown in Fig. 5E, the three miRNAs were
differentially decreased in the A549 cell models compared with
those in controls. To further validate the data, we tested these
miRNA expression levels on MRC-5 cells. The expression trend of
these miRNAs is similar pattern to those of miRNAs in A549 cell
models (Fig. 5F). Then, analysis
of the Pearson's correlation coefficient demonstrated that miR-30a
is inversely correlated with collagen I, and miR-128-3p and
miR-223-3p are inversely correlated with α-SMA (Fig. 5G-I).
Some researchers named the interaction among mRNA,
miRNA, and lncRNA functions ceRNA mechanism. A detailed study
should be carried out for determining whether the ceRNA mechanism
is involved in the interactions among coexpressed mRNAs, miRNAs,
and lncRNAs.
Interaction network of coexpressed
miRNA and targeted genes
The ceRNA mechanism proposed that all types of RNA
transcripts communicate through a new ‘language’ mediated by
miRNA-binding sites. The miRNAs bind to sequences with partial
complementarity on target RNA transcripts resulting in the
repression of target gene expression (17). Thus, we further analyzed the miRNAs
and their targeted genes in the interaction network of mRNAs,
miRNAs, and lncRNAs. First, the target genes of coexpressed miRNA
were predicted based on TargetScan, miRanda data, and miRbase.
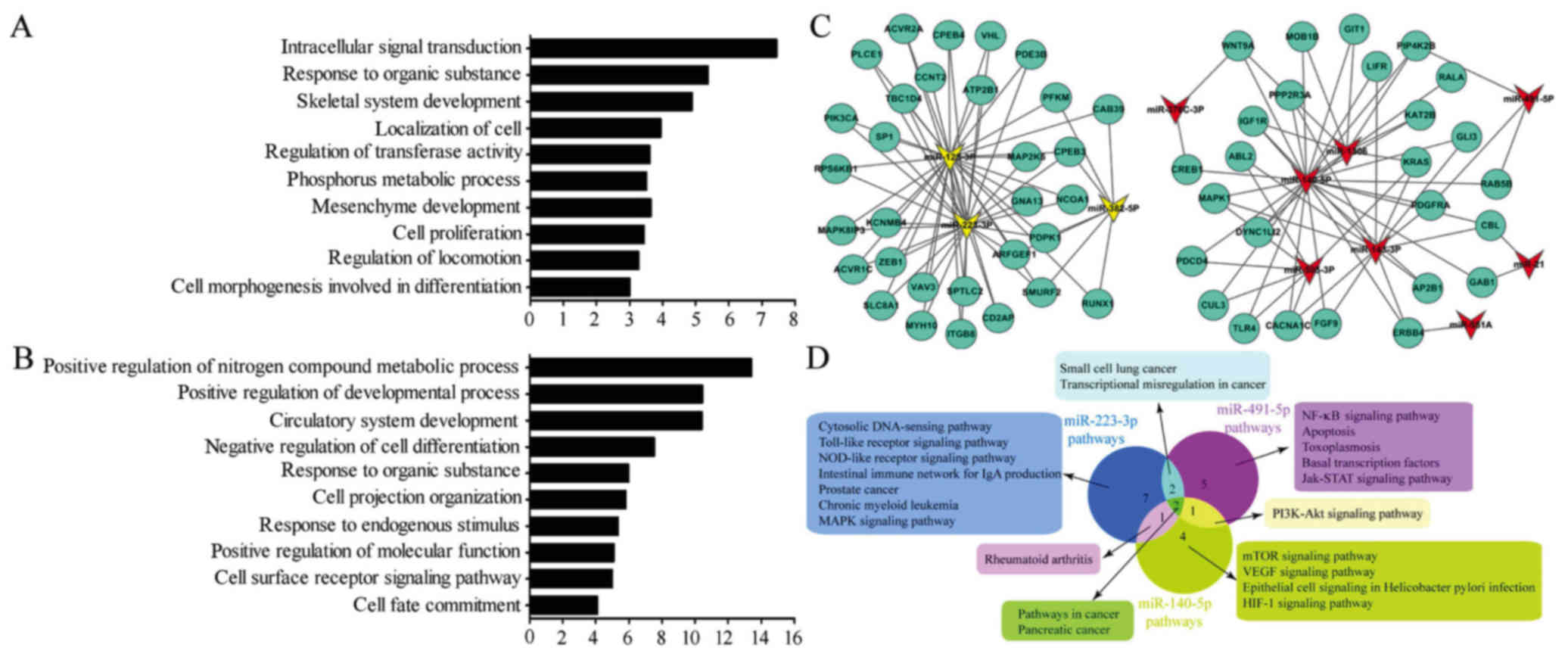
Subsequently, we analyzed the functional classification of these
target genes by GO analysis. The results showed that the
upregulated target genes were principally enriched for GO terms
related to organic substance response, skeletal system development,
transferase activity regulation, mesenchyme development, cell
proliferation, and cell morphogenesis (Fig. 6A). The downregulated coexpressed
miRNA target genes were principally enriched for GO terms related
to the positive regulation of development, negative regulation of
cell differentiation, positive regulation of molecular function,
and cell fate commitment (Fig.
6B). Most of these upregulated or downregulated miRNA target
genes are associated with EMT.
In the interaction network of coexpressed miRNAs and
their targeted genes, miR-223-3p, miR-376C-3p, miR-491-5p,
miR-505-3p, miR-140-5p, miR-551A, miR-21, miR-1306, miR-128-3p, and
miR-382-5p from the mRNA-miRNA-lncRNA interaction network were
considered as examples (Fig. 6C).
One miRNA had many target genes, whereas different miRNAs can have
the same target gene. One gene also can have different binding
sites with miRNAs. In total, 57 mRNAs might be the 10 miRNAs'
targeted genes. However, among these targeted genes, not a single
one is the same as the mRNA in mRNA-miRNA-lncRNA interaction
network. This finding means that the ceRNA mechanism does not exist
in the above coexpressed mRNA-miRNA-lncRNA interaction network.
Analysis with DIANA-miRPath determined 10 candidate miRNAs, which
were involved in possible disease-related pathways (P-value cutoff
at 0.05). miR-223-3p, miR-491-5p, and miR-140-5p were significantly
implicated in disease-related pathways and were found to be
associated in 12, 10, and eight pathways, respectively. The Venn
diagram revealed the intersection (Fig. 6D). Thus, cancer and pancreatic
cancer pathways are crossing pathways related to miR-223-3p,
miR-491-5p, and miR-140-5p.
In addition, our previous study demonstrated that
miR-30a may function as a novel therapeutic target for lung
fibrosis by blocking mitochondrial fission dependent on
dynamin-related protein1 (Drp-1). miR-30a exerts its regulatory
function through regulating Drp-1 promoter hydroxymethylation by
directly targeting TET1. To date, our further detailed study has
showed no existence of ceRNA mechanism in miR-30a regulatory mode
(28,29).
Discussion
A recent study found that pulmonary fibrosis
exhibited several cancer-like pathogenic features. Thus, pulmonary
fibrosis is considered to be a neoproliferative disorder of the
lung (31). Similar to cancer
cells, myofibroblasts show epigenetic and genetic abnormalities, as
well as functional features, such as uncontrolled proliferation,
resistance to apoptosis, and high migration rates (32). Recently, noncoding RNAs, such as
miRNA and lncRNA, have been found to be involved in the development
of diseases, especially tumor and pulmonary fibrosis (33–36).
However, the mechanism underlying the coexpression and interaction
among mRNA, miRNA, and lncRNA in disease regulation requires
elucidation. In the present study, we applied microarray and
experimental data to examine mRNA, miRNA, and lncRNA coexpression
profiles in human alveolar epithelial cells and interpret their
interactions. The results indicated that the abnormally expressed
mRNA, miRNA, and lncRNA function in the development of pulmonary
tumor or fibrosis.
In our previous studies on the EMT process, 10 ng/ml
of TGF-β1-activated 24 and 48 h cell samples were selected
(37,38). The coexpression profiles of mRNA,
miRNA, and lncRNA in TGF-β1-activated 24 and 48 h cell samples were
first investigated in this study. Some studies already reported
coexpressed mRNAs or miRNAs, such as COL1 (39), C3 (40), MMP1 (41), miR-30a (42), and miR-21 (43), that can regulate EMT induced by
TGF-β. However, these studies usually only focus on targeting a
single gene or protein in a single molecular pathway rather than on
systematic or coordinated expression of other genes during disease
development. Our data showed that the future challenge for an
effective pathway-based therapy is how to disrupt numerous altered
disease pathways. One specific mRNA, miRNA, or lncRNA may target
multiple genes potentially implicated in diseases.
Detailed information on the biological functions and
potential mechanisms of 242 mRNAs in EMT based on the results of GO
and KEGG pathways were obtained. Then, 29 mRNAs, which were
coexpressed and interacted with one another, were ranked. Many of
these genes are well known because they often occur in the majority
of fibrotic diseases and in various tumors (44,45).
These genes can be categorized according to their functions
recorded in the KEGG database during EMT. MMP1,
MMP2, and COL1A1 were associated
with ECM-receptor interaction pathway; ITGA5,
ITGA11, ITGB3, and CTNNA1, with
focal adhesion pathway; and CD38, CD46,
CD59, C3, and C5, with
hematopoietic cell lineage pathway. Thus, the hematopoietic cell
plays an important role in human activities, and drawing blood
becomes simpler and easier than obtaining tissue samples. As a
result, the biomarkers from peripheral blood have attracted
considerable attention from researchers investigating diseases,
such as tumor and fibrosis (46–48).
Our laboratory group reported that the apoptosis of alveolar
epithelial cells is suppressed by high miR-30a expression, which
can be detected in the peripheral blood of patients with idiopathic
pulmonary fibrosis (28,29). Biomarkers or key genes from
peripheral blood ideally providing predictive information for the
diagnosis, progression, treatment responses, and disease prognosis
are thus prospects of future disease research. The high-throughput
analysis of large-scale gene data, such as those presented in this
study can facilitate the identification of maximally upregulated
and downregulated genes, which are possible future targets
(49).
Basing on the 29 coexpression mRNAs, we analyzed
which coexpression miRNAs and lncRNAs can interact. The results
showed that 24 mRNAs, 11 miRNAs, and 33 lncRNAs, can coexpress and
interact with one another. Experimental evidence supports that
lncRNAs act as ceRNAs for miRNAs and play roles in physiological
and pathological processes. Kumar (50) reported that HMGA2 can operate as a
ceRNA for the let-7 family to promote lung cancer progression.
Karreth (51) also reported that
the BRAF pseudogene functions as a ceRNA and induces lymphoma in
vivo. Therefore, we further analyzed whether ceRNA is present
in the mRNA-miRNA-lncRNA interaction network. Data showed that 57
mRNAs might be the targeted genes of 10 miRNAs. However, no mRNA
comes from the mRNA-miRNA-lncRNA interaction network. This finding
suggests that no ceRNA exists in the mRNA-miRNA-lncRNA interaction
network. Living beings have evolved complex mechanisms for
efficient functioning. ceRNA theories can only illustrate the
possibility of regulation in living beings. Moreover, some other
functional modes, such as histone modification and transcription
factor recruitment in the interaction among mRNA, miRNA, and
lncRNA, occur. Wang (52) reported
that lncRNA-DC promotes STAT3 signaling by interacting with the C
terminus of STAT3 to control human dendritic cell differentiation.
However, the possibility of ceRNA existing in pulmonary tumor and
fibrosis cannot be ruled out because of the generally low abundance
of lncRNAs, and the frequent promiscuity of protein-RNA
interactions. Of course, this research is still in the exploratory
stage, and conclusions were obtained only through the changes in
their levels and bioinformatics analysis, needing further
experimental identification and validation.
The mRNA-miRNA-lncRNA interaction network showed
that during the occurrence and development of diseases, many mRNA,
miRNA, or lncRNA molecules are involved. The term interaction means
different things. Proteins encoded by the mRNAs may interact with
each other, with miRNAs, or with lncRNAs. miRNAs may interact and
regulate the stability or expression of mRNAs or lncRNAs. lncRNAs
may interact with miRNAs or mRNAs. Each gene regulates a broad
spectrum of target genes and affects many signaling pathways.
Therefore, disruption of a single gene expression may only limit an
inhibitory effect, whereas joint interference of multiple genes may
be effective. Thus, determining how to silence multiple abnormally
expressed genes simultaneously in order to enhance the efficacy of
disease treatments is important. Li (53) reported an artificially designed
interfering lncRNA, which simultaneously contains sequences that
can complementarily bind to multiple miRNAs.
The genes enlisted in the current study represent
only a small proportion of genes published on pulmonary tumor or
fibrosis. This condition may be correlated with the time point of
analysis or the specific condition of in vitro culture of
the cells. In conclusion, the present study provided new
information regarding the potential role of mRNAs, miRNAs, and
lncRNAs in EMT. These findings may provide basic mechanistic
information, possible biomarkers, and novel treatment of
EMT-targeted strategies for diseases, especially for pulmonary
tumor and fibrosis.
Acknowledgements
This study was supported by National Natural Science
Foundation of China (31670365, 31470415, 81670064), Important
Project of Science and Technology of Shandong Province
(2014GSF119014), Project of Science and Technology of Yantai
(2016ZH082). International Cooperation Program for Key Professors
by Shandong Provincial Education Deparment.
References
|
1
|
Fernandez IE and Eickelberg O: New
cellular and molecular mechanisms of lung injury and fibrosis in
idiopathic pulmonary fibrosis. Lancet. 380:680–688. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Alonso SR, Tracey L, Ortiz P, Pérez-Gómez
B, Palacios J, Pollán M, Linares J, Serrano S, Sáez-Castillo AI,
Sánchez L, et al: A high-throughput study in melanoma identifies
epithelial-mesenchymal transition as a major determinant of
metastasis. Cancer Res. 67:3450–3460. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Oruqaj G, Karnati S, Vijayan V, Kotarkonda
LK, Boateng E, Zhang W, Ruppert C, Günther A, Shi W and
Baumgart-Vogt E: Compromised peroxisomes in idiopathic pulmonary
fibrosis, a vicious cycle inducing a higher fibrotic response via
TGF-β signaling. Proc Natl Acad Sci USA. 112:pp. 2048–2057. 2015;
View Article : Google Scholar
|
|
4
|
Akhurst RJ and Hata A: Targeting the TGFβ
signalling pathway in disease. Nat Rev Drug Discov. 11:790–811.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Blackwell TS, Tager AM, Borok Z, Moore BB,
Schwartz DZ, Anstom KJ, Bar-Joseph Z, Bitterman P, Blackburn MR,
Bradford W, et al: Future directions in idiopathic pulmonary
fibrosis research. An NHLBI workshop report. Am J Respir Crit Care
Med. 189:pp. 214–222. 2014; View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iser IC, Pereira MB, Lenz G and Wink MR:
The epithelial-to-mesenchymal transition-like process in
glioblastoma: An updated systematic review and in silico
investigation. Med Res Rev. 37:271–313. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shenoy A and Blelloch RH: Regulation of
microRNA function in somatic stem cell proliferation and
differentiation. Nat Rev Mol Cell Biol. 15:565–576. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Holdt LM, Hoffmann S, Sass K, Langenberger
D, Scholz M, Krohn K, Finstermeier K, Stahringer A, Wilfert W,
Beutner F, et al: Alu elements in ANRIL non-coding RNA at
chromosome 9p21 modulate atherogenic cell functions through
trans-regulation of gene networks. PLoS Genet. 9:e10035882013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Payer B, Rosenberg M, Yamaji M, Yabuta Y,
Koyanagi-Aoi M, Hayashi K, Yamanaka S, Saitou M and Lee JT: Tsix
RNA and the germline factor, PRDM14, link X reactivation and stem
cell reprogramming. Mol Cell. 52:805–818. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Song R, Walentek P, Sponer N, Klimke A,
Lee JS, Dixon G, Harland R, Wan Y, Lishko P, Lize M, et al:
miR-34/449 miRNAs are required for motile ciliogenesis by
repressing cp110. Nature. 510:115–120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nicodemus-Johnson J, Laxman B, Stern RK,
Sudi J, Tierney CN, Norwick L, Hogarth DK, McConville JF, Naureckas
ET, Sperling AI, et al: Maternal asthma and microRNA regulation of
soluble HLA-G in the airway. J Allergy Clin Immunol. 131:1496–1503.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Caruso P, Dempsie Y, Stevens HC, McDonald
RA, Long L, Lu R, White K, Mair KM, McClure JD, Southwood M, et al:
A role for miR-145 in pulmonary arterial hypertension: Evidence
from mouse models and patient samples. Circ Res. 111:290–300. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sayed D and Abdellatif M: MicroRNAs in
development and disease. Physiol Rev. 91:827–887. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun Q, Csorba T, Skourti-Stathaki K,
Proudfoot NJ and Dean C: R-loop stabilization represses antisense
transcription at the Arabidopsis FLC locus. Science. 340:619–621.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tollervey JR, Curk T, Rogelj B, Briese M,
Cereda M, Kayikci M, König J, Hortobágyi T, Nishimura AL, Zupunski
V, et al: Characterizing the RNA targets and position-dependent
splicing regulation by TDP-43. Nat Neurosci. 14:452–458. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gontan C, Achame EM, Demmers J, Barakat
TS, Rentmeester E, van IJcken W, Grootegoed JA and Gribnau J: RNF12
initiates X-chromosome inactivation by targeting REX1 for
degradation. Nature. 485:386–390. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kino T, Hurt DE, Ichijo T, Nader N and
Chrousos GP: Noncoding RNA gas5 is a growth arrest- and
starvation-associated repressor of the glucocorticoid receptor. Sci
Signal. 3:ra82010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu X, Li D, Zhang W, Guo M and Zhan Q:
Long non-coding RNA gadd7 interacts with TDP-43 and regulates Cdk6
mRNA decay. EMBO J. 31:4415–4427. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hung T, Wang Y, Lin MF, Koegel AK, Kotake
Y, Grant GD, Horlings HM, Shah N, Umbricht C, Wang P, et al:
Extensive and coordinated transcription of noncoding RNAs within
cell-cycle promoters. Nat Genet. 43:621–629. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee S, Kopp F, Chang TC, Sataluri A, Chen
B, Sivakumar S, Yu H, Xie Y and Mendell JT: Noncoding RNA NORAD
regulates genomic stability by sequestering PUMILIO proteins. Cell.
164:69–80. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cao G, Zhang J, Wang M, Song X, Liu W, Mao
C and Lv C: Differential expression of long non-coding RNAs in
bleomycin-induced lung fibrosis. Int J Mol Med. 32:355–364.
2013.PubMed/NCBI
|
|
23
|
Song X, Cao G, Jing L, Lin S, Wang X,
Zhang J, Wang M, Liu W and Lv C: Analysing the relationship between
lncRNA and protein-coding gene and the role of lncRNA as ceRNA in
pulmonary fibrosis. J Cell Mol Med. 18:991–1003. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Foster KA, Oster CG, Mayer MM, Avery ML
and Audus KL: Characterization of the A549 cell line as a type II
pulmonary epithelial cell model for drug metabolism. Exp Cell Res.
243:359–366. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vockley CM, D'Ippolito AM, McDowell IC,
Majoros WH, Safi A, Song L, Crawford GE and Reddy TE: Direct GR
binding sites potentiate clusters of TF binding across the human
genome. Cell. 166:1269–1281. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pardo A, Gibson K, Cisneros J, Richards
TJ, Yang Y, Becerril C, Yousem S, Herrera I, Ruiz V, Selman M and
Kaminski N: Up-regulation and profibrotic role of osteopontin in
human idiopathic pulmonary fibrosis. PLoS Med. 2:e2512005.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Roberts JR, Perkins GD, Fujisawa T,
Pettigrew KA, Gao F, Ahmed A and Thickett DR: Vascular endothelial
growth factor promotes physical wound repair and is anti-apoptotic
in primary distal lung epithelial and A549 cells. Crit Care Med.
35:2164–2170. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mao C, Zhang J, Lin S, Jing L, Xiang J,
Wang M, Wang B, Xu P, Liu W, Song X and Lv C: miR-30a inhibits
AECs-II apoptosis by blocking mitochondrial fission dependent on
Drp-1. J Cell Mol Med. 18:2404–2416. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang S, Liu H, Liu Y, Zhang J, Li H, Liu
W, Cao G, Xv P, Zhang J, Lv C and Song X: miR-30a as potential
therapeutics by targeting TET1 through regulation of Drp-1 promoter
hydroxymethylation in idiopathic pulmonary fibrosis. Int J Mol Sci.
18:pii: E6332017. View Article : Google Scholar
|
|
30
|
Zhang YL: The value of disease. Guangxi
Normal University Press; Guilin: 2014
|
|
31
|
Vancheri C, Failla M, Crimi N and Raghu G:
Idiopathic pulmonary fibrosis: A disease with similarities and
links to cancer biology. Eur Respir J. 35:496–504. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vancheri C: Idiopathic pulmonary fibrosis:
An altered fibroblast proliferation linked to cancer biology. Proc
Am Thorac Soc. 9:pp. 153–157. 2012; View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Huang C, Yang Y and Liu L: Interaction of
long noncoding RNAs and microRNAs in the pathogenesis of idiopathic
pulmonary fibrosis. Physiol Genomics. 47:463–469. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sun H, Chen J, Qian W, Kang J, Wang J,
Jiang L, Qiao L, Chen W and Zhang J: Integrated long non-coding RNA
analyses identify novel regulators of epithelial-mesenchymal
transition in the mouse model of pulmonary fibrosis. J Cell Mol
Med. 20:1234–1246. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yue B, Cai D, Liu C, Fang C and Yan D:
Linc00152 functions as a competing endogenous RNA to confer
oxaliplatin resistance and holds prognostic values in colon cancer.
Mol Ther. 24:2064–2077. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xing Z, Lin A, Li C, Liang K, Wang S, Liu
Y, Park P, Qin L, Wei Y, Hawke D, et al: lncRNA directs cooperative
epigenetic regulation downstream of chemokine signals. Cell.
159:1110–1125. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang J, Xv P, Wang Y, Wang M, Li H, Lin
S, Mao C, Wang B, Song X and Lv C: Astaxanthin prevents pulmonary
fibrosis by promoting myofibroblast apoptosis dependent on
Drp1-mediated mitochondrial fission. J Cell Mol Med. 19:2215–2231.
2015.PubMed/NCBI
|
|
38
|
Wang M, Zhang J, Song X, Liu W, Zhang L,
Wang X and Lv C: Astaxanthin ameliorates lung fibrosis both in vivo
and in vitro through preventing transdifferentiation, inhibiting
proliferation, and promoting apoptosis of activated cells. Food
Chem Toxicol. 56:450–458. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Alipio ZA, Jones N, Liao W, Yang J,
Kulkarni S, Kumar K Sree, Hauer-Jensen M, Ward DC, Ma Y and Fink
LM: Epithelial to mesenchymal transition (EMT) induced by bleomycin
or TFG(b1)/EGF in murine induced pluripotent stem cell-derived
alveolar Type II-like cells. Differentiation. 82:89–98. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cho MS, Rupaimoole R, Choi HJ, Noh K, Chen
J, Hu Q, Sood AK and Afshar-Kharghan V: Complement component 3 is
regulated by TWIST1 and mediates epithelial-mesenchymal transition.
J Immunol. 196:1412–1418. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Uttamsingh S, Bao X, Nguyen KT, Bhanot M,
Gong J, Chan JL, Liu F, Chu TT and Wang LH: Synergistic effect
between EGF and TGF-beta1 in inducing oncogenic properties of
intestinal epithelial cells. Oncogene. 27:2626–2634. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kao CJ, Martiniez A, Shi XB, Yang J, Evans
CP, Dobi A, White RW deVere and Kung HJ: miR-30 as a tumor
suppressor connects EGF/Src signal to ERG and EMT. Oncogene.
33:2495–2503. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Braun J, Hoang-Vu C, Dralle H and
Hüttelmaier S: Downregulation of microRNAs directs the EMT and
invasive potential of anaplastic thyroid carcinomas. Oncogene.
29:4237–4244. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Selman M, Pardo A, Barrera L, Estrada A,
Watson SR, Wilson K, Aziz N, Kaminski N and Zlotnik A: Gene
expression profiles distinguish idiopathic pulmonary fibrosis from
hypersensitivity pneumonitis. Am J Respir Crit Care Med.
173:188–198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nakerakanti SS, Kapanadze B, Yamasaki M,
Markiewicz M and Trojanowska M: Fli1 and Ets1 have distinct roles
in connective tissue growth factor/CCN2 gene regulation and
induction of the profibrotic gene program. J Biol Chem.
281:25259–25269. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Noé G, Bellesoeur A, Thomas-Schoemann A,
Rangarajan S, Naji F, Puszkiel A, Huillard O, Saidu N, Golmard L,
Alexandre J, et al: Clinical and kinomic analysis identifies
peripheral blood mononuclear cells as a potential pharmacodynamic
biomarker in metastatic renal cell carcinoma patients treated with
sunitinib. Oncotarget. 7:67507–67520. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Fernandez IE, Greiffo FR, Frankenberger M,
Bandres J, Heinzelmann K, Neurohr C, Hatz R, Hartl D, Behr J and
Eickelberg O: Peripheral blood myeloid-derived suppressor cells
reflect disease status in idiopathic pulmonary fibrosis. Eur Respir
J. 48:1171–1183. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Roderburg C and Luedde T: Circulating
microRNAs as markers of liver inflammation, fibrosis and cancer. J
Hepatol. 61:1434–1437. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
du Bois RM: Strategies for treating
idiopathic pulmonary fibrosis. Nat Rev Drug Discov. 9:129–140.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kumar MS, Armenteros-Monterroso E, East P,
Chakravorty P, Matthews N, Winslow MM and Downward J: HMGA2
functions as a competing endogenous RNA to promote lung cancer
progression. Nature. 505:212–217. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Karreth FA, Reschke M, Ruocco A, Ng C,
Chapuy B, Léopold V, Sjoberg M, Keane TM, Verma A, Ala U, et al:
The BRAF pseudogene functions as a competitive endogenous RNA and
induces lymphoma in vivo. Cell. 161:319–332. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang P, Xue Y, Han Y, Lin L, Wu C, Xu S,
Jiang Z, Xu J, Liu Q and Cao X: The STAT3-binding long noncoding
RNA lnc-DC controls human dendritic cell differentiation. Science.
344:310–313. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li X, Su Y, Sun B, Ji W, Peng Z, Xu Y, Wu
M and Su C: An artificially-designed interfering lncRNA expressed
by oncolytic adenovirus competitively consumes oncomiRs to exert
antitumor efficacy in hepatocellular carcinoma. Mol Cancer Ther.
15:1436–1451. 2016. View Article : Google Scholar : PubMed/NCBI
|