Introduction
Osteosarcoma (OS) is a mesenchymal tumor, which is a
primary aggressive type of bone cancer typically presenting in
children and young adults. The causes of OS can be summarized as a
complex of genetic, epigenetic and biological factors. Although
there have been advances in treatment, the clinical outcomes for
patients with OS have not improved substantially and the survival
rate of patients remains at only ~15–30% (1–3).
Therefore, the development of more effective diagnostic and
treatment methods for OS is urgently required.
MicroRNAs (miRNAs) are non-coding, endogenous RNAs,
which show a high level of conservation in the genomes of the
majority of species. The dysregulation of miRNAs has been reported
to be involved in the progression of OS and OS-derived cells
through effects on the malignant phenotype of the cells or
sensitivity to chemotherapy (4,5).
miRNA (miR)-494 has been investigated in several types of tumor,
including non-small cell lung cancer, liver cancer,
cholangiocarcinoma, gastric cancer and pancreatic cancer (6–10).
However, the modulatory effect of miR-494 differs in different
types of tumor. For example, miR-494 can induce resistance of tumor
necrosis factor-related apoptosis-inducing ligand and enhance G1/S
transition leading to promotion of the malignant feather of liver
cancer (11). By contrast, miR-494
exerts tumor-suppressive effects in ovarian cancer and epithelial
ovarian cancer via inhibiting cell growth and promoting apoptosis
(12,13). miR-494 has also been reported to
induce the activation of drug resistance, which is mediated by the
suppression of bone marrow stromal cells in acute myeloid leukemia
cells (14). According to these
findings, the pathophysiological mechanisms underlying the effects
of miR-494 in tumors and other diseases are complex and numerous,
requiring further investigations.
In the present study, deregulated miR-494 was
identified in OS tissues and OS cells. miR-494 was found to
function as a tumor suppressor in the development of OS by
inhibiting proliferation and cell metastasis, and inducing cell
cycle arrest in OS cells. Previous studies have demonstrated that
cyclin-dependent kinase 6 (CDK6) is vital in the G1/S transition,
and the inhibition of CDK6 can lead to cell cycle arrest (15). In accordance, the data obtained in
the present study demonstrated that CDK6 is a potential target of
miR-494. An increase in the level of CDK6 was closely associated
with the malignant phenotype of OS. These findings indicated that
miR-494 exerted a tumor suppressive effect in OS, the function of
which may be mediated by CDK6, providing a potential diagnostic and
therapeutic target for the treatment of OS.
Materials and methods
Human OS tissues and cells
A total of 18 patients (male, 10; female, 8; age ≤18
years, 13; age >18 years, 5; tumor stage I+II, 15; tumor stage
III, 3) diagnosed with OS were recruited from the Second Affiliated
Hospital, Chongqing Medical University (Chongqing, China). These
patients were divided into two groups (metastasis, vs. no
metastasis) according to radiological results. All patients
provided written informed consent. The experimental protocols were
approved by the Ethics Committee of the Second Affiliated Hospital,
Chongqing Medical University. The OS tissues and the corresponding
normal tissues (5 cm from the tumor margin) were obtained from
resection and then immediately snap frozen in liquid nitrogen for
storage at −80°C.
The OS-derived human HOS, Saos2, U2OS and MG-63 cell
lines, and the NHOst normal osteoblast cell line were purchased
from Shanghai Cell Bank, Chinese Academy of Sciences (Shanghai,
China). The cells were maintained in DMEM with 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and incubated in a humidified atmosphere containing 5%
CO2 at 37°C.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from the tissues and cell
lines using an RNeasy Plus Mini kit (Qiagen GmbH, Hilden, Germany)
according to the manufacturer's instructions. Total RNA (2 µg) was
purified and reverse transcribed into cDNA using a PrimeScript RT
Reagent kit (Perfect Real-Time) from Takara Bio, Inc. (Otsu, Japan)
according to the manufacturer's instructions. The qPCR analysis was
performed on an ABI 7500 Fast Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) with a SYBR Premix Ex
Taq™ II kit (Takara Bio, Inc.). The reaction mix contained 2 µl
cDNA, 10 µl 2X Premix Ex Taq (Probe qPCR), 0.4 µl forward/reverse
primers, 0.8 µl Probe, 0.2 µl ROX Reference Dye II and 6.2 µl
ddH2O in a total volume of 20 µl. The sequences of the
primers were as follows: miR-494 forward,
5′-TGACCTGAAACATACACGGGA-3′, and reverse,
5′-TATCGTTGTACTCCACTCCTTGAC-3′; U6 forward,
5′-AAAGACCTGTACGCCAACAC-3′ and reverse,
5′-GTCATACTCCTGCTTGCTGAT-3′. The PCR cycling conditions were as
follows: 95°C for 3 min, followed by 40 cycles of 95°C for 30 sec,
62°C for 30 sec and 72°C for 30 sec. The results were calculated
using the 2−ΔΔCq method (16) and normalized to the expression of
U6.
Bioinformatics
PicTar (http://pictar.mdc-berlin.de/), TargetScan (http://www.targetscan.org) and miRBase (http://www.mirbase.org/) (17–19)
were used to investigate the potential binding sites of miR494.
Luciferase reporter gene assay
The wild-type 3′-untranslated region (3′-UTR) of
CDK6 containing the miR-494 binding site was cloned into the
pGL3-control vector (Ambion; Thermo Fisher Scientific, Inc.) to
construct the CDK6-3′-UTR-wild-type (WT). The CDK6 3′-UTR mutations
containing mutant (mut) sequences were obtained using a QuikChange
Lightning Site-Directed Mutagenesis kit (Agilent Technologies,
Inc., Palo Alto, CA, USA). All plasmids were confirmed by
sequencing. The miR-494 mimics were generated by Shanghai
GenePharma Co., Ltd. (Shanghai, China) and the scramble sequence
was purchased from Ambion; Thermo Fisher Scientific, Inc. Briefly,
the MG-63 (~2–3×106) and U2OS cells
(~2–3×106) were seeded into a 24-well plate 1 day prior
to transfection, and then co-transfected with miR-494 mimics
(sense, 5′UGA AAC AUA CAC GGG AAA CCU C3′ and antisense, 5′GGU UUC
CCG UGU AUG UUU CAU U3′; 100 nM) or scramble (sense, 5′UUC UCC GAA
CGU GUC ACG UUU 3′ and antisense, 5′ACG UAC ACG UUC GGA GAA UU3′;
50 nM), in addition to the CDK6-3′-UTR-WT or CDK6-3′-UTR-mutusing
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
After 48 h, the luciferase activity was determined using a
Dual-Luciferase reporter assay (Promega Corporation, Madison, WI,
USA), which was normalized to the activity of Renilla
luciferase.
Cell proliferation assays
In order to evaluate the effect of the
overexpression of miR-494 on cell proliferation, MTT assay and
colony formation assays were performed. Briefly, 1×103
cells (MG-63 and U2OS cells) were seeded in 96-well plates with
four repeats and then transfected with the miR-494 mimics or
scramble control. The cell viability was assessed 48 h following
transfection using an MTT kit (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). The results were determined at an optical
density of 570 nm on a Tecan Spectra Fluor microplate reader (Tecan
Group, Ltd., Männedorf, Switzerland). For the colony formation
assay, ~200 transfected cells (miR-494 mimics or Scramble) were
seeded in a 6-well plate with four repeats and cultured in DMEM
containing 10% FBS for 2 weeks at 37°C. The medium was replaced
every 3 days. The colonies were fixed with methanol, stained with
0.1% crystal violet (Sigma-Aldrich; Merck KGaA) and subsequently
counted under a light microscope.
Cell invasion and migration assay
In order to investigate the effect of the
overexpression of miR-494 on cell invasion and migration, the
present study examined metastasis using an assay with BD Matrigel
invasion chambers (BD Biosciences, San Jose, CA, USA). A total of
4×104 cells (MG-63 and U2OS cells) transfected with
miR-494 mimics or scramble control were cultured in serum-free DMEM
medium and subsequently placed into the upper chamber, which was
Matrigel-coated for the cell invasion assay or Matrigel-free for
the migration assay. Simultaneously, 10% FBS was added into the
lower chambers to function as a chemoattractant. Following
incubation for 48 h at 37°C, the cells on the upper surface of the
membranes were removed, and the cells on the lower surface were
fixed and stained (0.1% crystal violet). Images of the cells were
captured and the number of cells were counted under a microscope
(IX71; Olympus, Tokyo, Japan) in five randomly selected fields
(magnification, ×200).
Cell cycle analysis
A total of 1×104 MG-63 and U2OS cells
were first transfected with miR-494 mimics or scramble control.
After 48 h, the transfected cells were digested, collected and
washed with PBS. Following washing twice with PBS, the cells were
fixed with 70% ethanol at 4°C overnight. Propidium iodide (PI)
staining solution (50 µg/ml; 1 mg/ml of RNase A, 0.1% Triton X-100
in PBS) was added into the cell suspension. Finally the cells were
examined on a BD FACSCalibur flow cytometer (BD Biosciences). The
experiments were repeated three times.
In vivo growth assay
To examine the role of miR-494 in vivo, 20
female BALB/C-nu/nu mice (4–5 weeks old; weight, ~18–25 g) were
used, which were purchased from the Animal Center of the Cancer
Institute of Chinese Academy of Medical Science (Beijing, China).
The study protocol was approved by the Ethics Committee of the
Second Affiliated Hospital, Chongqing Medical University. The mice
were maintained on a standard 12:12 h light-dark cycle with free
access to food and water at 18–22°C and with 50–60% humidity. The
mice were randomly dived into two groups: the Lv-miR-494 group and
the Lv-control group. A total of 2×106 MG-63 cells
transfected with miR-494 (Lv-miR-494 group) or the corresponding
control (Lv-control) were subcutaneously injected into the flank
region of the female nude mice. The mice were monitored every 3
days. After 5 weeks, the mice were sacrificed and the tumor tissues
were measured on the basis of their width and length: (length ×
width2)/2.
Western blot analysis
Following transfection, the cells were washed three
times with PBS and lysed with radioimmunoprecipitation assay lysis
buffer to obtain the total protein. Protein concentration was
measured using a bicinchoninic acid protein assay kit (Pierce;
Thermo Fisher Scientific, Inc.). Total proteins (50 µg) were
separated by 10% SDS-PAGE and then transferred onto polyvinylidene
fluoride membranes, as previously described (20). The membranes were incubated with
blocking buffer (5% skimmed milk) for 1.5 h at room temperature and
then incubated with the following primary antibodies: anti-CDK6
(1:500, ab79454; Abcam, Cambridge, MA, USA) and anti-GAPDH antibody
(1:500; SAB4300645-100UG; Sigma-Aldrich, Merck KGaA) at 4°C
overnight. Following incubation with the primary antibody, blots
were washed with TBS-0.1% Tween and subsequently incubated with
horseradish peroxidase-conjugated goat anti-rabbit (1:2,000,
ab6721) or rabbit anti-mouse IgG (1:2,000, ab6709) (both from
Abcam) secondary antibodies at 37°C for 2 h. Band intensity was
quantified using Immobilon Western Chemiluminescent HRP Substrate
(EMD Millipore, Billerica, MA, USA). Experiments were performed in
triplicate.
Statistical analysis
Statistical analysis was performed using the SPSS
13.0 software package (SPSS, Inc., Chicago, IL, USA) using one-way
analysis of variance followed by the Student-Newman-Keuls post hoc
test. P≤0.05 was considered to indicate a statistically significant
difference.
Results
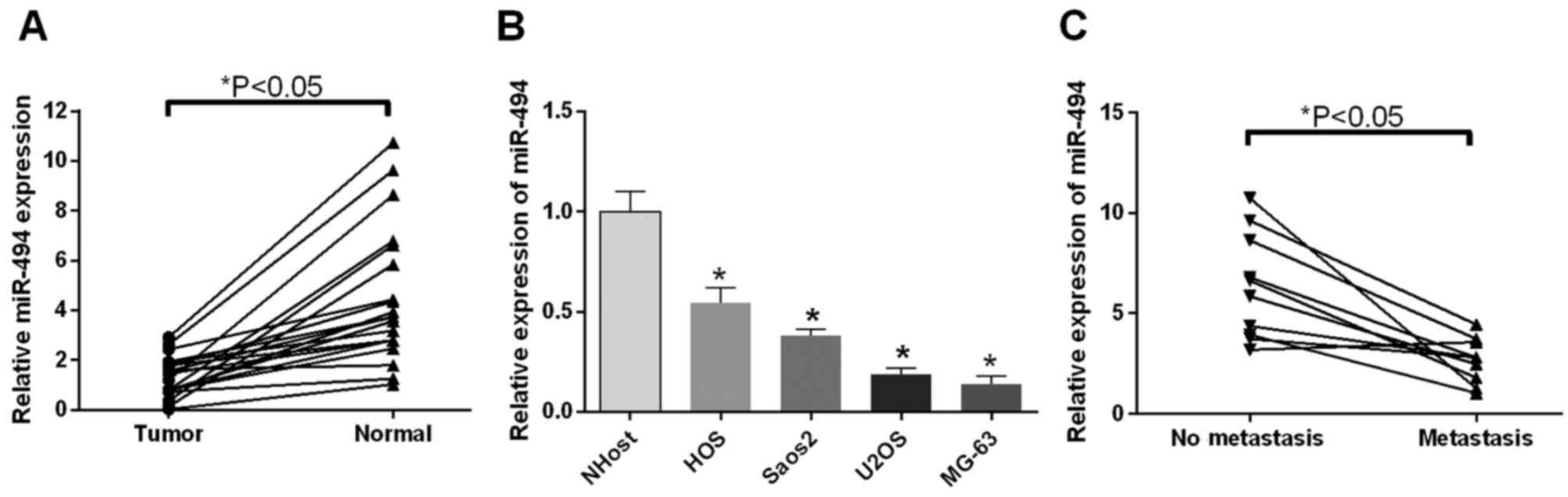
miR-494 is downregulated in OS tissues
and cells
The present study used RT-qPCR analysis to examine
the expression profiles of miR-494 in OS tumor tissues (metastatic,
vs. non-metastatic), cell lines and corresponding controls. It was
found that the expression levels of miR-494 in the 18 patients were
significantly downregulated, compared with those in the normal
tissues (P<0.05; Fig. 1A). In
accordance, the expression levels of miR-494 in the HOS, Saos2,
U2OS, MG-63 OS cells were significantly downregulated, compared
with that in the NHOst cells (P<0.05; Fig. 1B). The association between cell
metastasis and the expression of miR-494 was also investigated. The
results suggested that the expression of miR-494 was decreased in
the metastatic group, compared with the non-metastatic group
(P<0.05; Fig. 1C). These
findings indicated that the expression of miR-494, which was
downregulated in OS tissues and cells, was closely linked to tumor
metastasis.
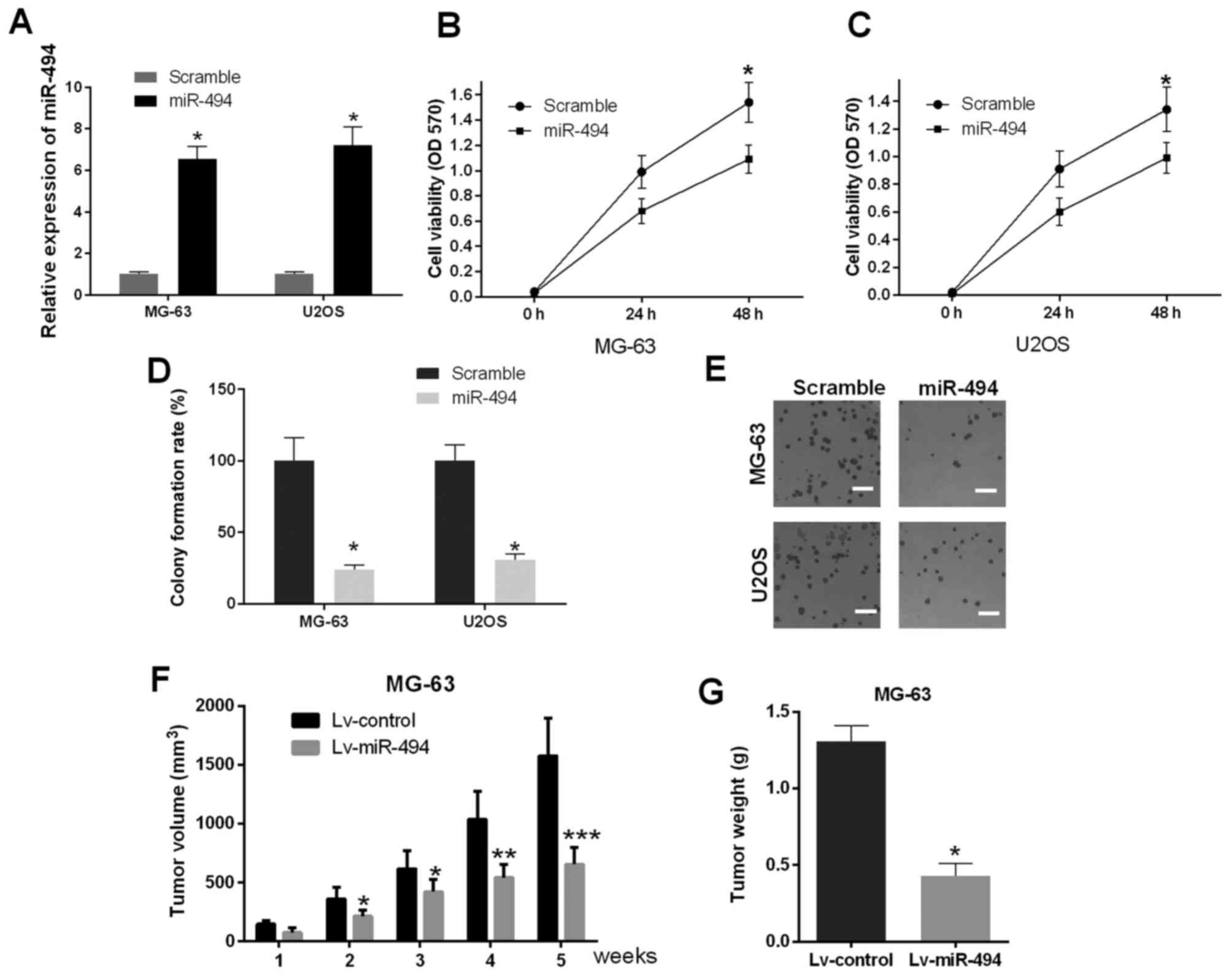
Restoration of the expression of
miR-494 inhibits proliferation in vitro and in vivo
As primary cells from human OS tissues are difficult
to culture, MG-63 and U2OS cells were selected for the subsequent
experiments, due to their routine maintenance in cell culture and
ubiquitous use in investigations of OS (21,22).
In addition, MG-63 and U2OS cells are OS-derived human cell lines
with differing proliferation potential, providing the opportunity
to examine different types of the disease: MG-63 cells exhibit a
low level of proliferation, whereas U2OS cells exhibit more
malignant proliferation potential (23). In the present study, miR-494 mimics
were transfected into MG-63 and U2OS cells, MTT and cell colony
formation assays were used to investigate the effect of the
promotion of miR-494 on cell proliferation in vitro (48 h
post-transfection). The results of the RT-qPCR analysis showed that
the expression of miR-494 was significantly elevated in the cells
transfected with miR-494 mimics (P<0.05; Fig. 2A). The results of the MTT assay
suggested that the ectopic expression of miR-494 resulted in cell
growth inhibition, compared with the scramble control group in
MG-63 and U2OS cells (P<0.05; Fig.
2B and C). The results of the cell colony formation assays
confirmed that the cells transfected with miR-494 mimics exhibited
decreased cell numbers (P<0.05; Fig. 2D and E), which was in accordance
with the MTT assays. To further determine the underlying regulatory
effects of miR-494 in vivo, female nude mice were
subcutaneously injected with miR-494 lentivirus (Lv-miR-494) and
Lv-control, respectively. The data indicated that injection of
miR-494 significantly suppressed tumor volume and weight, compared
with those in the Lv-control group, and this effect was
time-dependent. (P<0.05, P<0.01 and P<0.001; Fig. 2F and G). Taken together, these
findings demonstrated that the ectopic expression of miR-494 caused
marked cell growth inhibition in vitro and in
vivo.
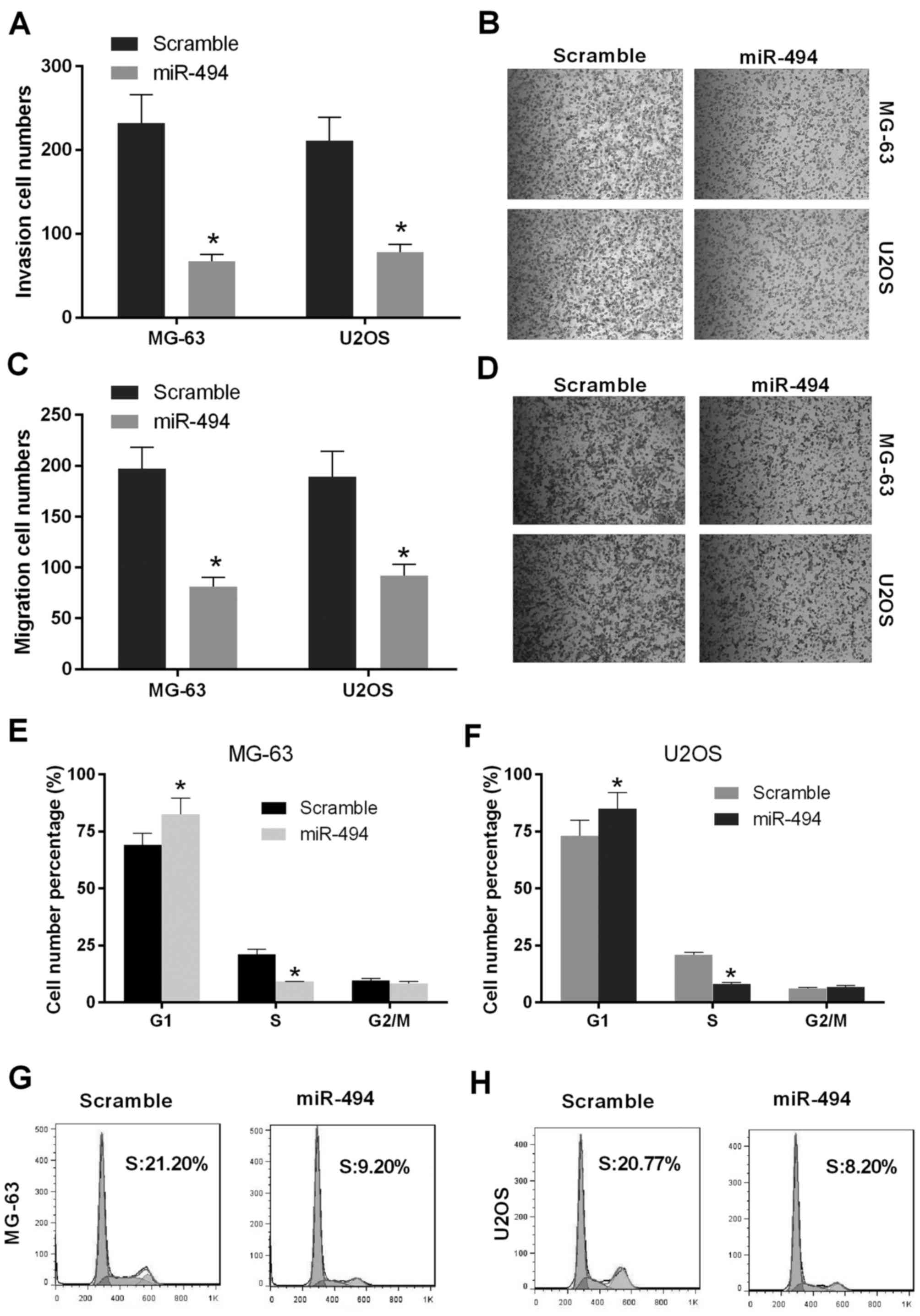
Overexpression of miR-494 inhibits
cell metastasis and induces cell cycle arrest in OS cells
As the overexpression of miR-494 inhibited
proliferation in vitro and in vivo, the present study
performed invasion and migration assays to evaluate the effect of
the restoration of miR-494 on cell metastasis. As shown in Fig. 3A and B, the numbers of MG-63 and
U2OS cells in the miR-494 group were decreased, compared with the
number in the scramble group (P<0.05), which indicated that cell
invasion in the miR-494 group was repressed relative to the
control. In addition, the numbers of migrated MG-63 and U2OS cells
in the miR-494 group were decreased, compared with the number in
the scramble group, which was consistent with the results of the
cell invasion assay (P<0.05; Fig.
3C and D). The effect of miR-494 on cell cycle progression was
then investigated in MG-63 and U2OS cells. The results showed that
the ectopic expression of miR-494 in the MG-63 cells and U2OS cells
significantly increased in the number of G1-phase cells and reduced
the number of S-phase cells (P<0.05; Fig. 3E-H). Taken together, these findings
suggested that miR-494 inhibited cell metastasis and induced cell
cycle arrest of the OS cells.
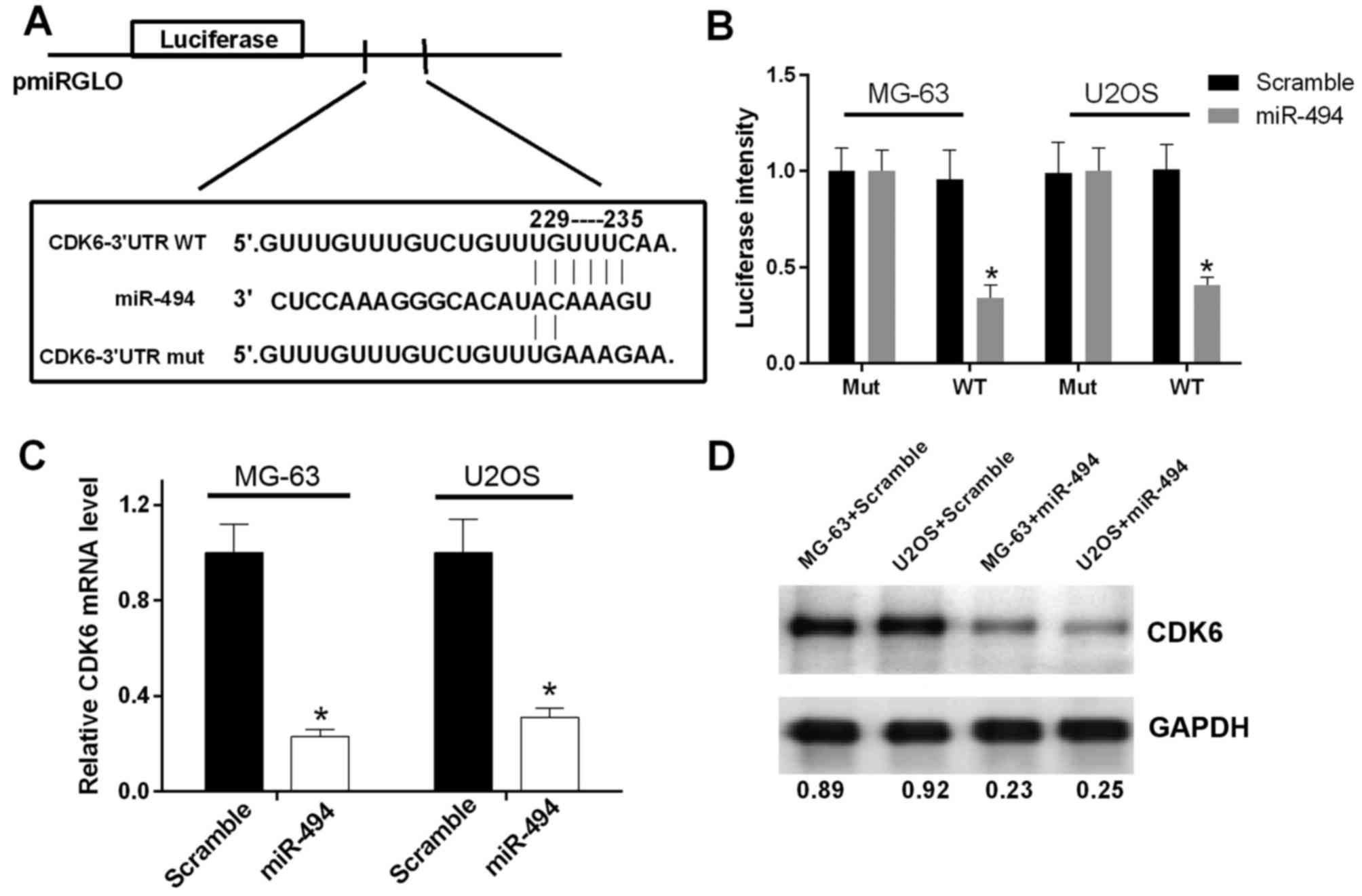
miR-494 directly targets CDK6 and
suppresses its expression
As miR-494 has been reported to be involved in tumor
progression via base-pairing to the 3′-UTR of its target genes,
bioinformatics analysis was performed using PicTar, TargetScan and
miRBase to predict the potential target of miR-494. The results
revealed an miR-494 binding site in the CDK6 3′-UTR (Fig. 4A). In order to validate whether the
3′-UTR of CDK6 is a functional target of miR-494, a luciferase
reporter assay was performed. The results showed that the promoted
expression of miR-494 suppressed the luciferase activity of the
CDK6 3′-UTR-WT construct, but not the CDK6 3′-UTR-mut construct in
OS cells (P<0.05; Fig. 4B). In
addition, RT-qPCR and western blot analyses were performed to
evaluate the effect of promoted miR-494 on the expression of CDK6.
The elevated expression of miR-494 resulted in reductions in the
mRNA and protein expression of CDK6 in the MG-63 and U2OS cells
(P<0.05; Fig. 4C and D). These
data indicated that miR-494 directly targeted CDK6 and negatively
regulated the expression of CDK6.
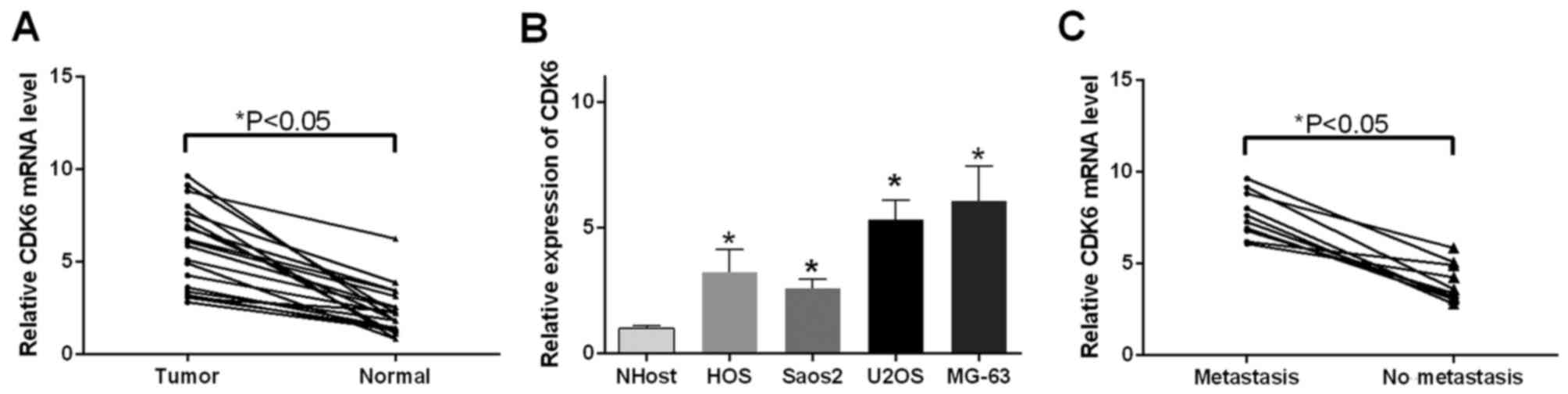
CDK6 is upregulated in OS tissues and
cells
As it was found that miR-494 directly targeted CDK6
and regulated the expression of CDK6, the present study
investigated the expression pattern of CDK6 in the OS tissue and
cells. The results of the RT-qPCR analysis indicated that CDK6 mRNA
was upregulated in the 18 OS tissue samples (P<0.05; Fig. 5A) and OS cells (P<0.05; Fig. 5B), compared with the corresponding
controls, respectively. In addition, the expression of CDK6 was
examined in tissues with differing features of metastasis. It was
observed that the tissues in the metastasis group exhibited
increased levels of CDK6, compared with those in the no metastasis
group (P<0.05; Fig. 5C).
Discussion
An increasing number of studies have focused on the
pathophysiological and physiological mechanisms of miRNAs in tumor
progression. miRNAs can either positively or negatively affect the
development of tumors depending on their specific downstream target
genes by base-pairing with their 3′-UTR. For example, miR-24 can
induce chemotherapy resistance and hypoxic advantage in breast
cancer through the downregulation of factor inhibiting HIF-1
(24). miR-497 is significantly
correlated with temozolomide-resistance in glioma cells by
regulating the insulin-like growth factor 1 receptor/insulin
receptor substrate 1 pathway (25). miR-93 is known to affect metastatic
spread in breast carcinoma through the regulation of protein kinase
WNK1 (26). miR-152 has been
demonstrated to target phosphatase and tensin homolog in
nasopharyngeal carcinoma cells by promoting cell migration and
inhibiting apoptosis (27). The
abnormal expression of miRNAs has also been shown to be involved in
the pathogenesis and progression of OS. For example, miR-150 has
been reported as a tumor suppressor in the development of OS, by
targeting insulin-like growth factor 2 mRNA-binding protein 1
(28). miR-92a functions as a
driver of tumor progression, which can promote tumor growth in OS
by suppressing F-box and WD repeat-containing protein 7 (29). The present study aimed to
investigate the potential role of miR-494 in the development of
OS.
The function ofmiR-494 as a tumor suppressor in OS
was identified in the present study, which induced cell growth
inhibition through the regulation of CDK6. The malignant phenotype
of tumors is closely linked to the irregular proliferation of
cells. Several studies have focused on the diagnostic and
therapeutic targets of cytological features. The results of these
investigations provide evidence that the ectopic expression of
miR-494-3p in PC-3 and DU145 cells inhibits proliferation and
metastasis, and sell apoptosis (30). In addition, miR-494 exhibits a
suppressive function in gastric cancer by inhibiting cell invasion
and proliferation (31). In
accordance, the findings of the present study indicated that the
overexpression of miR-494 resulted in cell growth inhibition and
cell cycle arrest at the G1/S phase in MG-63 and U2OS cells. In
addition, the data obtained from animal experiments indicated that
the injection of miR-494 significantly suppressed tumor volume and
weight in vivo. In separate experiments, patients were
divided into two groups according to metastasis, and radiological
results were examined to determine the correlation between the
expression of miR-494 and cell metastasis. The data indicated that
the group exhibiting metastasis exhibited inhibited expression of
miR-494, compared with the group without metastasis. Taken
together, these results indicated the decreased expression of
miR-494 was involved in the development of OS.
The present study also performed bioinformatics
analysis and luciferase reporter gene assay to examine the
potential targets of miR-494. The results showed that CDK6 was a
direct target of miR-494. CDK6 is vital in cell cycle progression,
and inhibition of CDK6 leads to uncontrolled tumor cell
proliferation, which is a major hallmark of cancer (32). The function of CDK6 in cell
progression has been found in several types of cancer, including
glioblastoma, myxofibrosarcoma and lymphoid malignancies (33,34).
In accordance with these findings, the results of the present study
suggested that CDK6 was significantly upregulated in OS tissues and
cells, compared with corresponding controls. The expression of CDK6
was further investigated in tissues of differing malignant
phenotypes (metastasis and no metastasis) and found that the
metastatic tumors exhibited elevated expression of CDK6. Taken
together, these data indicated that CDK6 promoted the progression
of OS. miR-494 was shown to exert a tumor suppressive function in
the development of OS, and CDK6, which functions as an oncogene in
OS, is a direct target of miR-494. The role of miR-494 in OS
tissues and cells may be mediated by CDK6. However, the exact
effect of CDK6 dysregulation was not investigated in the present
study.
In conclusion, the results of the present study
suggested that the miR-494-induced inhibited expression of CDK6 in
OS may be a useful epigenetic therapeutic approach, although
further experiments are required to determine this.
Acknowledgements
This work was supported by grants from the Natural
Science Foundation of China (NSFC 81672230 and 81501867) and the
Key Project of Science and Technology Commission of Chongqing (CTSC
2013jcyjA10090).
References
|
1
|
Luetke A, Meyers PA, Lewis I and Juergens
H: Osteosarcoma treatment-where do we stand? A state of the art
review. Cancer Treat Rev. 40:523–532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Benjamin RS: Osteosarcoma: Better
treatment through better trial design. Lancet Oncol. 16:12–13.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xiong Y, Wu S, Du Q, Wang A and Wang Z:
Integrated analysis of gene expression and genomic aberration data
in osteosarcoma (OS). Cancer Gene Ther. 22:524–529. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lulla RR, Costa FF, Bischof JM, Chou PM,
de F, Bonaldo M, Vanin EF and Soares MB: Identification of
differentially expressed micrornas in osteosarcoma. Sarcoma.
2011:7326902011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marina N, Gebhardt M, Teot L and Gorlick
R: Biology and therapeutic advances for pediatric osteosarcoma.
Oncologist. 9:422–441. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Romano F, Garancini M and Uggeri F,
Degrate L, Nespoli L, Gianotti L, Nespoli A and Uggeri F: Surgical
treatment of liver metastases of gastric cancer: State of the art.
World J Surg Oncol. 10:1572012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Olaru AV, Ghiaur G, Yamanaka S, Luvsanjav
D, An F, Popescu I, Alexandrescu S, Allen S, Pawlik TM, Torbenson
M, et al: MicroRNA down-regulated in human cholangiocarcinoma
control cell cycle through multiple targets involved in the G1/S
checkpoint. Hepatology. 54:2089–2098. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hong KJ, Wu DC, Cheng KH, Chen LT and Hung
WC: RECK inhibits stemness gene expression and tumorigenicity of
gastric cancer cells by suppressing ADAM-mediated Notch1
activation. J Cell Physiol. 229:191–201. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ma YB, Li GX, Hu JX, Liu X and Shi BM:
Correlation of miR-494 expression with tumor progression and
patient survival in pancreatic cancer. Genet Mol Res.
14:18153–18159. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tian C, Zheng G, Zhuang H, Li X, Hu D, Zhu
L, Wang T, You MJ and Zhang Y: MicroRNA-494 activation suppresses
bone marrow stromal cell-mediated drug resistance in acute myeloid
leukemia cells. J Cell Physiol. 232:1387–1395. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lim L, Balakrishnan A, Huskey N, Jones KD,
Jodari M, Ng R, Song G, Riordan J, Anderton B, Cheung ST, et al:
MicroRNA-494 within an oncogenic microRNA megacluster regulates
G1/S transition in liver tumorigenesis through suppression of
mutated in colorectal cancer. Hepatology. 59:202–215. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao X, Zhou Y, Chen YU and Yu F: miR-494
inhibits ovarian cancer cell proliferation and promotes apoptosis
by targeting FGFR2. OncolLett. 11:4245–4251. 2016.
|
|
13
|
Yuan J, Wang K and Xi M: miR-494 inhibits
epithelial ovarian cancer growth by targeting c-Myc. Med Sci Monit.
22:617–624. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tian C, Zheng G, Zhuang H, Li X, Hu D, Zhu
L, Wang T, You MJ and Zhang Y: MicroRNA-494 activation suppresses
bone marrow stromal cell-mediated drug resistance inacute myeloid
leukemia cells. J Cell Physiol. 232:1387–1395. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rader J, Russell MR, Hart LS, Nakazawa MS,
Belcastro LT, Martinez D, Li Y, Carpenter EL, Attiyeh EF, Diskin
SJ, et al: Dual CDK4/CDK6 inhibition induces cell-cycle arrest and
senescence in neuroblastoma. Clin Cancer Res. 19:6173–6182. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Krek A, Grün D, Poy MN, Wolf R, Rosenberg
L, Epstein EJ, MacMenamin P, da Piedade I, Gunsalus KC, Stoffel M
and Rajewsky N: Combinatorial microRNA target predictions. Nat
Genet. 37:495–500. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:2015. View Article : Google Scholar
|
|
19
|
Griffiths-Jones S, Saini HK, van Dongen S
and Enright AJ: miRBase: Tools for microRNA genomics. Nucleic Acids
Res. 36(Database issue): D154–D158. 2008.PubMed/NCBI
|
|
20
|
Liu S and Feng P: miR-203 determines poor
outcome and suppresses tumor growth by targeting tbk1 in
osteosarcoma. Cell Physiol Biochem. 37:1956–1966. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vanas V, Haigl B, Stockhammer V and
Sutterlüty-Fall H: MicroRNA-21 increases proliferation and
cisplatin sensitivity of osteosarcoma-derived cells. PLoS One.
11:e01610232016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yao J, Qin L, Miao S, Wang X and Wu X:
Overexpression of miR-506 suppresses proliferation and promotes
apoptosis of osteosarcoma cells by targeting astrocyte elevated
gene-1. Oncol Lett. 12:1840–1848. 2016.PubMed/NCBI
|
|
23
|
Li Y, Liu J, Liu ZZ and Wei WB:
MicroRNA-145 inhibits tumour growth and metastasis in osteosarcoma
by targeting cyclin-dependent kinase, CDK6. Eur Rev Med Pharmacol
Sci. 20:5117–5125. 2016.PubMed/NCBI
|
|
24
|
Roscigno G, Puoti I, Giordano I,
Donnarumma E, Russo V, Affinito A, Adamo A, Quintavalle C, Todaro
M, Vivanco MD and Condorelli G: miR-24 induces chemotherapy
resistance and hypoxic advantage in breast cancer. Oncotarget.
8:19507–19521. 2017.PubMed/NCBI
|
|
25
|
Zhu D, Tu M, Zeng B, Cai L, Zheng W, Su Z
and Yu Z: Up-regulation of miR-497 confers resistance to
temozolomide in human glioma cells by targeting mTOR/Bcl-2. Cancer
Med. 6:452–462. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shyamasundar S, Lim JP and Bay BH: miR-93
inhibits the invasive potential of triple-negative breast cancer
cells in vitro via protein kinase WNK1. Int J Oncol. 49:2629–2636.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang S, Li X and Zhu H: MicroRNA-152
targets phosphatase and tensin homolog to inhibit apoptosis and
promote cell migration of nasopharyngeal carcinoma cells. Med Sci
Monit. 22:4330–4337. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qu Y, Pan S, Kang M, Dong R and Zhao J:
MicroRNA-150 functions as a tumor suppressor in osteosarcomaby
targeting IGF2BP1. Tumour Biol. 37:5275–5284. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang X, Li X, Wu F, Gao H, Wang G, Zheng
H, Wang H, Li J and Chen C: Overexpression of miR-92a promotes the
tumor growth of osteosarcoma by suppressing F-box and WD
repeat-containing protein 7. Gene. 606:10–16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shen PF, Chen XQ, Liao YC, Chen N, Zhou Q,
Wei Q, Li X, Wang J and Zeng H: MicroRNA-494-3p targets CXCR4 to
suppress the proliferation, invasion and migration of prostate
cancer. Prostate. 74:756–767. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao XQ, Liang TJ and Fu JW: miR-494
inhibits invasion and proliferation of gastric cancer by targeting
IGF-1R. Eur Rev Med Pharmacol Sci. 20:3818–3824. 2016.PubMed/NCBI
|
|
32
|
Handschick K, Beuerlein K, Jurida L,
Bartkuhn M, Müller H, Soelch J, Weber A, Dittrich-Breiholz O,
Schneider H, Scharfe M, et al: Cyclin-dependent kinase 6 is a
chromatin-bound cofactor for NF-κB-dependent gene expression. Mol
Cell. 53:193–208. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wiedemeyer WR, Dunn IF, Quayle SN, Zhang
J, Chheda MG, Dunn GP, Zhuang L, Rosenbluh J, Chen S, Xiao Y, et
al: Pattern of retinoblastoma pathway inactivation dictates
response to CDK4/6 inhibition in GBM. Proc Natl Acad Sci USA.
107:pp. 11501–11506. 2010; View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tsai JW, Li CF, Kao YC, Wang JW, Fang FM,
Wang YH, Wu WR, Wu LC, Hsing CH, Li SH, et al: Recurrent
amplification at 7q21.2 Targets CDK6 gene in primary
myxofibrosarcomas and identifies CDK6 overexpression as an
independent adverse prognosticator. Ann Surg Oncol. 19:2716–2725.
2012. View Article : Google Scholar : PubMed/NCBI
|