Introduction
Ovarian cancer is the most lethal gynecologic cancer
(1): The five-year survival rate
of patients is <50% post diagnosis (2). First-line clinical treatments for
ovarian cancer patients are cytoreductive surgery and
paclitaxel-based chemotherapy (3).
However, because of multi-drug resistance to chemotherapy, systemic
chemotherapy produces a disappointingly low initial response in
most patients (4). Furthermore,
many commonly used anti-cancer chemotherapeutics have potent
cytotoxic effects in normal cells (5). Therefore, there is an urgent need to
develop effective, non-cytotoxic, chemotherapeutic approaches for
patients with ovarian cancer.
In recent years, many bioactive phytochemicals have
been observed to exhibit anti-cancer activities (6,7).
They demonstrate minimal general toxicity and adverse side effects
and thus may represent potential alternative medicine to
conventional cytotoxic chemotherapy (8,9).
Baicalin is a flavone glycoside found in Scutellaria
baicalensis Georgi, with a chemical formula of
C21H18O11 (10). Baicalin has been reported to have
anti-oxidation, anti-proliferation, anti-inflammation and antitumor
effects (11–13). With respect to cancer, it has been
reported to inhibit the proliferation of various cancer cells
through induction of apoptosis and inhibition of migration
(14–16). However, the effects of baicalin on
ovarian cancer cells and the underlying molecular mechanisms are
still not clear.
The present study aimed to evaluate whether baicalin
could exert antitumor effects on ovarian cancer cells and to
explore the molecular mechanism of this process. The data revealed
that baicalin dose-dependently induced apoptosis and significantly
reduced the migration of ovarian cancer cells. Baicalin may,
therefore, be an effective active ingredient for the development of
an effective drug for patients with ovarian cancer.
Materials and methods
Materials, reagents and chemicals
Antibodies against caspase-3 (cat. no. 19677-1-AP),
caspase-9 (cat. no. 10,380-1-AP), B-cell lymphoma 2 apoptosis
regulator (Bcl-2) (cat. no. 12789-1-AP), matrix metallopeptidase
(MMP)-2 (cat. no 10373-2-AP), MMP-9 (cat. no. 10375-1-AP) and
β-actin (cat. no. 20,536-1-AP) were obtained from ProteinTech
Group, Inc. (Chicago, IL, USA). Secondary polyclonal anti-rabbit
horseradish peroxidase (HRP)-conjugated antibodies (cat. no.
111-035-003) were purchased from Jackson ImmunoResearch
Laboratories, Inc. (West Grove, PA, USA). Radioimmunoprecipitation
assay (RIPA) lysis buffer (50 mM Tris pH 7.4, 150 mM NaCl, 1%
NP-40, 0.5% sodium deoxycholate, 0.1% SDS, sodium fluoride and
EDTA) was from Beyotime Institute of Biotechnology (Haimen, China).
The enhanced chemiluminescence (ECL) kit was from GE Healthcare
Life Sciences (Little Chalfont, UK). The Annexin V-conjugated
fluorescein isothiocyanate (FITC) apoptosis detection kit with
propidium iodide (PI) was purchased from Nanjing KeyGen Biotech
Co., Ltd. (Nanjing, China). Transwells were from BD Biosciences
(San Jose, American). MTT (3-(4, 5-dimethyl-2-yl)-2, 5-diphenyl
tetrazolium bromide) and DAPI were obtained from Sigma-Aldrich;
Merck KGaA (Darmstadt, Germany). Baicalin (concentration ≥ 98%) was
bought from the National Pharmaceutical Engineering Center
(Jiangxi, China).
Drug preparation
Baicalin was dissolved in 100% DMSO at a
concentration of 1 M as a stock solution and stored at 4°C, and
diluted in Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) to the required
concentration before each experiment. The final concentration of
DMSO was <0.1% in all baicalin groups.
Cell lines and cell culture
The ovarian cancer cell line A2780 and normal
ovarian cell line IOSE80 were purchased from American Type Culture
Collection (Manassas, VA, USA) and cultured in DMEM medium
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 100 U/ml penicillin (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C in a humidified atmosphere with 5%
CO2.
Cell viability assays
The effect of baicalin on the viability of cells was
detected by MTT assay. The cells (1×104 cells/well) were
seeded into 96-well plates and incubated for 24 h. Following 24 h
treatment with 0 (control) 40, 80, 120, 160, 200 and 240 µM
baicalin, cell viability was detected by adding 20 µl of MTT
solution (5 mg/ml in PBS) to each well and incubating the mixtures
for 4 h at 37°C. The MTT solution was then removed and 150 µl of
dimethyl sulfoxide (DMSO) was added to the wells. The absorbance
was measured using a Multiskan Ascent plate reader (Thermo Fisher
Scientific, Inc.) at a wavelength of 540 nm.
DAPI staining assay
To assess the effect of baicalin on the nuclei of
ovarian cancer cells, ~4×104 cells/well were treated
with baicalin at 0, 80 or 160 µM for 24 h. Cells in each well were
then stained with DAPI before fixation with 3.7% formaldehyde at
room temperature for 15 min. The cells were then washed with PBS
and detected by fluorescence microscopy. From each sample, 3 visual
fields were randomly selected for evaluation.
Cell apoptosis by flow cytometry
The extent of apoptosis was evaluated by flow
cytometry using an Annexin V-FITC/PI apoptosis detection kit (cat.
no. CA1020, Solarbio, Beijing, China). Following treatment with
either 0, 80 or 160 µM baicalin for 24 h, ovarian cancer cells
(1×106 cells/well) were harvested and washed thrice with
PBS, then incubated with Annexin V-FITC and PI at room temperature
for 10 min in the dark. The cells were detected using a BD Accuri™
C6 flow cytometer and analyzed using BD Accuri™ C6 Software version
1.0.264.21 (BD Biosciences, Franklin Lakes, NJ, USA).
Wound healing assays
A2780 cells (1×105 cells/well) were
seeded into 24-well plates and scraped with the end of 200 µl
pipette tips. The plates were washed with PBS to remove detached
cells and then incubated with the complete growth medium containing
either 0, 20 or 40 µM baicalin solution for 24 h. Cell migration
was observed under a phase-contrast microscope at 100×
magnification at 0 and 24 h post-induction of injury. Migrated
cells in the denuded area in each of six random fields were
measured and quantified using Image J software version 1.50
(National Institutes of Health, Bethesda, MD, USA).
Transwell migration assays
Cell migration was quantified by transwell assays.
Ovarian cancer cells were treated with 0, 20 or 40 µM baicalin for
24 h and harvested. A total of 2×104 cells in serum-free
DMEM were added to each upper chamber and DMEM medium with 10% FBS
was added to the lower chamber as a chemoattractant. After 24 h
incubation at 37°C, cells remaining on the upper surface of
membrane were removed and the cells that had migrated to the
underside of the membrane were stained with 0.1% crystal violet for
10 min. The migrated cells on the underside of the membrane were
counted under a light microscope under a 200× magnification field.
A total of 6 random fields of each transwell membrane were counted
and averaged.
Western blot analysis
Total protein was extracted from the cells samples
using RIPA lysis buffer (Beyotime Institute of Biotechnology) with
protease inhibitors (Biocolor Ltd., Beijing, China) in a proportion
of 1:100. Keep on ice for 5 min, swirling the plate occasionally
for uniform spreading. Centrifuge samples at 4°C, 12,000 × g for 15
min, transfer supernatant for further analysis. Equal amounts of
protein (25 µg) were loaded on a 10% SDS-PAGE gel. The lysates were
resolved by electrophoresis (80 V for 30 min and 120 V for 1.5 h)
and transferred onto polyvinylidene difluoride membranes (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Membranes were blocked in
5% nonfat milk for 1 h at room temperature and then incubated with
primary antibodies against caspase3 (1:1,000), caspase9 (1:1,000),
Bcl-2 (1:1,000), MMP-2 (1:200), MMP-9 (1:200) or β-actin (1:1,000)
in blocking buffer overnight at 4°C. This was followed by
incubation with relevant secondary polyclonal anti-rabbit
HRP-conjugated antibody (1:5,000) for 1 h at room temperature.
Protein bands were visualized using a Chemiluminescent ECL assay
kit (GE Healthcare Life Sciences) and the Bio-Rad ChemiDoc XRS+
image analyzer. Protein expression levels were quantitatively
determined using Image J software version 1.50 (National Institutes
of Health, Bethesda, MD, USA). β-actin was used as internal
reference for protein expression in the treated cells.
Statistical analysis
Data are presented as the mean ± standard deviation
of 3 independent experiments. For each independent experiment, the
assays were performed in duplicate. Statistical differences between
two groups were analyzed using a Student's t-test and multiple
comparison analyses were performed by one-way analysis of variance
followed by Tukey post-hoc testing. Statistical analysis was
performed using GraphPad Prism 5.0 (GraphPad Software, Inc., La
Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Baicalin inhibits ovarian cancer cell
viability
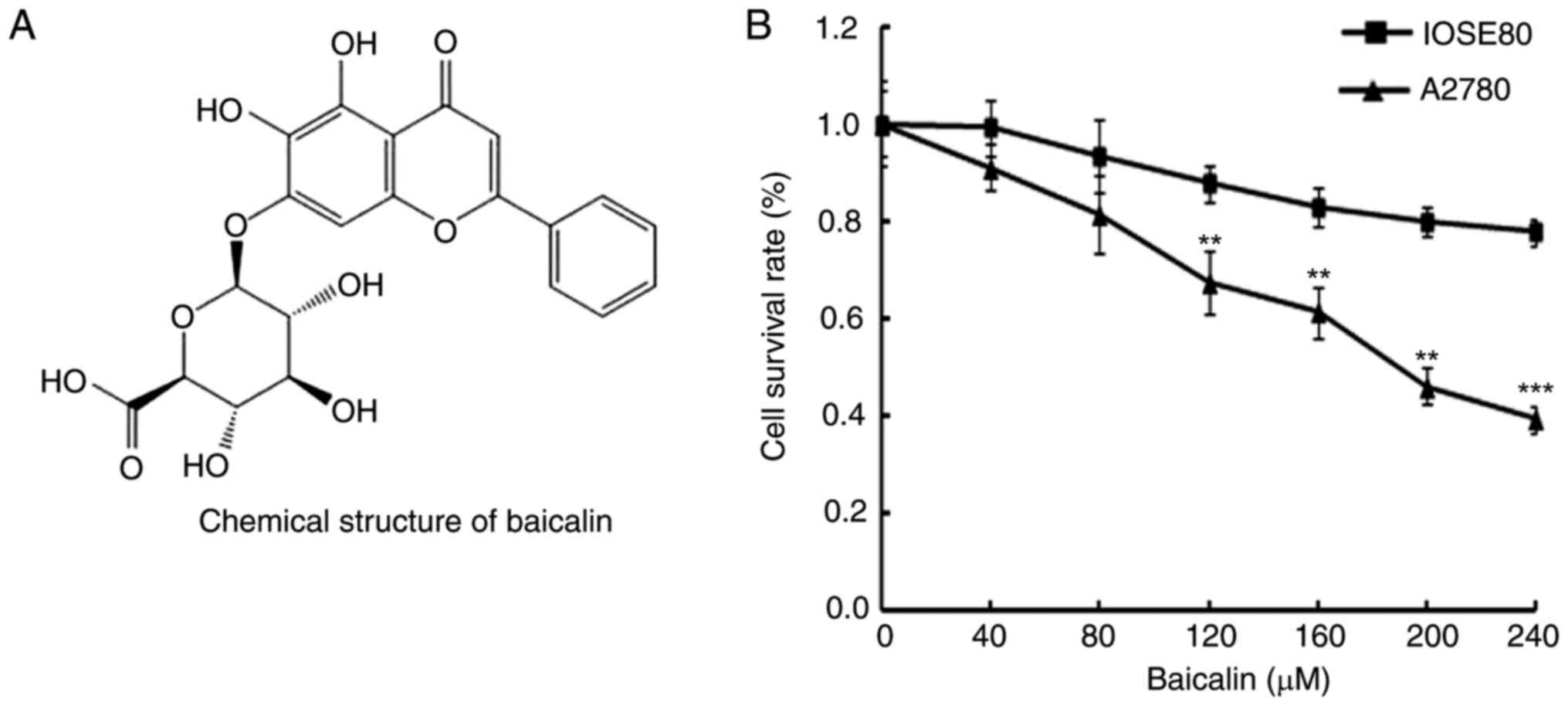
In order to determine the effects of baicalin
(Fig. 1A) on the viability of
A2780 ovarian cancer cells and IOSE80 normal ovarian cells, cells
were treated with 0, 40, 80, 120, 160, 200 and 240 µM baicalin for
24 h, then cell viability was determined by MTT assay. Cancer cells
treated with baicalin revealed significantly reduced viability
compared with untreated cells, in a dose-dependent manner (Fig. 1B). However, baicalin did not affect
the growth of normal ovarian cells (Fig. 1B). These data therefore indicated
that baicalin inhibited the growth of ovarian cancer cells.
Baicalin induces ovarian cancer cell
apoptosis
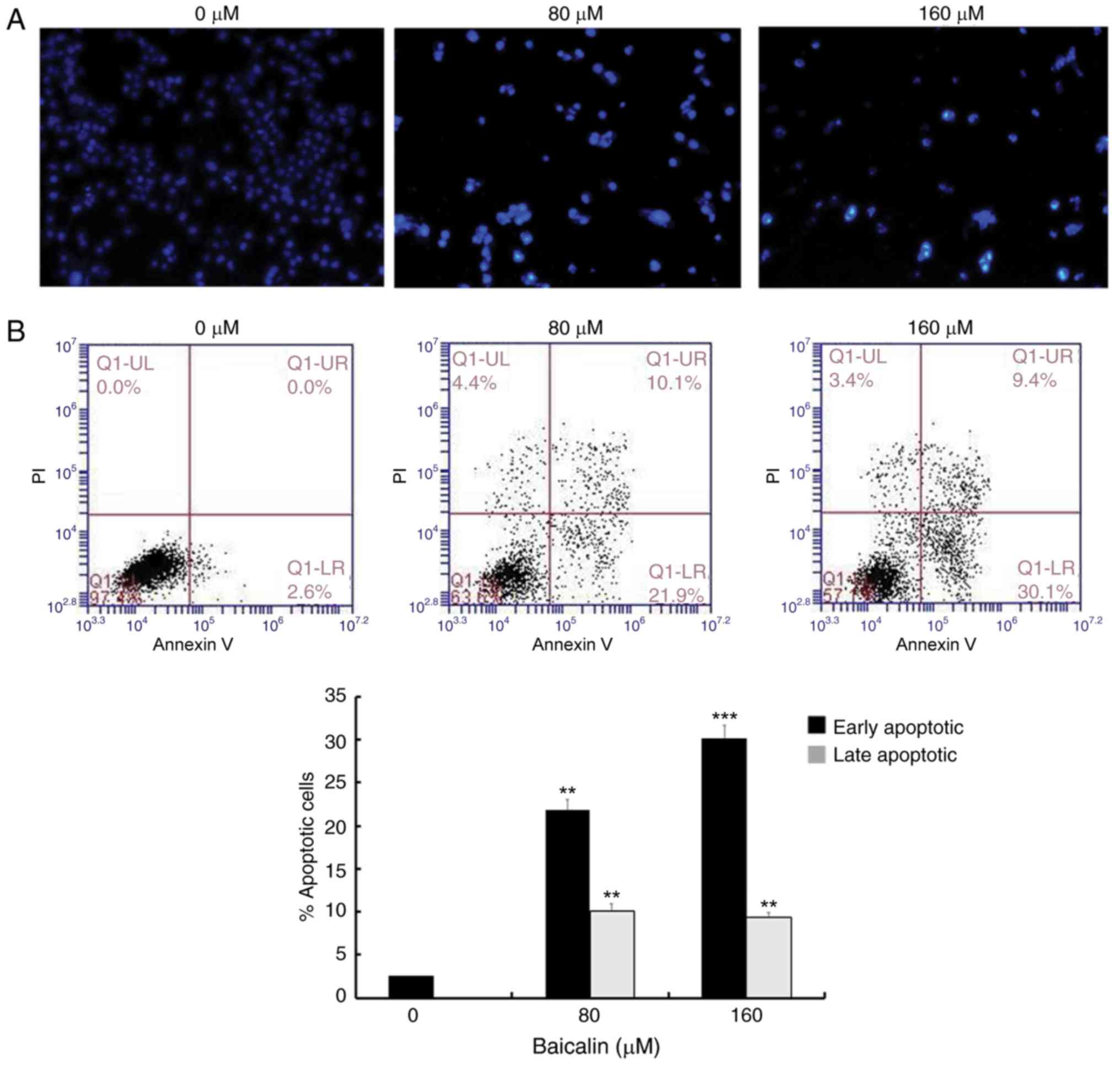
To assess whether the antitumor effects of baicalin
on A2780 ovarian cancer cells were associated with apoptosis, cells
were stained with DAPI and observed under a fluorescence microscope
(Fig. 2A). Nuclear chromatin
condensation and fragmented punctuate blue nuclear fluorescence
were observed in ovarian cancer cells treated with 80 and 160 µM
baicalin for 24 h, in a dose-dependent manner, while the control
cells displayed normal and intact nuclei. This suggested that
baicalin may induce ovarian cancer cell apoptosis. To further
investigate this, apoptosis was analyzed by flow cytometry
(Fig. 2B). The parentage of early-
and late-stage apoptotic cells significantly increased in groups
treated with baicalin compared with untreated control cells
(Fig. 2B): The percentage of total
apoptotic A2780 cells was 2.6% in the control cells (2.6%
early-stage and 0% late-stage), 32% in the cells treated with 80 µM
baicalin (21.9% early-stage and 10.9% late-stage) and 39.5% in the
cells treated with 160 µM baicalin (30.1% early-stage and 9.4%
late-stage). These results demonstrated that baicalin induces
apoptosis in A2780 ovarian cancer cells.
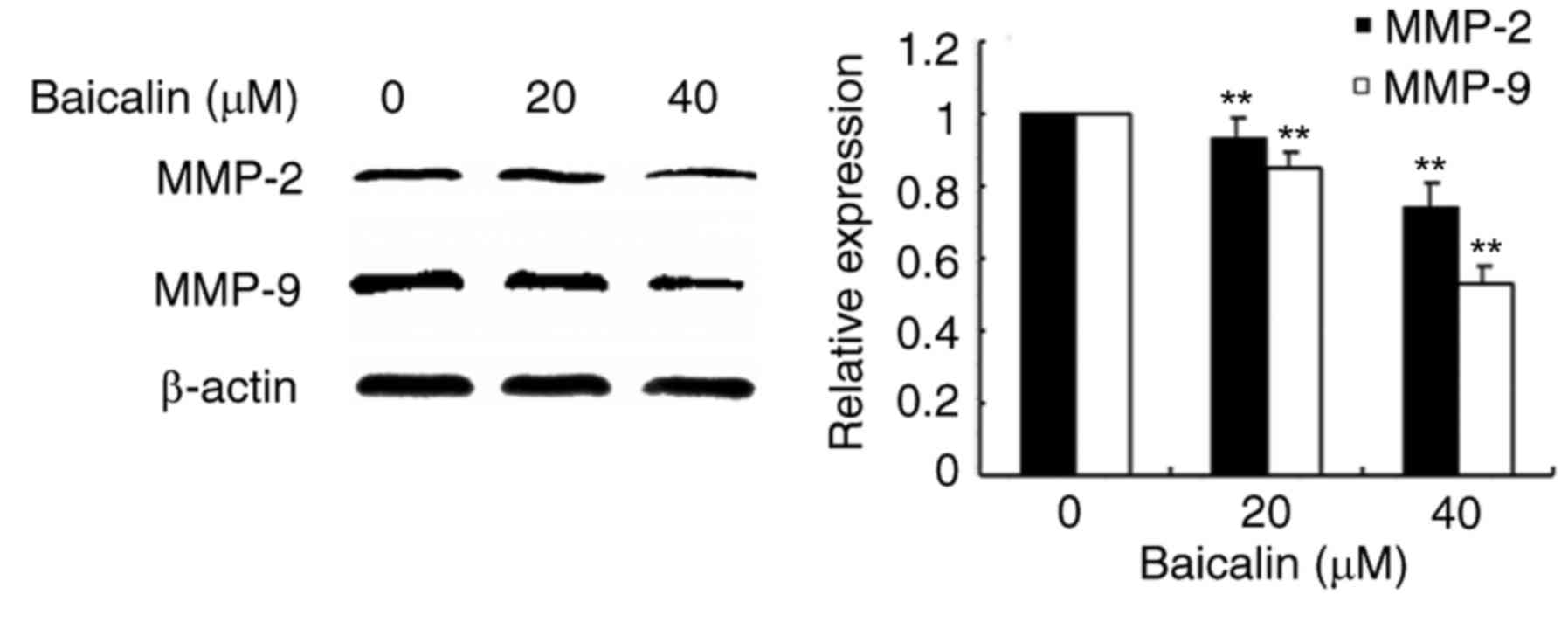
Baicalin suppresses migration of
ovarian cancer cells by antagonizing MMP-9 expression
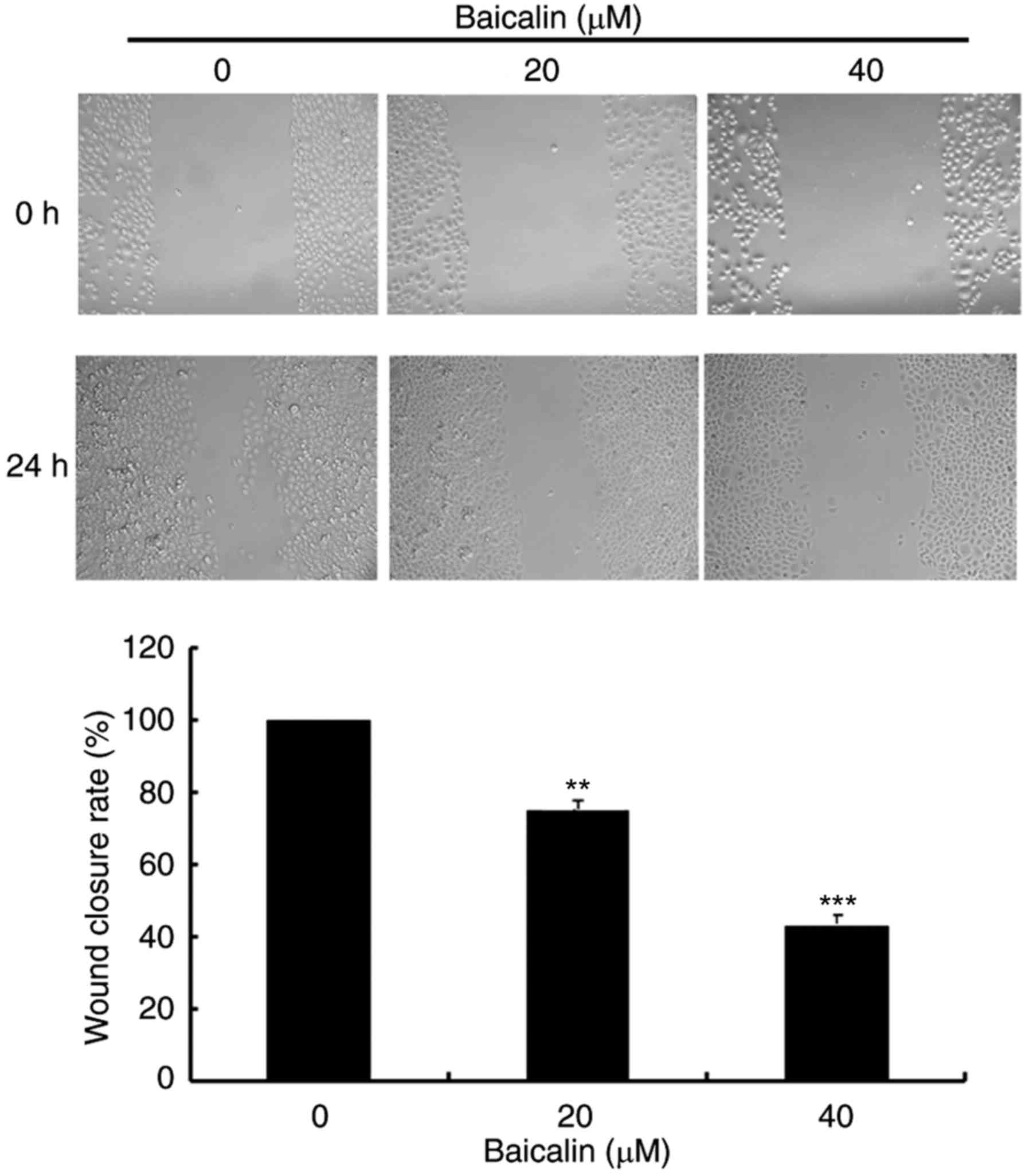
To evaluate the effects of baicalin on cell
migration, wound healing assays and transwell assays were
performed. In the wound healing assay, baicalin dose-dependently
significantly decreased the migration of A2780 cells compared with
the untreated control (Fig. 3).
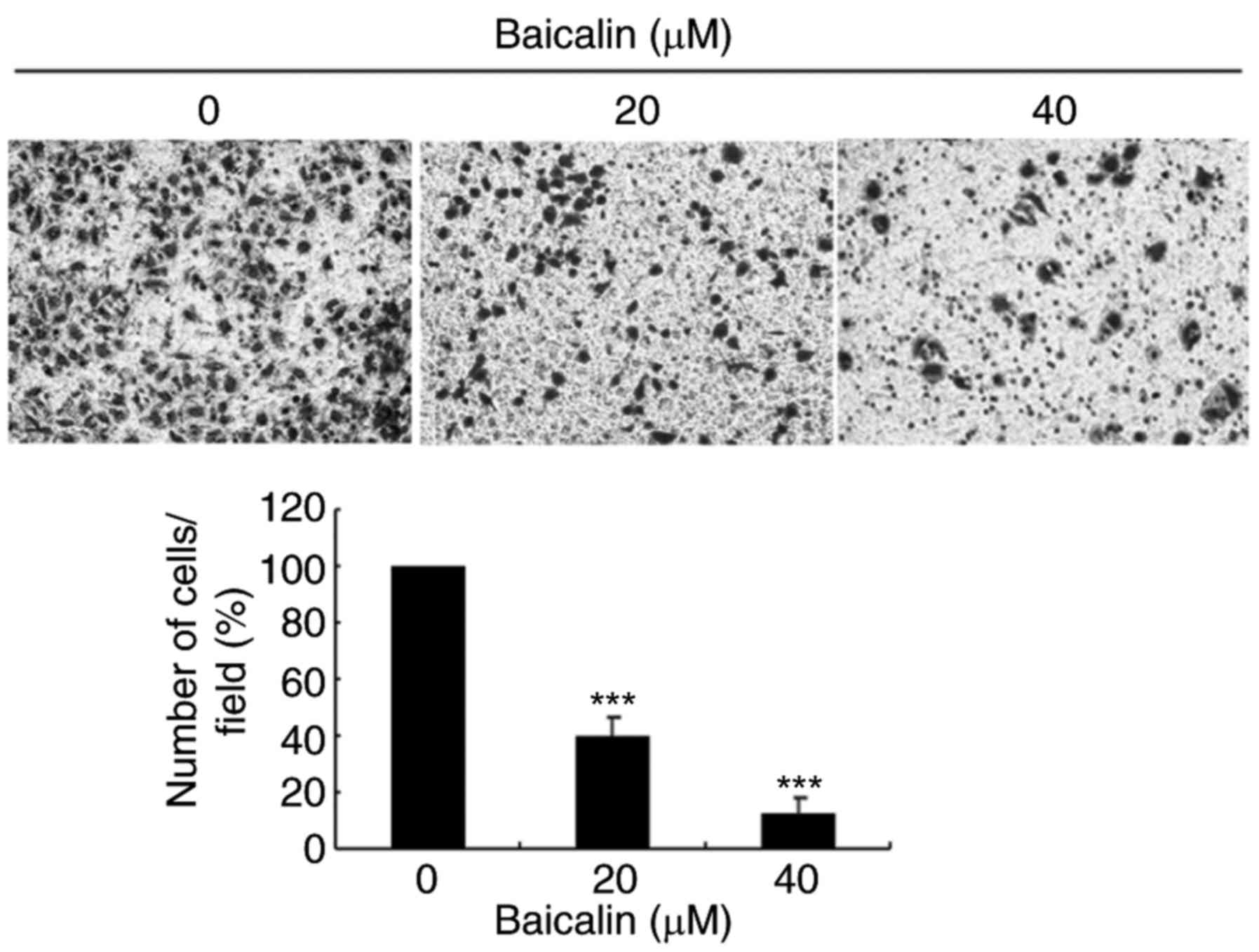
Likewise, baicalin significantly inhibited ovarian cancer cell
migration in a 24 h transwell assay, in a dose-dependent manner,
compared with untreated control cells (Fig. 4). Treatment with 20 and 40 µM
baicalin inhibited A2780 cells migration by 59 and 87% respectively
(P<0.001; Fig. 4). The wound
healing and transwell chamber assays both suggested that baicalin
suppresses the migration of ovarian cancer cells. Given the effects
of baicalin on ovarian cancer cell migration, the mechanisms of
this process were further investigated. Since MMPs plays an
important role in cancer metastasis, MMP-2 and MMP-9 protein
expression levels were detected by western blot. Baicalin
dose-dependently reduced MMP-2 and MMP-9 protein expression levels
in treated cells compared with untreated cells (Fig. 5). These data therefore suggested
that the inhibitory effect of baicalin on the migration of ovarian
cancer was at least partially associated with downregulation of
MMP-2 and MMP-9 expression.
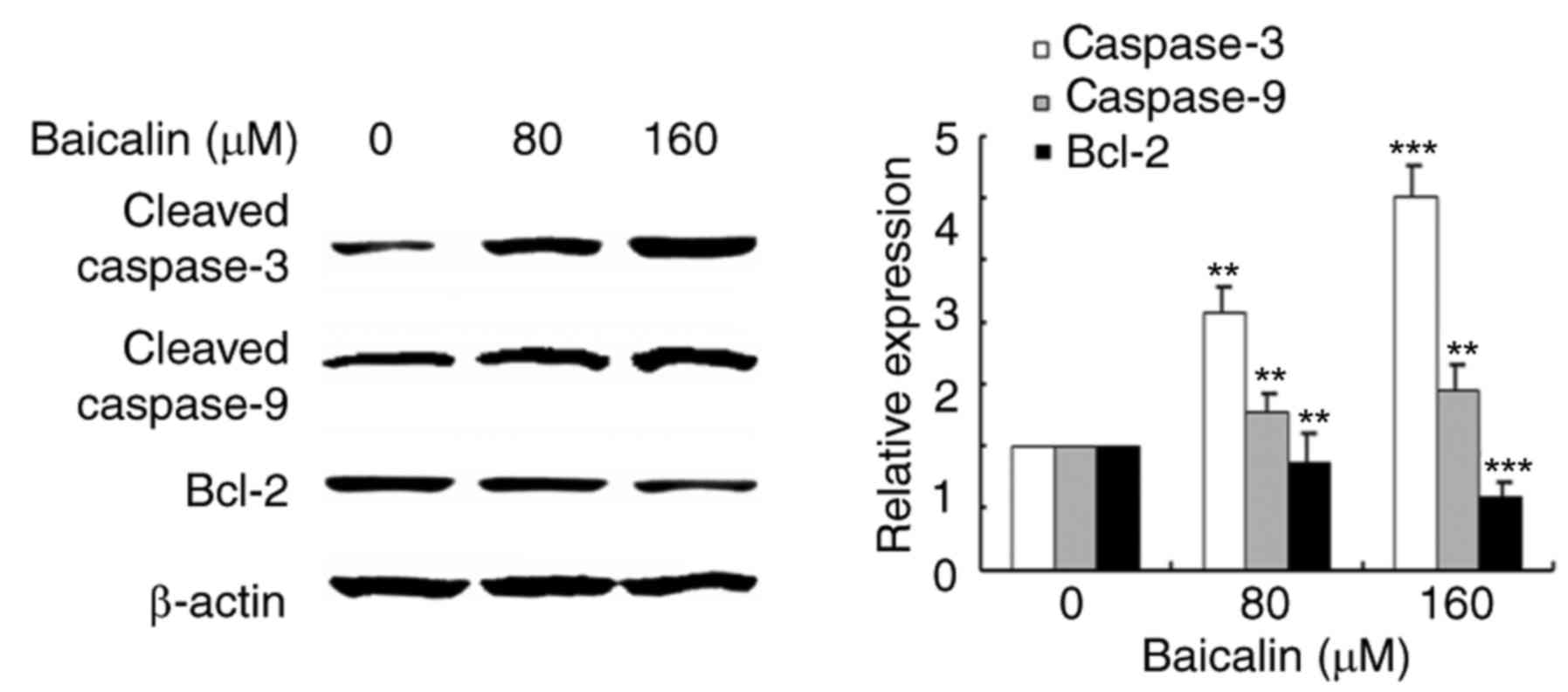
The effects of baicalin on
apoptosis-related proteins
Since baicalin was demonstrated to induce apoptosis
in ovarian cancer cells, the expression and activation of apoptosis
related proteins was investigated by western blotting analysis
(Fig. 6). Protein expression
levels of Bcl-2, an anti-apoptotic protein, decreased in ovarian
cancer cells treated with baicalin in a dose-dependent manner,
compared with untreated control cells. Cleavage of caspase-3 and
caspase-9 was also measured in the study: The results revealed that
cleaved-caspase-3 and cleaved-caspase-9 levels increased in
baicalin-treated ovarian cancer cells compared with untreated
control cells. These results suggested that baicalin may activate
the caspase-dependent apoptosis pathway in A2780 cells.
Discussion
Apoptosis is a fundamental life phenomenon through
the whole process of life (17).
It has been reported that in many human tumor cells, the
proliferation of cells is unrestricted if cells apoptosis is
definitely hindered (18).
Therefore, the antitumor effects of baicalin on ovarian cancer
cells was investigated as a strategy to identify new and effective
drugs for patients with ovarian cancer. Baicalin has previously
been demonstrated to inhibit platelet-derived growth
factor-BB-stimulated vascular smooth muscle cell proliferation
through suppressing β-type platelet-derived growth factor
receptor/extracellular signal-regulated kinase signaling (19). Baicalin, a phytochemical component
of Scutellaria baicalensis Georgi has widespread
applications as anti-inflammatory, anti-hepatitis and
anti-oxidation agent (11,20,21).
Furthermore, the anti-cancer effect of baicalin has also been
previously documented (22). The
present study aimed to explore the effects of baicalin on ovarian
cancer cells and analyze the mechanisms underlining the observed
effects.
Monomer compounds extracted from plants have
previously been reported to induce apoptosis (23–25).
Apoptosis is programmed cell death and plays a vital role in
eliminating mutated or hyper-growing cancer cells. Various natural
compounds have been shown to suppress the growth of tumor cells by
inducing apoptosis (26–28). Therefore, induction of apoptosis
has become the major target of most anti-cancer agents. It has been
reported that baicalin inhibits the proliferation of HeLa cells via
the induction of apoptosis through the intracellular mitochondrial
pathway (29). The present study
indicated that baicalin significantly reduces the viability of
ovarian cancer cells, with no significant effect observed in normal
ovarian cells, and that ovarian cancer cells treated with baicalin
displayed specific apoptotic morphological changes. In addition,
the percentage of early and late apoptotic ovarian cancer cells
significantly increased following treatment with baicalin. Thus,
baicalin may specifically and significantly induce apoptosis of
ovarian cancer cells without affecting normal ovarian cells.
Mitochondrial proteins directly activate cellular apoptotic
programs (30,31). Bcl-2 is involved in the
mitochondria-associated apoptotic pathway (32). Downregulation of Bcl-2 expression
could lead to loss of mitochondrial membrane potential and trigger
a series of apoptotic events such as activation of caspase-9 and
caspase-3, as observed in this study. Peng et al (29) indicated that baicalin-induces
apoptosis in HeLa cells through activation of caspase-3 through the
intracellular mitochondrial pathway and the surface death receptor
pathway, however, it did not show that baicalin could suppress
migration of ovarian cancer cells by antagonizing MMP2/9
expression. The present study demonstrated that baicalin not only
induced ovarian cancer cell apoptosis via the intracellular
mitochondrial pathway but also suppressed they migratory ability of
ovarian cancer cells by antagonizing MMP2/9 expression. These data
suggested that baicalin could induce cell death through the
mitochondria-associated apoptotic pathway in A2780 epithelial
ovarian cancer cells, but requires further confirmation in other
cell models of ovarian cancer.
Additionally, the present study demonstrated that
baicalin effectively suppressed ovarian cancer cell migration.
MMP-9 and MMP-2 belong to the gelatin enzyme class of proteases
(33,34). A recent study has suggested that
the expression of MMP-9 is associated with metastasis in ovarian
cancer (35). Also, the inhibition
of MMP-2 expression suppresses the metastatic potential of prostate
cancer cells (36). The present
study demonstrated that baicalin significantly inhibited ovarian
cancer cell migration and that MMP-9 and MMP-2 protein expression
was reduced by baicalin treatment in a dose-dependent manner. These
findings indicate that baicalin may suppress ovarian cancer cell
migration through downregulation of MMP-9 and MMP-2 expression.
In conclusion, to the best of our knowledge, the
present study is the first to demonstrate that baicalin may
function as a selective antitumor agent for ovarian cancer by
inhibiting cell viability, inducing apoptosis and suppressing
ovarian cancer cell migration. These data thus suggested that
baicalin may potentially be used in the formulation of a novel and
effective antitumor treatment for ovarian cancer patients.
Acknowledgements
We are grateful to Professor Nazir Ahmad from the
University of Agriculture, Faisalabad, Pakistan for detailed
correction of the manuscript. The present study was supported by
grants from the National Natural Science Foundation of China (grant
nos. 31572590, 31772815 and 31502138), Shandong Province (grant no.
BS2015NY001), and Higher Educational Science and Technology Program
of Shandong Province (grant no. J15LF03).
References
|
1
|
Temkin SM, Tanner EJ, Dewdney SB and
Minasian LM: Reducing overtreatment in gynecologic oncology: The
case for less in endometrial and ovarian cancer. Front Oncol.
6:1182016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vernooij F, Heintz P, Witteveen E and van
der Graaf Y: The outcomes of ovarian cancer treatment are better
when provided by gynecologic oncologists and in specialized
hospitals: A systematic review. Gynecol Oncol. 105:801–812. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Markman M: Current status and future
directions of platinum/paclitaxel-based chemotherapy of ovarian
cancer. Semin Oncol. 24(4 Suppl 11): S11–S27. 1997.PubMed/NCBI
|
|
4
|
Fujiwara Y, Takaishi K, Nakao J, Ikeda T,
Katabuchi H, Takeya M and Komohara Y: Corosolic acid enhances the
antitumor effects of chemotherapy on epithelial ovarian cancer by
inhibiting signal transducer and activator of transcription 3
signaling. Oncol Lett. 6:1619–1623. 2013.PubMed/NCBI
|
|
5
|
Roy J, Maltais R, Jegham H and Poirier D:
Libraries of 2β-(N-substituted piperazino)-5α-androstane-3α,
17β-diols: Chemical synthesis and cytotoxic effects on human
leukemia HL-60 cells and on normal lymphocytes. Mol Divers.
15:317–339. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou L, Xu T, Zhang Y, Zhu M, Zhu W, Wang
Z, Gu H, Wang H, Li P, Ying J, et al: Transcriptional network in
ovarian cancer cell line SKOV3 treated with Pinellia pedatisecta
Schott extract. Oncol Rep. 36:462–470. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yung MM, Ross FA, Hardie DG, Leung TH1,
Zhan J, Ngan HY and Chan DW: Bitter Melon (Momordica charantia)
extract inhibits tumorigenicity and overcomes cisplatin-resistance
in ovarian cancer cells through targeting AMPK signaling cascade.
Integr Cancer Ther. 15:376–389. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ahmed KM: The effect of olive leaf extract
in decreasing the expression of two pro-inflammatory cytokines in
patients receiving chemotherapy for cancer. A randomized clinical
trial. Saudi Dent J. 25:141–147. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang HB, Lu P, Cao WB, Zhang ZH and Meng
XL: The effect-enhancing and toxicity-reducing activity of
Hypericum japonicum Thunb. Extract in murine liver cancer
chemotherapy. Mol Clin Oncol. 1:395–399. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang CZ, Zhang CF, Chen L, Anderson S, Lu
F and Yuan CS: Colon cancer chemopreventive effects of baicalein,
an active enteric microbiome metabolite from baicalin. Int J Oncol.
47:1749–1758. 2015.PubMed/NCBI
|
|
11
|
Guo X, Chi S, Cong X, Li H, Jiang Z, Cao R
and Tian W: Baicalin protects sertoli cells from heat
stress-induced apoptosis via activation of the Fas/FasL pathway and
Hsp72 expression. Reprod Toxicol. 57:196–203. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun SJ, Wu XP, Song HL and Li GQ: Baicalin
ameliorates isoproterenol-induced acute myocardial infarction
through iNOS, inflammation, oxidative stress and P38MAPK pathway in
rat. Int J Clin Exp Med. 8:22063–22072. 2015.PubMed/NCBI
|
|
13
|
Yu Y, Pei M and Li L: Baicalin induces
apoptosis in hepatic cancer cells in vitro and suppresses tumor
growth in vivo. Int J Clin Exp Med. 8:8958–8967. 2015.PubMed/NCBI
|
|
14
|
Cortés-Castell E, Veciana-Galindo C,
Torró-Montell L, Palazón-Bru A, Sirvent-Segura E, Gil-Guillén V and
Rizo-Baeza M: Protection by polyphenol extract from olive stones
against apoptosis produced by oxidative stress in human
neuroblastoma cells. Nutr Hosp. 33:118–122. 2016. View Article : Google Scholar
|
|
15
|
Kim GT, Lee SH and Kim YM: Torilis
japonica extract-generated intracellular ROS induces apoptosis by
reducing the mitochondrial membrane potential via regulation of the
AMPK-p38 MAPK signaling pathway in HCT116 colon cancer. Int J
Oncol. 49:1088–1098. 2016.PubMed/NCBI
|
|
16
|
Hwang S, Cho GS, Ryu S, Kim HJ, Song HY,
Yune TY, Ju C and Kim WK: Post-ischemic treatment of WIB801C,
standardized Cordyceps extract, reduces cerebral ischemic injury
via inhibition of inflammatory cell migration. J Ethnopharmacol.
186:169–180. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Afford S and Randhawa S: Apoptosis. Mol
Pathol. 53:55–63. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Martin SJ, Lennon SV, Bonham AM and Cotter
TG: Induction of apoptosis (programmed cell death) in human
leukemic HL-60 cells by inhibition of RNA or protein synthesis. J
Immunol. 145:1859–1867. 1990.PubMed/NCBI
|
|
19
|
Dong LH, Wen JK, Miao SB, Jia Z, Hu HJ,
Sun RH, Wu Y and Han M: Baicalin inhibits PDGF-BB-stimulated
vascular smooth muscle cell proliferation through suppressing
PDGFRbeta-ERK signaling and increase in p27 accumulation and
prevents injury-induced neointimal hyperplasia. Cell Res.
20:1252–1262. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ye C, Li S, Yao W, Xu L, Qiu Y, Liu Y, Wu
Z and Hou Y: The anti-inflammatory effects of baicalin through
suppression of NLRP3 inflammasome pathway in LPS-challenged piglet
mononuclear phagocytes. Innate Immun. 22:196–204. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao W, Liu L, Wang Y, Mao T and Li J:
Effects of a combination of puerarin, baicalin and berberine on the
expression of proliferator-activated receptor-γ and insulin
receptor in a rat model of nonalcoholic fatty liver disease. Exp
Ther Med. 11:183–190. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lange I, Moschny J, Tamanyan K,
Khutsishvili M, Atha D, Borris RP and Koomoa DL: Scrophularia
orientalis extract induces calcium signaling and apoptosis in
neuroblastoma cells. Int J Oncol. 48:1608–1616. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee K, Hong S, Seong GJ and Kim CY:
Cigarette smoke extract causes injury in primary retinal ganglion
cells via apoptosis and autophagy. Curr Eye Res. 41:1367–1372.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ebrahimzadeh-Bideskan AR, Hami J, Alipour
F, Haghir H, Fazel AR and Sadeghi A: Protective effects of ascorbic
acid and garlic extract against lead-induced apoptosis in
developing rat hippocampus. Metab Brain Dis. 31:1123–1132. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim MK, Choi HS, Cho SG, Shin YC and Ko
SG: Rubus coreanus Miquel extract causes apoptosis of
doxorubicin-resistant NCI/ADR-RES ovarian cancer cells via JNK
phosphorylation. Mol Med Rep. 13:4065–4072. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lin R, Li Z, Lin J, Ye J, Cai Q, Chen L
and Peng J: Ethanolic extract of Tulipa edulis Bak induces
apoptosis in SGC-7901 human gastric carcinoma cells via the
mitochondrial signaling pathway. Oncol Lett. 10:2371–2377.
2015.PubMed/NCBI
|
|
27
|
Listyawati S, Sismindari, Mubarika S,
Murti YB and Ikawati M: Anti-proliferative activity and apoptosis
induction of an ethanolic extract of boesenbergia pandurata (Roxb.)
schlecht. Against HeLa and vero cell lines. Asian Pac J Cancer
Prev. 17:183–187. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang C and Wang B: Ginkgo biloba extract
attenuates oxidative stress and apoptosis in mouse cochlear neural
stem cells. Phytother Res. 30:774–780. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peng Y, Fu ZZ, Guo CS, Zhang YX, Di Y,
Jiang B and Li QW: Effects and mechanism of baicalin on apoptosis
of cervical cancer HeLa cells in-vitro. Iran J Pharm Res.
14:251–261. 2015.PubMed/NCBI
|
|
30
|
Liu M, Li SJ, Xin YN, Ji SS, Xie RJ and
Xuan SY: Ferric nitrilotriacetate (Fe-NTA)-induced reactive
oxidative species protects human hepatic stellate cells from
apoptosis by regulating Bcl-2 family proteins and mitochondrial
membrane potential. Int J Clin Exp Med. 8:18074–18081.
2015.PubMed/NCBI
|
|
31
|
Dejean LM, Martinez-Caballero S, Manon S
and Kinnally KW: Regulation of the mitochondrial apoptosis-induced
channel, MAC, by BCL-2 family proteins. Biochim Biophys Acta.
1762:191–201. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Soriano ME and Scorrano L: The interplay
between BCL-2 family proteins and mitochondrial morphology in the
regulation of apoptosis. Adv Exp Med Biol. 687:97–114. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou Y, Zeng YP, Zhou Q, Guan JX and Lu
ZN: The effect of captopril on the expression of MMP-9 and the
prognosis of neurological function in herpes simplex encephalitis
mice. Neurol Res. 38:733–739. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li NA, Wang H, Zhang J and Zhao E:
Knockdown of hypoxia inducible factor-2α inhibits cell invasion via
the downregulation of MMP-2 expression in breast cancer cells.
Oncol Lett. 11:3743–3748. 2016.PubMed/NCBI
|
|
35
|
Che YL, Luo SJ, Li G, Cheng M, Gao YM, Li
XM, Dai JM, He H, Wang J, Peng HJ, et al: The C3G/Rap1 pathway
promotes secretion of MMP-2 and MMP-9 and is involved in serous
ovarian cancer metastasis. Cancer Lett. 359:241–249. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu H, Chen A, Guo F and Yuan L: A
short-hairpin RNA targeting osteopontin downregulates MMP-2 and
MMP-9 expressions in prostate cancer PC-3 cells. Cancer Lett.
295:27–37. 2010. View Article : Google Scholar : PubMed/NCBI
|